
The Effect of Cold Stress on the Root-Specific Lipidome of Two Wheat Varieties with Contrasting Cold Tolerance

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Targeted Profiling Using Scheduled Parallel Reaction Monitoring Assays Enables the Profiling of the Complex Lipidome in Different Wheat Root Developmental Zones
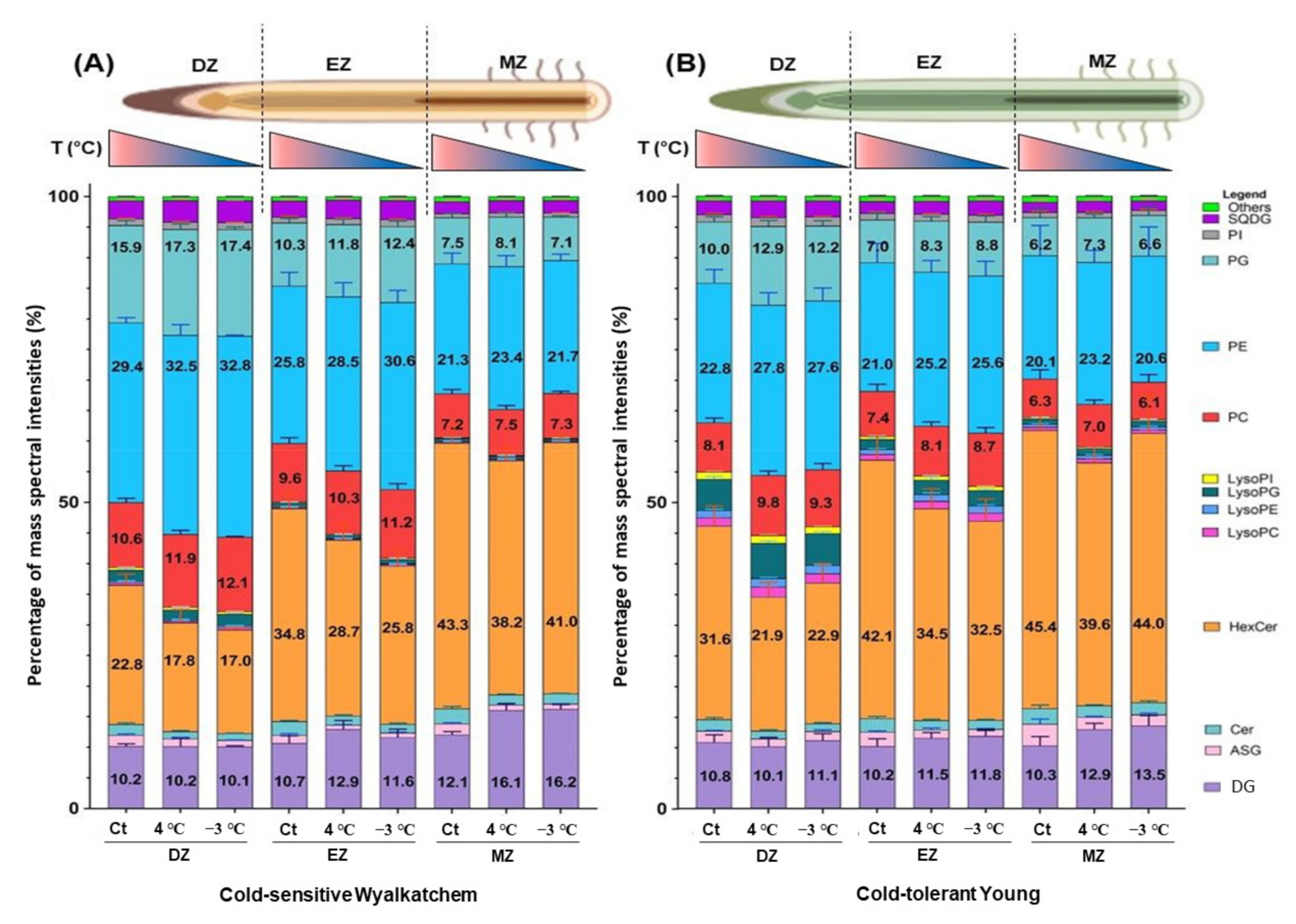
2.2. Overview of the Lipid Profiles in Each of the Developmental Root Zones of Wyalkatchem and Young Varieties at the Unstressed Stage, and after Chilling and Freezing Stress
2.3. Varietal Difference: Comparisons of Lipid Profiles of Each of the Developmental Root Zones of Cold-Tolerant Young with Cold-Sensitive Wyalkatchem after Chilling and Freezing Stress
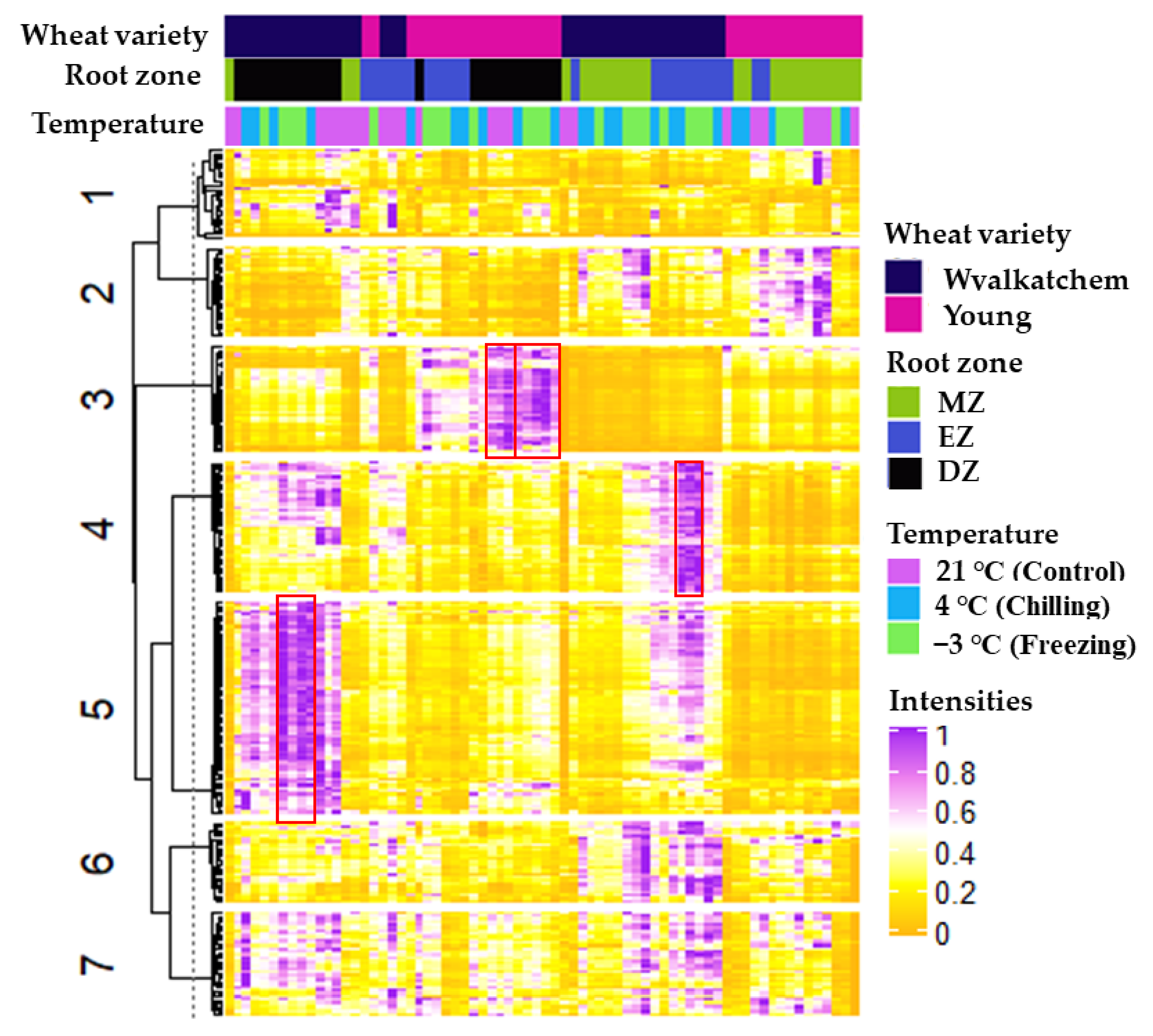
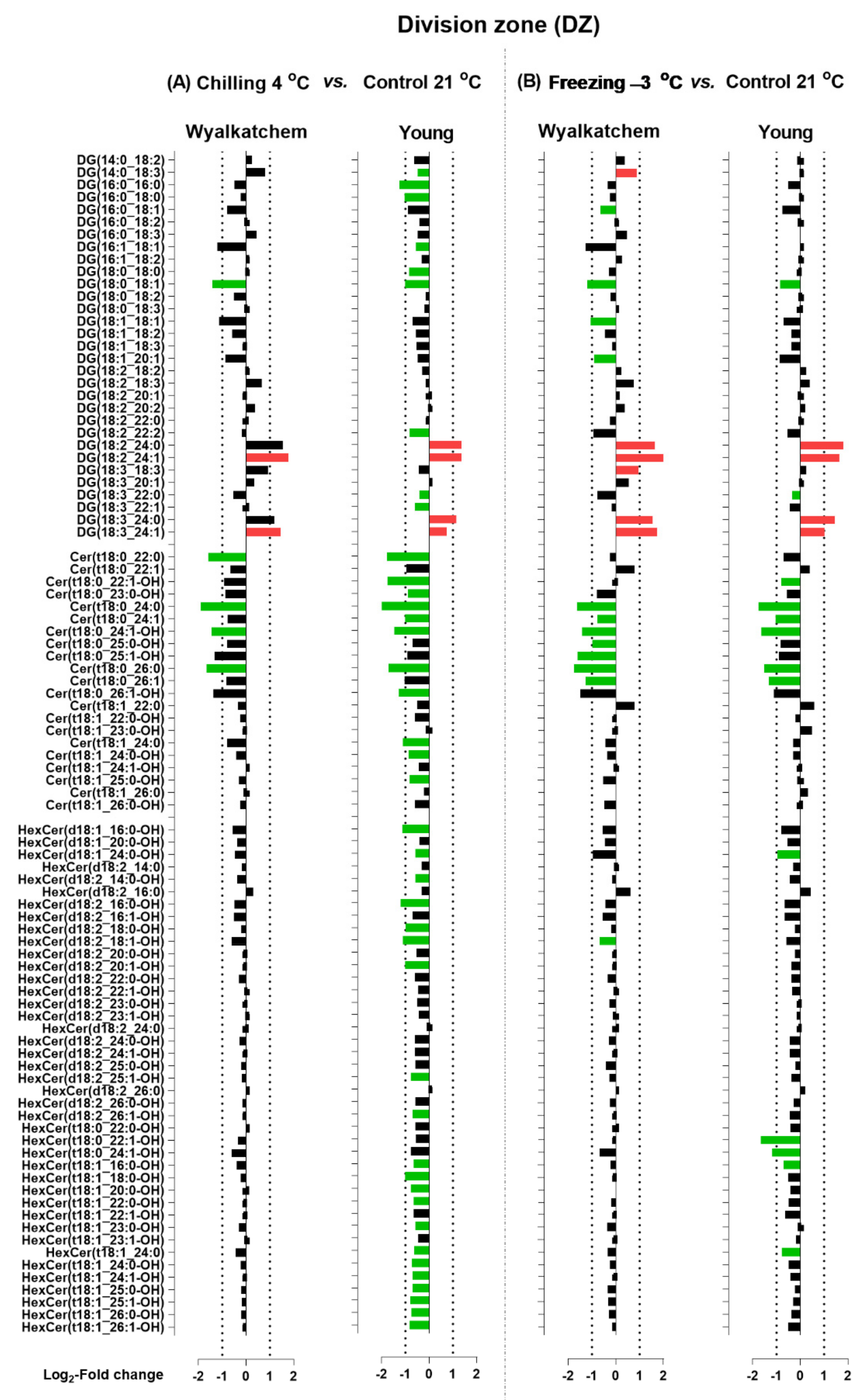
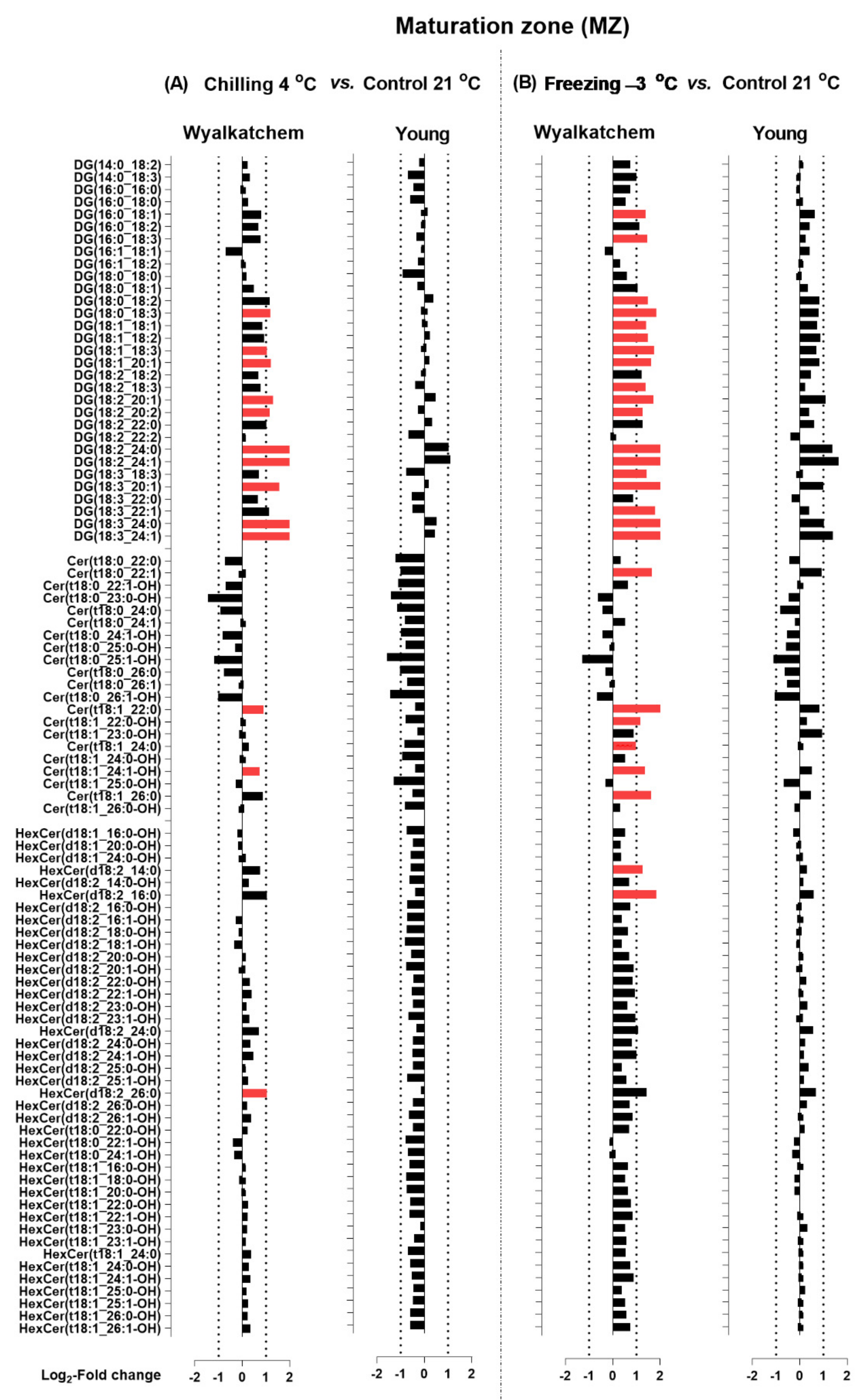
2.4. Lipid Species Contributing Changes in Lipid Profiles in the Three Developmental Root Zones of Both Wheat Varieties following Cold Stress
3. Discussion
3.1. Lipid Composition at the Unstressed Stage Reveal Varietal Differences among the Three Developmental Root Zones in Each Wheat Variety
3.2. Cold Stress Mainly Alters the Glycerolipid and Sphingolipid Compositions of the Root Zones, which May Reveal the Main Differences between the Cold-Sensitive and Cold-Tolerant Varieties
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Lipid Nomenclature, Annotation and Abbreviations
4.3. Plant Material and Growth Conditions
4.4. Stress Treatment and Harvest of Root Samples
4.5. Lipid Extraction
4.6. LC-ESI-MS/MS Analysis
4.7. Data Processing
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The International Wheat Genome Sequencing Consortium (IWGSC); Appels, R.; Eversole, K.; Feuillet, C.; Keller, B.; Rogers, J.; Stein, N.; Pozniak, C.J.; Stein, N.; Choulet, F.; et al. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, R.; Sayre, K.D.; Govaerts, B.; Gupta, R.; Subbarao, G.V.; Ban, T.; Hodson, D.; Dixon, J.M.; Ortiz-Monasterio, J.I.; Reynolds, M. Climate change: Can wheat beat the heat? Agric. Ecosyst. Environ. 2008, 126, 46–58. [Google Scholar] [CrossRef]
- Fowler, D.B.; Gusta, L.V. Selection for winter hardiness in wheat. I. Identification of genotypic variability. Crop Sci. 1979, 19, 769–772. [Google Scholar] [CrossRef]
- Fowler, D.B.; Chauvin, L.P.; Limin, A.E.; Sarhan, F. The regulatory role of vernalization in the expression of low-temperature-induced genes in wheat and rye. Theor. Appl. Genet. 1996, 93, 554–559. [Google Scholar] [CrossRef] [PubMed]
- Wanner, L.A.; Junttila, O. Cold-Induced Freezing Tolerance in Arabidopsis1. Plant Physiol. 1999, 120, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, W.; Li, M.; Welti, R. Profiling lipid changes in plant response to low temperatures. Physiol. Plant. 2006, 126, 90–96. [Google Scholar] [CrossRef]
