
Chlorophyll Fluorescence Imaging as a Tool for Evaluating Disease Resistance of Common Bean Lines in the Western Amazon Region of Colombia

 , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Results
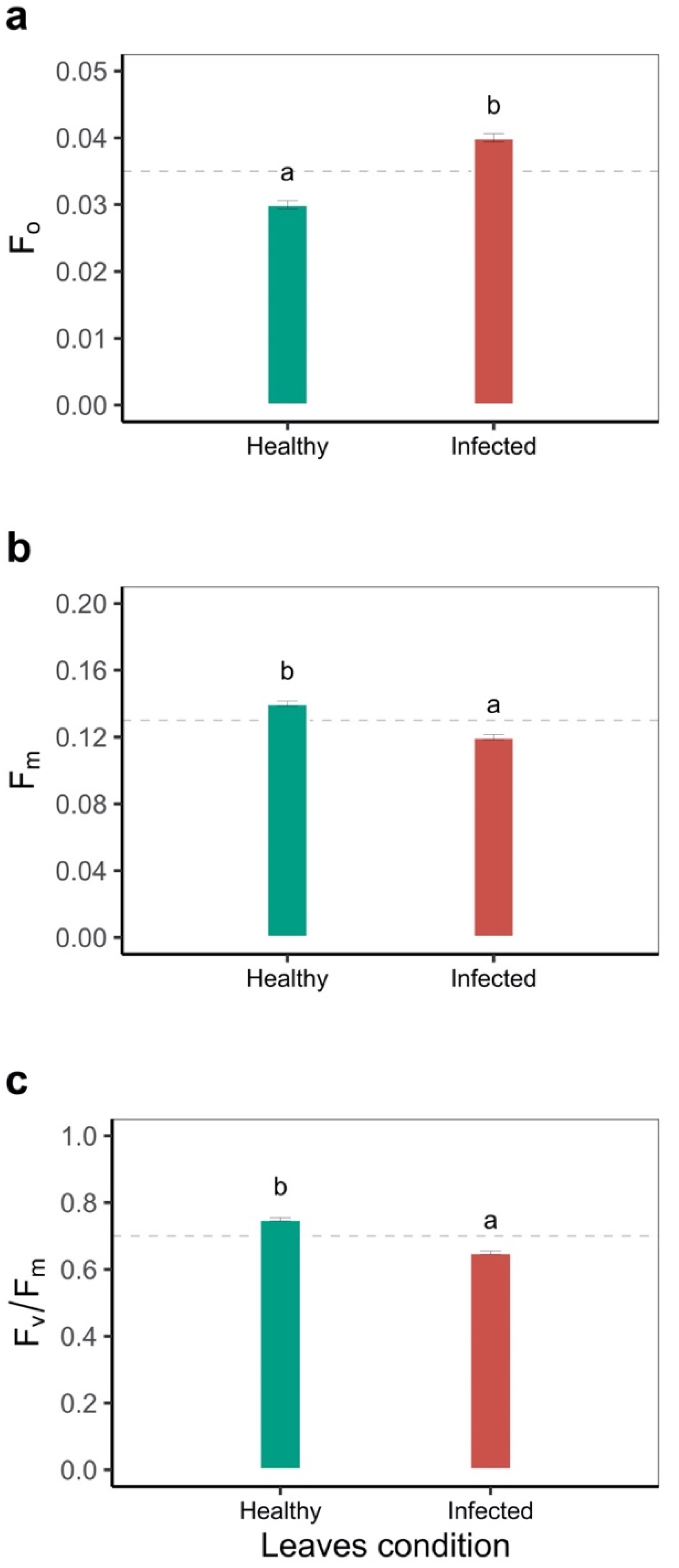
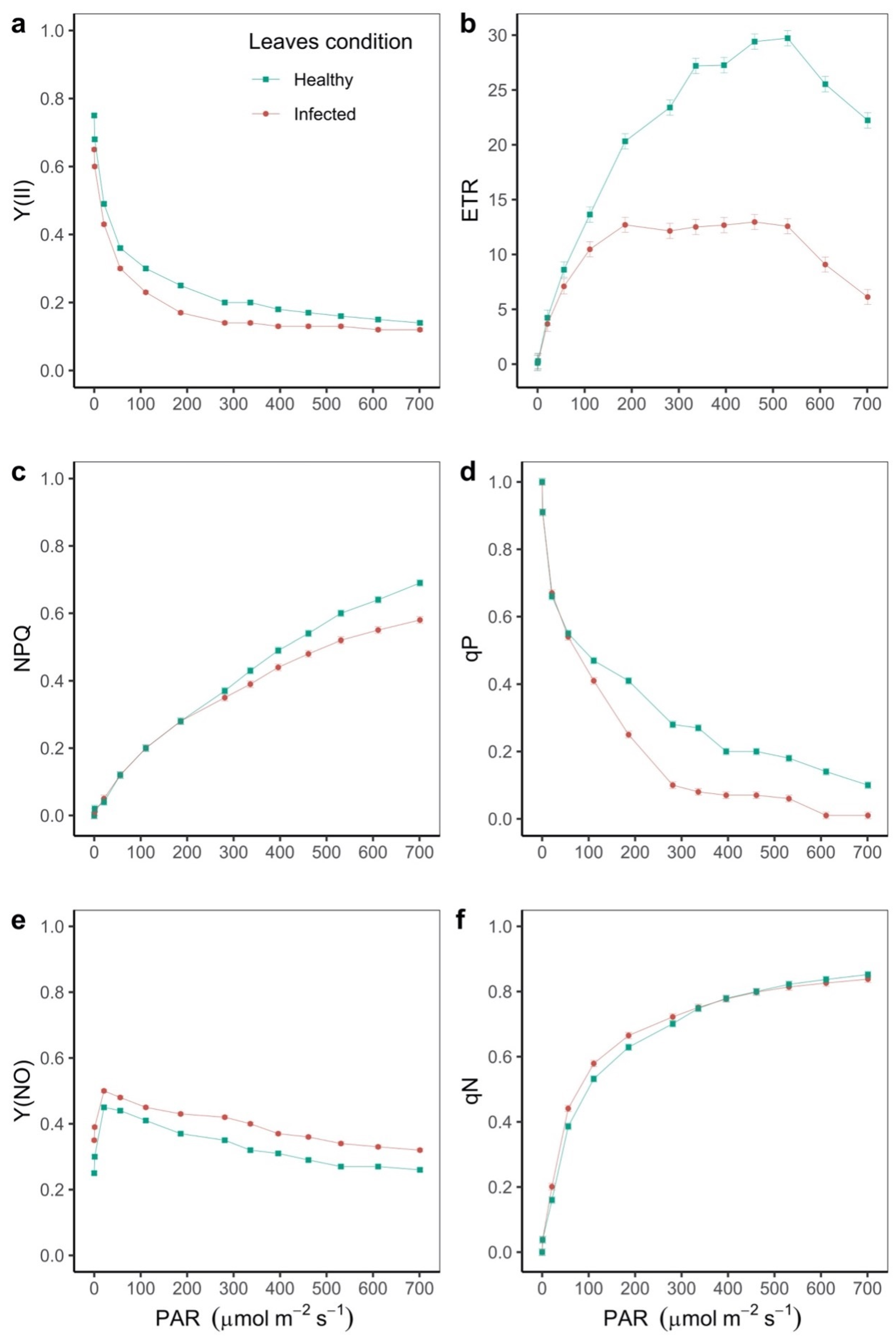
2.1. Response of the Photosynthetic Apparatus to Disease Stress in Leaves of Phaseolus vulgaris
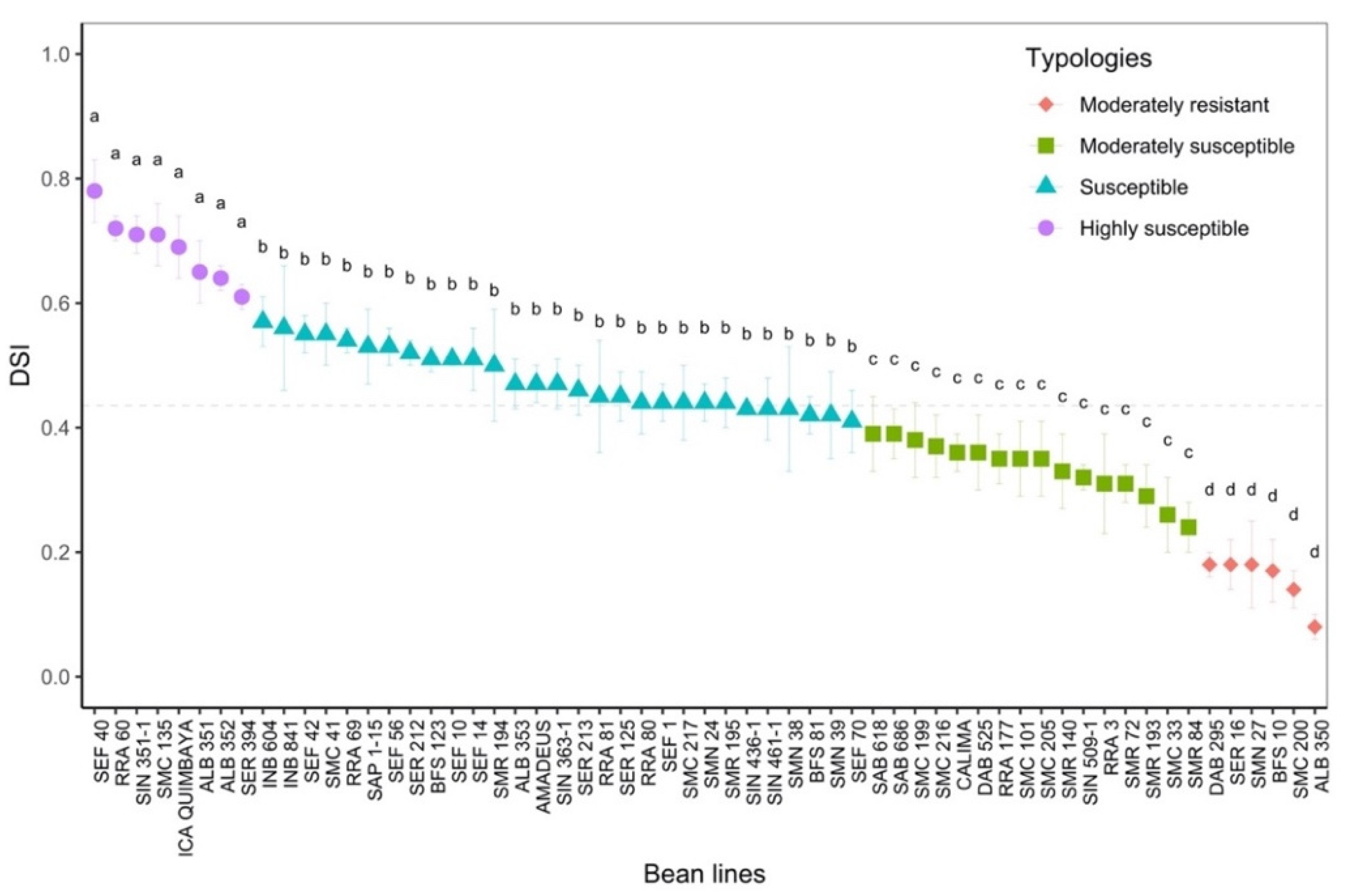
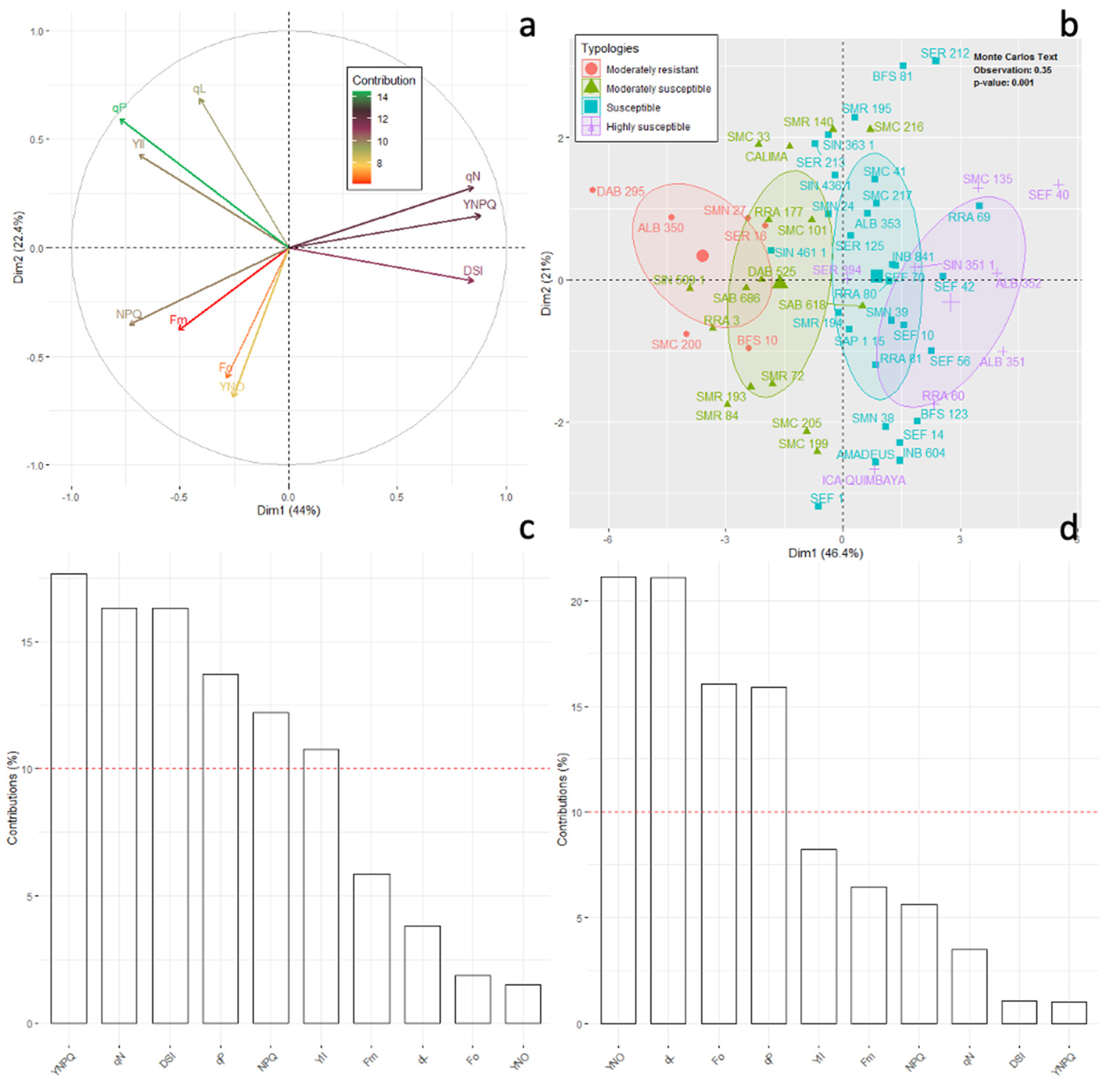
2.2. Phenotyping of Advanced Lines of Phaseolus vulgaris for Susceptibility to Disease Stress
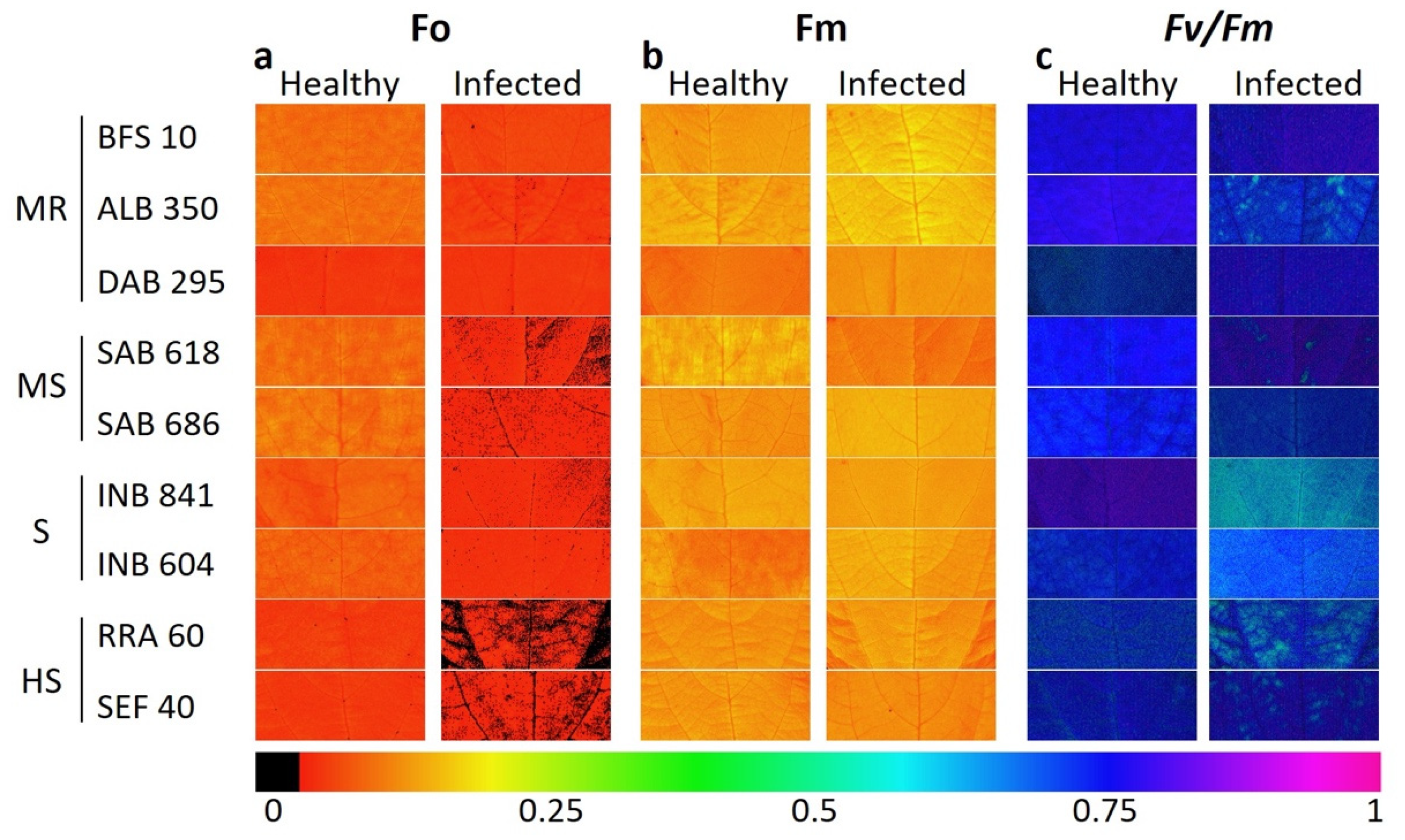
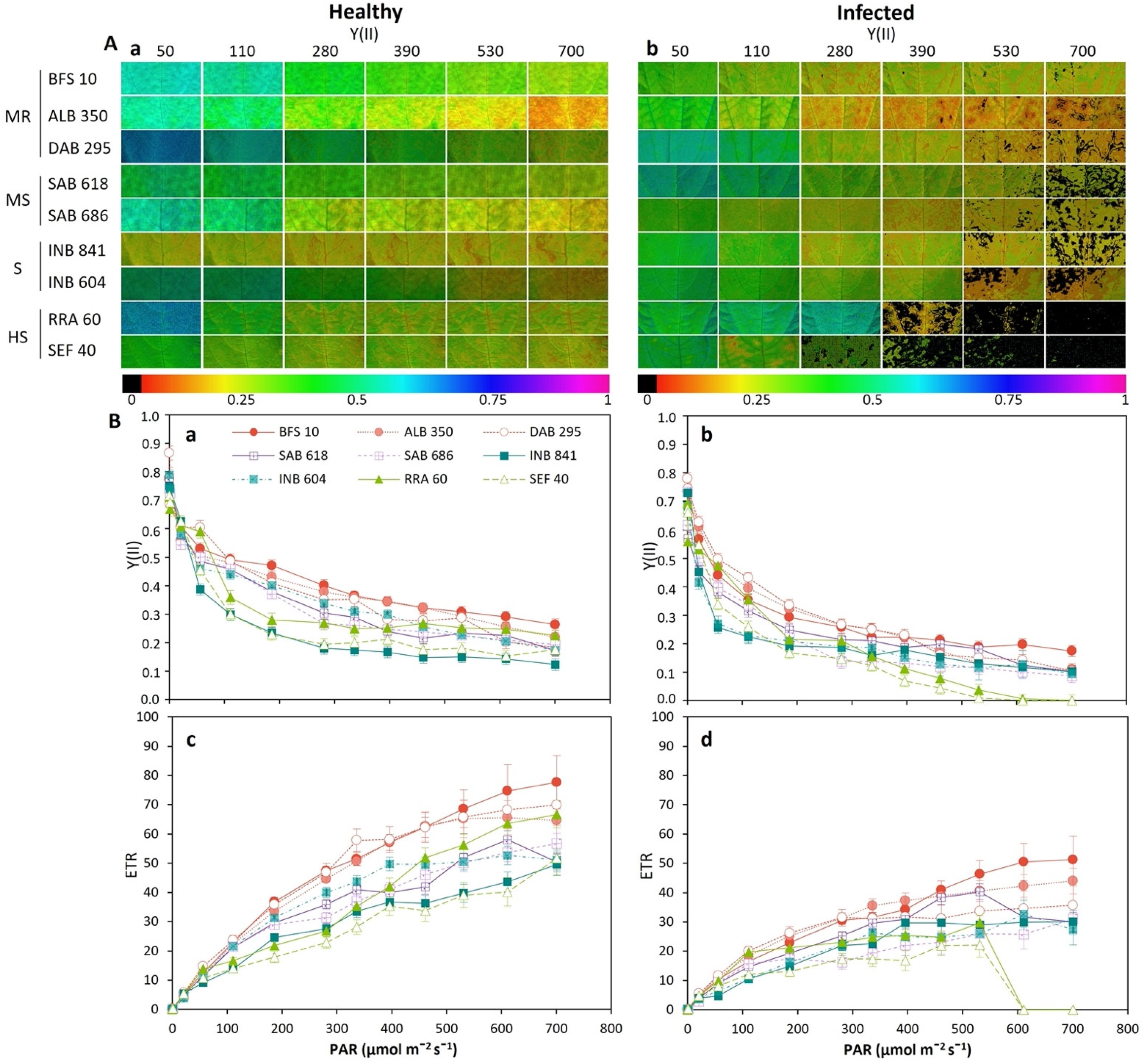
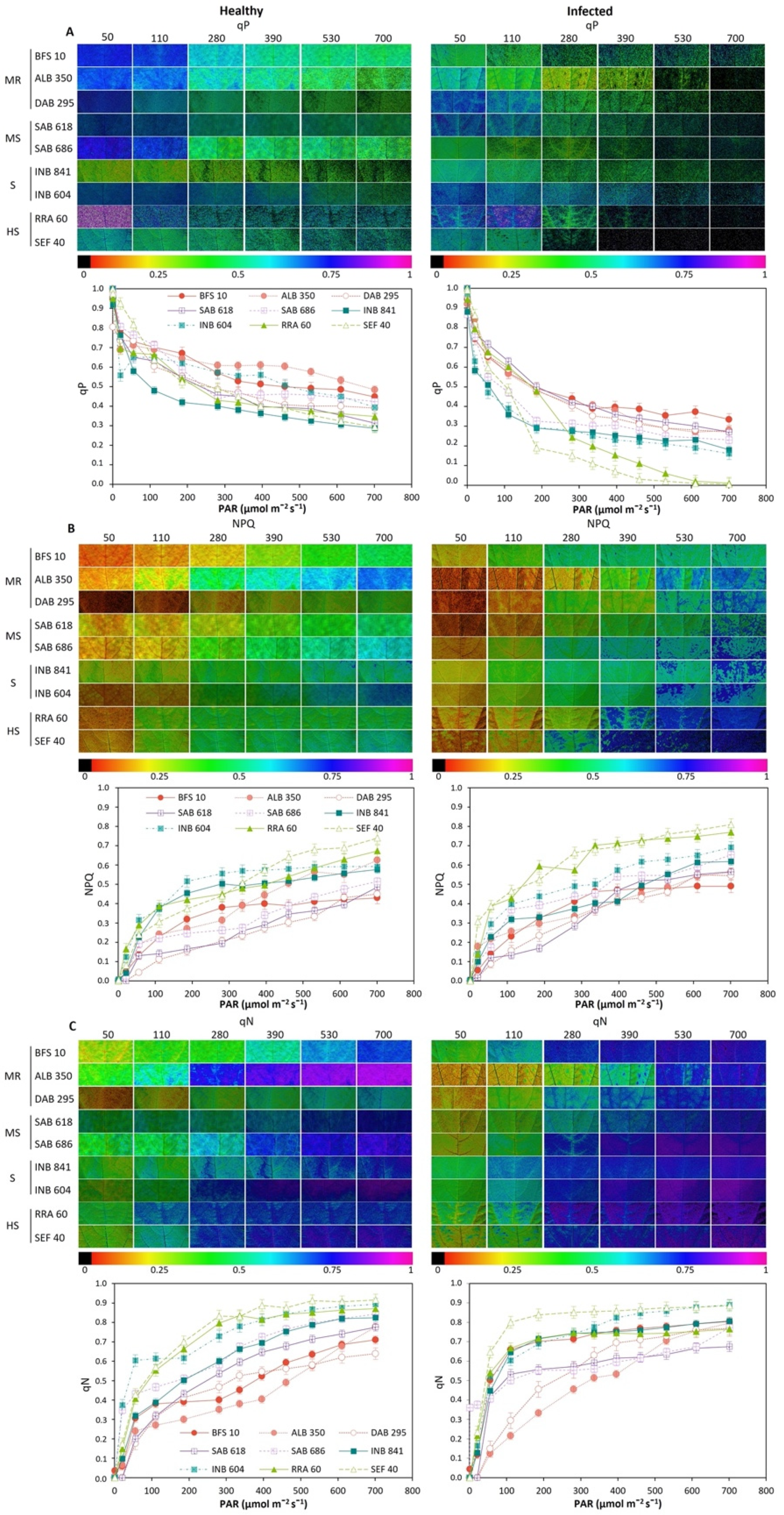
- Moderately resistant (MR; n = 6; 21.8% of the total genotypes evaluated) was a typology characterized by low and high levels of Fo and Fm, respectively, in the dark adaptation phase. At the level of energy distribution in the different fractions, this typology assigned the highest proportion to qP (p < 0.05) and, likewise, to efficiency in the