
Thaumatin-like Protein (TLP) Genes in Garlic (Allium sativum L.): Genome-Wide Identification, Characterization, and Expression in Response to Fusarium proliferatum Infection



Abstract
:1. Introduction
2. Results
2.1. In Silico Genome-Wide Identification of TLP Genes in A. sativum cv. Ershuizao
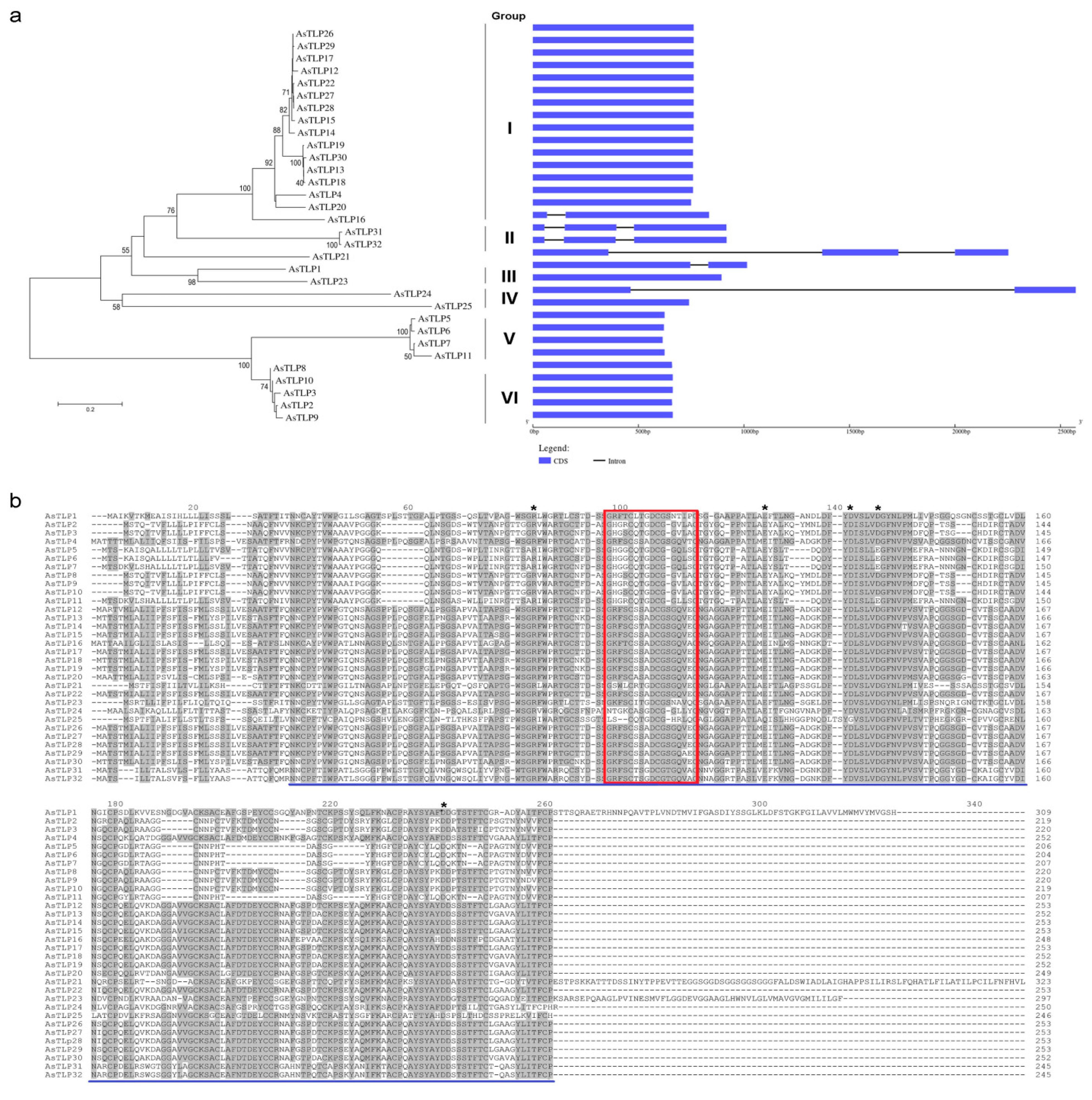
2.2. Structural and Phylogenetic Analyses of AsTLPs
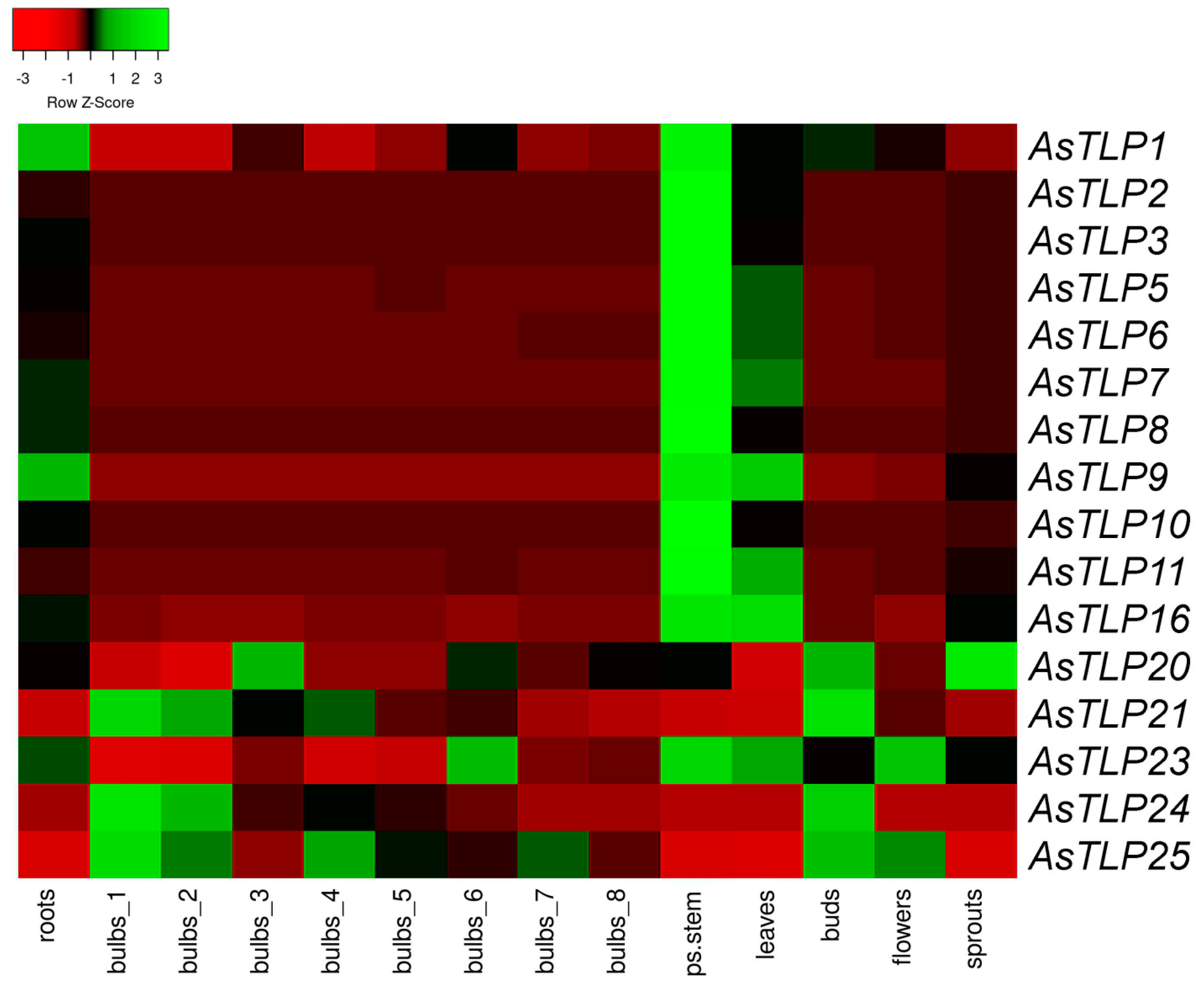
2.3. In Silico Analysis of AsTLP Transcription
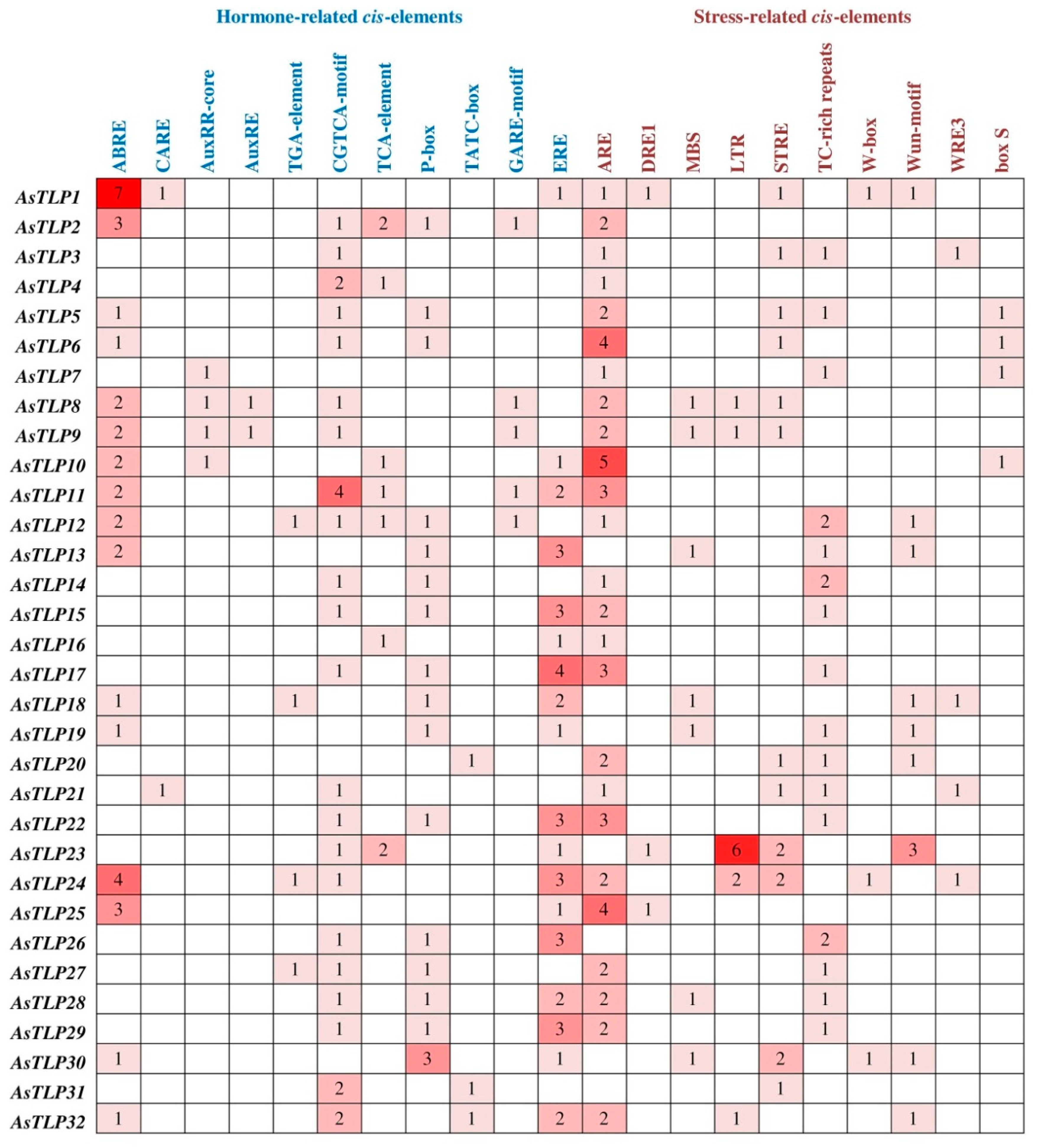
2.4. AsTLP Promoter Analysis
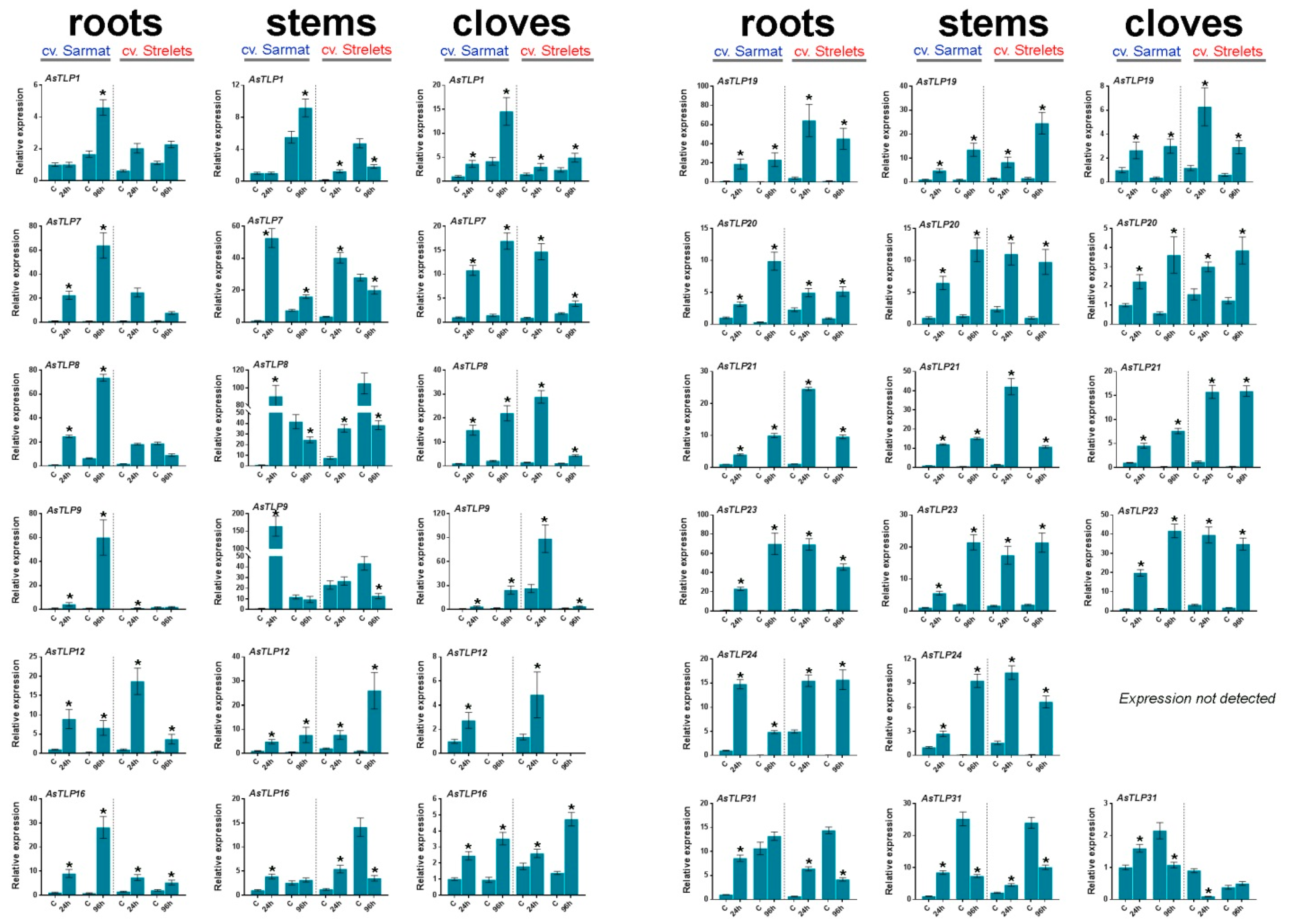
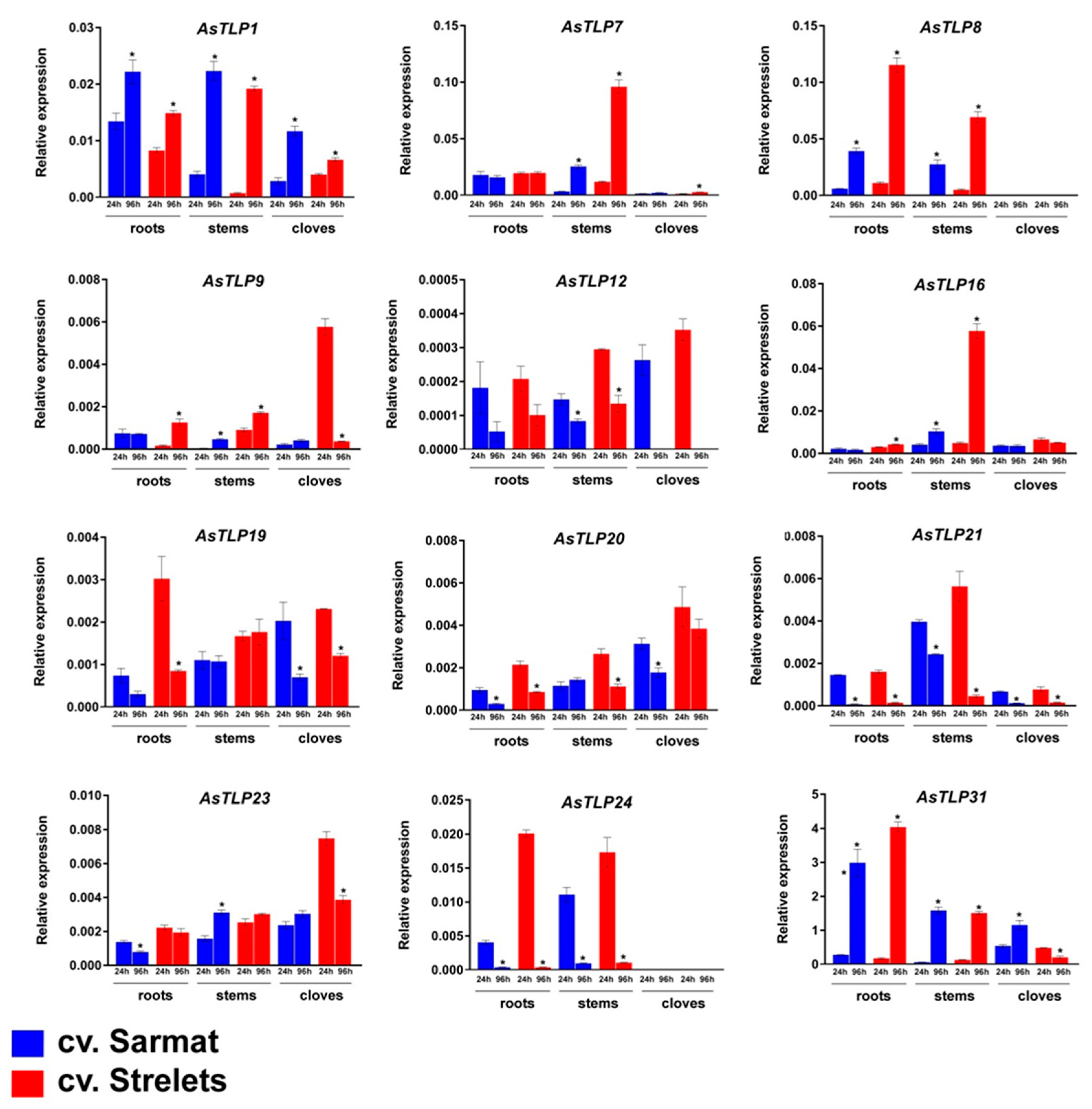
2.5. AsTLP1-32 Expression in cv. Sarmat and Strelets Infected with F. proliferatum
2.6. Cloning and Characterization of CDSs of AsTLP Genes Differentially Expressed in FBR-Sensitive and -Resistant Cultivars
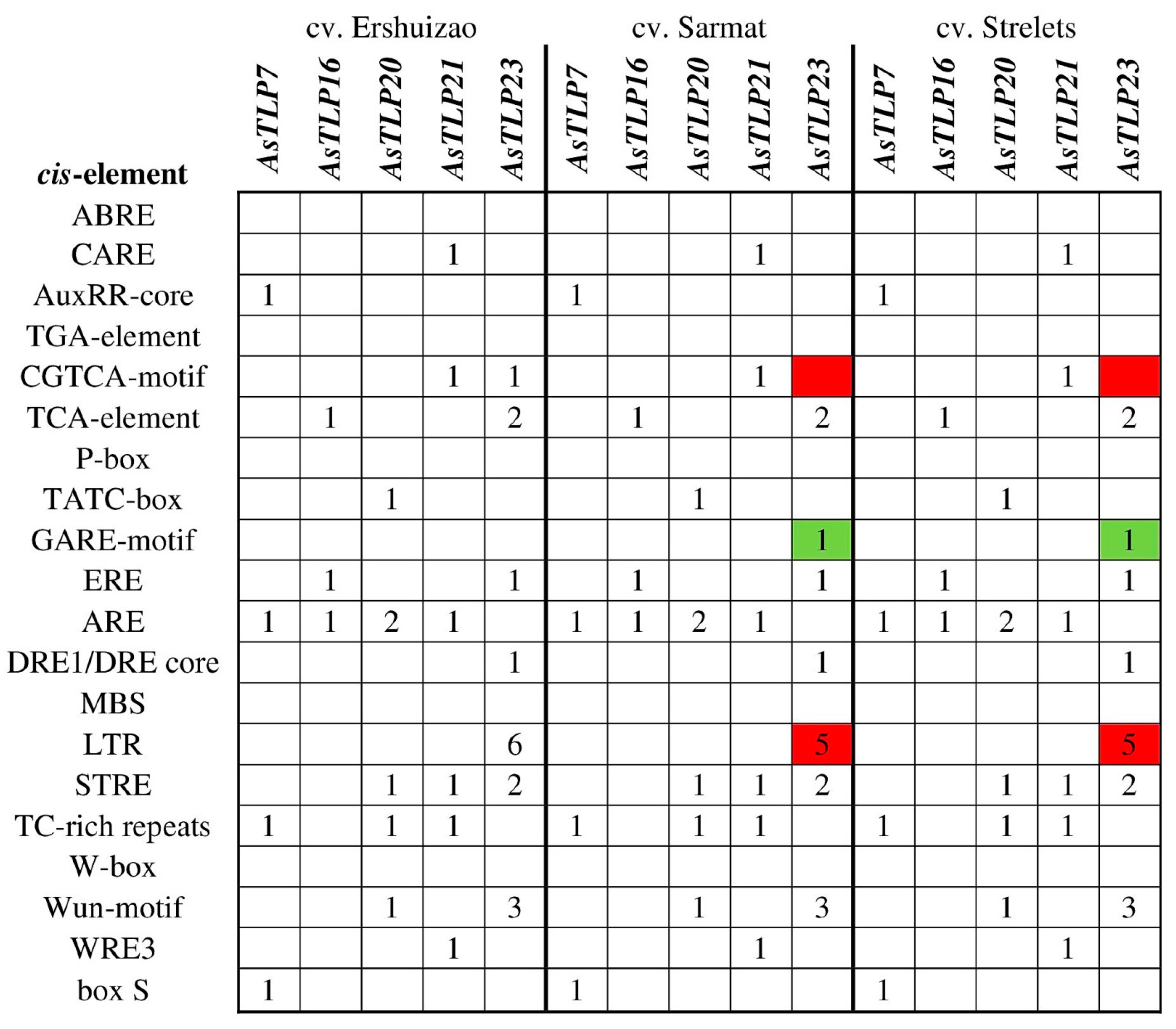
2.7. Analysis of Regulatory Regions in the AsTLP Genes Differentially Expressed in cv. Sarmat and Strelets after F. proliferatum Infection
3. Discussion
4. Materials and Methods
4.1. In Silico Identification and Structural Characterization of AsTLP Genes
4.2. In Silico mRNA Expression Analysis
4.3. Gene Identification
4.4. Plants, Fungi, and F. proliferatum Infection Assay
4.5. RNA Extraction and Quantitative Real-Time Reverse Transcription PCR (qRT-PCR)
4.6. Promoter and 5′-UTR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Selitrennikoff, C.P. Antifungal proteins. Appl. Environ. Microbiol. 2001, 67, 2883–2894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vander Wel, H.; Loeve, K. Isolation and characterization of thaumatin I & II, the sweet-tasting proteins from Thaumatoccus danielli. Eur. J. Biochem. 1972, 31, 221–225. [Google Scholar] [CrossRef]
- De Jesús-Pires, C.; Ferreira-Neto, J.R.C.; Pacifico Bezerra-Neto, J.; Kido, E.A.; de Oliveira Silva, R.L.; Pandolfi, V.; Wanderley-Nogueira, A.C.; Binneck, E.; da Costa, A.F.; Pio-Ribeiro, G.; et al. Plant Thaumatin-like Proteins: Function, Evolution and Biotechnological Applications. Curr. Protein Pept. Sci. 2020, 21, 36–51. [Google Scholar] [CrossRef] [PubMed]
- Vitali, A.; Pacini, L.; Bordi, E.; De Mori, P.; Pucillo, L.; Maras, B.; Botta, B.; Brancaccio, A.; Giardina, B. Purification and characterization of an antifungal thaumatin-like protein from Cassia didymobotrya cell culture. Plant Physiol. Biochem. 2006, 44, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Basu, A.; Kundu, S. Cloning, characterization, and bacterial over-expression of an osmotin-like protein gene from Solanum nigrum L. with antifungal activity against three necrotrophic fungi. Mol. Biotechnol. 2015, 57, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Saeidi, M.; Zareie, R. Prediction, isolation, overexpression and antifungal activity analysis of Medicago truncatula var. truncatula putative thaumatin like proteins (TLP-1, -2, -3, -4 and -5). Turk. J. Biol. 2020, 44, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, T.; Jan, A.; Kakishima, M.; Komatsu, S. Proteomic analysis of bacterial-blight defense-responsive proteins in rice leaf blades. Proteomics 2006, 6, 6053–6065. [Google Scholar] [CrossRef]
- Rodrigo, I.; Vera, P.; Frank, R.; Conejero, V. Identification of the viroid-induced tomato pathogenesis-related (PR) protein P23 as the thaumatin-like tomato protein NP24 associated with osmotic stress. Plant Mol. Biol. 1991, 16, 931–934. [Google Scholar] [CrossRef] [PubMed]
