
Stocky1, a Novel Gene Involved in Maize Seedling Development and Cuticle Integrity


 ,
,  ,
,
Abstract
:1. Introduction
1.1. Embryogenesis and Seedling Architecture
1.2. The Cuticle Role
1.3. Mutants for the Study of Seedling Development in Maize
2. Results
2.1. Origin and Isolation of Defective Seedlings “des” Mutants
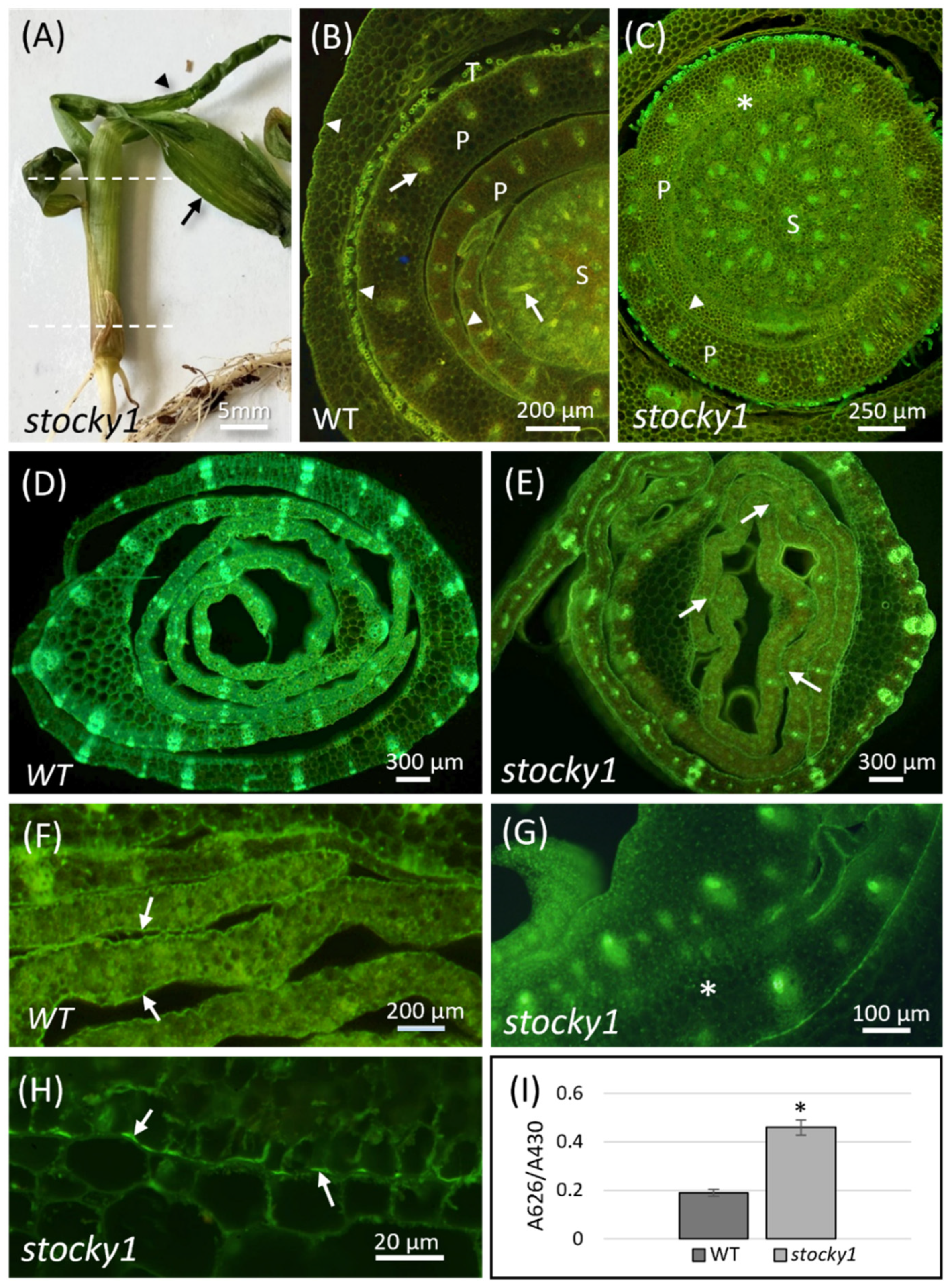
2.2. Phenotypic Characterization of Stocky1
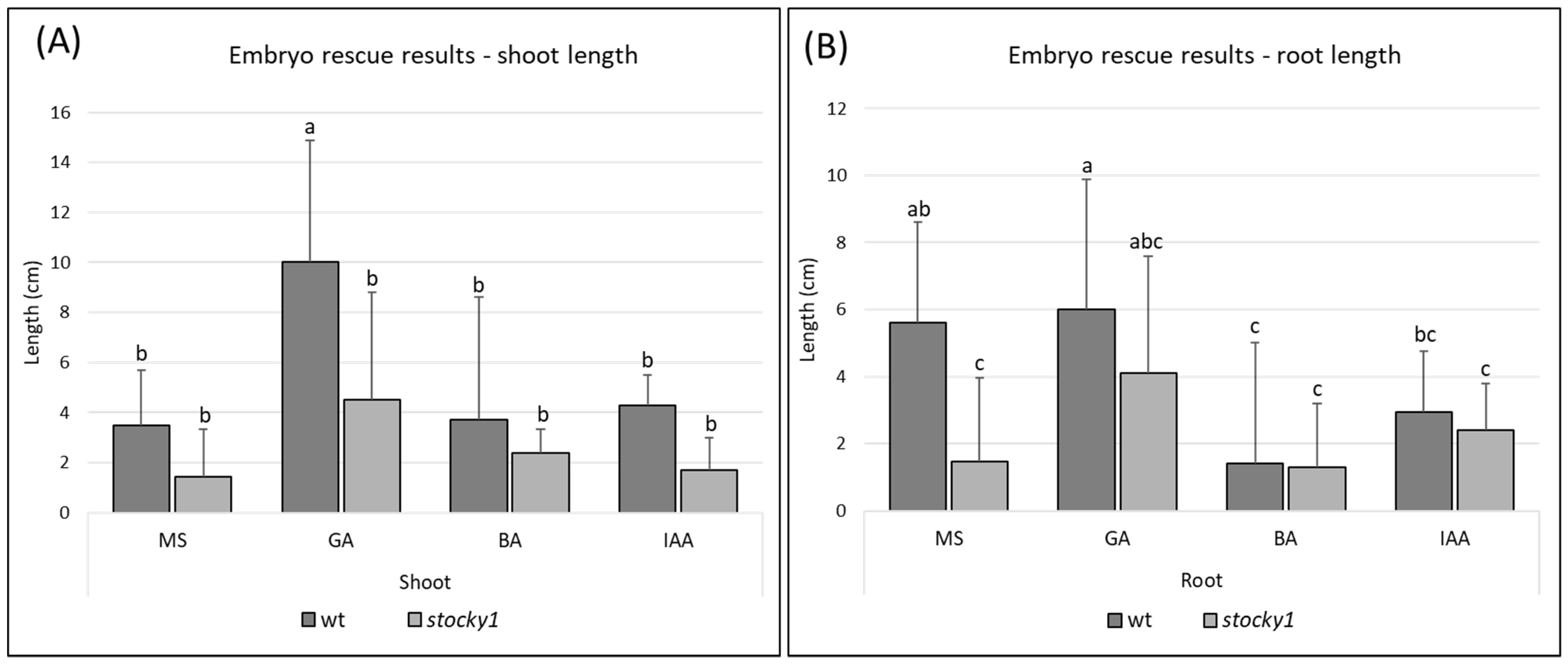
2.3. Role of Hormones
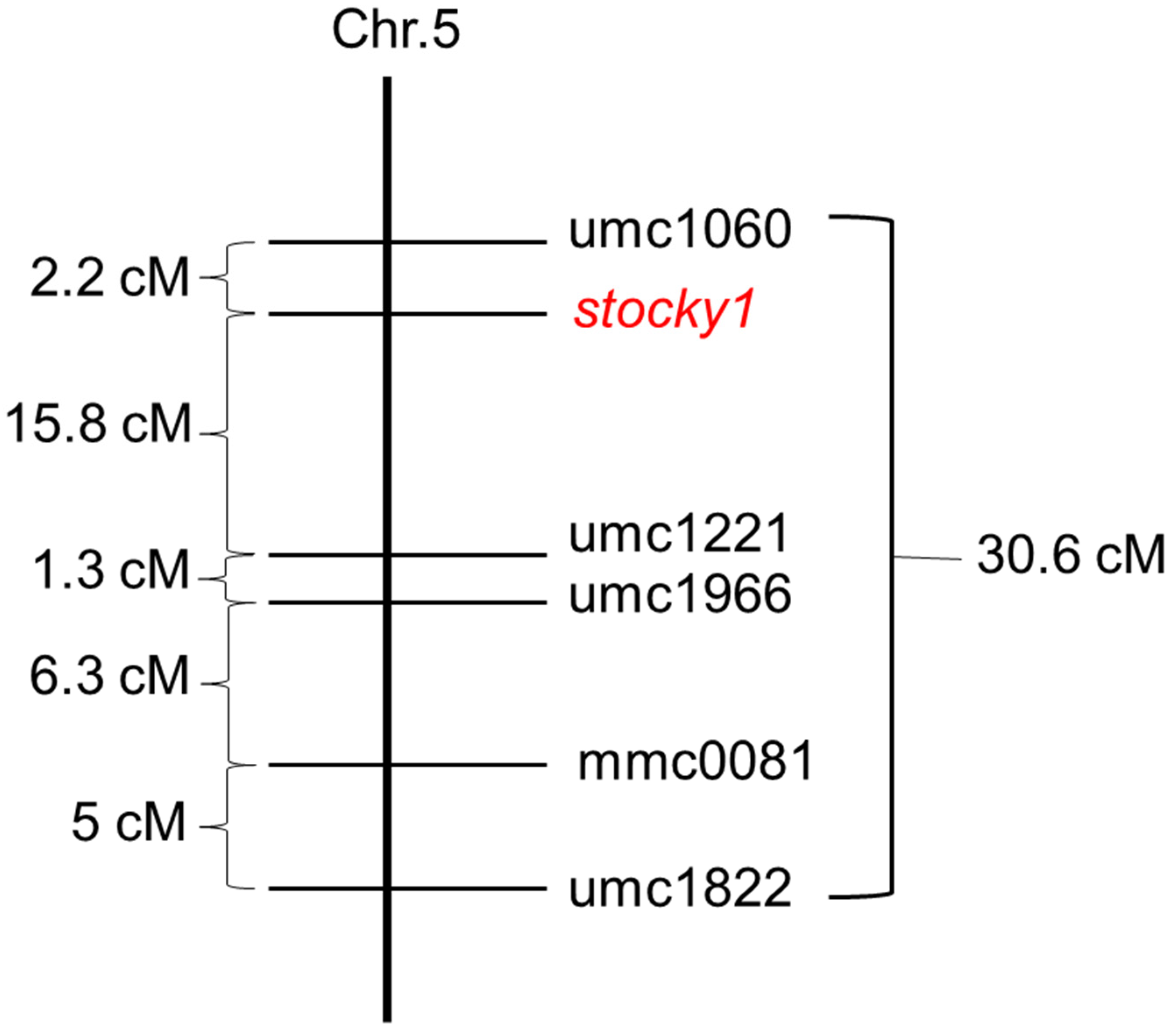
2.4. Molecular Genetic Analysis of Stocky1
2.5. Does Stocky1 Encode for a 3-ketoacyl-CoA Synthase (KCS) Gene?
3. Discussion
3.1. Stocky1 Organs’ Fusion and the Cuticle
3.2. Genetic Analysis and Gene Candidate
4. Materials and Methods
4.1. Isolation of the Mutants and Genetic Analysis
4.2. Gametophytic Selection
4.3. Fluorescence Microscopy
4.4. Toluidine Blue Permeability Test
4.5. Embryo Culture
4.6. Mapping of Stocky1
4.7. Candidate Gene Sequence Analysis
4.8. RNA, cDNA Preparation, and Quantitative Gene Expression Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Doll, N.M.; Depège-Fargeix, N.; Rogowsky, P.M.; Widiez, T. Signaling in Early Maize Kernel Development. Mol. Plant 2017, 10, 375–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandler, J.; Nardmann, J.; Werr, W. Plant Development Revolves around Axes. Trends Plant Sci. 2008, 13, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, D.; Widiez, T.; Di Tommaso, S.; Rositi, H.; Adrien, J.; Maire, E.; Langer, M.; Olivier, C.; Peyrin, F.; Rogowsky, P. Fast Virtual Histology Using X-Ray in-Line Phase Tomography: Application to the 3D Anatomy of Maize Developing Seeds. Plant Methods 2015, 11, 55. [Google Scholar] [CrossRef]
- Harada, J.J. Seed Maturation and Control of Germination. In Cellular and Molecular Biology of Plant Seed Development; Larkins, B.A., Vasil, I.K., Eds.; Advances in Cellular and Molecular Biology of Plants; Springer: Dordrecht, The Netherlands, 1997; pp. 545–592. ISBN 978-94-015-8909-3. [Google Scholar]