electron transfer rate (ETR, p < 0.05). This typology had a lower DSI with an overall average of 0.16 ± 0.02 (represented by lines ALB 350, SMC 200, BFS 10, DAB 295 and SER 16). In terms of agronomic response (Table 1), these lines accumulated a greater amount of biomass in different organs, such as the root, stem, leaves and canopy biomass (p < 0.001), and showed a greater number of pods (4.6 ± 0.29 g). The plants accumulated canopy biomass twice as much compared to the highly susceptible type.
- Moderately susceptible (MS; DSI = 0.33, n = 16; 27.1%) was a typology that was characterized by an increase in the fraction of non-regulated energy (qN), and its reaction centers were moderately reduced so that its electron transfer rate (ETR) was lower compared to the moderately resistant bean lines. The best and lowest rated lines in terms of DSI were SMR 84 and SAB 686, respectively. This typology reduced its pod biomass; however, a strong presence of flowers was found with an increase in root biomass.
- Susceptible (S; DSI = 0.48, n = 29; 49.1%) was a typology where Y(II) was reduced by 40% and the fraction of energy dissipated as heat (NPQ) and non-regulated (qN) was increased by 46% and 40%, respectively, in relation to the moderately resistant bean lines. Likewise, this typology showed a considerable reduction in the fraction of energy distributed to the photosynthetic process (qP, 44.8%) and in the electron transport rate (ETR, 53.2%). The lines that conformed to this typology experienced a greater effect from disease stress on pod production, as well as on the other plant organs.
- Highly susceptible (HS; DSI = 0.69, n = 8; 13.5%) was a typology where the energy was distributed in a higher proportion in the form of heat dissipation (NPQ) and as non-regulated (qN), and the efficiency of the photosynthetic apparatus was very low (qP and ETR). At the agronomic level, a significant effect was found since several plant organs did not show adequate development; however, an increase in root biomass was found at the expense of shoot growth.