- Looze, Y.; Boussard, P.; Huet, J.; Vandenbusche, G.; Azarkan, M.; Raussens, V.; Wintjens, R. Purification and characterization of a wound-inducible thaumatin-like protein from the latex of Carica papaya. Phytochemistry 2009, 70, 970–978. [Google Scholar] [CrossRef]
- Park, E.J.; Kim, T.H. Thaumatin-like genes function in the control of both biotic stress signaling and ABA signaling pathways. Biochem. Biophys. Res. Commun. 2021, 567, 17–21. [Google Scholar] [CrossRef]
- Wang, X.; Tang, C.; Deng, L.; Cai, G.; Liu, X.; Liu, B.; Han, Q.; Buchenauer, H.; Wei, G.; Han, D.; et al. Characterization of a pathogenesis-related thaumatin-like protein gene TaPR5 from wheat induced by stripe rust fungus. Physiol. Plant. 2010, 139, 27–38. [Google Scholar] [CrossRef]
- He, L.; Li, L.; Zhu, Y.; Pan, Y.; Zhang, X.; Han, X.; Li, M.; Chen, C.; Li, H.; Wang, C. BolTLP1, a Thaumatin-like Protein Gene, Confers Tolerance to Salt and Drought Stresses in Broccoli (Brassica oleracea L. var. Italica). Int. J. Mol. Sci. 2021, 22, 11132. [Google Scholar] [CrossRef] [PubMed]
- Pluskota, W.E.; Pupel, P.; Głowacka, K.; Okorska, S.B.; Jerzmanowski, A.; Nonogaki, H.; Górecki, R.J. Jasmonic acid and ethylene are involved in the accumulation of osmotin in germinating tomato seeds. J. Plant Physiol. 2019, 232, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Futamura, N.; Tani, N.; Tsumura, Y.; Nakajima, N.; Sakaguchi, M.; Shinohara, K. Characterization of genes for novel thaumatin-like proteins in Cryptomeria japonica. Tree Physiol. 2006, 26, 51–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, M.; Tong, Z.; Ding, C.; Wang, Z.; Sun, H.; Xia, Z.; Qi, M.; Wu, Y.; Liang, Y. Molecular Characterization of the Thaumatin-like Protein PR-NP24 in Tomato Fruits. J. Agric. Food Chem. 2019, 67, 13001–13009. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.P.; Chen, P.D.; Liu, D.J.; Kynast, R.; Friebe, B.; Velazhahan, R.; Muthukrishnan, S.; Gill, B.S. Development of wheat scab symptoms is delayed in transgenic wheat plants that constitutively express a rice thaumatin-like protein gene. Theor. Appl. Genet. 1999, 99, 755–760. [Google Scholar] [CrossRef]
- Velazhahan, R.; Muthu Krishnan, S. Transgenic tobacco plants constitutively overexpressing a rice thaumatin-like protein (PR-5) show enhanced résistance to Alternaria alternate. Biol. Plant. 2003, 47, 347–354. [Google Scholar] [CrossRef]
- El-kereamy, A.; El-sharkawy, I.; Ramamoorthy, R.; Taheri, A.; Errampalli, D.; Kumar, P.; Jayasankar, S. Prunus domestica pathogenesis-related protein-5 activates the defense response pathway and enhances the resistance to fungal infection. PLoS ONE 2011, 6, e17973. [Google Scholar] [CrossRef] [Green Version]
- Acharya, K.; Pal, A.K.; Gulati, A.; Kumar, S.; Singh, A.K.; Ahuja, P.S. Overexpression of Camellia sinensis thaumatin-like protein, CsTLP in potato confers enhanced resistance to Macrophomina phaseolina and Phytophthora infestans infection. Mol. Biotechnol. 2013, 54, 609–622. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Wu, J.; Zhang, Y.; Agüero, C.B.; Li, X.; Liu, S.; Wang, C.; Walker, M.A.; Lu, J. Overexpression of a thaumatin-like protein gene from Vitis amurensis improves downy mildew resistance in Vitis vinifera grapevine. Protoplasma 2017, 254, 1579–1589. [Google Scholar] [CrossRef]
