
In Vitro Anti-Photoaging and Skin Protective Effects of Licania macrocarpa Cuatrec Methanol Extract

 , , , , , , ,
, , , , , , ,  and
and 
Abstract
:1. Introduction
2. Results
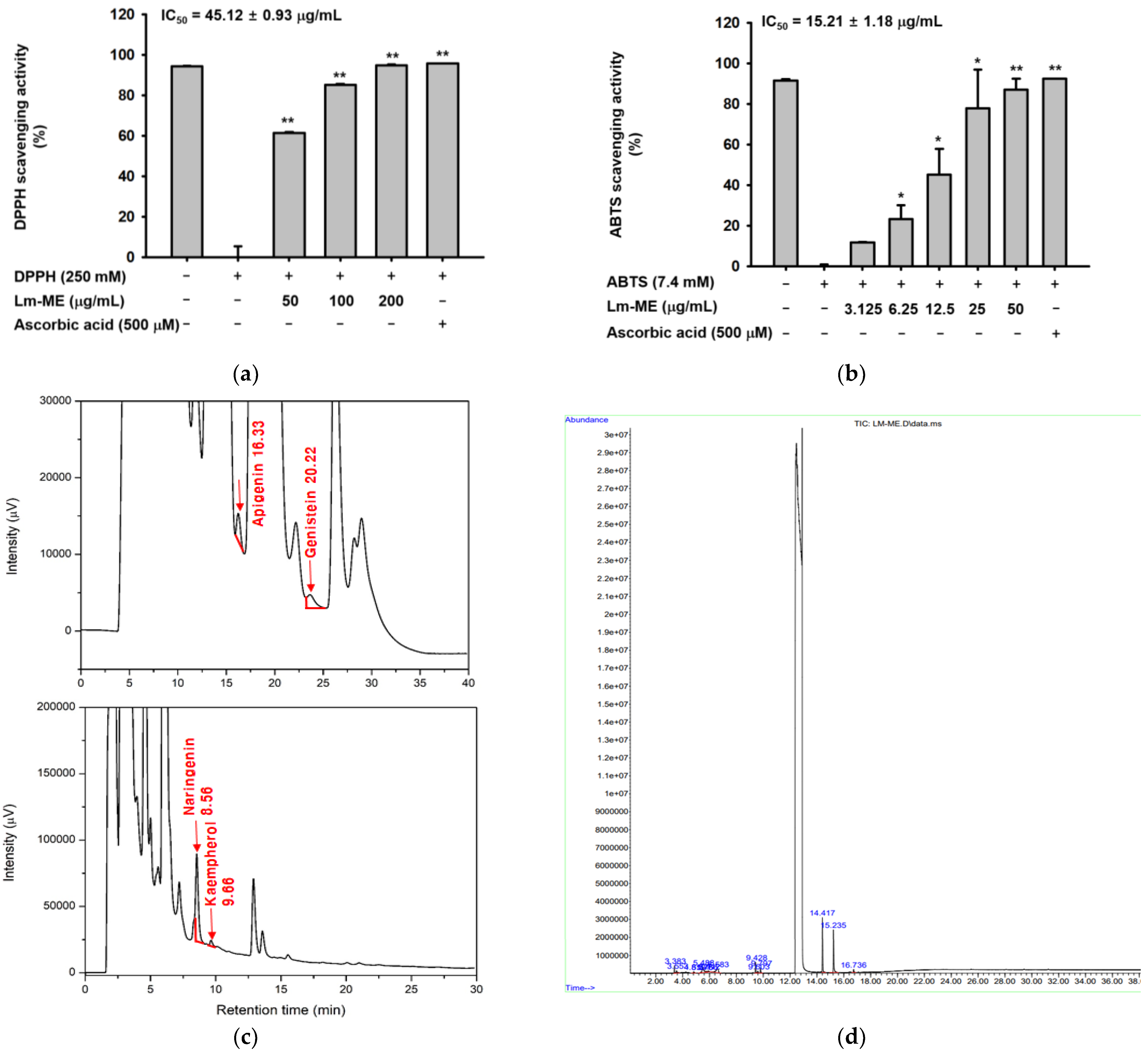
2.1. Antioxidatve Activity of Lm-ME and Its Phytochemical Fingerprinting Profiles
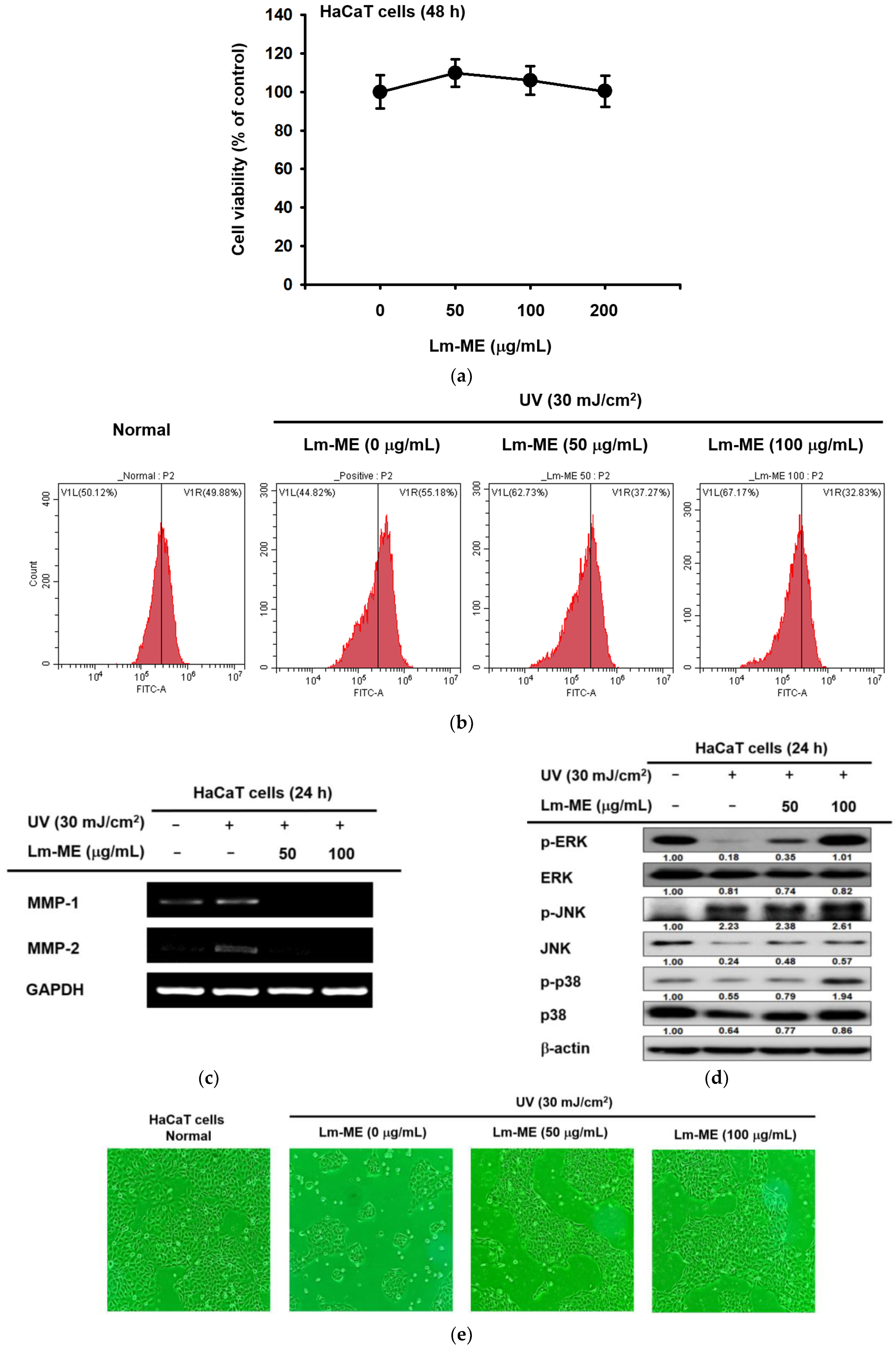
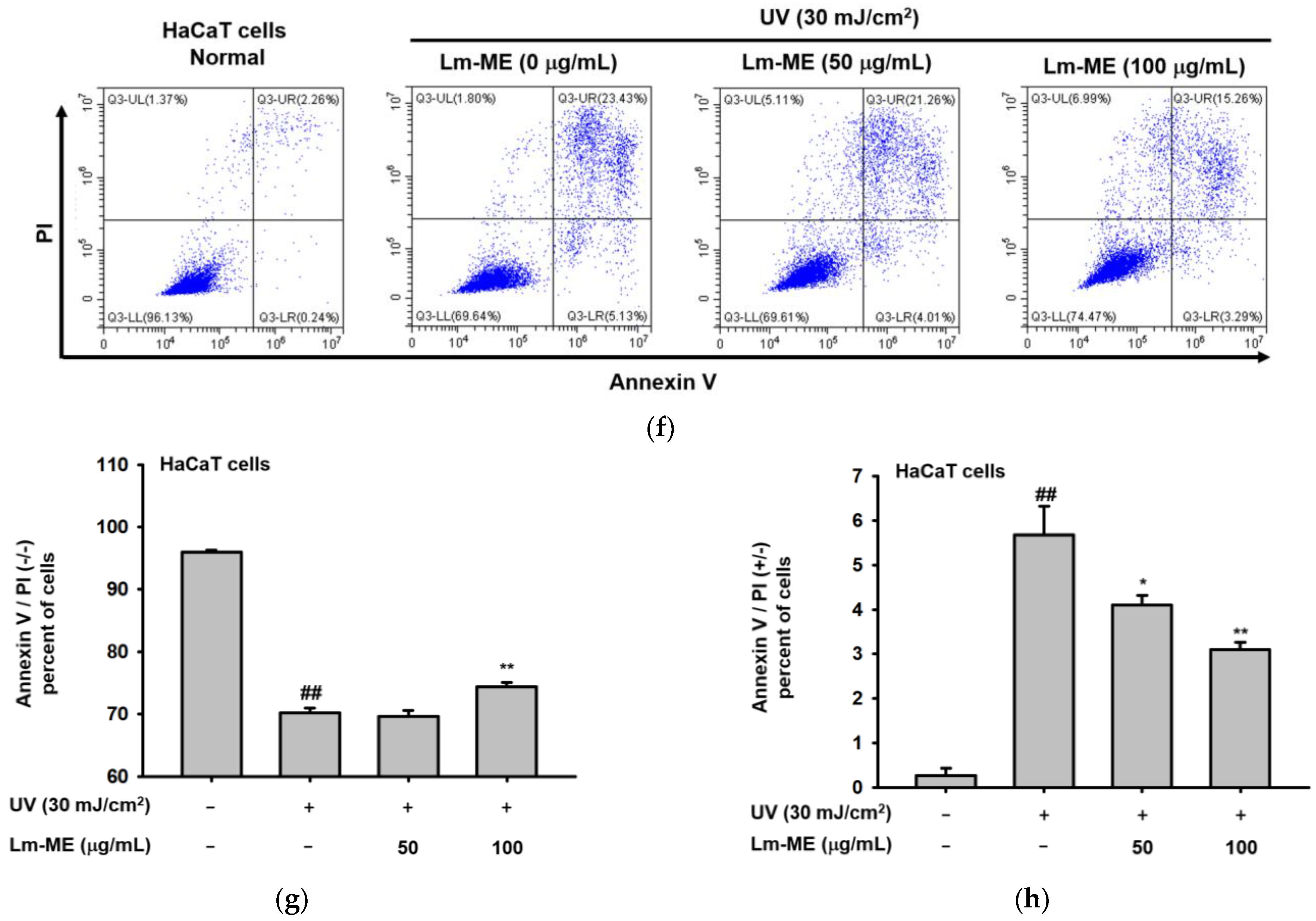
2.2. Lm-ME Showed the Role of UVB Protection on Human Keratinocytes
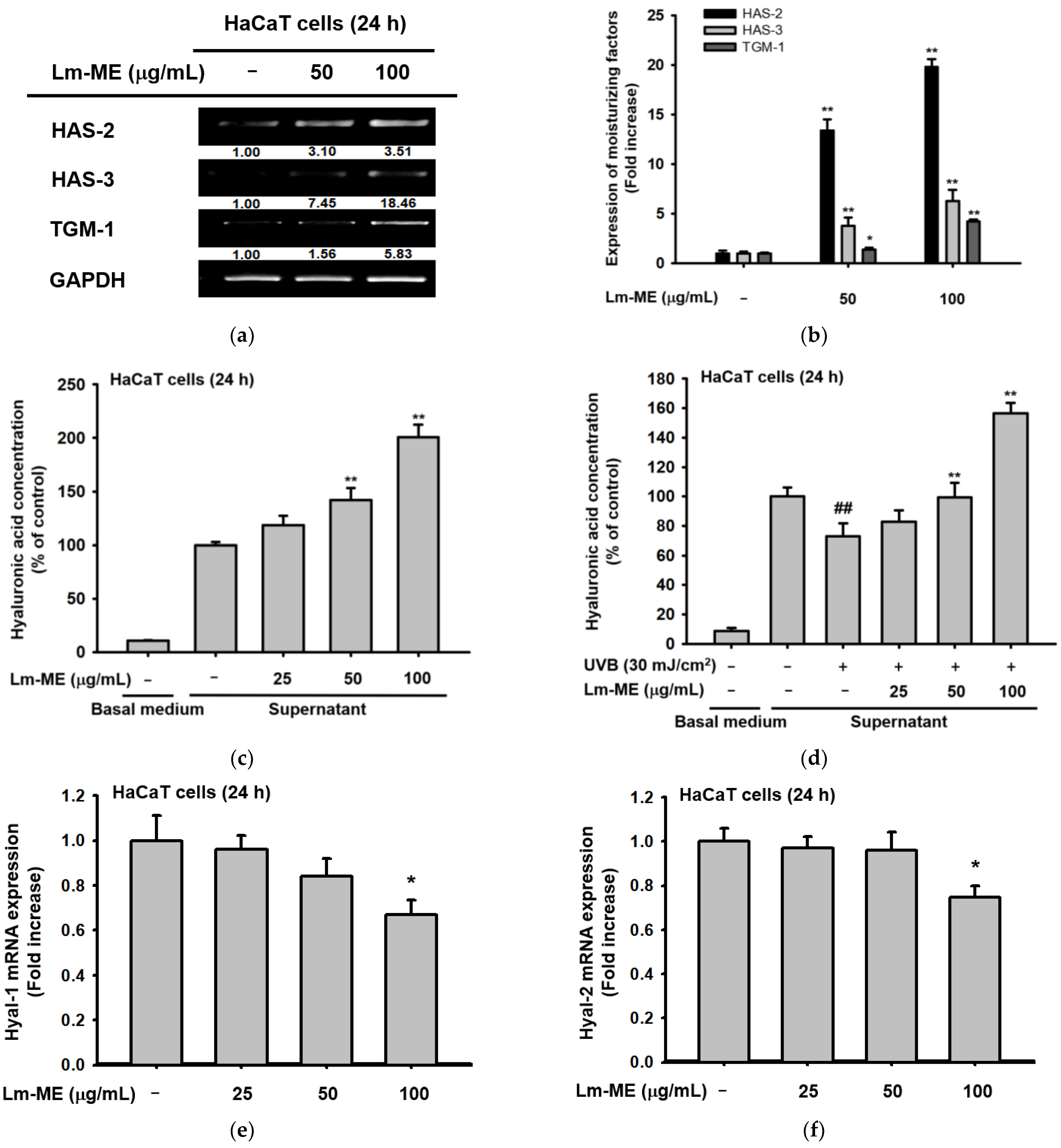
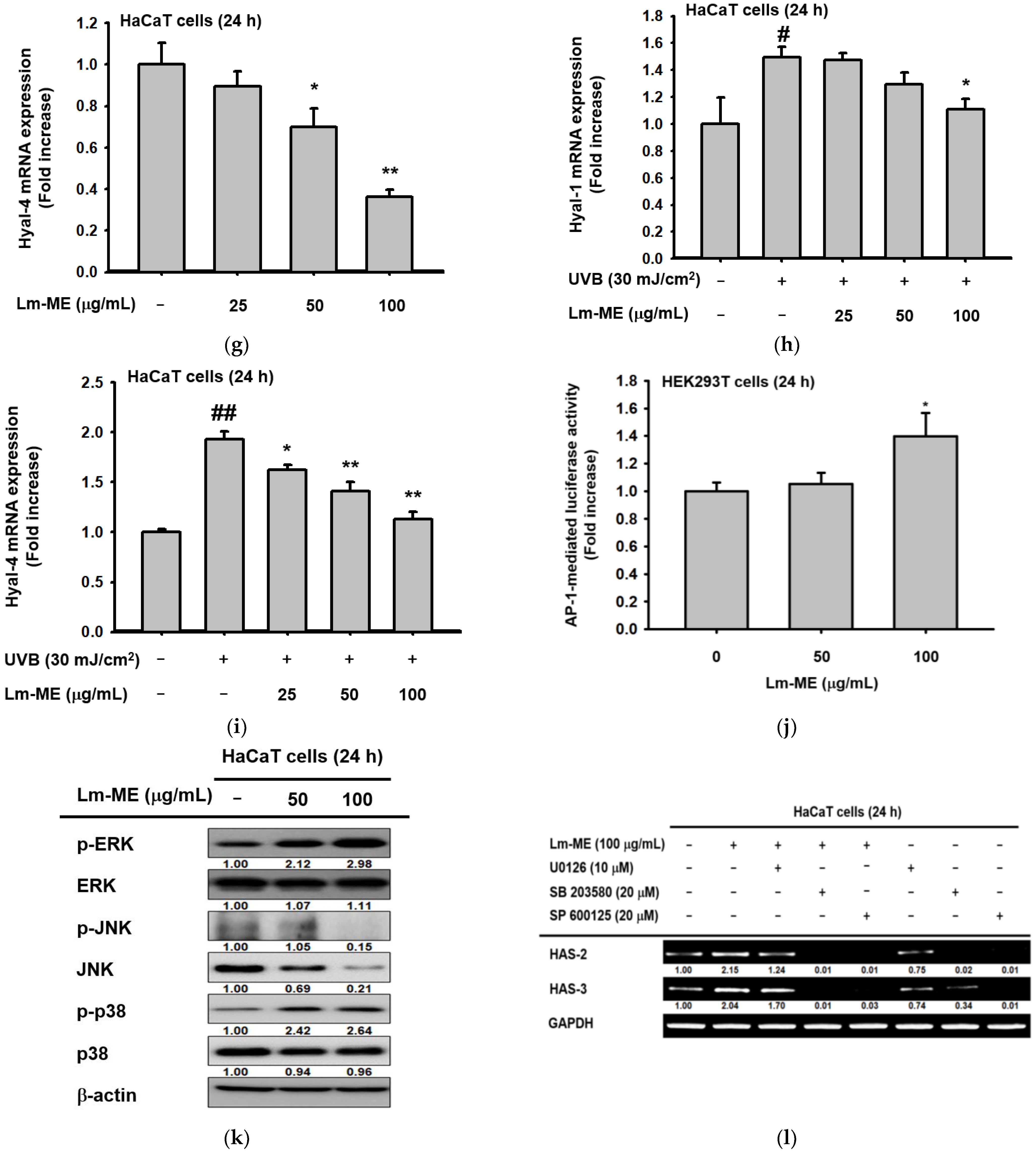
2.3. The Effects of Lm-ME on Hydration/Moisturization of HaCaT Cells
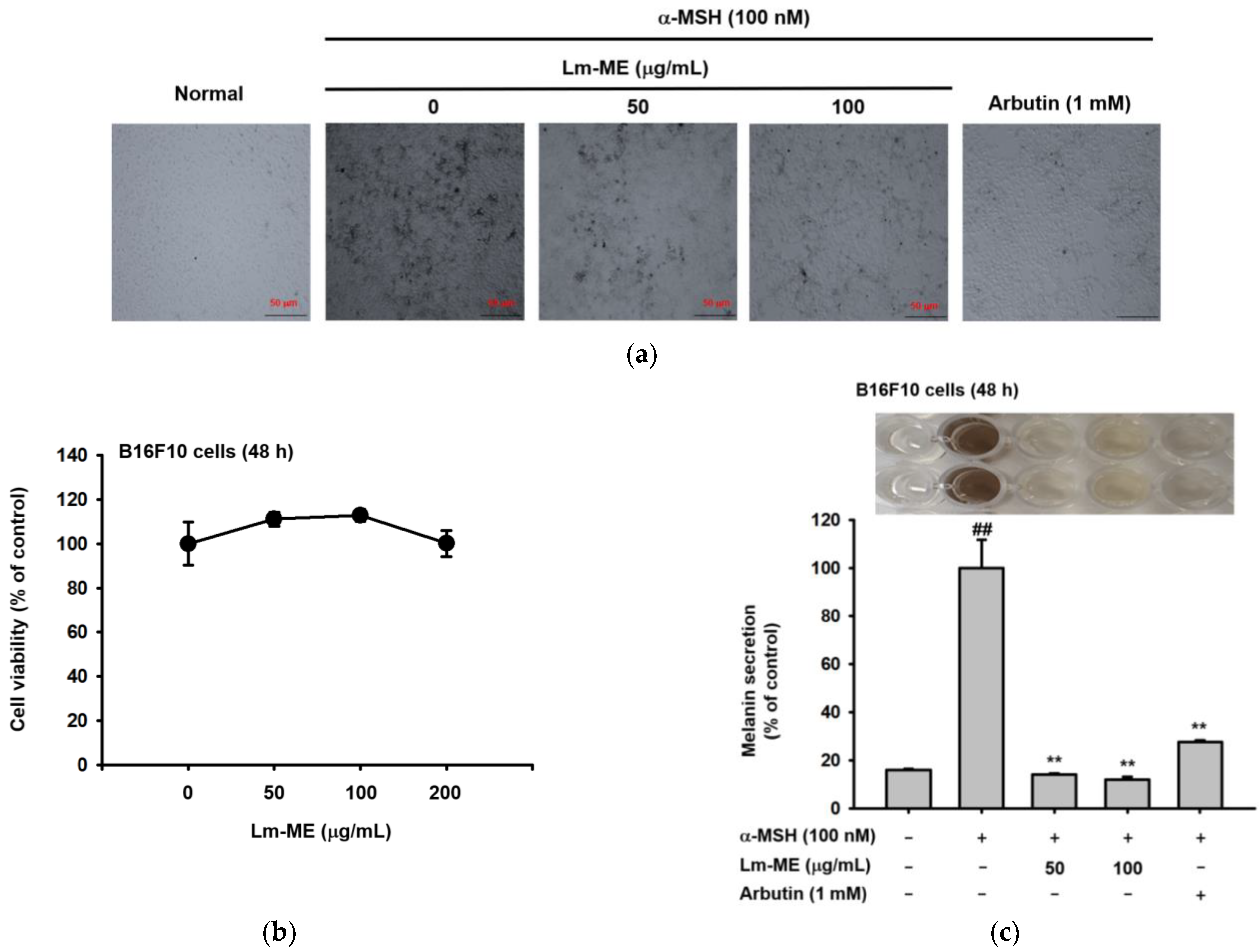
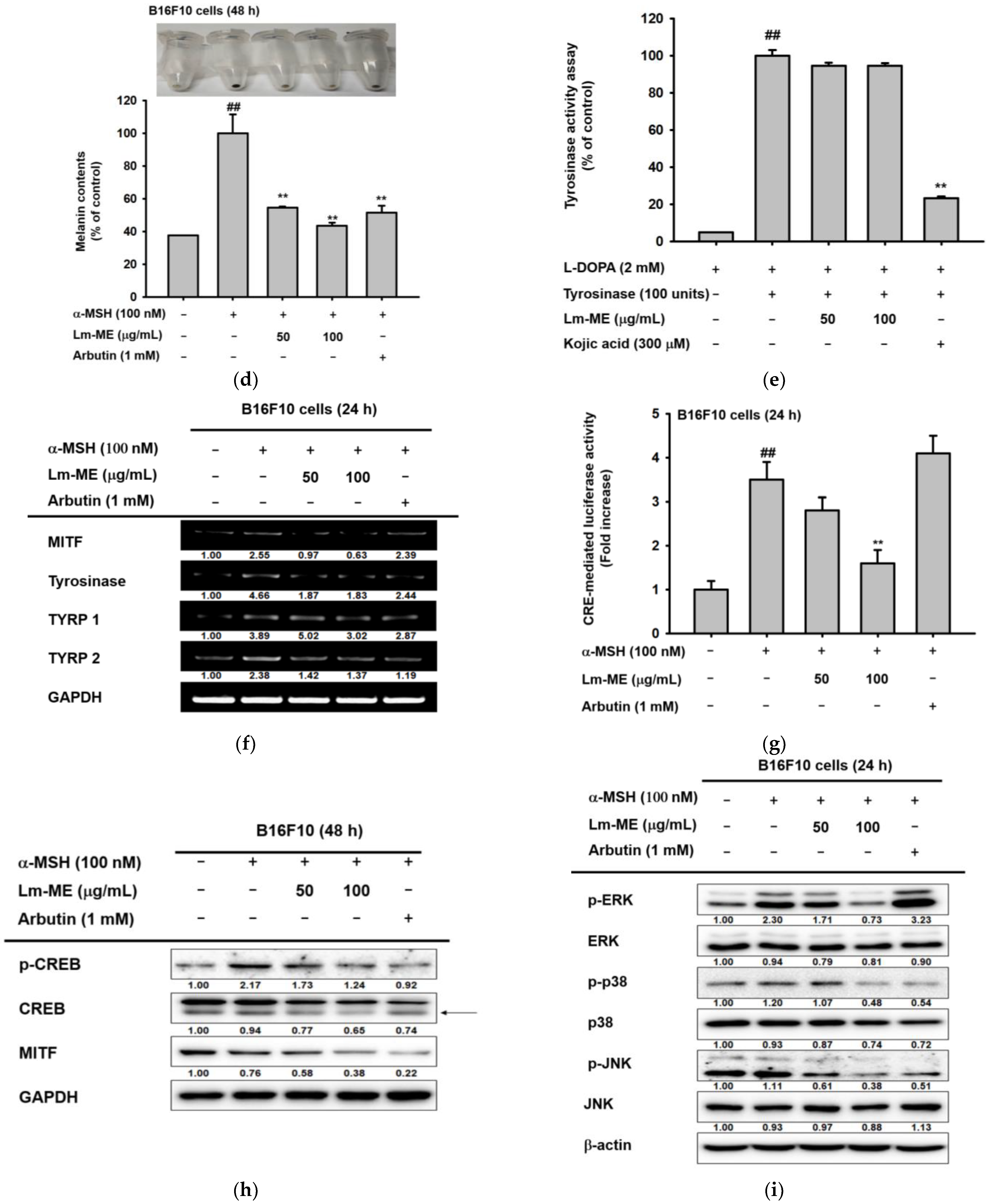
2.4. Lm-ME Suppressed Pigmentation Induced by α-MSH in B16F10 Cells
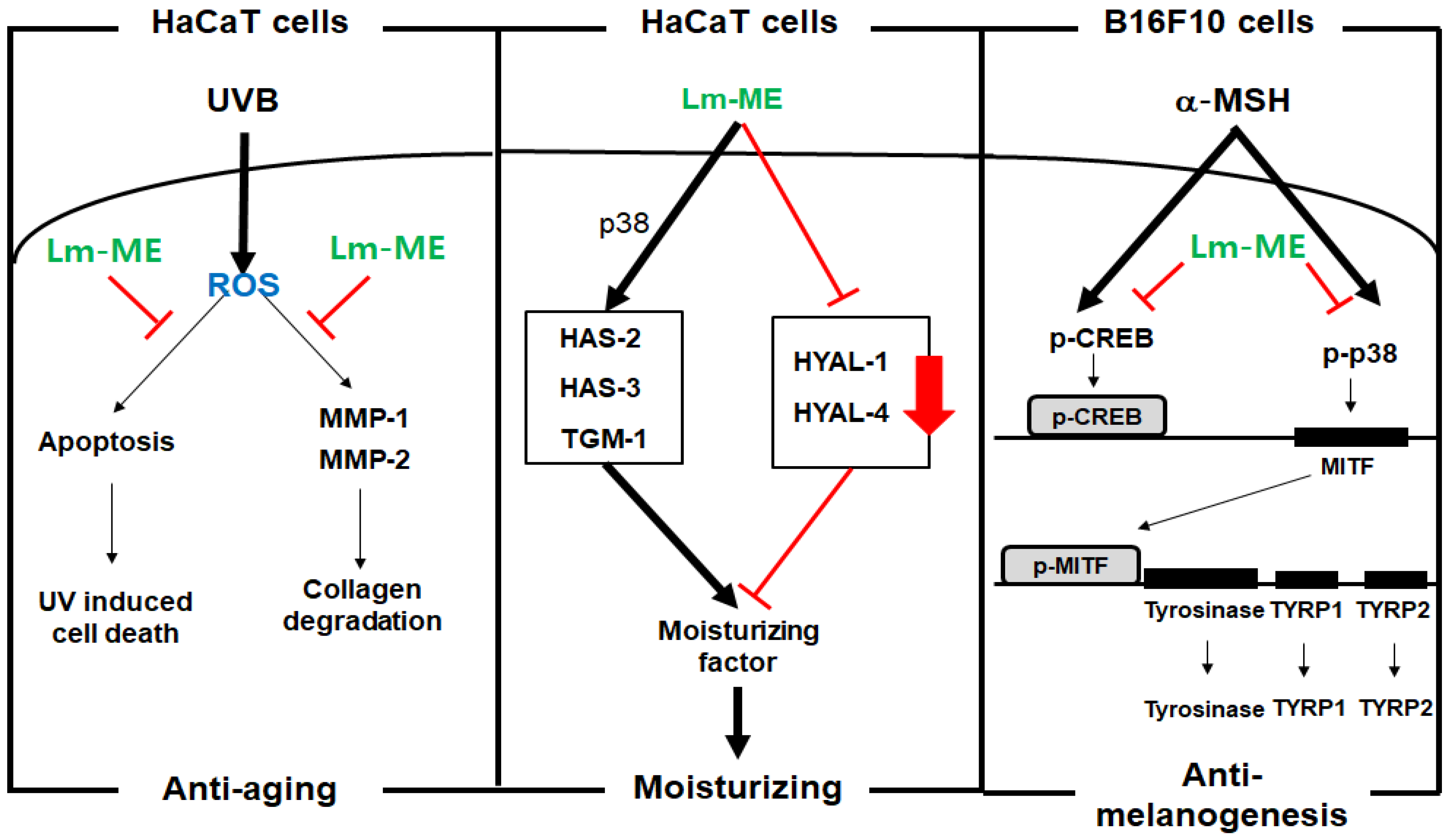
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Compound Treatment
4.4. High-Performance Liquid Chromatography and Gas Chromatography–Mass Spectrometry Analyses
4.5. MTT Assay
4.6. UVB Irradiation
4.7. UVB-Induced Apoptosis Analysis
4.8. Cellular ROS Analysis
4.9. DPPH Assay
4.10. ABTS Assay
4.11. Semi-Quantitative Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR) and Quantitative Real-Time PCR
4.12. Western Blot Analysis
4.13. Carbazole Assay for Hyaluronan Quantification
4.14. Melanogenesis and Melanin Secretion
4.15. Tyrosinase Assay
4.16. Luciferase Reporter Gene Assay
4.17. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Lm-ME | Licania macrocarpa Cuatrec methanol extract |
| CREB | cAMP response element (CRE) |
| DCDFA | 2′,7′-dichlorodihydrofluorescein diacetate |
| ERK | extracellular signal-regulated kinase |
| JNK | c-Jun N-terminal kinase |
| MAPKs | mitogen-activated protein kinases |
| MMP | matrix metalloproteinases |
| MITF | microphthalmia-associated transcription factor |
| α-MSH | α-melanocyte-stimulating hormone |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| ROS | reactive oxygen species |
| RT-PCR | reverse transcription-polymerase chain reaction |
| TYRP-1 | tyrosinase-related protein 1 |
| TYRP-2 | tyrosinase-related protein 2 |
| UV | ultraviolet |
| HPLC | high-performance liquid chromatography |
| HA | hyaluronic acid |
References