- Widiez, T.; Ingram, G.; Gutiérrez-Marcos, J.F. Embryo-Endosperm-Sporophyte Interactions in Maize Seeds. In Maize Kernel Development; Cabi: Wallingford, UK, 2017. [Google Scholar]
- Bewley, D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-4692-7. [Google Scholar]
- Ingram, G.; Nawrath, C. The Roles of the Cuticle in Plant Development: Organ Adhesions and Beyond. J. Exp. Bot. 2017, 68, 5307–5321. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Kong, L.; Zhi, P.; Chang, C. Update on Cuticular Wax Biosynthesis and Its Roles in Plant Disease Resistance. Int. J. Mol. Sci. 2020, 21, 5514. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Suh, M.C. Advances in the Understanding of Cuticular Waxes in Arabidopsis Thaliana and Crop Species. Plant Cell Rep. 2015, 34, 557–572. [Google Scholar] [CrossRef]
- Kunst, L.; Samuels, L. Plant Cuticles Shine: Advances in Wax Biosynthesis and Export. Curr. Opin. Plant Biol. 2009, 12, 721–727. [Google Scholar] [CrossRef]
- Liu, X.; Bourgault, R.; Strable, J.; Galli, M.; Chen, Z.; Dong, J.; Molina, I.; Gallavotti, A. The FUSED LEAVES1/ADHERENT1 Regulatory Module Is Required For Maize Cuticle Development And Organ Separation. New Phytol. 2020, 229, 388–402. [Google Scholar] [CrossRef]
- Bourgault, R.; Matschi, S.; Vasquez, M.; Qiao, P.; Sonntag, A.; Charlebois, C.; Mohammadi, M.; Scanlon, M.J.; Smith, L.G.; Molina, I. Constructing Functional Cuticles: Analysis of Relationships between Cuticle Lipid Composition, Ultrastructure and Water Barrier Function in Developing Adult Maize Leaves. Ann. Bot. 2020, 125, 79–91. [Google Scholar] [CrossRef]
- Qiao, P.; Bourgault, R.; Mohammadi, M.; Matschi, S.; Philippe, G.; Smith, L.G.; Gore, M.A.; Molina, I.; Scanlon, M.J. Transcriptomic Network Analyses Shed Light on the Regulation of Cuticle Development in Maize Leaves. Proc. Natl. Acad. Sci. USA 2020, 117, 12464–12471. [Google Scholar] [CrossRef]
- Bianchi, A.; Marchesi, G. The Surface of the Leaf in Normal and Glossy Maize Seedlings. Z. Vererb. 1960, 91, 214–219. [Google Scholar] [CrossRef]