- Odeny Ojola, P.; Nyaboga, E.N.; Njiru, P.N.; Orinda, G. Overexpression of rice thaumatin-like protein (Ostlp) gene in transgenic cassava results in enhanced tolerance to Colletotrichum gloeosporioides f. sp. manihotis. J. Genet. Eng. Biotechnol. 2018, 16, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Wardhan, V.; Pandey, A.; Chakraborty, S.; Chakraborty, N. Chickpea transcription factor CaTLP1 interacts with protein kinases, modulates ROS accumulation and promotes ABA-mediated stomatal closure. Sci. Rep. 2016, 6, 38121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.K.; Kumar, K.R.R.; Kumar, D.; Shukla, P.; Kirti, P.B. Characterization of a pathogen induced thaumatin-like protein gene AdTLP from Arachis diogoi, a wild peanut. PLoS ONE 2013, 8, 83963. [Google Scholar] [CrossRef] [PubMed]
- Munis, M.F.; Tu, L.; Deng, F.; Tan, J.; Xu, L. A thaumatin-like protein gene involved in cotton fiber secondary cell wall development enhances resistance against Verticillium dahliae and other stresses in transgenic tobacco. Biochem. Biophys. Res. Commun. 2010, 393, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.; Lei, Z.; Sumner, L.; Mysore, K.; Arakane, Y. Apoplastic extracts from a transgenic wheat line exhibiting lesion-mimic phenotype have multiple pathogenesis-related proteins that are antifungal. Mol. Plant Microbe Interact. 2004, 17, 1306–1317. [Google Scholar] [CrossRef] [Green Version]
- Grenier, J.; Potvin, C.; Asselin, A. Barley pathogenesis-related proteins with fungal cell wall lytic activity inhibit the growth of yeasts. Plant Physiol. 1993, 103, 1277–1283. [Google Scholar] [CrossRef] [PubMed]
- Mitchum, M.G.; Wang, X.; Wang, J.; Davis, E.L. Role of nematode peptides and other small molecules in plant parasitism. Annu. Rev. Phytopathol. 2012, 50, 175–195. [Google Scholar] [CrossRef]
- Kirino, H.; Yoshimoto, K.; Shinya, R. Thaumatin-like proteins and a cysteine protease inhibitor secreted by the pine wood nematode Bursaphelenchus xylophilus induce cell death in Nicotiana benthamiana. PLoS ONE 2020, 15, e0241613. [Google Scholar] [CrossRef]
- Shinya, R.; Morisaka, H.; Kikuchi, T.; Takeuchi, Y.; Ueda, M.; Futai, K. Secretome Analysis of the Pine Wood Nematode Bursaphelenchus xylophilus Reveals the Tangled Roots of Parasitism and Its Potential for Molecular Mimicry. PLoS ONE 2013, 8, e67377. [Google Scholar] [CrossRef]
- Breiteneder, H. Thaumatin-like proteins—A new family of pollen and fruit allergens. Allergy 2004, 59, 479–481. [Google Scholar] [CrossRef]
- Izumi, E.; Hidaka, S.; Hiroi, A.; Kinugasa, S.; Yano, E.; Zaima, N.; Moriyama, T. Thaumatin-Like Protein (Pru av 2) Is a Cherry Allergen That Triggers Percutaneous Sensitization in Mice. Foods 2021, 10, 134. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.; Chakrabarti, C. Crystal structure analysis of NP24-I: A thaumatin-like protein. Planta 2008, 228, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shumayla; Tyagi, S.; Alok, A.; Singh, K.; Upadhyay, S.K. Thaumatin-like protein kinases: Molecular characterization and transcriptional profiling in five cereal crops. Plant Sci. 2020, 290, 110317. [Google Scholar] [CrossRef]
- Chan, Y.W.; Tung, W.L.; Griffith, M.; Chow, K.C. Cloning of a cDNA encoding the thaumatin-like protein of winter rye (Secale cereale L. Musketeer) and its functional characterization. J. Exp. Bot. 1999, 50, 1627–1628. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.J.; Sturrock, R.; Ekramoddoullah, A.K.M. The superfamily of thaumatin-like proteins: Its origin, evolution, and expression towards biological function. Plant Cell Rep. 2010, 29, 419–436. [Google Scholar] [CrossRef] [PubMed]
- Petre, B.; Major, I.; Rouhier, N.; Duplessis, S. Genome-wide analysis of eukaryote thaumatin-like proteins (TLPs) with an emphasis on poplar. BMC Plant Biol. 2011, 11, 33. [Google Scholar] [CrossRef] [Green Version]
- Shiu, S.H.; Bleecker, A.B. Plant receptor-like kinase Gene family: Diversity, function, and signaling. Sci. Signal. 2001, 98, re22. [Google Scholar] [CrossRef]
