
An Identification System Targeting the SRK Gene for Selecting S-Haplotypes and Self-Compatible Lines in Cabbage

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Self-Pollination and Identification of Self-Compatible Index
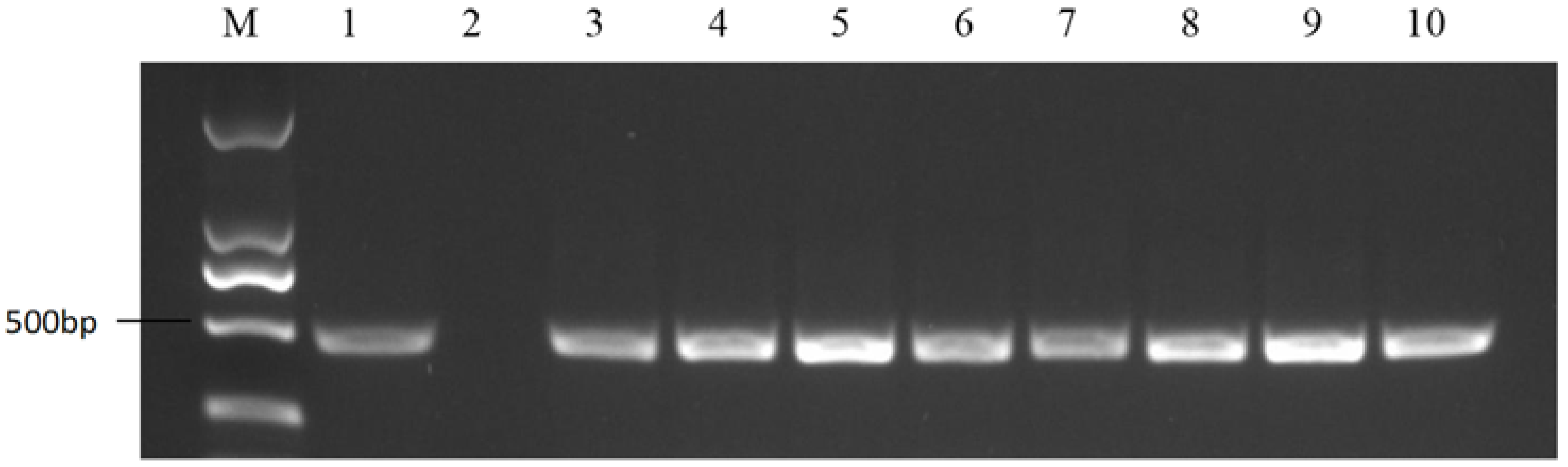
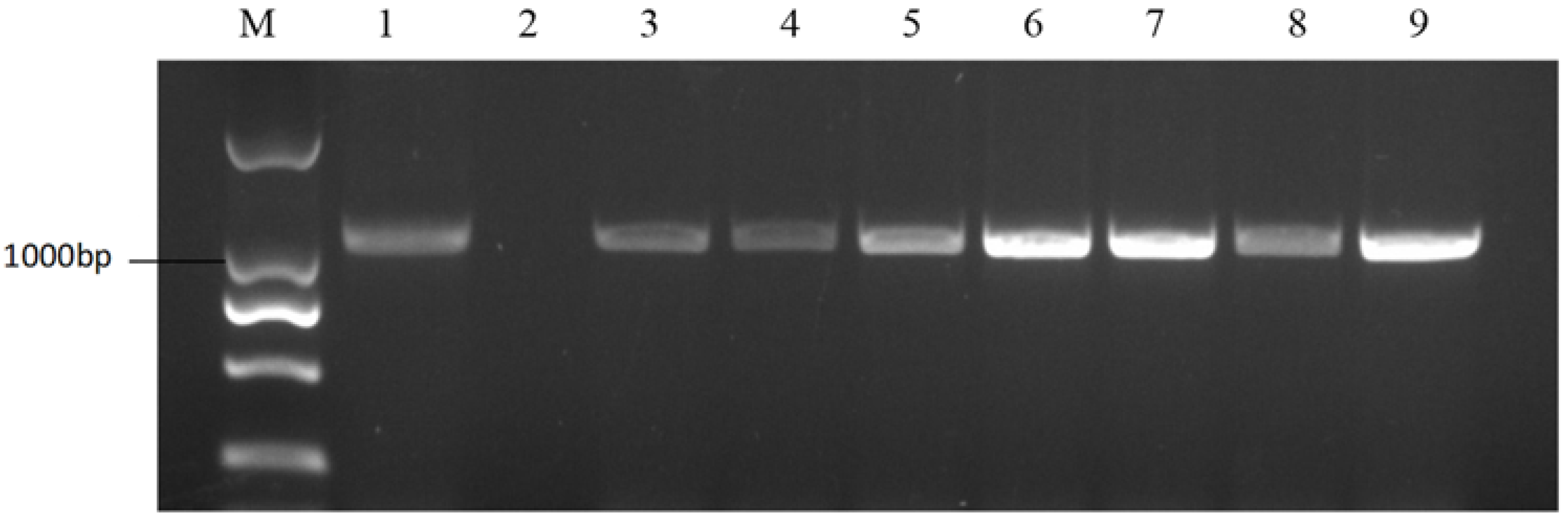
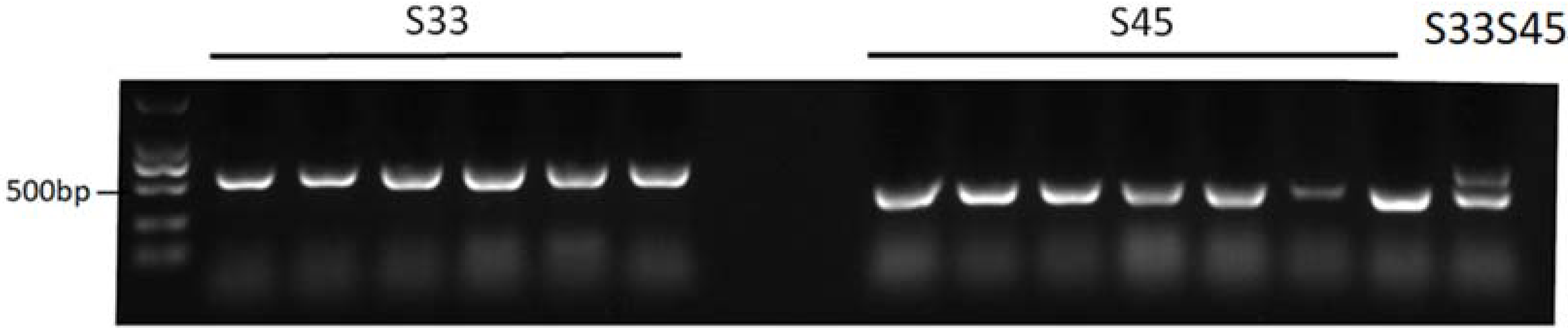
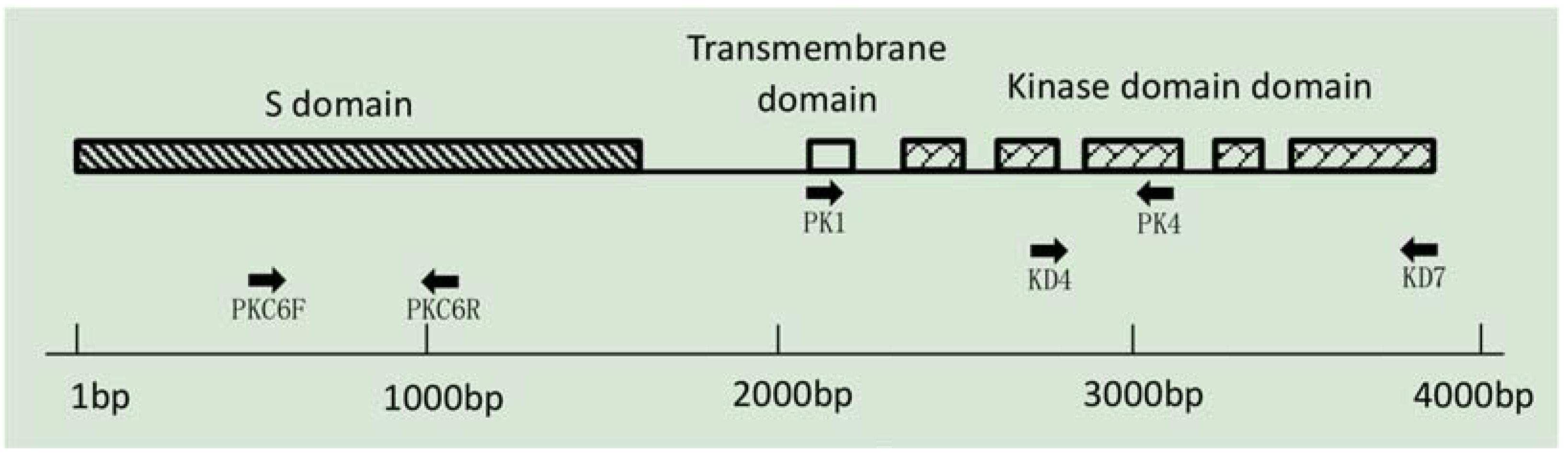
2.2. Marker Development for S-Haplotype Identification
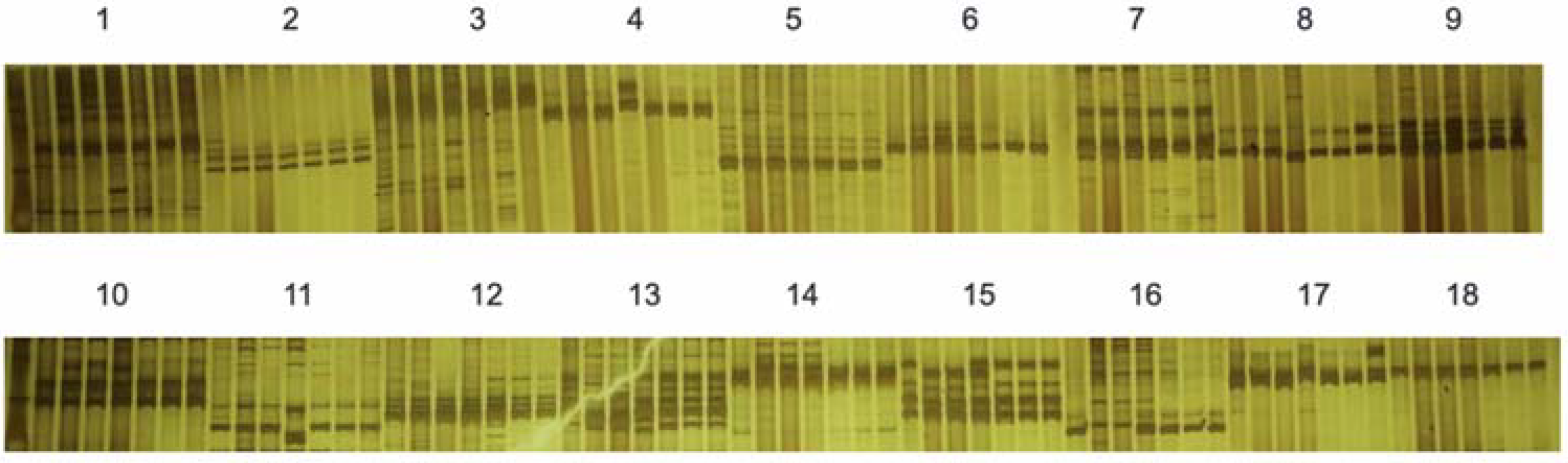
2.3. Identification of S-Haplotypes of 58 Cabbage Inbred Lines
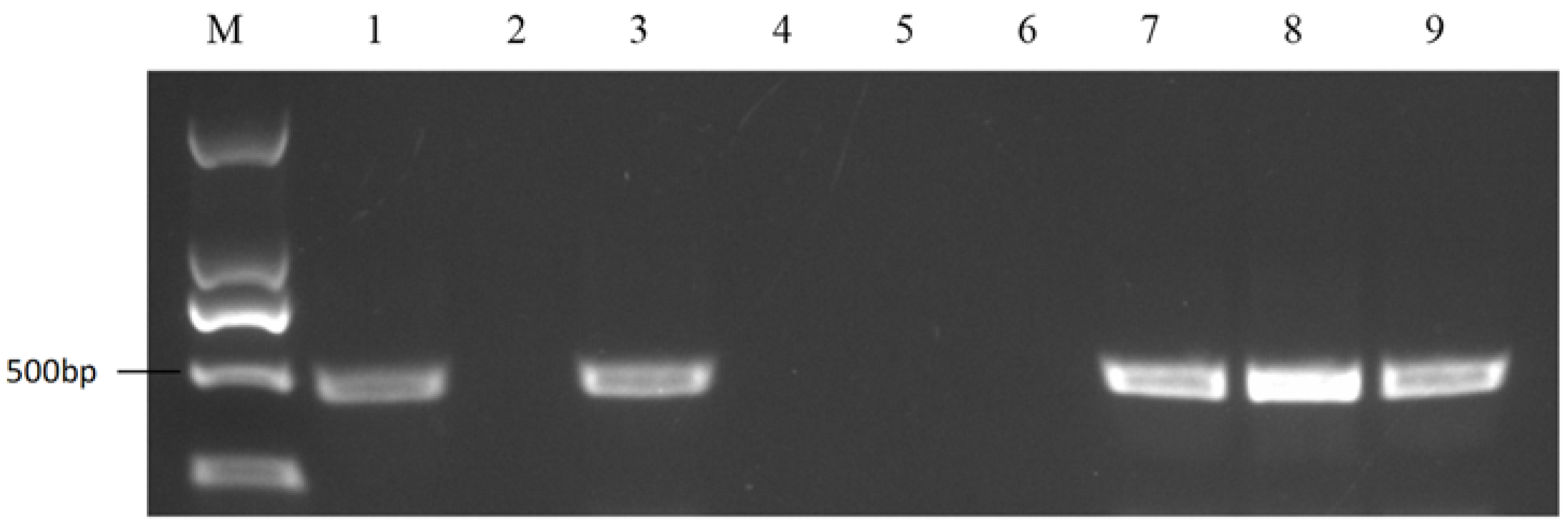
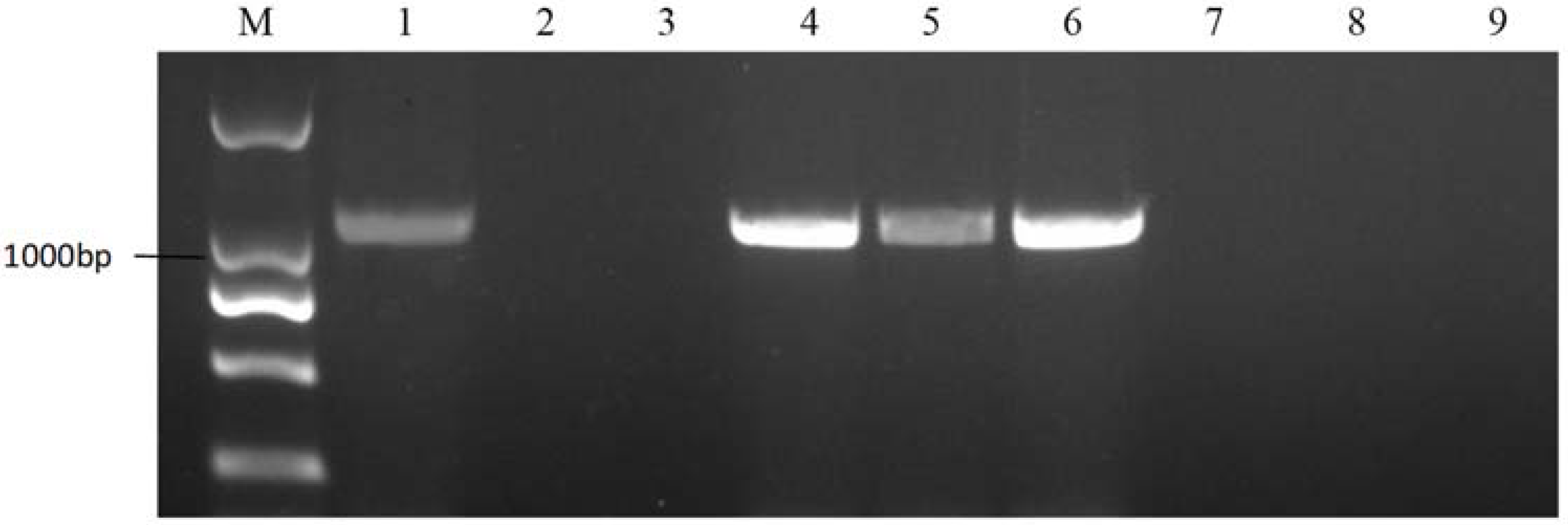
2.4. Self-Compatible Line Identification among the Sister Lines of 18-512
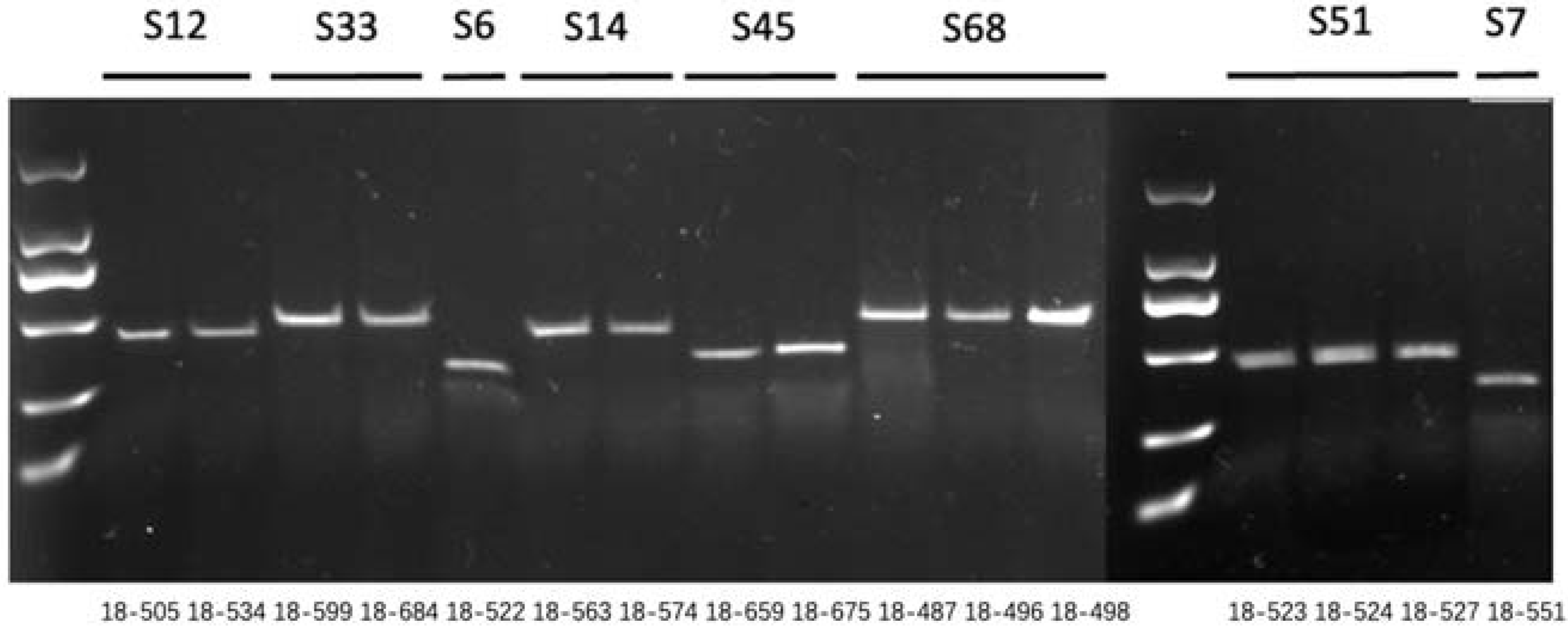
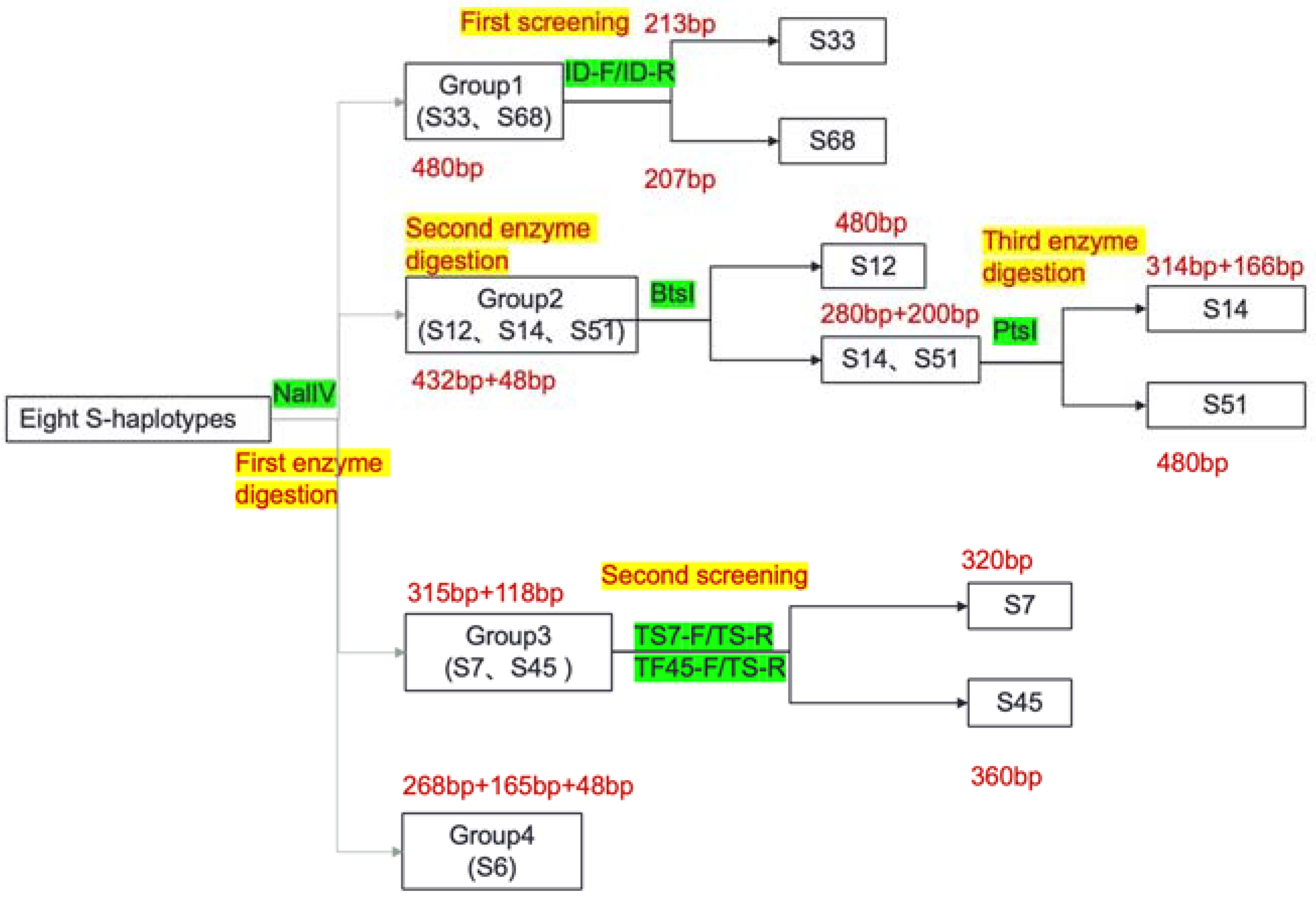
2.5. Development of the Class I S-Haplotype Primer Identification System
3. Discussion
3.1. Consistency of the Molecular Identification and Compatible Index (CI)
3.2. Distribution of S-Haplotypes in Winter Cabbage Materials
3.3. Comparison of Different S-Haplotypes Identification Methods
4. Materials and Methods
4.1. Plant Materials
4.2. Identification of Self-Compatible Index
4.3. Novel Marker Development for Class I S-Haplotype Identification
4.4. Identification of S-Haplotypes of 58 Cabbage Inbred Lines
4.5. Self-Compatible Line Screening among the Sister Lines of 18-512
4.6. Development of the Class I S-Haplotype Primer Identification System