- Zheng, J.; He, C.; Qin, Y.; Lin, G.; Park, W.D.; Sun, M.; Li, J.; Lu, X.; Zhang, C.; Yeh, C.-T.; et al. Co-Expression Analysis Aids in the Identification of Genes in the Cuticular Wax Pathway in Maize. Plant J. 2019, 97, 530–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Rocca, N.; Manzotti, P.S.; Cavaiuolo, M.; Barbante, A.; Dalla Vecchia, F.; Gabotti, D.; Gendrot, G.; Horner, D.S.; Krstajic, J.; Persico, M.; et al. The Maize Fused Leaves1 (Fdl1) Gene Controls Organ Separation in the Embryo and Seedling Shoot and Promotes Coleoptile Opening. J. Exp. Bot. 2015, 66, 5753–5767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, N. Organ and Cell Fusions in the Adherent1 Mutant in Maize. Int. J. Plant Sci. 1998, 159, 702–715. [Google Scholar] [CrossRef]
- Guo, H.-S.; Zhang, Y.-M.; Sun, X.-Q.; Li, M.-M.; Hang, Y.-Y.; Xue, J.-Y. Evolution of the KCS Gene Family in Plants: The History of Gene Duplication, Sub/Neofunctionalization and Redundancy. Mol. Genet. Genom. 2016, 291, 739–752. [Google Scholar] [CrossRef]
- Lin, M.; Matschi, S.; Vasquez, M.; Chamness, J.; Kaczmar, N.; Baseggio, M.; Miller, M.; Stewart, E.L.; Qiao, P.; Scanlon, M.J.; et al. Genome-Wide Association Study for Maize Leaf Cuticular Conductance Identifies Candidate Genes Involved in the Regulation of Cuticle Development. G3 Genes Genomes Genet. 2020, 10, 1671–1683. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Du, Y.; He, C.; Dietrich, C.R.; Li, J.; Ma, X.; Wang, R.; Liu, Q.; Liu, S.; Wang, G.; et al. Maize Glossy6 Is Involved in Cuticular Wax Deposition and Drought Tolerance. J. Exp. Bot. 2019, 70, 3089–3099. [Google Scholar] [CrossRef] [Green Version]
- Dolfini, S.; Consonni, G.; Viotti, C.; Dal Prà, M.; Saltini, G.; Giulini, A.; Pilu, R.; Malgioglio, A.; Gavazzi, G. A Mutational Approach to the Study of Seed Development in Maize. J. Exp. Bot. 2007, 58, 1197–1205. [Google Scholar] [CrossRef]