- Wang, X.; Zafian, P.; Choudhary, M.; Lawton, M. The PR5K receptor protein kinase from Arabidopsis thaliana is structurally related to a family of plant defense proteins. Proc. Natl. Acad. Sci. USA 1996, 93, 2598–2602. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Bonos, S.; Meyer, W.A.; Day, P.R.; Belanger, F.C. Transgenic creeping bentgrass with delayed dollar spot symptoms. Mol. Breed. 2003, 11, 95–101. [Google Scholar] [CrossRef]
- Wan, Q.; Hongbo, S.; Zhaolong, X.; Jia, L.; Dayong, Z.; Yihong, H. Salinity Tolerance Mechanism of Osmotin and Osmotin-like Proteins: A Promising Candidate for Enhancing Plant Salt Tolerance. Curr. Genom. 2017, 18, 553–556. [Google Scholar] [CrossRef]
- Liu, J.; Han, D.; Shi, Y. Gene Cloning, Expression, and Antifungal Activities of Permatin from Naked Oat (Avena nuda). Probiotics Antimicrob. Proteins 2019, 11, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Skadsen, R.W.; Sathish, P.; Kaeppler, H.F. Expression of thaumatin-like permatin PR-5 genes switches from the ovary wall to the aleurone in developing barley and oat seeds. Plant Sci. 2000, 156, 11–22. [Google Scholar] [CrossRef]
- Thimme Gowda, C.; Purama, S.N.S.; Kammara, R. TLPdb: A Resource for Thaumatin-Like Proteins. Protein J. 2020, 39, 301–307. [Google Scholar] [CrossRef]
- Osmond, R.I.; Hrmova, M.; Fontaine, F.; Imberty, A.; Fincher, G.B. Binding interactions between barley thaumatin-like proteins and (1,3)-beta-D-glucans. Kinetics, specificity, structural analysis and biological implications. Eur. J. Biochem. 2001, 268, 4190–4199. [Google Scholar] [CrossRef]
- Snowdon, A.L. A Colour Atlas of Post-Harvest Diseases and Disorders of Fruits and Vegetables. Volume 1: General Introduction and Fruits; Wolfe Scientific Ltd.: London, UK, 1990. [Google Scholar]
- Akhter, A.; Hage-Ahmed, K.; Soja, G.; Steinkellner, S. Potential of Fusarium wilt-inducing chlamydospores, in vitro behaviour in root exudates and physiology of tomato in biochar and compost amended soil. Plant Soil 2016, 406, 425–440. [Google Scholar] [CrossRef] [Green Version]
- Gálvez, L.; Urbaniak, M.; Waśkiewicz, A.; Stępień, Ł.; Palmero, D. Fusarium proliferatum—Causal agent of garlic bulb rot in Spain: Genetic variability and mycotoxin production. Food Microbiol. 2017, 67, 41–48. [Google Scholar] [CrossRef]
- Chand, S.K.; Nanda, S.; Mishra, R.; Joshi, R.K. Multiple garlic (Allium sativum L.) microRNAs regulate the immunity against the basal rot fungus Fusarium oxysporum f. sp. cepae. Plant Sci. 2017, 257, 9–21. [Google Scholar] [CrossRef]
- Rout, E.; Nanda, S.; Joshi, R.K. Molecular characterization and heterologous expression of a pathogen induced PR5 gene from garlic (Allium sativum L.) conferring enhanced resistance to necrotrophic fungi. Eur. J. Plant Pathol. 2016, 144, 345–360. [Google Scholar] [CrossRef]
- Anisimova, O.K.; Shchennikova, A.V.; Kochieva, E.Z.; Filyushin, M.A. Pathogenesis-Related Genes of PR1, PR2, PR4, and PR5 Families Are Involved in the Response to Fusarium Infection in Garlic (Allium sativum L.). Int. J. Mol. Sci. 2021, 22, 6688. [Google Scholar] [CrossRef]
- Smole, U.; Bublin, M.; Radauer, C.; Ebner, C.; Breiteneder, H. Mal d 2, the thaumatin-like allergen from apple, is highly resistant to gastrointestinal digestion and thermal processing. Int. Arch. Allergy Immunol. 2008, 147, 289–298. [Google Scholar] [CrossRef]