- Miquel, M.; James, D., Jr.; Dooner, H.; Browse, J. Arabidopsis requires polyunsaturated lipids for low-temperature survival. Proc. Natl. Acad. Sci. USA 1993, 90, 6208–6212. [Google Scholar] [CrossRef] [Green Version]
- Welti, R.; Li, W.; Li, M.; Sang, Y.; Biesiada, H.; Zhou, H.-E.; Rajashekar, C.B.; Williams, T.D.; Wang, X. Profiling membrane lipids in plant stress responses: Role of phospholipase Dα in freezing-induced lipid changes in Arabidopsis. J. Biol. Chem. 2002, 277, 31994–32002. [Google Scholar] [CrossRef] [Green Version]
- Cacas, J.-L.; Furt, F.; Le Guédard, M.; Schmitter, J.-M.; Buré, C.; Gerbeau-Pissot, P.; Moreau, P.; Bessoule, J.-J.; Simon-Plas, F.; Mongrand, S. Lipids of plant membrane rafts. Prog. Lipid Res. 2012, 51, 272–299. [Google Scholar] [CrossRef]
- Dörmann, P. Galactolipids in Plant Membranes. eLS 2013. [Google Scholar] [CrossRef]
- Bates, P.D.; Browse, J. The Significance of Different Diacylgycerol Synthesis Pathways on Plant Oil Composition and Bioengineering. Front. Plant Sci. 2012, 3, 147. [Google Scholar] [CrossRef] [Green Version]
- Hou, Q.; Ufer, G.; Bartels, D. Lipid signalling in plant responses to abiotic stress. Plant Cell Environ. 2016, 39, 1029–1048. [Google Scholar] [CrossRef]
- Sperling, P.; Franke, S.; Luthje, S.; Heinz, E. Are glucocerebrosides the predominant sphingolipids in plant plasma membranes? Plant Physiol. Biochem. 2005, 43, 1031–1038. [Google Scholar] [CrossRef]
- Markham, J.E.; Lynch, D.V.; Napier, J.; Dunn, T.M.; Cahoon, E.B. Plant sphingolipids: Function follows form. Curr. Opin. Plant Biol. 2013, 16, 350–357. [Google Scholar] [CrossRef]
- Luttgeharm, K.D.; Kimberlin, A.N.; Cahoon, E.B. Plant Sphingolipid Metabolism and Function. Lipids Plant Algae Dev. 2016, 86, 249–286. [Google Scholar] [CrossRef]
- Simon-Plas, F.; Perraki, A.; Bayer, E.; Gerbeau-Pissot, P.; Mongrand, S. An update on plant membrane rafts. Curr. Opin. Plant Biol. 2011, 14, 642–649. [Google Scholar] [CrossRef]
- Yang, H.; Richter, G.L.; Wang, X.; Młodzińska, E.; Carraro, N.; Ma, G.; Jenness, M.; Chao, D.-Y.; Peer, W.A.; Murphy, A.S. Sterols and sphingolipids differentially function in trafficking of the Arabidopsis ABCB19 auxin transporter. Plant J. 2012, 74, 37–47. [Google Scholar] [CrossRef]
- Testerink, C.; Munnik, T. Phosphatidic acid: A multifunctional stress signaling lipid in plants. Trends Plant Sci. 2005, 10, 368–375. [Google Scholar] [CrossRef]
- Wang, X.; Chapman, K.D. Lipid signalling in plants. Front. Plant Sci. 2013, 4, 216. [Google Scholar] [CrossRef] [Green Version]