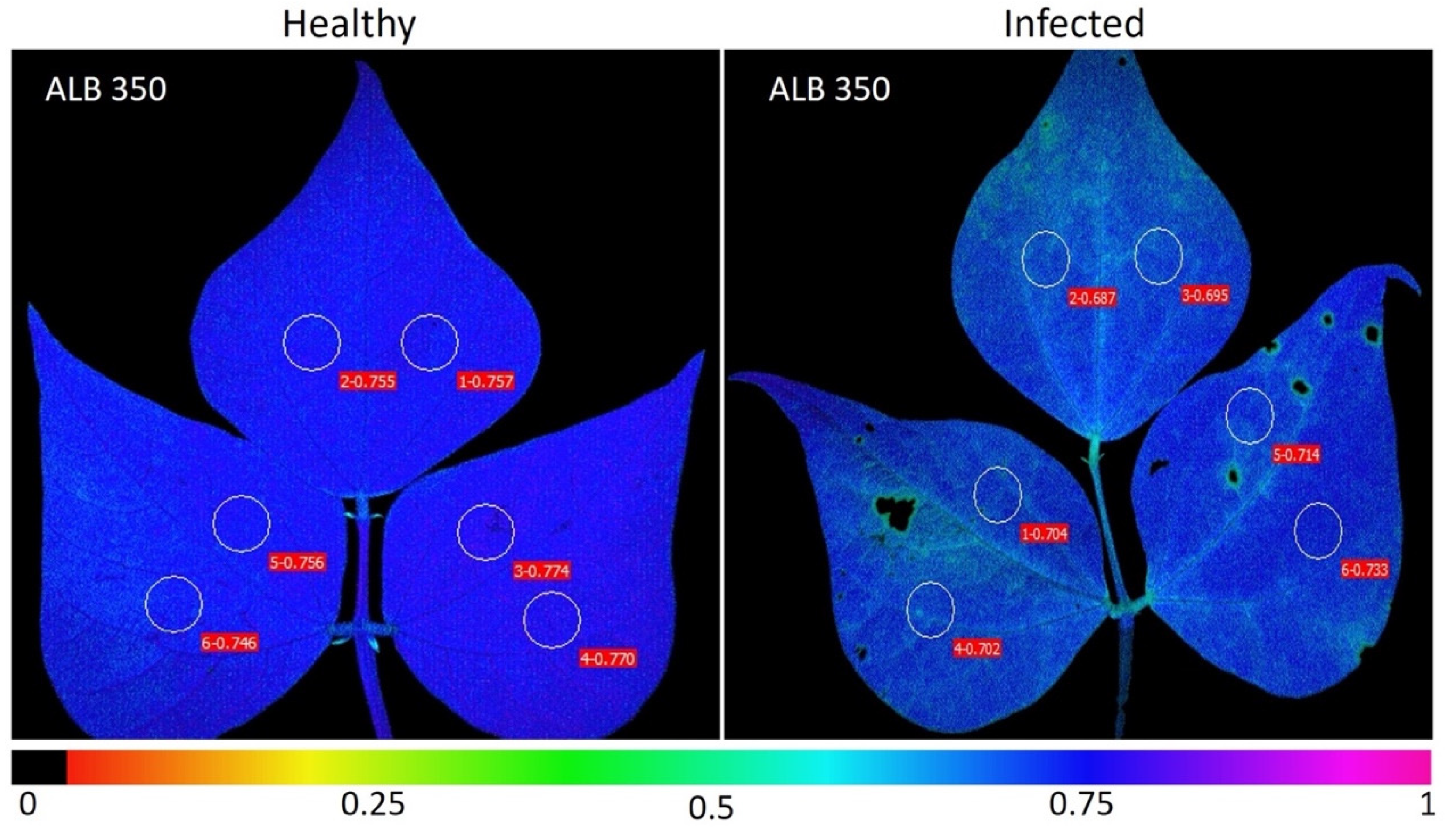
2.3. Fluorescence and Images of Chlorophyll A (Chla) Fluorescence Parameters for Phenotyping of Advanced Lines of Phaseolus vulgaris
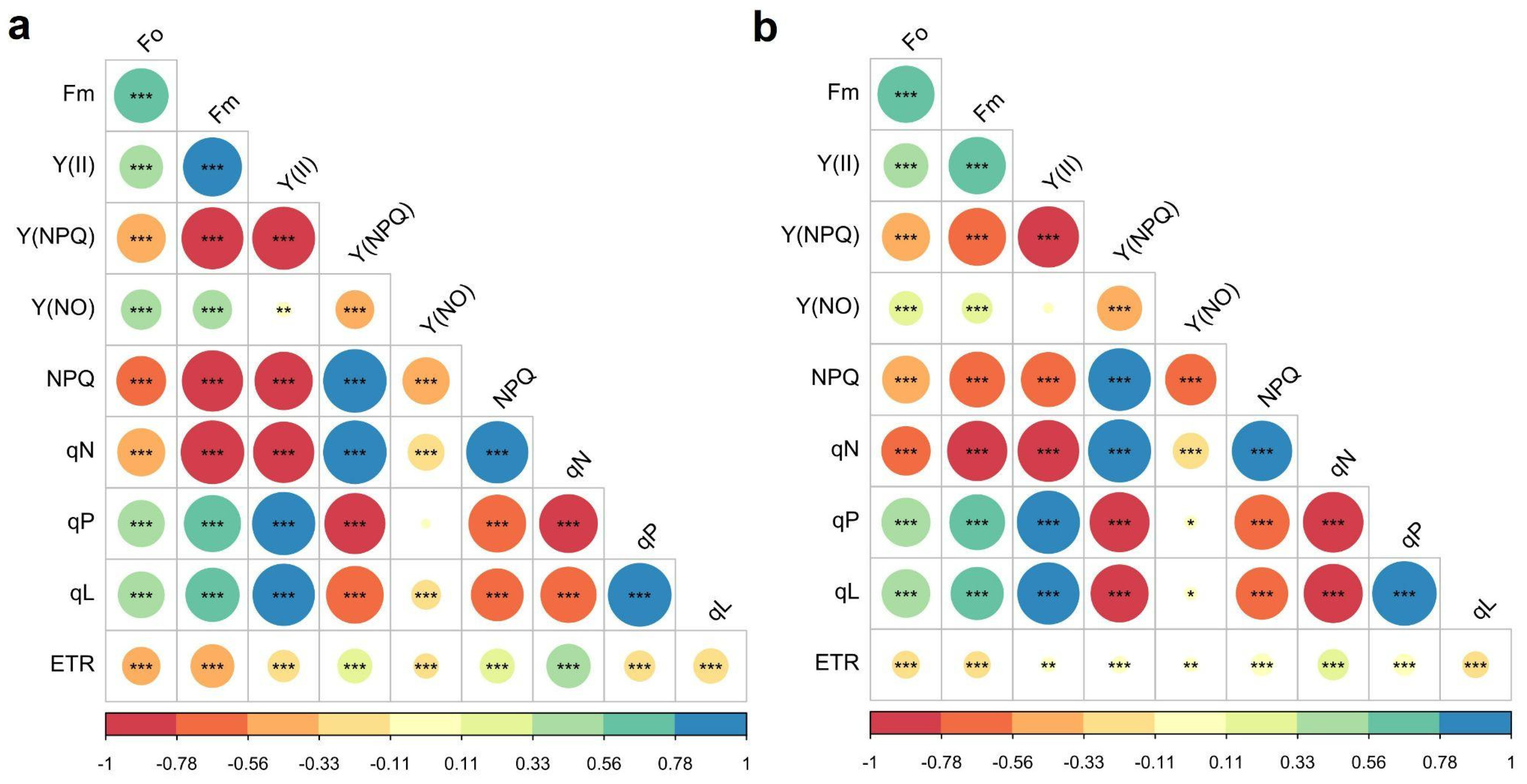
2.4. Correlations of Disease Susceptibility Index (DSI) with Chlorophyll A (Chla) Fluorescence Parameters and Agronomic Variables
3. Discussion
3.1. Magnitude of Leaf Disease Severity Modifies Energy Pathways within the Photosynthetic Apparatus
3.2. Physiological Response Differences among Common Bean Lines Reflect Possible Disease Resistance Mechanisms
3.3. Identification of Mechanisms of Energy Use among Moderately Resistant and Susceptible Bean Lines
3.4. Traits Associated with Lower Values of DSI Favor Better Agronomic Performance
4. Materials and Methods
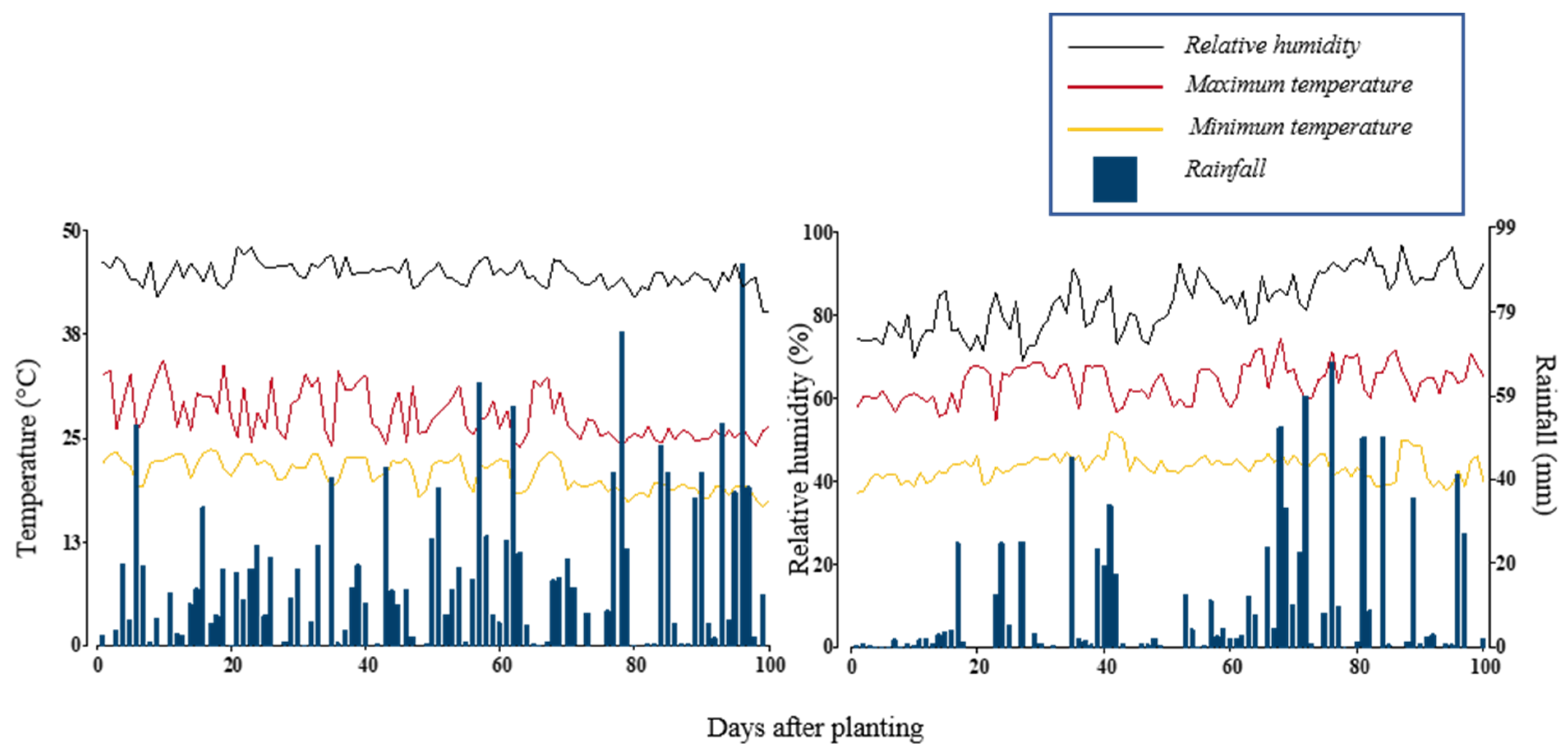
4.1. Experimental Site and Meteorological Conditions
4.2. Plant Material and Experimental Design
4.3. Selection of Infected and Healthy Leaves for Physiological Evaluation
4.4. Chlorophyll (Chla) Fluorescence and Imaging for the Chla Parameters
4.5. Physiological, Environmental and Agronomic Performance Variables
4.6. Developing and Validating the Disease Susceptibility Index
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nay, M.M.; Mukankusi, C.M.; Studer, B.; Raatz, B. Haplotypes at the Phg-2 Locus Are Determining Pathotype-Specificity of Angular Leaf Spot Resistance in Common Bean. Front. Plant Sci. 2019, 10, 1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmutz, J.; McClean, P.E.; Mamidi, S.; Wu, G.A.; Cannon, S.B.; Grimwood, J.; Jenkins, J.; Shu, S.; Song, Q.; Chavarro, C.; et al. A reference genome for common bean and genome-wide analysis of dual domestications. Nat. Genet. 2014, 46, 707–713. [Google Scholar] [CrossRef] [Green Version]
- Beebe, S.E.; Rao, I.M.; Blair, M.W.; Acosta-Gallegos, J.A. Phenotyping common beans for adaptation to drought. Front. Physiol. 2013, 4, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperdouli, I.; Mellidou, I.; Moustakas, M. Harnessing Chlorophyll Fluorescence for Phenotyping Analysis of Wild and Cultivated Tomato for High Photochemical Efficiency under Water Deficit for Climate Change Resilience. Climate 2021, 9, 154. [Google Scholar] [CrossRef]
- Bassi, D.; Briñez, B.; Rosa, J.S.; Oblessuc, P.R.; Almeida, C.; Nucci, S.M.; Da Silva, L.C.D.; Chiorato, A.F.; Vianello, R.P.; Camargo, L.E.A.; et al. Linkage and mapping of quantitative trait loci associated with angular leaf spot and powdery mildew resistance in common beans. Genet. Mol. Biol. 2017, 40, 109–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, J.; Solarte, D.; Lobaton, J.D.; Mayor, V.; Barrera, S.; Jara, C.; Beebe, S.; Raatz, B. Fine-mapping of angular leaf spot resistance gene Phg-2 in common bean and development of molecular breeding tools. Theor. Appl. Genet. 2019, 132, 2003–2016. [Google Scholar] [CrossRef] [Green Version]
- de Almeida, C.E.; Bonfante, G.F.; de Carvalho Paulino, J.F.; de Deus, B.C.; Patricio, F.A.; Carbonell, S.E.; Chiorato, A.F.; Benchimol-Reis, L.L. Precise assessment of angular leaf spot severity using the Leaf Doctor app for common beans. African J. Biotechnol. 2021, 20, 169–174. [Google Scholar]
- Bitocchi, E.; Nanni, L.; Bellucci, E.; Rossi, M.; Giardini, A.; Zeuli, P.S.; Logozzo, G.; Stougaard, J.; McClean, P.; Attene, G.; et al. Mesoamerican origin of the common bean (Phaseolus vulgaris L.) is revealed by sequence data. Proc. Natl. Acad. Sci. USA 2012, 109, E788–E796. [Google Scholar] [CrossRef] [Green Version]
- Guzmán, P.; Gilbertson, R.L.; Nodari, R.; Johnson, W.C.; Temple, S.R.; Mandala, D.; Mkandawire, A.B.; Gepts, P. Characterization of variability in the fungus Phaeoisariopsis griseola suggests coevolution with the common bean (Phaseolus vulgaris). Phytopathology 1995, 85, 600–607. [Google Scholar] [CrossRef]
- Contreras-Medina, L.M.; Osornio-Rios, R.A.; Torres-Pacheco, I.; Romero-Troncoso, R.D.J.; Guevara-González, R.G.; Millan-Almaraz, J.R. Smart Sensor for Real-Time Quantification of Common Symptoms Present in Unhealthy Plants. Sensors 2012, 12, 784–805. [Google Scholar] [CrossRef] [Green Version]
- Bock, C.H.; Parker, P.E.; Cook, A.Z.; Gottwald, T.R. Characteristics of the Perception of Different Severity Measures of Citrus Canker and the Relationships Between the Various Symptom Types. Plant Dis. 2008, 92, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Van Schoonhoven, A.; Corrales, M.A.P. Sistema Estándar Para la Evaluación de Germoplasma de Fríjol. Cali (Colombia); Centro Internacional de Agricultura Tropical: Cali, Colombia, 1987; 56p. [Google Scholar]
- Tung, J.; Goodwin, P.H.; Hsiang, T. Chlorophyll fluorescence for quantification of fungal foliar infection and assessment of the effectiveness of an induced systemic resistance activator. Eur. J. Plant Pathol. 2013, 136, 301–315. [Google Scholar] [CrossRef]