- Landoni, M.; Gavazzi, G.; Rascio, N.; Vecchia, F.D.; Consonni, G.; Dolfini, S. A Maize Mutant with an Altered Vascular Pattern. Ann. Bot. 2000, 85, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Dolfini, S.; Landoni, M.; Consonni, G.; Rascio, N.; Vecchia, F.D.; Gavazzi, G. The Maize Lilliputian Mutation Is Responsible for Disrupted Morphogenesis and Minute Stature. Plant J. 1999, 17, 11–17. [Google Scholar] [CrossRef]
- Gavazzi, G.; Dolfini, S.; Galbiati, M.; Helentjaris, T.; Landoni, M.; Pelucchi, N.; Todesco, G. Mutants Affecting Germination and Early Seedling Development in Maize. Maydica 1993, 38, 265–274. [Google Scholar]
- Xue, D.; Zhang, X.; Lu, X.; Chen, G.; Chen, Z.-H. Molecular and Evolutionary Mechanisms of Cuticular Wax for Plant Drought Tolerance. Front. Plant Sci. 2017, 8, 621. [Google Scholar] [CrossRef] [PubMed]
- Sinha, N.; Lynch, M. Fused Organs in the Adherent1 Mutation in Maize Show Altered Epidermal Walls with No Perturbations in Tissue Identities. Planta 1998, 206, 184–195. [Google Scholar] [CrossRef]
- Lolle, S.J.; Cheung, A.Y.; Sussex, I.M. Fiddlehead: An Arabidopsis Mutant Constitutively Expressing an Organ Fusion Program That Involves Interactions between Epidermal Cells. Dev. Biol. 1992, 152, 383–392. [Google Scholar] [CrossRef]
- Lolle, S.J.; Berlyn, G.P.; Engstrom, E.M.; Krolikowski, K.A.; Reiter, W.-D.; Pruitt, R.E. Developmental Regulation of Cell Interactions in TheArabidopsis Fiddlehead-1Mutant: A Role for the Epidermal Cell Wall and Cuticle. Dev. Biol. 1997, 189, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Pruitt, R.E.; Vielle-Calzada, J.-P.; Ploense, S.E.; Grossniklaus, U.; Lolle, S.J. FIDDLEHEAD, a Gene Required to Suppress Epidermal Cell Interactions in Arabidopsis, Encodes a Putative Lipid Biosynthetic Enzyme. Proc. Natl. Acad. Sci. USA 2000, 97, 1311–1316. [Google Scholar] [CrossRef] [Green Version]
- Kurdyukov, S.; Faust, A.; Nawrath, C.; Bär, S.; Voisin, D.; Efremova, N.; Franke, R.; Schreiber, L.; Saedler, H.; Métraux, J.-P.; et al. The Epidermis-Specific Extracellular BODYGUARD Controls Cuticle Development and Morphogenesis in Arabidopsis. Plant Cell 2006, 18, 321–339. [Google Scholar] [CrossRef] [Green Version]