- Xiong, L.; Schumaker, K.S.; Zhu, J.-K. Cell Signaling during Cold, Drought, and Salt Stress. Plant Cell 2002, 14 (Suppl. S1), S165–S183. [Google Scholar] [CrossRef] [Green Version]
- Arisz, S.A.; Van Van Wijk, R.; Roels, W.; Zhu, J.-K.; Haring, M.A.; Munnik, T. Rapid phosphatidic acid accumulation in response to low temperature stress in Arabidopsis is generated through diacylglycerol kinase. Front. Plant Sci. 2013, 4, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bargmann, B.; Munnik, T. The role of phospholipase D in plant stress responses. Curr. Opin. Plant Biol. 2006, 9, 515–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbaglia, A.M.; Hoffmann-Benning, S. Long-Distance Lipid Signaling and its Role in Plant Development and Stress Response. 2016, 86, 339–361. Lipids Plant Algae Dev. 2016, 86, 339–361. [Google Scholar] [CrossRef]
- Guo, L.; Mishra, G.; Taylor, K.; Wang, X. Phosphatidic Acid Binds and Stimulates Arabidopsis Sphingosine Kinases. J. Biol. Chem. 2011, 286, 13336–13345. [Google Scholar] [CrossRef] [Green Version]
- Dutilleul, C.; Benhassaine-Kesri, G.; Demandre, C.; Reze, N.; Launay, A.; Pelletier, S.; Renou, J.-P.; Zachowski, A.; Baudouin, E.; Guillas, I. Phytosphingosine-phosphate is a signal for AtMPK6 activation and Arabidopsis response to chilling. New Phytol. 2012, 194, 181–191. [Google Scholar] [CrossRef]
- Ruelland, E.; Cantrel, C.; Gawer, M.; Kader, J.-C.; Zachowski, A. Activation of Phospholipases C and D Is an Early Response to a Cold Exposure in Arabidopsis Suspension Cells. Plant Physiol. 2002, 130, 999–1007. [Google Scholar] [CrossRef] [Green Version]
- Moellering, E.R.; Muthan, B.; Benning, C. Freezing tolerance in plants requires lipid remodelling at the outer chloroplast membrabe. Science 2010, 330, 226–228. [Google Scholar] [CrossRef] [Green Version]
- Degenkolbe, T.; Giavalisco, P.; Zuther, E.; Seiwert, B.; Hincha, D.K.; Willmitzer, L. Differential remodeling of the lipidome during cold acclimation in natural accessions of Arabidopsis thaliana. Plant J. 2012, 72, 972–982. [Google Scholar] [CrossRef]
- Li, Q.; Zheng, Q.; Shen, W.; Cram, D.; Fowler, D.B.; Wei, Y.; Zou, J. Understanding the Biochemical Basis of Temperature-Induced Lipid Pathway Adjustments in Plants. Plant Cell 2015, 27, 86–103. [Google Scholar] [CrossRef] [Green Version]
- Barnes, A.C.; Benning, C.; Roston, R.L. Chloroplast Membrane Remodeling during Freezing Stress Is Accompanied by Cytoplasmic Acidification Activating SENSITIVE TO FREEZING2. Plant Physiol. 2016, 171, 2140–2149. [Google Scholar] [CrossRef] [Green Version]