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Nettancourt, D. Incompatibility and Incongruity Barriers Between Different Species. In Incompatibility and Incongruity in Wild and Cultivated Plants; Springer: Berlin/Heidelberg, Germany, 2001; pp. 217–251. [Google Scholar]
- Takayama, S.; Isogai, A. Self-incompatibility in plants. Annu. Rev. Plant Biol. 2005, 56, 467–489. [Google Scholar] [CrossRef] [Green Version]
- Hinata, K.; Watanabe, M.; Yamakawa, S.; Satta, Y.; Isogai, A. Evolutionary aspects of the S-related genes of the Brassica self-incompatibility system: Synonymous and nonsynonymous base substitutions. Genetics 1995, 140, 1099–1104. [Google Scholar] [CrossRef]
- Bateman, A. Self-incompatibility systems in angiosperms. Heredity 1952, 6, 285–310. [Google Scholar] [CrossRef] [Green Version]
- Nasrallah, J.B.; Nasrallah, M.E. Pollen-Stigma Signaling in the Sporophytic Self-Incompatibility Response. Plant Cell 1993, 5, 1325. [Google Scholar] [CrossRef]
- Nasrallah, J.; Kao, T.-H.; Goldberg, M.; Nasrallah, M. A cDNA clone encoding an S-locus-specific glycoprotein from Brassica oleracea. Nature 1987, 326, 523. [Google Scholar] [CrossRef] [Green Version]
- Thompson, K. Self-incompatibility in marrow-stem kale, Brassica oleracea var acephala I. Demonstration of a sporophytic system. J. Genet. 1957, 55, 45–60. [Google Scholar]
- Schopfer, C.R.; Nasrallah, M.E.; Nasrallah, J.B. The male determinant of self-incompatibility in Brassica. Science 1999, 286, 1697–1700. [Google Scholar] [CrossRef]