- Rousseau, C.; Belin, E.; Bove, E.; Rousseau, D.; Fabre, F.; Berruyer, R.; Guillaumès, J.; Manceau, C.; Jacques, M.-A.; Boureau, T. High throughput quantitative phenotyping of plant resistance using chlorophyll fluorescence image analysis. Plant Methods 2013, 9, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamers, J.; van der Meer, T.; Testerink, C. How Plants Sense and Respond to Stressful Environments. Plant Physiol. 2020, 182, 1624–1635. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, Z.; Iqbal, M.S.; Hashem, A.; Abd_Allah, E.F.; Ansari, M.I. Plant Defense Responses to Biotic Stress and Its Interplay with Fluctuating Dark/Light Conditions. Front. Plant Sci. 2021, 12, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Sheng, L.; Zhang, G.; Gu, Q.; Zheng, K. Influence of bacterial leaf blight on the photosynthetic characteristics of resistant and susceptible rice. J. Phytopathol. 2018, 166, 547–554. [Google Scholar] [CrossRef]
- Bilgin, D.D.; Zavala, J.A.; Zhu, J.; Clough, S.J.; Ort, D.R.; De Lucia, E.H. Biotic stress globally downregulates photosynthesis genes. Plant Cell Environ. 2010, 33, 1597–1613. [Google Scholar] [CrossRef] [Green Version]
- Aucique-Perez, C.E.; Daza, E.S.; Ávila-Diazgranados, R.A.; Romero, H.M. Chlorophyll a fluorescence and leaf temperature are early indicators of oil palm diseases. Sci. Agricola 2020, 77, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Wang, M.; Ling, N.; Shen, Q.; Guo, S. Potential role of photosynthesis-related factors in banana metabolism and defense against Fusarium oxysporum f. sp. cubense. Environ. Exp. Bot. 2016, 129, 4–12. [Google Scholar] [CrossRef]
- de Ribou, S.d.B.; Douam, F.; Hamant, O.; Frohlich, M.W.; Negrutiu, I. Plant science and agricultural productivity: Why are we hitting the yield ceiling? Plant Sci. 2013, 210, 159–176. [Google Scholar] [CrossRef]
- Bueno, A.C.S.O.; Castro, G.L.S.; Junior, D.D.S.; Pinheiro, H.A.; Filippi, M.C.C.; Silva, G.B. Response of photosynthesis and chlorophyll a fluorescence in leaf scald-infected rice under influence of rhizobacteria and silicon fertilizer. Plant Pathol. 2017, 66, 1487–1495. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, T.; Feng, B.; Zhang, C.; Peng, S.; Zhang, X.; Fu, G.; Tao, L. Non-photochemical Quenching Plays a Key Role in Light Acclimation of Rice Plants Differing in Leaf Color. Front. Plant Sci. 2017, 7, 1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, D.A.; Bernards, M.A. Chlorophyll fluorescence imaging as a tool to monitor the progress of a root pathogen in a perennial plant. Planta 2015, 243, 263–279. [Google Scholar] [CrossRef] [PubMed]
- Rios, J.; Aucique-Pérez, C.; Debona, D.; Neto, L.C.; Rios, V.; Rodrigues, F. Changes in leaf gas exchange, chlorophyll a fluorescence and antioxidant metabolism within wheat leaves infected by Bipolaris sorokiniana. Ann. Appl. Biol. 2016, 170, 189–203. [Google Scholar] [CrossRef]
- Zlatev, Z.; Petrova, Z.; Penchev, E. Changes in the Chlorophyll fluorescence of common winter wheat depending on the weed infestation and the date of application of a set of herbicides. Agric. Sci. 2016, VIII, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Lehto, K.; Tikkanen, M.; Hiriart, J.-B.; Paakkarinen, V.; Aro, E.-M. Depletion of the Photosystem II Core Complex in Mature Tobacco Leaves Infected by the Flavum Strain of Tobacco mosaic virus. Mol. Plant-Microbe Interact. 2003, 16, 1135–1144. [Google Scholar] [CrossRef] [Green Version]
- Almon, E.; Horowitz, M.; Wang, H.L.; Lucas, W.J.; Zamski, E.; Wolf, S. Phloem-Specific Expression of the Tobacco Mosaic Virus Movement Protein Alters Carbon Metabolism and Partitioning in Transgenic Potato Plants. Plant Physiol. 1997, 115, 1599–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otulak, K.; Chouda, M.; Bujarski, J.; Garbaczewska, G. The evidence of Tobacco rattle virus impact on host plant organelles ultrastructure. Micron 2015, 70, 7–20. [Google Scholar] [CrossRef]
- Rajendran, D.K.; Park, E.; Nagendran, R.; Hung, N.B.; Cho, B.-K.; Kim, K.-H.; Lee, Y.H. Visual Analysis for Detection and Quantification of Pseudomonas cichorii Disease Severity in Tomato Plants. Plant Pathol. J. 2016, 32, 300–310. [Google Scholar] [CrossRef] [Green Version]
- Berdugo, C.A.; Zito, R.; Paulus, S.; Mahlein, A.-K. Fusion of sensor data for the detection and differentiation of plant diseases in cucumber. Plant Pathol. 2014, 63, 1344–1356. [Google Scholar] [CrossRef]
- Hu, H.; Tang, M.; Guo, H.; Zhang, G.; Gu, Q.; Yuan, Y.; Sheng, L.; Zhou, H.; Liu, Z. Photosynthetic rate, chlorophyll fluorescence in anthracnose infected tea plants. Int. J. Agric. Biol. 2020, 24, 531–537. [Google Scholar]
- Mahlein, A.-K.; Oerke, E.-C.; Steiner, U.; Dehne, H.-W. Recent advances in sensing plant diseases for precision crop protection. Eur. J. Plant Pathol. 2012, 133, 197–209. [Google Scholar] [CrossRef]
- Banks, J.M. Continuous excitation chlorophyll fluorescence parameters: A review for practitioners. Tree Physiol. 2017, 37, 1128–1136. [Google Scholar] [CrossRef]