- You, L.; Cho, J.Y. The regulatory role of korean ginseng in skin cells. J. Ginseng Res. 2021, 45, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Lorz, L.R.; Lee, J.; Cho, J.Y. In vitro photoprotective, anti-inflammatory, moisturizing, and antimelanogenic effects of a methanolic extract of Chrysophyllum lucentifolium cronquist. Plants 2021, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.; Qomaladewi, N.P.; Lee, J.; Park, S.H.; Cho, J.Y. The role of autophagy in skin fibroblasts, keratinocytes, melanocytes, and epidermal stem cells. J. Investig. Dermatol. 2020, 140, 1691–1697. [Google Scholar] [CrossRef] [PubMed]
- Saba, E.; Kim, S.H.; Lee, Y.Y.; Park, C.K.; Oh, J.W.; Kim, T.H.; Kim, H.K.; Roh, S.S.; Rhee, M.H. Korean red ginseng extract ameliorates melanogenesis in humans and induces antiphotoaging effects in ultraviolet B-irradiated hairless mice. J. Ginseng Res. 2020, 44, 496–505. [Google Scholar] [CrossRef]
- Truong, V.L.; Keum, Y.S.; Jeong, W.S. Red ginseng oil promotes hair growth and protects skin against UVC radiation. J. Ginseng Res. 2021, 45, 498–509. [Google Scholar] [CrossRef]
- Ujfaludi, Z.; Tuzesi, A.; Majoros, H.; Rothler, B.; Pankotai, T.; Boros, I.M. Coordinated activation of a cluster of MMP genes in response to UVB radiation. Sci. Rep. 2018, 8, 2660. [Google Scholar] [CrossRef]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of matrix metalloproteinases in photoaging and photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef] [Green Version]
- Ahn, Y.; Kim, M.G.; Choi, Y.J.; Lee, S.J.; Suh, H.J.; Jo, K. Photoprotective effects of sphingomyelin-containing milk phospholipids in ultraviolet B–irradiated hairless mice by suppressing NF-κB expression. J. Dairy Sci. 2022, 105, 1929–1939. [Google Scholar] [CrossRef]
- Lorz, L.R.; Kim, M.Y.; Cho, J.Y. Medicinal potential of Panax ginseng and its ginsenosides in atopic dermatitis treatment. J. Ginseng Res. 2020, 44, 8–13. [Google Scholar] [CrossRef]
- Brozyna, A.; Zbytek, B.; Granese, J.; Carlson, J.A.; Ross, J.; Slominski, A. Mechanism of UV-related carcinogenesis and its contribution to nevi/melanoma. Expert. Rev. Dermatol. 2007, 2, 451–469. [Google Scholar] [CrossRef] [Green Version]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. Dermatoendocrinology 2012, 4, 253–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Videira, I.F.; Moura, D.F.; Magina, S. Mechanisms regulating melanogenesis. An. Bras. Dermatol. 2013, 88, 76–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Mello, S.A.