- Zheng, G.; Li, L.; Li, W. Glycerolipidome responses to freezing- and chilling-induced injuries: Examples in Arabidopsis and rice. BMC Plant Biol. 2016, 16, 70. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; He, L.; Zhao, C.; Wang, F.; Yan, B.; Gao, Y.; Li, Z.; Yang, K.; Xu, J. Biochemical and Transcriptional Regulation of Membrane Lipid Metabolism in Maize Leaves under Low Temperature. Front. Plant Sci. 2017, 8, 2053. [Google Scholar] [CrossRef] [Green Version]
- Noblet, A.; Leymarie, J.; Bailly, C. Chilling temperature remodels phospholipidome of Zea mays seeds during imbibition. Sci. Rep. 2017, 7, 8886. [Google Scholar] [CrossRef] [Green Version]
- Margutti, M.P.; Reyna, M.; Meringer, M.V.; Racagni, G.E.; Villasuso, A.L. Lipid signalling mediated by PLD/PA modulates proline and H 2 O 2 levels in barley seedlings exposed to short- and long-term chilling stress. Plant Physiol. Biochem. 2017, 113, 149–160. [Google Scholar] [CrossRef]
- Margutti, M.P.; Reyna, M.; Vilchez, A.C.; Villasuso, A.L. Lipid profiling shows tissue-specific differences in barley for glycerolipid composition in response to chilling. Environ. Exp. Bot. 2018, 158, 150–160. [Google Scholar] [CrossRef]
- De La Roche, I.A.; Andrews, C.J.; Pomeroy, M.K.; Weinberger, P.; Kates, M. Lipid changes in winter wheat seedlings (Triticum aestivum) at temperatures inducing cold hardiness. Can. J. Bot. 1972, 50, 2401–2409. [Google Scholar] [CrossRef]
- Ashworth, E.N.; Christiansen, M.N.; John, J.B.S.; Patterson, G.W. Effect of Temperature and BASF 13 338 on the Lipid Composition and Respiration of Wheat Roots. Plant Physiol. 1981, 67, 711–715. [Google Scholar] [CrossRef]
- Vigh, L.; Horvàth, I.; Van Hasselt, P.R.; Kuiper, P.J.C. Effect of Frost Hardening on Lipid and Fatty Acid Composition of Chloroplast Thylakoid Membranes in Two Wheat Varieties of Contrasting Hardiness. Plant Physiol. 1985, 79, 756–759. [Google Scholar] [CrossRef] [Green Version]
- Kendall, E.J.; McKersie, B.D. Free radical and freezing injury to cell membranes of winter wheat. Physiol. Plant. 1989, 76, 86–94. [Google Scholar] [CrossRef]
- Bohn, M.; Lüthje, S.; Sperling, P.; Heinz, E.; Dörffling, K. Plasma membrane lipid alterations induced by cold acclimation and abscisic acid treatment of winter wheat seedlings differing in frost resistance. J. Plant Physiol. 2007, 164, 146–156. [Google Scholar] [CrossRef]
- Li, H.-X.; Xiao, Y.; Cao, L.-L.; Yan, X.; Li, C.; Shi, H.-Y.; Wang, J.-W.; Ye, Y.-H. Cerebroside C Increases Tolerance to Chilling Injury and Alters Lipid Composition in Wheat Roots. PLoS ONE 2013, 8, e73380. [Google Scholar] [CrossRef] [Green Version]
- Cheong, B.E.; Ho, W.W.H.; Biddulph, B.; Wallace, X.; Rathjen, T.; Rupasinghe, T.W.T.; Roessner, U.; Dolferus, R. Phenotyping reproductive stage chilling and frost tolerance in wheat using targeted metabolome and lipidome profiling. Metabolomics 2019, 15, 144. [Google Scholar] [CrossRef] [Green Version]