- Nath, K.; O’Donnell, J.P.; Lu, Y. Chlorophyll Fluorescence for High-Throughput Screening of Plants During Abiotic Stress, Aging, and Genetic Perturbation. In Photosynthesis: Structures, Mechanisms, and Applications; Springer: Cham, Switzerland, 2017; pp. 261–273. [Google Scholar] [CrossRef]
- Moustakas, M.; Calatayud, Á.; Guidi, L. Editorial: Chlorophyll Fluorescence Imaging Analysis in Biotic and Abiotic Stress. Front. Plant Sci. 2021, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chaerle, L.; Hagenbeek, D.; De Bruyne, E.; Van Der Straeten, M.; Straeten, D. Chlorophyll fluorescence imaging for disease-resistance screening of sugar beet. Plant Cell Tissue Organ Cult. (PCTOC) 2007, 91, 97–106. [Google Scholar] [CrossRef]
- Naumann, J.C.; Young, D.R.; Anderson, J.E. Leaf chlorophyll fluorescence, reflectance, and physiological response to freshwater and saltwater flooding in the evergreen shrub, Myrica cerifera. Environ. Exp. Bot. 2008, 63, 402–409. [Google Scholar] [CrossRef]
- Legendre, R.; Basinger, N.; van Iersel, M. Low-Cost Chlorophyll Fluorescence Imaging for Stress Detection. Sensors 2021, 21, 2055. [Google Scholar] [CrossRef]
- Pérez-Bueno, M.L.; Pineda, M.; Barón, M. Phenotyping Plant Responses to Biotic Stress by Chlorophyll Fluorescence Imaging. Front. Plant Sci. 2019, 10, 1135. [Google Scholar] [CrossRef]
- Mandal, K.; Saravanan, R.; Maiti, S.; Kothari, I.L. Effect of downy mildew disease on photosynthesis and chlorophyll fluorescence in Plantago ovata Forsk. J. Plant Dis. Prot. 2009, 116, 164–168. [Google Scholar] [CrossRef]
- Guidi, L.; Lo Piccolo, E.; Landi, M. Chlorophyll fluorescence, photoinhibition and abiotic stress: Does it make any difference the fact to be a C3 or C4 species? Front. Plant Science. Front. Media 2019, 10, 174. [Google Scholar] [CrossRef]
- Cen, H.; Weng, H.; Yao, J.; He, M.; Lv, J.; Hua, S.; Li, H.-Y.; He, Y. Chlorophyll Fluorescence Imaging Uncovers Photosynthetic Fingerprint of Citrus Huanglongbing. Front. Plant Sci. 2017, 8, 1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gortari, F.; Guiamet, J.J.; Graciano, C. Plant–pathogen interactions: Leaf physiology alterations in poplars infected with rust (Melampsora medusae). Tree Physiol. 2018, 38, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Yordanov, I.; Tsonev, T.; Goltsev, V.; Kruleva, L.; Velikova, V. Interactive effect of water deficit and high temperature on photosynthesis of sunflower and maize plants 1. Changes in parameters of chlorophyll fluorescence induction kinetics and fluorescence quenching. Photosynthetica 1997, 33, 391–402. [Google Scholar]
- Kalaji, H.M.; Schansker, G.; Brestic, M.; Bussotti, F.; Calatayud, A.; Ferroni, L.; Goltsev, V.; Guidi, L.; Jajoo, A.; Li, P.; et al. Frequently asked questions about chlorophyll fluorescence, the sequel. Photosynth. Res. 2016, 132, 13–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Botany. Oxf. Acad. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [Green Version]
- Suárez, J.C.; Urban, M.O.; Contreras, A.T.; Noriega, J.E.; Deva, C.; Beebe, S.E.; Polanía, J.A.; Casanoves, F.; Rao, I.M. Water Use, Leaf Cooling and Carbon Assimilation Efficiency of Heat Resistant Common Beans Evaluated in Western Amazonia. Front. Plant Sci. 2021, 12, 1–17. [Google Scholar] [CrossRef]
- Rodríguez-Moreno, L.; Pineda, M.; Soukupová, J.; Macho, A.P.; Beuzón, C.R.; Barón, M.; Ramos, C. Early detection of bean infection by Pseudomonas syringae in asymptomatic leaf areas using chlorophyll fluorescence imaging. Photosynth. Res. 2007, 96, 27–35. [Google Scholar] [CrossRef]
- Glowacka, K.; Kromdijk, J.; Kucera, K.; Xie, J.; Cavanagh, A.; Leonelli, L.; Leakey, A.D.B.; Ort, D.R.; Niyogi, K.K.; Long, S.P. Photosystem II Subunit S overexpression increases the efficiency of water use in a field-grown crop. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Barón, M.; Pineda, M.; Pérez-Bueno, M.L. Picturing pathogen infection in plants. Z. Fur Nat. Sect C J. Biosci. 2016, 71, 355–3568. [Google Scholar] [CrossRef]
- Fernández-Calleja, M.; Monteagudo, A.; Casas, A.M.; Boutin, C.; Pin, P.A.; Morales, F.; Igartua, E. Rapid On-Site Phenotyping via Field Fluorimeter Detects Differences in Photosynthetic Performance in a Hybrid—Parent Barley Germplasm Set. Sensors 2020, 20, 1486. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, U.; Klughammer, C.; Walz, H. Complementary PS II Quantum Yields Calculated from Simple Fluorescence Parameters Measured by PAM Fluorometry and the Saturation Pulse Method In Vivo Redox Changes of Ferredoxin, P700 and Plastocyanin View Project Assessment of Wavelength-Dependent Properties of Photosynthesis by Chlorophyll Fluorescence Measurements View Project Complementary PS II Quantum Yields Calculated from Simple Fluorescence Parameters Measured by PAM Fluorometry and the Saturation Pulse Method. 2008, Volume 1. Available online: http://www.walz.com/http://www.walz.com/ (accessed on 22 April 2022).