- Stein, J.C.; Howlett, B.; Boyes, D.C.; Nasrallah, M.E.; Nasrallah, J.B. Molecular cloning of a putative receptor protein kinase gene encoded at the self-incompatibility locus of Brassica oleracea. Proc. Natl. Acad. Sci. USA 1991, 88, 8816–8820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusaba, M.; Matsushita, M.; Okazaki, K.; Satta, Y.; Nishio, T. Sequence and structural diversity of the S locus genes from different lines with the same self-recognition specificities in Brassica oleracea. Genetics 2000, 154, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Takasaki, T.; Hatakeyama, K.; Suzuki, G.; Watanabe, M.; Isogai, A.; Hinata, K. The S receptor kinase determines self-incompatibility in Brassica stigma. Nature 2000, 403, 913–916. [Google Scholar] [CrossRef]
- Okazaki, K.; Kusaba, M.; Ockendon, D.; Nishio, T. Characterization of S tester lines in Brassica oleracea: Polymorphism of restriction fragment length of SLG homologues and isoelectric points of S-locus glycoproteins. Theor. Appl. Genet. 1999, 98, 1329–1334. [Google Scholar] [CrossRef]
- Ockendon, D. The S-allele collection of Brassica oleracea. In Proceedings of the III International Symposium on Brassicas and XII Crucifer Genetics Workshop, Wellesbourne, UK, 5–9 September 2000; pp. 25–30. [Google Scholar]
- Fang, Z.; Sun, P.; Liu, Y.; Yang, L.; Wang, X.; Hou, A.; Bian, C. A male sterile line with dominant gene (Ms) in cabbage (Brassica oleracea var. capitata) and its utilization for hybrid seed production. Euphytica 1997, 97, 265–268. [Google Scholar] [CrossRef]
- Brace, J.; King, G.J.; Ockendon, D. A molecular approach to the identification of S-alleles in Brassica oleracea. Sex. Plant Reprod. 1994, 7, 203–208. [Google Scholar] [CrossRef]