- Cheong, B.E.; Onyemaobi, O.; Ho, W.W.H.; Ben Biddulph, T.; Rupasinghe, T.W.T.; Roessner, U.; Dolferus, R. Phenotyping the Chilling and Freezing Responses of Young Microspore Stage Wheat Spikes Using Targeted Metabolome and Lipidome Profiling. Cells 2020, 9, 1309. [Google Scholar] [CrossRef]
- Sarabia, L.D.; Hill, C.B.; Boughton, B.; Roessner, U. Advancs of metabolite profiling of plants in challenging environments. In Annual Plant Reviews; Roberts, J., Ed.; John Wiley and Sons: New York, NY, USA, 2018; Volume 1, pp. 1–45. [Google Scholar]
- Sarabia, L.D.; Boughton, B.A.; Rupasinghe, T.; van de Meene, A.M.L.; Callahan, D.L.; Hill, C.B.; Roessner, U. High-mass-resolution MALDI mass spectrometry imaging reveals detailed spatial distribution of metabolites and lipids in roots of barley seedlings in response to salinity stress. Metabolomics 2018, 14, 63. [Google Scholar] [CrossRef] [Green Version]
- Ho, W.W.H.; Hill, C.B.; Doblin, M.S.; Shelden, M.C.; van de Meene, A.; Rupasinghe, T.; Bacic, A.; Roessner, U. Integrative Multi-omics Analyses of Barley Rootzones under Salinity Stress Reveal Two Distinctive Salt Tolerance Mechanisms. Plant Commun. 2020, 1, 100031. [Google Scholar] [CrossRef]
- Nielsen, K.F.; Humphries, E.C. Effect of temperature on plant growth. Soils Fert. 1966, 29, 1–7. [Google Scholar]
- Petr, J. (Ed.) Weather and Yield; Elsevier: Amsterdam, The Netherlands, 1991; p. 288. [Google Scholar]
- Somssich, M.; Khan, G.A.; Persson, S. Cell Wall Heterogeneity in Root Development of Arabidopsis. Front. Plant Sci. 2016, 7, 1242. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Rupasinghe, T.W.T.; Boughton, B.A.; Natera, S.H.A.; Hill, C.B.; Tarazona, P.; Feussner, I.; Roessner, U. A high-resolution HPLC-QqTOF platform using parallel reaction monitoring for in-depth lipid discovery and rapid profiling. Anal. Chim. Acta 2018, 1026, 87–100. [Google Scholar] [CrossRef]
- Yu, D.; Boughton, B.A.; Hill, C.B.; Feussner, I.; Roessner, U.; Rupasinghe, T.W.T. Insights into oxidized lipid modification in barley roots as an adaptation mechanism to salinity stress. Front. Plant Sci. 2020, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, R.; Shimodaira, H. Pvclust: An R package for assessing the uncertainty in hierarchical clustering. Bioinformatics 2006, 22, 1540–1542. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y. Plant Phospholipid Diversity: Emerging Functions in Metabolism and Protein–Lipid Interactions. Trends Plant Sci. 2017, 22, 1027–1040. [Google Scholar] [CrossRef] [PubMed]
- Lynch, D.V. Sphingolipids. In Lipid Metabolism in Plants; Moore, T.S., Jr., Ed.; CRC Press: Boca Raton, FL, USA, 1993; pp. 285–308. [Google Scholar]