- Yang, H.; Luo, P. Changes in Photosynthesis Could Provide Important Insight into the Interaction between Wheat and Fungal Pathogens. Int. J. Mol. Sci. 2021, 22, 8865. [Google Scholar] [CrossRef]
- Serrano, I.; Audran, C.; Rivas, S. Chloroplasts at work during plant innate immunity. J. Exp. Bot. 2016, 67, 3845–3854. [Google Scholar] [CrossRef]
- Francesconi, S.; Balestra, G.M. The modulation of stomatal conductance and photosynthetic parameters is involved in Fusarium head blight resistance in wheat. PLoS ONE 2020, 15, e0235482. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, C.P.; Paulino, J.F.D.C.; Barbosa, C.C.F.; Gonçalves, G.D.M.C.; Fritsche-Neto, R.; Carbonell, S.A.M.; Chiorato, A.F.; Benchimol-Reis, L.L. Genome-wide association mapping reveals race-specific SNP markers associated with anthracnose resistance in carioca common beans. PLoS ONE 2021, 16, e0251745. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Leon-Reyes, A.; Van der Ent, S.; Van Wees, S.C.M. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, J.C.S.; Polanía, J.A.; Bastidas, A.T.C.; Suárez, L.R.; Beebe, S.; Rao, I.M. Agronomical, phenological and physiological performance of common bean lines in the Amazon region of Colombia. Theor. Exp. Plant Physiol. 2018, 30, 303–320. [Google Scholar] [CrossRef]
- Suárez, J.C.; Polanía, J.A.; Anzola, J.A.; Contreras, A.T.; Méndez, D.L.; Vanegas, J.I.; Noriega, J.E.; Rodríguez, L.; Urban, M.O.; Beebe, S.; et al. Influence of nitrogen supply on gas exchange, chlorophyll fluorescence and grain yield of breeding lines of common bean evaluated in the Amazon region of Colombia. Acta Physiol. Plant. 2021, 43, 1–15. [Google Scholar] [CrossRef]
- Chaerle, L.; Leinonen, I.; Jones, H.; Van Der Straeten, D. Monitoring and screening plant populations with combined thermal and chlorophyll fluorescence imaging. J. Exp. Bot. 2006, 58, 773–784. [Google Scholar] [CrossRef] [Green Version]
- Barczak-Brzyżek, A.K.; Kiełkiewicz, M.; Gawroński, P.; Kot, K.; Filipecki, M.; Karpińska, B. Cross-talk between high light stress and plant defence to the two-spotted spider mite in Arabidopsis thaliana. Exp. Appl. Acarol. 2017, 73, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Reinoso, A.D.; Ligarreto-Moreno, G.A.; Restrepo-Díaz, H. Physiological and biochemical expressions of a determinated growth common bean genotype (Phaseolus vulgaris L.) to water deficit stress periods. J. Anim. Plant Sci. 2018, 28, 119–127. [Google Scholar]
- Wang, L.; Poque, S.; Valkonen, J.P.T. Phenotyping viral infection in sweetpotato using a high-throughput chlorophyll fluorescence and thermal imaging platform. Plant Methods 2019, 15, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Xu, H.; Hao, H.; An, S.; Lu, C.; Wu, R.; Su, W. Effects of bensulfuron-methyl residue on photosynthesis and chlorophyll fluorescence in leaves of cucumber seedlings. PLoS ONE 2019, 14, e0215486. [Google Scholar] [CrossRef] [Green Version]
- Sofo, A. Drought stress tolerance and photoprotection in two varieties of olive tree. Acta Agric. Scand. Sect. B Soil Plant Sci. 2011, 61, 711–720. [Google Scholar] [CrossRef]
- Song, X.; Zhou, G.; Ma, B.; Wu, W.; Ahmad, I.; Zhu, G.; Yan, W.; Jiao, X. Nitrogen Application Improved Photosynthetic Productivity, Chlorophyll Fluorescence, Yield and Yield Components of Two Oat Genotypes under Saline Conditions. Agronomy 2019, 9, 115. [Google Scholar] [CrossRef] [Green Version]
- Mehta, P.; Jajoo, A.; Mathur, S.; Bharti, S. Chlorophyll a fluorescence study revealing effects of high salt stress on Photosystem II in wheat leaves. Plant Physiol. Biochem. 2010, 48, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Suancha, S.C.; Álvarado, S.O.H.; Balaguera-López, H.E. Fluorescencia como indicador de estrés en Helianthus annuus L. Una revisión. Rev. Colomb. Cienc. Hortícolas 2015, 9, 149. [Google Scholar] [CrossRef] [Green Version]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [Green Version]
- Aro, E.-M.; Virgin, I.; Andersson, B. Photoinhibition of Photosystem II. Inactivation, protein damage and turnover. Biochim. et Biophys. Acta 1993, 1143, 113–134. [Google Scholar] [CrossRef]
- Osmond, C.B. What is photoinhibition? Some insights from comparisons of shade and sun plants. In Photoinhibition of Photosynthesis from Molecular Mechanisms to the Field; Oxford, UK, 1994; pp. 1–19. Available online: https://www.semanticscholar.org/paper/What-is-photoinhibition-Some-insights-from-of-shade-Osmond/211682b35071523d7d4d2d0b39a25f945f600ec5 (accessed on 17 March 2022).
- Peña-Olmos, J.E.; Casierra-Posada, F. Photochemical efficiency of photosystem II (PSII) in broccoli plants (Brassica oleracea var Italica) affected by excess iron. Orinoquia 2013, 17, 15–22. [Google Scholar] [CrossRef]
- Hou, S.; Hou, Z.; Ye, J. Cotton Growth and Nitrogen Uptake in Response to Rates of Water and Nitrogen Under drip Irrigation with Saline Water. Xinjiang Agric Sci. 2010, 42, 1882–1887. [Google Scholar]
- Fracheboud, Y.; Leipner, J. The Application of Chlorophyll Fluorescence to Study Light, Temperature, and Drought Stress; Springer: Boston, MA, USA, 2003; pp. 125–150. [Google Scholar] [CrossRef]
- Suárez, J.C.; Polanía, J.A.; Contreras, A.T.; Rodríguez, L.; Machado, L.; Ordoñez, C.; Beebe, S.; Rao, I.M. Adaptation of common bean lines to high temperature conditions: Genotypic differences in phenological and agronomic performance. Euphytica 2020, 216, 1–20. [Google Scholar] [CrossRef]
- Wang, Y.; Bao, Z.; Zhu, Y.; Hua, J. Analysis of Temperature Modulation of Plant Defense Against Biotrophic Microbes. Mol. Plant-Microbe Interact. 2009, 22, 498–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of Combined Abiotic and Biotic Stresses on Plant Growth and Avenues for Crop Improvement by Exploiting Physio-morphological Traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef] [Green Version]
- Godoy-Lutz, G.; Kuninaga, S.; Steadman, J.R.; Powers, K. Phylogenetic analysis of Rhizoctonia solani subgroups associated with web blight symptoms on common bean based on ITS-5.8S rDNA. J. Gen. Plant Pathol. 2007, 74, 32–40. [Google Scholar] [CrossRef]
- Prabhu, A.S.; Silva, J.F.; Correa, J.R.; Polaro, R.H.; Lima, E.F. Murcha da teia micélica do feijoeiro comum: Epidemiologia e aplicação de fungicidas. Pesqui Agropecu Bras 1983, 18, 1323–1332. [Google Scholar]
- Villamar-Torres, R.; Montiel, L.G.; MuÃoz-Rengifo, J.; Montes, S.Z.; Jazayeri, S.M.; Maddela, N.R.; Torres, L.A.; Matute, A.M.; Pinos, M.H.; Salia, O.I.; et al. Agronomic evaluation and web blight resilience of common bean genotypes in the littoral region of Ecuador. African J. Biotechnol. 2018, 17, 328–336. [Google Scholar]
- Costa-Coelho, G.R.; Filho, A.C.C.; Lobo, M. A comparison of web blight epidemics on common bean cultivars with different growth habits. Crop Prot. 2014, 55, 16–20. [Google Scholar] [CrossRef]
- Ogawa, T.; Sonoike, K. Screening of mutants using chlorophyll fluorescence. J. Plant Res. 2021, 134, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New Fluorescence Parameters for the Determination of QARedox State and Excitation Energy Fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef]
- Brooks, M.D.; Niyogi, K.K. Use of a Pulse-Amplitude Modulated Chlorophyll Fluorometer to Study the Efficiency of Photosynthesis in Arabidopsis Plants. In Chloroplast Research in Arabidopsis; Jarvis, R.P., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2011; Volume 775, pp. 299–310. ISBN 978-1-61779-236-6. [Google Scholar]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Ritchie, R.J. Fitting light saturation curves measured using modulated fluorometry. Photosynth. Res. 2008, 96, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Gupta, C.; Gupta, M.; Gupta, S.; Salgotra, R. Screening of common bean (Phaseolus vulgaris L.) germplasm against Colletotrichum lindemuthianum inciting bean anthracnose. Res. J. Biotechnol. 2021, 7, 13–18. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; ISBN 3-900051-07-0. Available online: http://www.R-project.org/ (accessed on 17 March 2022).
- Di Rienzo, J.; Casanoves, F.; Balzarini, M.; González, L. InfoStat Versión 2019; Centro de Transferencia InfoStat, FCA, Universidad Nacional de Córdoba: Córdoba, Argentina, 2019; Available online: http://www.infostat.com.ar (accessed on 17 March 2022).