- Palacín, A.; Tordesillas, L.; Gamboa, P.; Sanchez-Monge, R.; Cuesta-Herranz, J.; Sanz, M.L.; Barber, D.; Salcedo, G.; Díaz-Perales, A. Characterization of peach thaumatin-like proteins and their identification as major peach allergens. Clin. Exp. Allergy 2010, 40, 1422–1430. [Google Scholar] [CrossRef] [PubMed]
- Gavrović-Jankulović, M.; Spasić, M.; Cirković Velicković, T.; Stojanović, M.; Inić-Kanada, A.; Dimitrijević, L.; Lindner, B.; Petersen, A.; Becker, W.M.; Jankov, R.M. Quantification of the thaumatin-like kiwi allergen by a monoclonal antibody-based ELISA. Mol. Nutr. Food Res. 2008, 52, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Yamaguchi-Shinozaki, K. ABA signaling in stress-response and seed development. Plant Cell Rep. 2013, 32, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Lin, T.; Wang, M.; Qi, X. An AC-Rich Bean Element Serves as an Ethylene-Responsive Element in Arabidopsis. Plants 2020, 9, 1033. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Carbajosa, J.; Moose, S.P.; Parsons, R.L.; Schmidt, R.J. A maize zinc-finger protein binds the prolamin box in zein gene promoters and interacts with the basic leucine zipper transcriptional activator Opaque2. Proc. Natl. Acad. Sci. USA 1997, 94, 7685–7690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Sun, M.; Jiang, X.; Sun, H.; Dang, X.; Cong, H.; Qiao, F. Glycinebetaine Biosynthesis in Response to Osmotic Stress Depends on Jasmonate Signaling in Watermelon Suspension Cells. Front. Plant Sci. 2018, 9, 1469. [Google Scholar] [CrossRef] [Green Version]
- Geffers, R.; Sell, S.; Cerff, R.; Hehl, R. The TATA box and a Myb binding site are essential for anaerobic expression of a maize GapC4 minimal promoter in tobacco. Biochim. Biophys. Acta 2001, 1521, 120–125. [Google Scholar] [CrossRef]
- Zhao, J.P.; Jiang, X.L.; Zhang, B.Y.; Su, X.H. Involvement of microRNA-mediated gene expression regulation in the pathological development of stem canker disease in Populus trichocarpa. PLoS ONE 2012, 7, e44968. [Google Scholar] [CrossRef]
- Filyushin, M.A.; Anisimova, O.K.; Kochieva, E.Z.; Shchennikova, A.V. Genome-Wide Identification and Expression of Chitinase Class I Genes in Garlic (Allium sativum L.) Cultivars Resistant and Susceptible to Fusarium proliferatum. Plants 2021, 10, 720. [Google Scholar] [CrossRef]
- Hauenschild, F.; Favre, A.; Schnitzler, J.; Michalak, I.; Freiberg, M.; Muellner-Riehl, A.N. Spatio-temporal evolution of Allium L. in the Qinghai-Tibet-Plateau region: Immigration and in situ radiation. Plant Divers. 2017, 39, 167–179. [Google Scholar] [CrossRef]
- Fritsch, R.M.; Friesen, N. Evolution, domestication and taxonomy. In Allium Crop Science: Recent Advances; Rabinowitch, H.D., Currah, L., Eds.; CAB International: Wallingford, UK, 2002; pp. 5–30. [Google Scholar]
- Taylor, A.; Vagany, V.; Barbara, D.J.; Thomas, B.; Pink, D.A.C.; Jones, J.E.; Clarkson, J.P. Identification of differential resistance to six Fusarium oxysporum f. sp. cepae isolates in commercial onion cultivars through the development of a rapid seedling assay. Plant Pathol. 2013, 62, 103–111. [Google Scholar] [CrossRef]
- Borde, M.; Dudhane, M.; Jite, P. Growth, water use efficiency and antioxidant defense responses of mycorrhizal and non mycorrhizal Allium sativum L. under drought stress condition. Ann. Plant Sci. 2012, 1, 6–11. [Google Scholar]
- Son, J.H.; Park, K.C.; Lee, S.I.; Kim, H.H.; Kim, J.H.; Kim, S.H.; Kim, N.S. Isolation of cold-responsive genes from garlic, Allium sativum. Genes Genomes 2012, 34, 93–101. [Google Scholar] [CrossRef]