- Kurdyukov, S.; Faust, A.; Trenkamp, S.; Bär, S.; Franke, R.; Efremova, N.; Tietjen, K.; Schreiber, L.; Saedler, H.; Yephremov, A. Genetic and Biochemical Evidence for Involvement of HOTHEAD in the Biosynthesis of Long-Chain α-,ω-Dicarboxylic Fatty Acids and Formation of Extracellular Matrix. Planta 2006, 224, 315–329. [Google Scholar] [CrossRef]
- Panikashvili, D.; Savaldi-Goldstein, S.; Mandel, T.; Yifhar, T.; Franke, R.B.; Höfer, R.; Schreiber, L.; Chory, J.; Aharoni, A. The Arabidopsis DESPERADO/AtWBC11 Transporter Is Required for Cutin and Wax Secretion. Plant Physiol. 2007, 145, 1345–1360. [Google Scholar] [CrossRef] [Green Version]
- Wellesen, K.; Durst, F.; Pinot, F.; Benveniste, I.; Nettesheim, K.; Wisman, E.; Steiner-Lange, S.; Saedler, H.; Yephremov, A. Functional Analysis of the LACERATA Gene of Arabidopsis Provides Evidence for Different Roles of Fatty Acid Omega-Hydroxylation in Development. Proc. Natl. Acad. Sci. USA 2001, 98, 9694–9699. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Goodwin, S.M.; Boroff, V.L.; Liu, X.; Jenks, M.A. Cloning and Characterization of the WAX2 Gene of Arabidopsis Involved in Cuticle Membrane and Wax Production. Plant Cell 2003, 15, 1170–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, H.; Molina, I.; Shockey, J.; Browse, J. Organ Fusion and Defective Cuticle Function in a Lacs1 Lacs2 Double Mutant of Arabidopsis. Planta 2010, 231, 1089–1100. [Google Scholar] [CrossRef] [PubMed]
- Javelle, M.; Vernoud, V.; Rogowsky, P.M.; Ingram, G.C. Epidermis: The Formation and Functions of a Fundamental Plant Tissue. New Phytol. 2011, 189, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Joubès, J.; Raffaele, S.; Bourdenx, B.; Garcia, C.; Laroche-Traineau, J.; Moreau, P.; Domergue, F.; Lessire, R. The VLCFA Elongase Gene Family in Arabidopsis Thaliana: Phylogenetic Analysis, 3D Modelling and Expression Profiling. Plant Mol. Biol. 2008, 67, 547–566. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A.A.; Stenback, K.E.; Flyckt, K.; Hoang, T.; Perera, M.A.D.; Nikolau, B.J. A Single-Cell Platform for Reconstituting and Characterizing Fatty Acid Elongase Component Enzymes. PLoS ONE 2019, 14, e0213620. [Google Scholar] [CrossRef] [Green Version]
- Riederer, M.; Muller, C. Annual Plant Reviews, Biology of the Plant Cuticle; John Wiley & Sons: Hoboken, NJ, USA, 2008; Volume 23, ISBN 1-4051-7157-X. [Google Scholar]
- Pilu, R.; Panzeri, D.; Gavazzi, G.; Rasmussen, S.K.; Consonni, G.; Nielsen, E. Phenotypic, Genetic and Molecular Characterization of a Maize Low Phytic Acid Mutant (Lpa241). Theor. Appl. Genet. 2003, 107, 980–987. [Google Scholar] [CrossRef]
- Nadiminti, P.P.; Rookes, J.E.; Boyd, B.J.; Cahill, D.M. Confocal Laser Scanning Microscopy Elucidation of the Micromorphology of the Leaf Cuticle and Analysis of Its Chemical Composition. Protoplasma 2015, 252, 1475–1486. [Google Scholar] [CrossRef]
- Tanaka, T.; Tanaka, H.; Machida, C.; Watanabe, M.; Machida, Y. A New Method for Rapid Visualization of Defects in Leaf Cuticle Reveals Five Intrinsic Patterns of Surface Defects in Arabidopsis. Plant J. 2004, 37, 139–146. [Google Scholar] [CrossRef]
- Beckett, J.B. B-A Translocations in Maize: I. Use in Locating Genes by Chromosome Arms. J. Hered. 1978, 69, 27–36. [Google Scholar] [CrossRef]
- Vejlupkova, Z.; Fowler, J.E. Maize DNA Preps for Undergraduate Students: A Robust Method for PCR Genotyping. Maize Genet. Coop. Newsl. 2003, 77, 24–25. [Google Scholar]
- Lander, E.S.; Green, P.; Abrahamson, J.; Barlow, A.; Daly, M.J.; Lincoln, S.E.; Newberg, L.A.; Newburg, L. MAPMAKER: An Interactive Computer Package for Constructing Primary Genetic Linkage Maps of Experimental and Natural Populations. Genomics 1987, 1, 174–181. [Google Scholar] [CrossRef]