- Tian, L.; Miao, W.; Liu, J.; Fang, Z.; Liu, Y.; Yang, L.; Zhang, Y.; Zhuang, M. Identification of S haplotypes in cabbage inbred lines (Brassica oleracea var. capitata L.). Sci. Hortic. 2013, 164, 400–408. [Google Scholar] [CrossRef]
- Zheng, M.; Zhu, C.; Liu, M.; Xu, B.; Ma, C.; Li, Q.; Ren, X.; Si, J.; Song, H. Haplotype identification and validation of cabbage self-incompatible lines based on SRK gene sequence analysis. China Veg. 2018, 03, 32–39. [Google Scholar]
- Hu, J.; Xu, Q.; Liu, C.; Liu, B.; Deng, C.; Chen, C.; Wei, Z.; Ahmad, M.H.; Peng, K.; Wen, H. Downregulated expression of S2-RNase attenuates self-incompatibility in “Guiyou No. 1” pummelo. Hortic. Res. 2021, 8, 199. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, C.; Zhang, B.; Tang, F.; Li, F.; Liao, Q.; Tang, D.; Peng, Z.; Jia, Y.; Gao, M. A nonS-locus F-box gene breaks self-incompatibility in diploid potatoes. Nat. Commun. 2021, 12, 4142. [Google Scholar] [CrossRef]
- Park, J.; Lee, S.; Watanabe, M.; Takahata, Y.; Nou, I. Identification of S-alleles using polymerase chain reaction-cleaved amplified polymorphic sequence of the S-locus receptor kinase in inbreeding lines of Brassica oleracea. Plant Breed. 2002, 121, 192–197. [Google Scholar] [CrossRef]
- Qi-Guo, G.; Ming, S.; Yi, N.; Kun, Y.; Li-Quan, Z.; Xiao-Jia, W. Test of Interaction Between SRK and THL1 from Brassica oleracea L. in Self-Incompatibility Signaling Process. Acta Agron. Sin. 2008, 34, 934–943. [Google Scholar]