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Moderately Resistant | Moderately Susceptible | Susceptible | Highly Susceptible | p-Value |
|---|---|---|---|---|---|
| Disease susceptibility index (DSI) | 0.16 ± 0.02 a | 0.33 ± 0.01 b | 0.48 ± 0.01 c | 0.69 ± 0.02 d | <0.0001 |
| Initial fluorescence (F0) | 0.02 ± 0.01 a | 0.02 ± 0.01 a | 0.02 ± 0.01 a | 0.01 ± 0.01 b | 0.0028 |
| Maximum fluorescence (Fm) | 0.04 ± 0.01 a | 0.04 ± 0.01 a | 0.03 ± 0.01 b | 0.02 ± 0.01 c | <0.0001 |
| Effective quantum yield of photochemical energy conversion in PSII (Y(II)) | 0.15 ± 0.02 a | 0.12 ± 0.01 a | 0.09 ± 0.01 b | 0.08 ± 0.02 b | 0.0008 |
| Quantum yield of regulated energy dissipation (Y(NPQ)) | 0.13 ± 0.02 b | 0.15 ± 0.01 b | 0.19 ± 0.01 a | 0.21 ± 0.02 a | 0.0003 |
| Quantum yield of non-regulated energy dissipation (Y(NO)) | 0.13 ± 0.01 | 0.13 ± 0.01 | 0.12 ± 0.01 | 0.12 ± 0.02 | |
| Non-photochemical quenching (NPQ) | 0.25 ± 0.02 | 0.23 ± 0.01 | 0.21 ± 0.01 | 0.2 ± 0.02 | |
| Non-photochemical quenching coefficient (qN) | 0.25 ± 0.03 b | 0.28 ± 0.02 b | 0.35 ± 0.01 a | 0.36 ± 0.04 a | 0.0004 |
| Coefficient of photochemical quenching (qP) | 0.29 ± 0.02 a | 0.23 ± 0.01 b | 0.16 ± 0.01 c | 0.09 ± 0.01 d | <0.0001 |
| Coefficient of photochemical quenching (qL) | 0.17 ± 0.02 a | 0.14 ± 0.01 b | 0.12 ± 0.01 b | 0.07 ± 0.01 c | <0.0001 |
| Electron transport rate (ETR; μmol e− m−2 s−1) | 10.26 ± 0.93 a | 8.08 ± 0.65 a | 4.8 ± 0.41 b | 3.07 ± 0.67 b | <0.0001 |
| ETRmax (ETR maximum; μmol e− m−2 s−1) | 65.66 ± 9.59 a | 49.40 ± 6.18 b | 46.84 ± 5.33 b | 19.71 ± 4.18 c | <0.0001 |
| Maximum saturating irradiance (Ek; μmol m−2 s−1) | 458.1 ± 22.47 a | 365.31 ± 17.747 b | 358.17 ± 18.8 b | 163 ± 12.82 c | <0.0001 |
| Dark respiration rates (Rd; μmol CO2 m−2 s−1) | 7.49 ± 0.02 c | 9.6 ± 0.29 b | 10.27 ± 0.18 b | 23 ± 0.98 a | <0.0001 |
| Root biomass (g) | 1.05 ± 0.24 a | 2.1 ± 0.38 b | 1.4 ± 0.25 a | 1.35 ± 0.64 a | <0.0001 |
| Stem biomass (g) | 8.45 ± 0.12 a | 7.12 ± 0.94 a | 5.35 ± 0.7 b | 3.48 ± 0.48 c | <0.0001 |
| Leaf biomass (g) | 8.62 ± 0.29 a | 9.26 ± 0.61 a | 7.8 ± 0.23 c | 5.87 ± 0.15 c | <0.0001 |
| Flower bud biomass (g) | 0.01 ± 0.01 | 0.04 ± 0.01 | 0.01 ± 0.01 | ||
| Flower biomass (g) | 0.04 ± 0.04 | 0.34 ± 0.13 | 0.05 ± 0.02 | 0.1 ± 0.10 | |
| Pods biomass (g) | 4.6 ± 0.29 a | 3.36 ± 0.45 b | 0.72 ± 0.44 c | 0.34 ± 0.22 c | <0.0001 |
| Canopy biomass (CB, g) | 22.98 ± 2.01 a | 19.75 ± 2.38 a | 15.22 ± 1.93 b | 10.9 ± 1.25 c | <0.0001 |
| Variables | Variables | ||||
|---|---|---|---|---|---|
| DSI | LT | ETR | qP | NPQ | |
| Disease susceptibility index (DSI) | 0.32 * | −0.72 *** | −0.82 *** | −0.38 *** | |
| Initial fluorescence (F0) | −0.49 *** | −0.19 | 0.06 | 0 | 0.04 |
| Maximum fluorescence (Fm) | −0.66 *** | −0.39 ** | 0.39 *** | 0.27 * | 0.1 |
| Effective quantum yield of photochemical energy conversion in PSII (Y(II)) | −0.52 *** | −0.42 *** | 0.8 *** | 0.67 *** | 0.3 ** |
| Quantum yield of regulated energy dissipation (Y(NPQ)) | 0.53 *** | 0.07 | −0.69 *** | −0.48 *** | −0.88 *** |
| Quantum yield of non-regulated energy dissipation (Y(NO)) | −0.04 | 0.36 ** | −0.19 | −0.18 | 0.69 *** |
| Non-photochemical quenching (NPQ) | −0.38 *** | 0.1 | 0.41 *** | 0.23 * | |
| Non-photochemical cooling coefficient (qN) | 0.48 *** | 0.2 | −0.65 *** | −0.36 ** | −0.77 *** |
| Coefficient of photochemical quenching (qP) | −0.82 *** | −0.3 ** | 0.78 *** | 0.32 ** | |
| Coefficient of photochemical quenching (qL) | −0.57 *** | −0.09 | 0.32 ** | 0.8 *** | 0.1 |
| Electron transport rate (ETR, μmol e− m−2 s−1) | −0.72 *** | −0.37 ** | 0.77 *** | 0.41 *** | |
| ETRmax (ETR maximum; μmol e− m−2 s−1) | −0.13 | −0.09 | 0.31 * | 0.29 * | 0.07 |
| Maximum saturating irradiance (Ek; μmol m−2 s−1) | −0.15 | 0.04 | 0.31 * | 0.12 | 0.08 |
| Dark respiration rates (Rd; μmol CO2 m−2 s−1) | −0.22 | −0.05 | 0.01 | 0.17 | −0.02 |
| Light compensation point (LCP; μmol m−2 s−1) | −0.15 | −0.06 | −0.02 | 0.08 | −0.01 |
| Photosynthetic efficiency (α; μmol CO2 μmol protons−1) | 0.27 * | −0.03 | −0.33 ** | −0.12 | −0.22 |
| Chlorophyll content index (CCI; SPAD) | 0.13 | −0.41 ** | −0.13 | −0.15 | −0.09 |
| Leaf temperature (LT; °C) | 0.32 * | −0.37 ** | −0.38 ** | 0.1 | |
| Stomatal conductance (gs; μmol H2O m−2 s−1) | −0.13 | −0.23 | 0.17 | −0.03 | 0.01 |
| Ambient temperature (°C) | −0.19 | −0.9 *** | 0.27 * | 0.07 | −0.09 |
| Relative humidity (%) | −0.07 | −0.52 *** | 0.14 | −0.1 | 0 |
| Leaf temperature differential (LTD; °C) | −0.29 * | −0.98 *** | 0.35 ** | 0.19 | −0.08 |
| Root biomass (g) | 0.28 * | −0.1 | −0.13 | 0.05 | −0.31 * |
| Stem biomass (g) | −0.4 *** | −0.15 | 0.36 ** | 0.22 | 0 |
| Leaf biomass (g) | −0.2 | 0.04 | 0.2 | 0.03 | 0.04 |
| Flower bud biomass (g) | −0.19 | −0.07 | 0.17 | 0.15 | 0.15 |
| Flower biomass (g) | −0.1 | 0.07 | 0.01 | −0.01 | 0 |
| Pod biomass (g) | −0.29 * | −0.11 | 0.23 | 0.3 ** | 0.11 |
| Viable seeds (number) | −0.03 | −0.43 * | 0.27 | 0.01 | −0.01 |
| Non-viable seeds (number) | −0.03 | 0.36 * | −0.02 | −0.06 | 0.15 |
| Canopy biomass (CB, g) | −0.33 ** | −0.11 | 0.32 * | 0.15 | −0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suárez, J.C.; Vanegas, J.I.; Contreras, A.T.; Anzola, J.A.; Urban, M.O.; Beebe, S.E.; Rao, I.M. Chlorophyll Fluorescence Imaging as a Tool for Evaluating Disease Resistance of Common Bean Lines in the Western Amazon Region of Colombia. Plants 2022, 11, 1371. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101371
Suárez JC, Vanegas JI, Contreras AT, Anzola JA, Urban MO, Beebe SE, Rao IM. Chlorophyll Fluorescence Imaging as a Tool for Evaluating Disease Resistance of Common Bean Lines in the Western Amazon Region of Colombia. Plants. 2022; 11(10):1371. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101371
Chicago/Turabian StyleSuárez, Juan Carlos, José Iván Vanegas, Amara Tatiana Contreras, José Alexander Anzola, Milan O. Urban, Stephen E. Beebe, and Idupulapati M. Rao. 2022. "Chlorophyll Fluorescence Imaging as a Tool for Evaluating Disease Resistance of Common Bean Lines in the Western Amazon Region of Colombia" Plants 11, no. 10: 1371. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101371