- Warnecke, D.; Heinz, E. Recently discovered functions of glucosylceramides in plants and fungi. Cell. Mol. Life Sci. 2003, 60, 919–941. [Google Scholar] [CrossRef] [PubMed]
- Lynch, D.V.; Dunn, T.M. An introduction to plant sphingolipids and a review of recent advances in understanding their metabolism and function. New Phytol. 2004, 161, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Lester, R.L.; Dickson, R.C. Sphingolipids with inositolphosphate-containing head groups. Adv. Lipid Res. 1993, 26, 253–274. [Google Scholar]
- Liang, H.; Yao, N.; Song, J.T.; Luo, S.; Lu, H.; Greenberg, J.T. Ceramide phosphorylation modulates programmed cell death in plants. Genes Dev. 2003, 17, 2636–2641. [Google Scholar] [CrossRef] [Green Version]
- Merrill, A.H., Jr.; Sweeley, C.C. Sphingolipids: Metabolism and cell signalling. In New Comprehensive Biochemistry: Biochemistry of Lipids, Lipoproteins and Membranes; Vance, D.E., Vance, J.E., Eds.; Elsevier: Amsterdam, The Netherlands, 1996; pp. 309–340. [Google Scholar]
- Michaelson, L.V.; Napier, J.A.; Molino, D.; Faure, J.-D. Plant sphingolipids: Their importance in cellular organization and adaption. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2016, 1861, 1329–1335. [Google Scholar] [CrossRef]
- Boggs, J.M. Lipid intermolecular hydrogen bonding: Influence on structural organization and membrane function. Biochim. Biophys. Acta (BBA)-Rev. Biomembr. 1987, 906, 353–404. [Google Scholar] [CrossRef]
- Curatolo, W. The physical properties of glycolipids. Biochim. Biophys. Acta 1987, 906, 111–136. [Google Scholar] [CrossRef]
- Karlsson, K.-A. Glycosphingolipids and surface membranes. In Biological Membranes; Chapman, D., Ed.; Academic Press: New York, NY, USA, 1982; pp. 1–74. [Google Scholar]
- Steponkus, P.L. Role of the plasma membrane in freezing injury and cold acclimation. Annu. Rev. Plant Physiol. 1984, 35, 543–584. [Google Scholar] [CrossRef]
- Coursol, S.; Fan, L.-M.; Le Stunff, H.; Spiegel, S.; Gilroy, S.; Assmann, S.M. Sphingolipid signalling in Arabidopsis guard cells involves heterotrimeric G proteins. Nature 2003, 423, 651–654. [Google Scholar] [CrossRef]
- Coursol, S.; Le Stunff, H.; Lynch, D.V.; Gilroy, S.; Assmann, S.M.; Spiegel, S. Arabidopsis Sphingosine Kinase and the Effects of Phytosphingosine-1-Phosphate on Stomatal Aperture. Plant Physiol. 2005, 137, 724–737. [Google Scholar] [CrossRef] [Green Version]
- Schiefelbein, J.W.; Benfey, P.N. The development of plant roots: New approaches to underground problems. Plant Cell 1991, 3, 1147–1154. [Google Scholar]