- Mazzurco, M.; Sulaman, W.; Elina, H.; Cock, J.M.; Goring, D.R. Further analysis of the interactions between the Brassica S receptor kinase and three interacting proteins (ARC1, THL1 and THL2) in the yeast two-hybrid system. Plant Mol. Biol. 2001, 45, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Murase, K.; Shiba, H.; Iwano, M.; Che, F.-S.; Watanabe, M.; Isogai, A.; Takayama, S. A membrane-anchored protein kinase involved in Brassica self-incompatibility signaling. Science 2004, 303, 1516–1519. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.L.; Arnoldo, M.; Goring, D.R. A breakdown of Brassica self-incompatibility in ARC1 antisense transgenic plants. Science 1999, 286, 1729–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takada, Y.; Sato, T.; Suzuki, G.; Shiba, H.; Takayama, S.; Watanabe, M. Involvement of MLPK pathway in intraspecies unilateral incompatibility regulated by a single locus with stigma and pollen factors. G3 Genes Genomes Genet. 2013, 3, 719–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Z.; Han, F.; Hu, Y.; Xue, Y.; Fang, Z.; Yang, L.; Zhang, Y.; Liu, Y.; Li, Z.; Wang, Y. Overcoming cabbage crossing incompatibility by the development and application of self-compatibility-QTL-specific markers and genome-wide background analysis. Front. Plant Sci. 2019, 10, 189. [Google Scholar] [CrossRef] [Green Version]
- Ockendon, D. An S-allele survey of cabbage (Brassica oleracea var. capitata). Euphytica 1982, 31, 325–331. [Google Scholar] [CrossRef]
- Ruffio-Chable, V.; Gaude, T. S-haplotype polymorphism in Brassica oleracea. In Proceedings of the International Symposium on Molecular Markers for Characterizing Genotypes and Identifying Cultivars in Horticulture, Montpellier, France, 6–8 March 2000; pp. 257–261. [Google Scholar]