- Sun, X.; Zhu, S.; Li, N.; Cheng, Y.; Zhao, J.; Qiao, X.; Lu, L.; Liu, S.; Wang, Y.; Liu, C.; et al. A Chromosome-Level Genome Assembly of Garlic (Allium sativum) Provides Insights into Genome Evolution and Allicin Biosynthesis. Mol. Plant 2020, 13, 1328–1339. [Google Scholar] [CrossRef] [PubMed]
- Anisimova, O.K.; Seredin, T.M.; Danilova, O.A.; Filyushin, M. First Report of Fusarium proliferatum Causing Garlic clove Rot in Russian Federation. Plant Dis. 2021, 105, 3308. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Lyu, H.M.; Zhu, K.; Van de Peer, Y.; Max Cheng, Z.M. The emergence and evolution of intron-poor and intronless genes in intron-rich plant gene families. Plant J. 2021, 105, 1072–1082. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Park, J.Y.; Kim, K.S.; Ko, M.K.; Cheong, S.J.; Oh, B.J. A thaumatin-like gene in nonclimacteric pepper fruits used as molecular marker in probing disease resistance, ripening, and sugar accumulation. Plant Mol. Biol. 2002, 49, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Armentia, A.; Martín-Armentia, S.; Pineda, F.; Martín-Armentia, B.; Castro, M.; Fernández, S.; Moro, A.; Castillo, M. Allergic hypersensitivity to garlic and onion in children and adults. Allergol. Immunopathol. 2020, 48, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C.; Felix, G. Plants and animals: A different taste for microbes? Curr. Opin. Plant Biol. 2005, 8, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Akbudak, M.A.; Yildiz, S.; Filiz, E. Pathogenesis related protein-1 (PR-1) genes in tomato (Solanum lycopersicum L.): Bioinformatics analyses and expression profiles in response to drought stress. Genomics 2020, 112, 4089–4099. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, W.; Sang, X.; Wang, T.; Gong, H.; Zhao, Y.; Zhao, P.; Wang, H. Genome-Wide Identification of the Thaumatin-like Protein Family Genes in Gossypium barbadense and Analysis of Their Responses to Verticillium dahliae Infection. Plants 2021, 10, 2647. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Zhao, X.; Geng, L.; Fu, L.; Lu, Y.; Liu, Q.; Jiang, X. Analysis of the thaumatin-like genes of Rosa chinensis and functional analysis of the role of RcTLP6 in salt stress tolerance. Planta 2021, 254, 118. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, H.; Zhu, J.K. Osmotic stress signaling via protein kinases. Cell Mol. Life Sci. 2012, 69, 3165–3173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deihimi, T.; Niazi, A.; Ebrahimi, M.; Kajbaf, K.; Fanaee, S.; Bakhtiarizadeh, M.R.; Ebrahimie, E. Finding the undiscovered roles of genes: An approach using mutual ranking of coexpressed genes and promoter architecture-case study: Dual roles of thaumatin like proteins in biotic and abiotic stresses. Springerplus 2012, 1, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, H.; Zhu, W.; Li, W.; Wang, F. Transcriptome Analysis Reveals the Effects of Chinese Chive (Allium tuberosum R.) Extract on Fusarium oxysporum f. sp. radicis-lycopersici Spore Germination. Cur. Microbiol. 2020, 77, 855–864. [Google Scholar] [CrossRef]
- Zuo, G.W.; Li, C.Y.; Li, B.; Wei, Y.R.; Hu, C.H.; Yang, Q.S.; Yang, J.; Sheng, O.; Kuang, R.B.; Deng, G.M.; et al. The toxic mechanism and bioactive components of Chinese leek root exudates acting against Fusarium oxysporum f. sp. cubense, tropical race 4. Eur. J. Plant Pathol. 2015, 143, 447–460. [Google Scholar] [CrossRef]