- Uemura, M.; Steponkus, P.L. Effect of Cold Acclimation on the Lipid Composition of the Inner and Outer Membrane of the Chloroplast Envelope Isolated from Rye Leaves. Plant Physiol. 1997, 114, 1493–1500. [Google Scholar] [CrossRef] [Green Version]
- Uemura, M.; Joseph, R.A.; Steponkus, P.L. Cold Acclimation of Arabidopsis thaliana (Effect on Plasma Membrane Lipid Composition and Freeze-Induced Lesions). Plant Physiol. 1995, 109, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Xu, Y.; Wang, J.; Singer, S.D.; Chen, G. The Role of Triacylglycerol in Plant Stress Response. Plants 2020, 9, 472. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhu, H.; Zhang, Q.; Li, M.; Yan, M.; Wang, R.; Wang, L.; Welti, R.; Zhang, W.; Wang, X. Phospholipase D_1 and phosphatidic acid regulate NADPH oxidase activity and production of reactive oxygen species in ABA-mediated stomatal closure in Arabidopsis. Plant Cell 2009, 21, 2357–2377. [Google Scholar] [CrossRef] [Green Version]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2013, 65, 1229–1240. [Google Scholar] [CrossRef]
- Webb, M.S.; Green, B.R. Biochemical and biophysical properties of thylakoid acyl lipids. Biochim. Biophys. Acta 1991, 1060, 133–158. [Google Scholar] [CrossRef]
- Arisz, S.A.; Heo, J.-Y.; Koevoets, I.T.; Zhao, T.; Van Egmond, P.; Meyer, A.J.; Zeng, W.; Niu, X.; Wang, B.; Mitchell-Olds, T.; et al. Diacylglycerol acyltransferase1 Contributes to Freezing Tolerance. Plant Physiol. 2018, 177, 1410–1424. [Google Scholar] [CrossRef] [Green Version]
- Tan, W.; Yang, Y.; Huang, L.; Xu, L.; Chen, Q.; Yu, L.; Xiao, S. Diacylglycerol acyltranferase and diacylglycerol kinase modulate triacylglycerol and phosphatidic acid production in the plant response to freezing stress. Plant Physiol. 2018, 177, 1303–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, D.V.; Steponkus, P.L. Plasma Membrane Lipid Alterations Associated with Cold Acclimation of Winter Rye Seedlings (Secale cereale L. cv Puma). Plant Physiol. 1987, 83, 761–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uemura, M.; Steponkus, P.L. A Contrast of the Plasma Membrane Lipid Composition of Oat and Rye Leaves in Relation to Freezing Tolerance. Plant Physiol. 1994, 104, 479–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steponkus, P.L.; Lynch, D.V. Freeze/thaw-induced destabilization of the plasma membrane and the effects of cold acclimation. J. Bioenerg. Biomembr. 1989, 21, 21–41. [Google Scholar] [CrossRef]
- Steponkus, P.L.; Lynch, D.V.; Uemura, M. The influence of cold acclimation on the lipid composition and cryobehavior of the plasma membrane of isolated rye protoplasts. Philos. Trans. R. Soc. Lond. B 1990, 326, 571–583. [Google Scholar]