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling pathways in melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, Y.; Hearing, V.J. Melanocytes and their diseases. Cold Spring Harb. Perspect. Med. 2014, 4, a017046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, N.; Vicanova, J.; Pavel, S. The hunt for natural skin whitening agents. Int. J. Mol. Sci. 2009, 10, 5326–5349. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.K.; Park, J.G.; Hong, Y.H.; Aziz, N.; Park, S.H.; Kim, S.; Kim, E.; Cho, J.Y. Anti-inflammatory effects of Licania macrocarpa cuatrec methanol extract target Src- and TAK1- mediated pathways. Evid. Based Complement. Alternat. Med. 2019, 2019, 4873870. [Google Scholar] [CrossRef] [Green Version]
- Feitosa, E.A.; Xavier, H.S.; Randau, K.P. Chrysobalanaceae: Traditional uses, phytochemistry and pharmacology. Rev. Bras. Farmacogn. 2012, 22, 1181–1186. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Shin, K.K.; Kim, H.G.; Jo, M.; Kim, J.K.; Lee, J.S.; Choung, E.S.; Li, W.Y.; Lee, S.W.; Kim, K.H.; et al. Src/NF-κB-targeted anti-inflammatory effects of Potentilla glabra var. Mandshurica (maxim.) hand.-mazz. ethanol extract. Biomolecules 2020, 10, 648. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Terazawa, S.; Mori, S.; Nakajima, H.; Yasuda, M.; Imokawa, G. The UVB-stimulated expression of transglutaminase 1 is mediated predominantly via the NF-κB signaling pathway: New evidence of its significant attenuation through the specific interruption of the p38/MSK1/NFκBp65 Ser276 Axis. PLoS ONE 2015, 10, e0136311. [Google Scholar] [CrossRef] [Green Version]
- Rauhala, L.; Jokela, T.; Karna, R.; Bart, G.; Takabe, P.; Oikari, S.; Tammi, M.I.; Pasonen-Seppanen, S.; Tammi, R.H. Extracellular ATP activates hyaluronan synthase 2 (HAS2) in epidermal keratinocytes via P2Y2, Ca2+ signaling, and MAPK pathways. Biochem. J. 2018, 475, 1755–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-O.; Kim, E.; Kim, J.H.; Hong, Y.H.; Kim, H.G.; Jeong, D.; Kim, J.; Kim, S.H.; Park, C.; Seo, D.B. Antimelanogenesis and skin-protective activities of panax ginseng calyx ethanol extract. J. Ginseng Res. 2018, 42, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Amaro-Ortiz, A.; Yan, B.; D’Orazio, J.A. Ultraviolet radiation, aging and the skin: Prevention of damage by topical cAMP manipulation. Molecules 2014, 19, 6202–6219. [Google Scholar] [CrossRef] [PubMed]