- Abdelrahman, M.; El-Sayed, M.; Sato, S.; Hirakawa, H.; Ito, S.I.; Tanaka, K.; Mine, Y.; Sugiyama, N.; Suzuki, Y.; Yamauchi, N.; et al. RNA-sequencing-based transcriptome and biochemical analyses of steroidal saponin pathway in a complete set of Allium fistulosum-A. cepa monosomic addition lines. PLoS ONE 2017, 12, e0181784. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Sugui, J.A.; Deising, H.B. Isolation of infection-specific sequence tags expressed during early stages of maize anthracnose disease development. Mol. Plant Pathol. 2002, 3, 197–203. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Genomic Location (Strand) | A. thaliana Homolog | Transcript ID in RNA-Seq Database | Size (bp) | Exons | CDS (bp) | Protein | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Size (aa) | MW (kDa) | pI | Signal Peptide | GH64 Domain | GRAVY | |||||||
| AsTLP1 | Chr1:491780133..491781147 (+) | Pathogenesis-related thaumatin superfamily protein NP_001031792.1 | Asa2G02997.1 | 1013 | 2 | 930 | 309 | 32.29 | 4.93 | 1–24 | 31–245 | 0.126 |
| AsTLP2 | Chr1:996536859..996536201 (−) | Osmotin 34 NP_192902.1 | Asa2G01042.1 | 660 | 1 | 660 | 219 | 23.46 | 5.52 | 1–20 | 23–219 | −0.157 |
| AsTLP3 (AsPR5a) | Chr2:821219552..821220214 (−) | Asa2G01043.1 | 663 | 1 | 663 | 220 | 23.46 | 4.71 | 1–21 | 24–220 | −0.085 | |
| AsTLP4 | Chr2:1584165307..1584166065 (−) | TLP AAD02499.1 | Asa3G06018.1 | 759 | 1 | 759 | 252 | 25.92 | 4.79 | 1–27 | 30–251 | 0.069 |
| AsTLP5 | Chr2:1617805168..1617805788 (−) | Osmotin 34 NP_192902.1 | Asa4G02103.1 Asa4G02102.1 | 621 | 1 | 621 | 206 | 21.89 | 4.76 | 1–26 | 29–206 | −0.289 |
| AsTLP6 | Chr2:1617847011..1617847625 (−) | 615 | 1 | 615 | 204 | 21.74 | 4.76 | 1–24 | 27–204 | −0.263 | ||
| AsTLP7 | Chr2:1617850920..1617851543 (−) | Asa4G02101.1 | 624 | 1 | 624 | 207 | 22.02 | 4.77 | 1–27 | 30–207 | −0.295 | |
| AsTLP8 (AsPR5b) | Chr2:1618362333..1618362995 (+) | Asa4G02099.1 | 663 | 1 | 663 | 220 | 23.50 | 4.71 | 1–21 | 24–220 | −0.126 | |
| AsTLP9 (AsPR5c) | Chr2:1618377059..1618377721 (+) | Asa4G02100.1 | 663 | 1 | 663 | 220 | 23.59 | 4.74 | 1–21 | 24–220 | −0.164 | |
| AsTLP10 | Chr2:1618405507..1618406165 (−) | Asa3G05858.1 | 660 | 1 | 660 | 219 | 23.43 | 4.71 | 1–20 | 23–219 | −0.126 | |
| AsTLP11 | Chr2:1622416758..1622417381 (−) | Asa3G05840.1 | 624 | 1 | 624 | 207 | 22.08 | 5.70 | 1–27 | 30–207 | −0.211 | |
| AsTLP12 | Chr2:1684781475..1684782236 (+) | TLP AAD02499.1 | Asa3G06302.1 | 762 | 1 | 762 | 253 | 26.16 | 4.27 | 1–29 | 32–252 | 0.056 |
| AsTLP13 | Chr2:1685003219..1685003978 (−) | Asa3G06331.1 | 759 | 1 | 759 | 252 | 26.28 | 4.24 | 1–28 | 31–251 | 0.012 | |
| AsTLP14 | Chr2:1685049653..1685050414 (−) | no detected | 762 | 1 | 762 | 253 | 26.06 | 4.18 | 1–29 | 32–252 | 0.087 | |
| AsTLP15 | Chr2:1685078657..1685079418 (−) | no detected | 762 | 1 | 762 | 253 | 26.06 | 4.18 | 1–29 | 32–252 | 0.068 | |
| AsTLP16 | Chr2:1685746745..1685747579 (+) | Asa3G06310.1 | 835 | 2 | 747 | 248 | 25.30 | 4.59 | 1–25 | 27–247 | 0.031 | |
| AsTLP17 | Chr2:1686634047..1686634808 (+) | Pathogenesis-related thaumatin superfamily protein NP_173432.2 | Asa3G06319.1 | 762 | 1 | 762 | 253 | 26.09 | 4.18 | 1–29 | 32–252 | 0.054 |
| AsTLP18 | Chr2:1687188296..1687189055 (−) | TLP AAD02499.1 | Asa3G06331.1 | 759 | 1 | 759 | 252 | 26.30 | 4.24 | 1–28 | 31–251 | 0.013 |
| AsTLP19 | Chr2:1687209664..1687210422 (−) | Asa3G06329.1 | 759 | 1 | 759 | 252 | 26.21 | 4.27 | 1–28 | 31–251 | 0.025 | |
| AsTLP20 | Chr2:1724629365..1724630114 (+) | Pathogenesis-related thaumatin superfamily protein NP_973870.1 | Asa7G02264.1 | 750 | 1 | 750 | 249 | 25.42 | 4.39 | 1–26 | 28–248 | 0.081 |
| AsTLP21 | Chr4:777618126..777620378 (+) | Pathogenesis-related thaumatin superfamily protein NP_568046.1 | Asa4G00901.1 | 2253 | 3 | 972 | 323 | 33.86 | 4.50 | 1–19 | 26–232 | −0.008 |