- Ockendon, D. Distribution of self-incompatibility alleles and breeding structure of open-pollinated cultivars of Brussels sprouts. Heredity 1974, 33, 159–171. [Google Scholar] [CrossRef] [Green Version]
- Martin, F.W. Staining and observing pollen tubes in the style by means of fluorescence. Stain Technol. 1959, 34, 125–128. [Google Scholar] [CrossRef]
- Hartman, E.; Levy, C.; Kern, D.M.; Johnson, M.A.; Basu, A. A rapid, inexpensive, and semi-quantitative method for determining pollen tube extension using fluorescence. Plant Methods 2014, 10, 3. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, K.; Kusaba, M.; Nishio, T. Single-seed PCR-RFLP analysis for the identification of S haplotypes in commercial F1 hybrid cultivars of broccoli and cabbage. Plant Cell Rep. 2000, 19, 400–406. [Google Scholar] [CrossRef]
- Nishio, T.; Kusaba, M.; Sakamoto, K.; Ockendon, D. Polymorphism of the kinase domain of the S-locus receptor kinase gene (SRK) in Brassica oleracea L. Theor. Appl. Genet. 1997, 95, 335–342. [Google Scholar] [CrossRef]
- Nishio, T.; Sakamoto, K.; Yamaguchi, J. PCR-RFLP of S locus for identification of breeding lines in cruciferous vegetables. Plant Cell Rep. 1994, 13, 546–550. [Google Scholar] [CrossRef]
- Sakamoto, K.; Nishio, T. Distribution of S haplotypes in commercial cultivars of Brassica rapa. Plant Breed. 2001, 120, 155–161. [Google Scholar] [CrossRef]