- Draelos, Z.D. Facial skin care products and cosmetics. Clin. Dermatol. 2014, 32, 809–812. [Google Scholar] [CrossRef] [PubMed]
- Kanakis, C.; Tarantilis, P.; Polissiou, M.; Diamantoglou, S.; Tajmir-Riahi, H. DNA interaction with naturally occurring antioxidant flavonoids quercetin, kaempferol, and delphinidin. J. Biomol. Struct. Dyn. 2005, 22, 719–724. [Google Scholar] [CrossRef]
- Vellosa, J.C.R.; Regasini, L.O.; Khalil, N.M.; Bolzani, V.d.S.; Khalil, O.A.; Manente, F.A.; Pasquini Netto, H.; Oliveira, O.M. Antioxidant and cytotoxic studies for kaempferol, quercetin and isoquercitrin. Eclética Quimica 2011, 36, 7–20. [Google Scholar] [CrossRef]
- Wang, H.; Kochevar, I.E. Involvement of UVB-induced reactive oxygen species in TGF-β biosynthesis and activation in keratinocytes. Free Radic Biol. Med. 2005, 38, 890–897. [Google Scholar] [CrossRef]
- Fagot, D.; Asselineau, D.; Bernerd, F. Direct role of human dermal fibroblasts and indirect participation of epidermal keratinocytes in MMP-1 production after UV-B irradiation. Arch. Dermatol. Res. 2002, 293, 576–583. [Google Scholar] [CrossRef]
- Lim, T.-G.; Jeon, A.J.; Yoon, J.H.; Song, D.; Kim, J.-E.; Kwon, J.Y.; Kim, J.R.; Kang, N.J.; Park, J.-S.; Yeom, M.H. 20-O-β-D-glucopyranosyl-20 (s)-protopanaxadiol, a metabolite of ginsenoside rb1, enhances the production of hyaluronic acid through the activation of ERK and AKT mediated by Src tyrosin kinase in human keratinocytes. Int. J. Mol. Med. 2015, 35, 1388–1394. [Google Scholar] [CrossRef] [Green Version]
- Siiskonen, H.; Oikari, S.; Pasonen-Seppänen, S.; Rilla, K. Hyaluronan synthase 1: A mysterious enzyme with unexpected functions. Front. Immunol. 2015, 6, 43. [Google Scholar] [CrossRef] [Green Version]
- Jing, H.; Sun, W.; Fan, J.; Zhang, Y.; Yang, J.; Jia, J.; Li, J.; Guo, J.; Luo, S.; Zheng, Y. Shikonin induces apoptosis of HaCaT cells via the mitochondrial, ERK and AKT pathways. Mol. Med. Rep. 2016, 13, 3009–3016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, R.; Zhao, X.; Yu, X.; Sun, Q. Metformin promotes HaCaT cell apoptosis through generation of reactive oxygen species via Raf-1-ERK1/2-NRF2 inactivation. Inflammation 2018, 41, 948–958. [Google Scholar] [CrossRef] [PubMed]