| AsTLP22 | Chr6:2008642583..2008643344 (+) | TLP AAD02499.1 | Asa3G06325.1 | 762 | 1 | 762 | 253 | 26.09 | 4.18 | 1–29 | 32–252 | 0.085 |
| AsTLP23 | Chr7:682376071..682376964 (+) | Pathogenesis-related thaumatin superfamily protein NP_001119140.1 | Asa5G05974.1 | 894 | 1 | 894 | 297 | 31.13 | 4.68 | 1–21 | 23–242 | 0.115 |
| AsTLP24 | Chr8:1265530915..1265533486 (−) | Pathogenesis-related thaumatin superfamily protein NP_177503.1 | Asa7G04908.1 | 2572 | 2 | 753 | 250 | 25.97 | 9.07 | 1–26 | 31–247 | 0.081 |
| AsTLP25 | Chr8:1394582992..1394583731 (+) | Pathogenesis-related thaumatin superfamily protein NP_001324474.1 | Asa7G04378.1 | 741 | 1 | 741 | 246 | 26.03 | 8.10 | 1–21 | 24–245 | −0.005 |
| AsTLP26 | Scaffold9088: 51512..52273 | Pathogenesis-related thaumatin superfamily protein NP_173432.2 | Asa3G06323.1 | 762 | 1 | 762 | 253 | 26.09 | 4.18 | 1–29 | 32–252 | 0.054 |
| AsTLP27 | Scaffold9089: 39596..38835 | TLP AAD02499.1 | Asa3G06322.1 | 762 | 1 | 762 | 253 | 26.12 | 4.18 | 1–29 | 32–252 | 0.075 |
| AsTLP28 | Scaffold9089: 40346..41107 | 762 | 1 | 762 | 253 | 26.12 | 4.18 | 1–29 | 32–252 | 0.075 | ||
| AsTLP29 | Scaffold9089: 89114..89875 | Asa3G06320.1 | 762 | 1 | 762 | 253 | 26.06 | 4.18 | 1–29 | 32–252 | 0.044 | |
| AsTLP30 | Scaffold9091: 149728..150486 | Asa3G06315.1 | 759 | 1 | 759 | 252 | 26.28 | 4.24 | 1–28 | 31–251 | 0.006 | |
| AsTLP31 | Scaffold12619: 87332..88248 | no detected | 917 | 3 | 738 | 245 | 26.47 | 5.75 | 1–22 | 25–244 | −0.134 | |
| AsTLP32 | Scaffold12619: 99641..100558 | no detected | 918 | 3 | 738 | 245 | 26.46 | 5.75 | 1–22 | 25–244 | −0.149 | |
| Gene | NCBI ID cv. Sarmat/cv. Strelets | cv. Sarmat | cv. Strelets |
|---|---|---|---|
| SNPs (aa Substitution) | |||
| AsTLP7 | OM386716/OM386717 | c.183A>C, c.218G>C (p.G73A), c.219A>C (p.G73A), c.225A>C, c.233A>G (p.N78S), c.243G>C, c.249G>A, c.252T>C, c.276T>C, c.279T>C, c.303A>C, c.333T>C, c.357T>C, c.378C>G, c.384C>T, c.399 >T, c.479>T (p.T160M), c.615C>T | |
| AsTLP16 | OM386718/OM386719 | c.141T>C, c.162T>C, c.406C>T, c.476C>G (p. A159G) | |
| AsTLP20 | OM386720/OM386721 | c.49T>C (p.S17P), c.53G>T (p.C18F), c.67T>C (p.S23P), c.70A>G, c.75A>T, c.184G>C (p.G62R) | |
| c. 670T>C (p.Y224H) | |||
| AsTLP21 | OM386722/OM386723 | c.22A>G, c.141G>A, c.453A>G, c.554G>A, c.570C>T, c.573T>G, c.738A>G, c.804A>T, c.929C>G (p.A310G) | |
| AsTLP23 | OM386724/OM386725 | c.T30>C, c.A45>T (p.Q15H), c.A99>G, c.T147>G, c.C165>G, c.C588>G, c.C684>T, c.C821>G (p. A274G), c.C847>T, c.C849>T, c.T851>C | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anisimova, O.K.; Kochieva, E.Z.; Shchennikova, A.V.; Filyushin, M.A. Thaumatin-like Protein (TLP) Genes in Garlic (Allium sativum L.): Genome-Wide Identification, Characterization, and Expression in Response to Fusarium proliferatum Infection. Plants 2022, 11, 748. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11060748
Anisimova OK, Kochieva EZ, Shchennikova AV, Filyushin MA. Thaumatin-like Protein (TLP) Genes in Garlic (Allium sativum L.): Genome-Wide Identification, Characterization, and Expression in Response to Fusarium proliferatum Infection. Plants. 2022; 11(6):748. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11060748
Chicago/Turabian StyleAnisimova, Olga K., Elena Z. Kochieva, Anna V. Shchennikova, and Mikhail A. Filyushin. 2022. "Thaumatin-like Protein (TLP) Genes in Garlic (Allium sativum L.): Genome-Wide Identification, Characterization, and Expression in Response to Fusarium proliferatum Infection" Plants 11, no. 6: 748. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11060748