- Doyle, J.J. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Haseyama, Y.; Kitashiba, H.; Okamoto, S.; Tonouchi, E.; Sakamoto, K.; Nishio, T. Nucleotide sequence analysis of S-locus genes to unify S haplotype nomenclature in radish. Mol. Breed. 2018, 38, 116. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S-Haplotype | GI Number in NCBI Database | PK1/PK4 | PKC6F/PKC6R |
|---|---|---|---|
| S2-b | AB298890 | - | 79.07% |
| S4 | M76647 | 95.56% | 95.35% |
| S5 | AB180898 | - | 74.42% |
| S6 | AB054708 | 100.00% | 95.35% |
| S7 | AB180901 | 95.56% | 95.35% |
| S8 | AB024420 | - | 97.67% |
| S12 | AB298891 | 93.33% | 100.00% |
| S13 | AB054710 | 95.56% | 97.67% |
| S14 | AB298892 | 97.78% | 97.67% |
| S15 | D85229 | - | 74.42% |
| S16 | AB013720 | - | 97.67% |
| S17 | AB054712 AB298894 | 97.78% | - |
| S22 | AB054713 AB298895 | - | 97.67% |
| S23 | AB054714 AB298897 | 95.56% | 93.02% |
| S24 | AB190355 | 88.89% | 95.35% |
| S25 | AB054715 | 95.56% | 96.67% |
| S28 | AB054716 | 95.56% | 96.67% |
| S33 | AB054718 AB298899 | 93.33% | 93.02% |
| S35 | AB054719 | - | 96.67% |
| S36 | AB054720 | - | 93.02% |
| S39 | AB298900 | - | 81.40% |
| S45 | AB298901 | - | 100.00% |
| S50 | AB054722 | - | 95.35% |
| S51 | AB298903 | - | 100.00% |
| S52 | AB054725 | 93.33% | 81.40% |
| S57 | AB298905 | - | 100.00% |
| S63 | AB024416 | 95.56% | - |
| S64 | Y18259 | - | 83.72% |
| S68 | AB180903 | 88.89% | 100.00% |
| Class | Primers | Sequences of Primers (5′-3′) | Tm/°C | Amplified Band |
|---|---|---|---|---|
| S7/S33 | TS33-F | CCCCGAAAACTTCCGGAGTT | 56 | S7: 320 bp S33: 600 bp |
| TS7-F | CGAACCACCAGTGCGATATGTACAG | |||