- Yardman-Frank, J.M.; Fisher, D.E. Skin pigmentation and its control: From ultraviolet radiation to stem cells. Exp. Dermatol. 2020, 30, 560–571. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Weng, Q.Y.; Fisher, D.E. UV signaling pathways within the skin. J. Investig. Dermatol. 2014, 134, 2080–2085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, D.; Park, S.H.; Kim, M.H.; Lee, S.; Cho, Y.K.; Kim, Y.A.; Park, B.J.; Lee, J.; Kang, H.; Cho, J.Y. Anti-melanogenic effects of ethanol extracts of the leaves and roots of Patrinia villosa (thunb.) Juss through their inhibition of CREB and induction of ERK and autophagy. Molecules 2020, 25, 5375. [Google Scholar] [CrossRef]
- Qomaladewi, N.P.; Kim, M.Y.; Cho, J.Y. Rottlerin reduces cAMP/CREB-mediated melanogenesis via regulation of autophagy. Int. J. Mol. Sci. 2019, 20, 2081. [Google Scholar] [CrossRef] [Green Version]
- Sangkaew, O.; Yompakdee, C. Fermented unpolished black rice (Oryza sativa L.) inhibits melanogenesis via ERK, p38, and AKT phosphorylation in B16F10 melanoma cells. J. Microbiol. Biotechnol. 2020, 30, 1184–1194. [Google Scholar] [CrossRef]
- Chung, Y.C.; Hyun, C.G. Inhibitory effects of pinostilbene hydrate on melanogenesis in B16F10 melanoma cells via ERK and p38 signaling pathways. Int. J. Mol. Sci. 2020, 21, 4732. [Google Scholar] [CrossRef]
- Xu, Z.; Chen, L.; Jiang, M.; Wang, Q.; Zhang, C.; Xiang, L.F. CCN1/Cyr61 stimulates melanogenesis through integrin α6β1, p38 MAPK, and ERK1/2 signaling pathways in human epidermal melanocytes. J. Investig. Dermatol. 2018, 138, 1825–1833. [Google Scholar] [CrossRef] [Green Version]
- Qomaladewi, N.; Aziz, N.; Mi Yeon, K.; Cho, J.Y. Piper cubeba L. methanol extract has anti-inflammatory activity targeting Src/Syk via NF- κB inhibition. Evid. Based Complement. Alternat. Med. 2019, 2019, 1548125. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.P.; Kim, D.S.; Park, S.H.; Shin, C.Y.; Woo, J.J.; Kim, J.W.; An, R.B.; Lee, C.; Cho, J.Y. Antioxidant capacity of Potentilla paradoxa nutt. and its beneficial effects related to anti-aging in hacat and B16F10 cells. Plants 2022, 11, 873. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.O.; Hwang, S.H.; Shen, T.; Kim, J.H.; You, L.; Hu, W.; Cho, J.Y. Enhancement of skin barrier and hydration-related molecules by protopanaxatriol in human keratinocytes. J. Ginseng Res. 2021, 45, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.O.; Kim, J.H.; Kim, S.; Kim, M.Y.; Hong, Y.H.; Kim, H.G.; Cho, J.Y. Gastroprotective effects of the nonsaponin fraction of Korean red ginseng through cyclooxygenase-1 upregulation. J. Ginseng Res. 2020, 44, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Park, H.R.; Cha, S.Y.; Lee, S.H.; Jo, J.W.; Go, J.N.; Lee, K.H.; Lee, S.Y.; Shin, S.S. Effect of red ginseng naturalgel on skin aging. J. Ginseng Res. 2020, 44, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Eruslanov, E.; Kusmartsev, S. Identification of ROS using oxidized DCFDA and flow-cytometry. Methods Mol. Biol. 2010, 594, 57–72. [Google Scholar]
- Molyneux, P. The use of the stable free radical diphenylpicrylhydrazyl (DPPH) for estimating antioxidant activity. Songklanakarin J. Sci. Technol. 2004, 26, 211–219. [Google Scholar]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Byrne, D.H. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compost. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Han, S.Y.; Kim, J.; Kim, E.; Kim, S.H.; Seo, D.B.; Kim, J.H.; Shin, S.S.; Cho, J.Y. AKT-targeted anti-inflammatory activity of Panax ginseng calyx ethanolic extract. J. Ginseng Res. 2018, 42, 496–503. [Google Scholar] [CrossRef]
- Lee, Y.G.; Byeon, S.E.; Kim, J.Y.; Lee, J.Y.; Rhee, M.H.; Hong, S.; Wu, J.C.; Lee, H.S.; Kim, M.J.; Cho, D.H.; et al. Immunomodulatory effect of Hibiscus cannabinus extract on macrophage functions. J. Ethnopharmacol. 2007, 113, 62–71. [Google Scholar] [CrossRef]
- You, L.; Kim, M.-Y.; Cho, J.Y. Protective effect of Potentilla glabra in UVB-induced photoaging process. Molecules 2021, 26, 5408. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.Y.; Kim, E.; Park, S.H.; Hwang, K.H.; Kim, D.; Jung, Y.J.; Kopalli, S.R.; Hong, Y.D.; Sung, G.H.; Cho, J.Y. Antimelanogenesis effects of theasinensin a. Int. J. Mol. Sci. 2021, 22, 7453. [Google Scholar] [CrossRef] [PubMed]
- Lorz, L.R.; Yoo, B.C.; Kim, M.-Y.; Cho, J.Y. Anti-wrinkling and anti-melanogenic effect of Pradosia mutisii methanol extract. Int. J. Mol. Sci. 2019, 20, 1043. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.A.; Lee, C.Y.; Mitra, A.; Kim, H.; Woo, B.Y.; Hong, Y.D.; Noh, J.K.; Yi, D.K.; Kim, H.G.; Cho, J.Y. Anti-inflammatory effects of Huberia peruviana cogn. methanol extract by inhibiting Src activity in the NF-κB pathway. Plants 2021, 10, 2335. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | RT | Name of the Compound | Corrected Area | % of Total |