- Woodhouse, M.R.; Cannon, E.K.; Portwood, J.L.; Harper, L.C.; Gardiner, J.M.; Schaeffer, M.L.; Andorf, C.M. A Pan-Genomic Approach to Genome Databases Using Maize as a Model System. BMC Plant Biol. 2021, 21, 385. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutants | WT | m | Total | % of Mutants | X2 Value for 3:1 | Sig. | P |
|---|---|---|---|---|---|---|---|
| des*-4 | 440 | 147 | 587 | 25.0 | 0.0006 | NS | 0.980 |
| stocky1 | 424 | 117 | 541 | 21.6 | 3.2834 | NS | 0.700 |
| des*-47 | 254 | 71 | 325 | 21.8 | 1.7241 | NS | 0.189 |
| des*-85 | 86 | 24 | 110 | 21.8 | 0.5939 | NS | 0.441 |
| des*-217 | 104 | 38 | 142 | 26.8 | 0.2347 | NS | 0.6281 |
| des*-256 | 114 | 38 | 152 | 25.0 | 0.0000 | NS | 1 |
| ↓ | → | des*-4 | stocky1 | des*-47 | des*-85 | des*-217 | des*-256 |
|---|---|---|---|---|---|---|---|
| des*-4 | - | + | + | + | + | + | |
| stocky1 | - | + | + | + | + | ||
| des*-47 | - | + | + | + | |||
| des*-85 | - | + | + | ||||
| des*-217 | - | + | |||||
| des*-256 | - | ||||||
| TB-A Employed in the Crosses with Stocky1 | Translocation Uncovering the Mutant |
|---|---|
| TB 1Sb | + |
| TB 1La | + |
| TB 3La-2S | + |
| TB 3Sb | + |
| TB 3La | + |
| TB 5Sc | - |
| TB 5La | + |
| TB 7Sc | + |
| TB 7Lb | + |
| TB 8Lc | + |
| TB 9Sb | + |
| TB 10L19 | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaiti, A.; Sangiorgio, S.; Faoro, F.; Pozzi, C.M.; Gavazzi, G.; Pilu, S.R. Stocky1, a Novel Gene Involved in Maize Seedling Development and Cuticle Integrity. Plants 2022, 11, 847. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11070847
Gaiti A, Sangiorgio S, Faoro F, Pozzi CM, Gavazzi G, Pilu SR. Stocky1, a Novel Gene Involved in Maize Seedling Development and Cuticle Integrity. Plants. 2022; 11(7):847. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11070847
Chicago/Turabian StyleGaiti, Angelo, Stefano Sangiorgio, Franco Faoro, Carlo Massimo Pozzi, Giuseppe Gavazzi, and Salvatore Roberto Pilu. 2022. "Stocky1, a Novel Gene Involved in Maize Seedling Development and Cuticle Integrity" Plants 11, no. 7: 847. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11070847