| TS-R | ACCATTCCGGATATCCGCATTT | |||
| S33/S45 | TF33-F | ATGGAATGAGGACTTTCCAATGCATC | 56 | S33: 573 bp S45: 364 bp |
| TF45-F | CCCCGAACTCAATTCAATGGAACT | |||
| TS-R | ACCATTCCGGATATCCGCATTT | |||
| S68/S33 | ID-F | CATGTAAGTGCAGGTAGCTC | 56 | S68: 207 bp |
| ID-R | CATGCATTTGAGCTTCCCACTA | S33: 213 bp |
| Class | Primers | Sequences of Primers (5′-3′) | Tm/°C | References |
|---|---|---|---|---|
| Class I | PKC6F | CAATTTCACAGAGAATAGTGA | 56 | This study |
| PKC6R | ACCATTCCGGATATCCGCATTT | |||
| Class II | KD4 | GAGGGCGAGAAGATCTTAATT | 60 | Park et al., 2002 [20] |
| KD7 | AAGACKATCATATTACCGAGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, W.; Zhang, B.; Ren, W.; Chen, L.; Fang, Z.; Yang, L.; Zhuang, M.; Lv, H.; Wang, Y.; Zhang, Y. An Identification System Targeting the SRK Gene for Selecting S-Haplotypes and Self-Compatible Lines in Cabbage. Plants 2022, 11, 1372. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101372
Chen W, Zhang B, Ren W, Chen L, Fang Z, Yang L, Zhuang M, Lv H, Wang Y, Zhang Y. An Identification System Targeting the SRK Gene for Selecting S-Haplotypes and Self-Compatible Lines in Cabbage. Plants. 2022; 11(10):1372. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101372
Chicago/Turabian StyleChen, Wendi, Bin Zhang, Wenjing Ren, Li Chen, Zhiyuan Fang, Limei Yang, Mu Zhuang, Honghao Lv, Yong Wang, and Yangyong Zhang. 2022. "An Identification System Targeting the SRK Gene for Selecting S-Haplotypes and Self-Compatible Lines in Cabbage" Plants 11, no. 10: 1372. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101372