- Imai, H.; Ohnishi, M.; Kinoshita, M.; Kojima, M.; Ito, S. Structure and Distribution of Cerebroside Containing Unsaturated Hydroxy Fatty Acids in Plant Leaves. Biosci. Biotechnol. Biochem. 1995, 59, 1309–1313. [Google Scholar] [CrossRef] [Green Version]
- Imai, H.; Ohnishi, M.; Hotsubo, K.; Kojima, M.; Ito, S. Sphingoid Base Composition of Cerebrosides from Plant Leaves. Biosci. Biotechnol. Biochem. 1997, 61, 351–353. [Google Scholar] [CrossRef] [Green Version]
- Imai, H. Glucocerebrosides containing unsaturated hydroxy fatty acids in Arabidopsis thaliana. In Advances in Plant Lipid Research; Sanchez, J., Cerda-Olmedo, E., Martinez-Force, E., Eds.; University of Seville Publications: Seville, Spain, 1998; pp. 38–40. [Google Scholar]
- Douce, R.; Joyard, J. Plant galactolipids. In Lipids: Structure and Function; Academic Press: Cambridge, MA, USA, 1980; pp. 321–362. [Google Scholar]
- McCarty, R.E.; Jagendorf, A.T. Chloroplast damage due to enzymatic hydrolysis of endogenous lipids. Plant Physiol. 1965, 40, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Moellering, E.R.; Benning, C. Galactoglycerolipid metabolism under stress: A time for remodeling. Trends Plant Sci. 2011, 16, 98–107. [Google Scholar] [CrossRef]
- Hölzl, G.; Dörmann, P. Chloroplast Lipids and Their Biosynthesis. Annu. Rev. Plant Biol. 2019, 70, 51–81. [Google Scholar] [CrossRef]
- Vu, H.S.; Shiva, S.; Roth, M.R.; Tamura, P.; Zheng, L.; Li, M.; Sarowar, S.; Honey, S.; McEllhiney, D.; Hinkes, P.; et al. Lipid changes after leaf wounding in Arabidopsis thaliana: Expanded lipidomic data form the basis for lipid co-occurrence analysis. Plant J. 2014, 80, 728–743. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Ihaka, R.; Gentleman, R. R: A Language for Data Analysis and Graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheong, B.E.; Yu, D.; Martinez-Seidel, F.; Ho, W.W.H.; Rupasinghe, T.W.T.; Dolferus, R.; Roessner, U. The Effect of Cold Stress on the Root-Specific Lipidome of Two Wheat Varieties with Contrasting Cold Tolerance. Plants 2022, 11, 1364. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101364
Cheong BE, Yu D, Martinez-Seidel F, Ho WWH, Rupasinghe TWT, Dolferus R, Roessner U. The Effect of Cold Stress on the Root-Specific Lipidome of Two Wheat Varieties with Contrasting Cold Tolerance. Plants. 2022; 11(10):1364. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101364
Chicago/Turabian StyleCheong, Bo Eng, Dingyi Yu, Federico Martinez-Seidel, William Wing Ho Ho, Thusitha W. T. Rupasinghe, Rudy Dolferus, and Ute Roessner. 2022. "The Effect of Cold Stress on the Root-Specific Lipidome of Two Wheat Varieties with Contrasting Cold Tolerance" Plants 11, no. 10: 1364. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101364