|---|---|---|---|---|
| 1 | 3.383 | Piperidine | 6,769,119 | 2.77% |
| 2 | 3.553 | Carbonic acid gas | 6,631,886 | 2.71% |
| 3 | 4.819 | Ethanol | 2,818,376 | 1.15% |
| 4 | 5.376 | Dimethyl sulfide | 625,946 | 0.26% |
| 5 | 5.488 | 2-Thiapropane | 25,021,273 | 10.22% |
| 6 | 5.756 | Methylthiomethane | 2,364,524 | 0.97% |
| 7 | 5.912 | Methyl sulfide | 3,966,607 | 1.62% |
| 8 | 6.583 | Dimethyl monosulfide | 27,990,880 | 11.44% |
| 9 | 9.428 | 2,3-Dithiabutane | 14,268,284 | 5.83% |
| 10 | 9.603 | Chloromethylmethyl sulfide | 5,181,894 | 2.12% |
| 11 | 9.797 | Methanesulphinic acid methyl ester | 7,917,119 | 3.23% |
| 12 | 14.417 | Dimethyl sulfone | 71,638,885 | 29.27% |
| 13 | 15.235 | Methyl-d3-hydrazine sulfate | 62,574,639 | 26.67% |
| 14 | 16.736 | S-Methyl methanethiosulphonate | 4,303,891 | 1.76% |
| Instrument | Condition A | Condition B | ||||
|---|---|---|---|---|---|---|
| Column | CAPCELL PAK C18 MG, 4.6 mm I.D. × 250 mm | |||||
| Detector | UV-Vis Detector | |||||
| Wavelength | 254 nm | 350 nm | ||||
| Compounds for analysis | Naringenin, kaempferol, and genistein | apigenin, hesperidin, and rutin | ||||
| Analyzed period | 30 min | 40 min | ||||
| Solvent | Solvent A | 2% acetic acid in water | Solvent A | 0.1% formic acid in MeOH:water = 10:90 | ||
| Solvent B | 0.5% acetic acid in water: ACN = 50:50 | Solvent B | 0.1% formic acid in MeOH:water = 90:10 | |||
| Flow rate | 1 mL/min | 0.4 mL/min | ||||
| Volume | 10 μL | 10 μL | ||||
| Gradient | Time (min) | Composition (%) | Time (min) | Composition (%) | ||
| A | B | A | B | |||
| 0 | 28 | 72 | 0 | 40 | 60 | |
| 20 | 0 | 100 | 20 | 40 | 60 | |
| 30 | 0 | 100 | 25 | 70 | 30 | |
| - | - | - | 40 | 70 | 30 | |
| PCR Type | Gene Name | Sequence (5′-3′) | |
|---|---|---|---|
| RT-PCR (human) | MMP-1 | F | TCTGACGTTGATCCCAGAGAGCAG |
| R | CAGGGTGACACCAGTGACTGCAC | ||
| MMP-2 | F | AAAACGGACAAAGAGTTGGCA | |
| R | CTGGGGCAGTCCAAAGAACT | ||
| HAS-2 | F | CCACCCAGTACAGCGTCAAC | |
| R | CATGGTGCTTCTGTCGCTCT | ||
| HAS-3 | F | TTCTTTATGTGACTCATCTGTCTCACCG | |
| R | ATTGTTGGCTACCAGTTTATCCAAACG | ||
| TGM-1 | F | AGGGAAGATCCAAGAGCCCA | |
| R | ACTCTGGATCCCCTACGCTT | ||
| HYAL-1 | F | TGT GGA CGT GGA TGT CAG TG | |
| R | GTA GTA GGG GTA GGT GCC CA | ||
| HYAL-2 | F | ATG TGC AGA ACT GGG AGA GC | |
| R | GGA AGC AAG TGT CTC GTC CA | ||
| HYAL-3 | F | TCT GGG CAT CAT AGC CAA CC | |
| R | AGA GGC CGA GTT GGT TCT TG | ||
| HYAL-4 | F | TCC TGT GAT TGG AAG CCC AC | |
| R | TAA TGG GGA CCC CCT GTG AT | ||
| GAPDH | F | GGTCACCAGGGCTGCTTTTA | |
| R | CACCGAGGAACTACCTGAT | ||
| Real-time PCR (human) | HYAL-1 | F | TGTGGACGTGGATGTCAGTG |
| R | GTAGTAGGGGTAGGTGCCCA | ||
| HYAL-2 | F | ATGTGCAGAACTGGGAGAGC | |
| R | GGAAGCAAGTGTCTCGTCCA | ||
| HYAL-3 | F | TCTGGGCATCATAGCCAACC | |
| R | AGAGGCCGAGTTGGTTCTTG | ||
| HYAL-4 | F | TCCTGTGATTGGAAGCCCAC | |
| R | TAATGGGGACCCCCTGTGAT | ||
| HAS-2 | F | TGACAGGCATCTCACGAACC | |
| R | TGGCGGGAAGTAAACTCGAC | ||
| HAS-3 | F | TATACCGCGCGCTCCAA | |
| R | GCCACTCCCGGAAGTAAGACT | ||
| TGM-1 | F | GAAATGCGGCAGATGACGAC | |
| R | AACTCCCCAGCGTCTGATTG | ||
| GAPDH | F | GAC AGT CAG CCG CAT CTT CT | |
| R | GCG CCC AAT ACG ACC AAA TC | ||
| RT-PCR (mouse) | MITF | F | TCTGACGTTGATCCCAGAGAGCAG |
| R | CAGGGTGACACCAGTGACTGCAC | ||
| Tyrosinase | F | AAAACGGACAAAGAGTTGGCA | |
| R | CTGGGGCAGTCCAAAGAACT | ||
| TYRP-1 | F | ATGGAACGGGAGGACAAACC | |
| R | TCCTGACCTGGCCATTGAAC | ||
| TYRP-2 | F | CAGTTTCCCCGAGTCTGCAT | |
| R | GTCTAAGGCGCCCAAGAACT | ||
| GAPDH | F | CACTCACGGCAAATTCAACGGCAC | |
| R | GACTCCACGACATACTCAGCAC | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, K.K.; Park, S.H.; Lim, H.Y.; Lorza, L.R.; Qomaladewia, N.P.; You, L.; Aziz, N.; Kim, S.A.; Lee, J.S.; Choung, E.S.; et al. In Vitro Anti-Photoaging and Skin Protective Effects of Licania macrocarpa Cuatrec Methanol Extract. Plants 2022, 11, 1383. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101383
Shin KK, Park SH, Lim HY, Lorza LR, Qomaladewia NP, You L, Aziz N, Kim SA, Lee JS, Choung ES, et al. In Vitro Anti-Photoaging and Skin Protective Effects of Licania macrocarpa Cuatrec Methanol Extract. Plants. 2022; 11(10):1383. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101383
Chicago/Turabian StyleShin, Kon Kuk, Sang Hee Park, Hye Yeon Lim, Laura Rojas Lorza, Nurinanda Prisky Qomaladewia, Long You, Nur Aziz, Soo Ah Kim, Jong Sub Lee, Eui Su Choung, and et al. 2022. "In Vitro Anti-Photoaging and Skin Protective Effects of Licania macrocarpa Cuatrec Methanol Extract" Plants 11, no. 10: 1383. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101383