
Uncertainty, Complexity and Constraints: How Do We Robustly Assess Biological Responses under a Rapidly Changing Climate?

 ,
,
Abstract
:1. Introduction
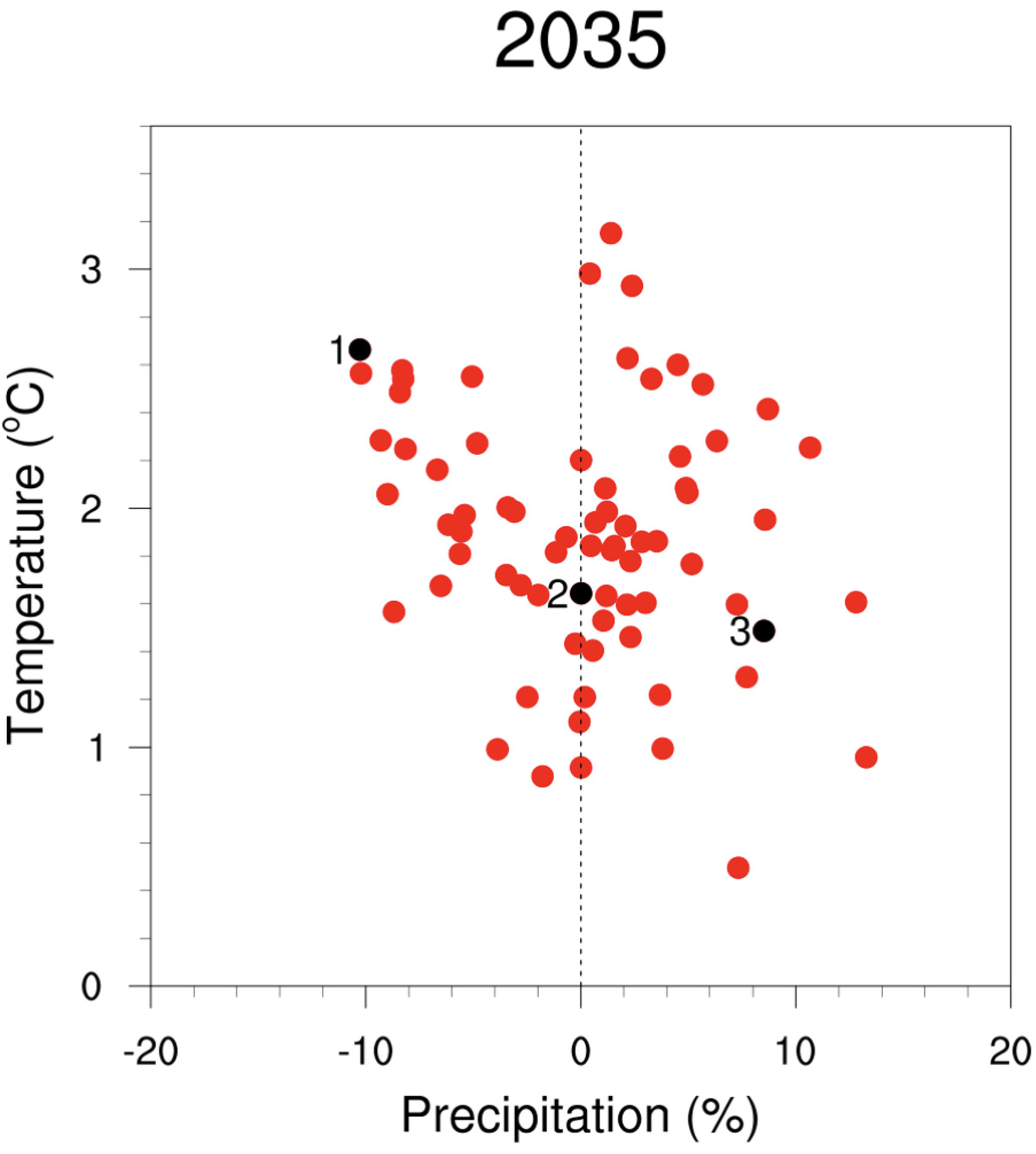
2. Uncertainty Associated with Climate Projections
2.1. Uncertainty from Greenhouse Gas Emissions Scenarios
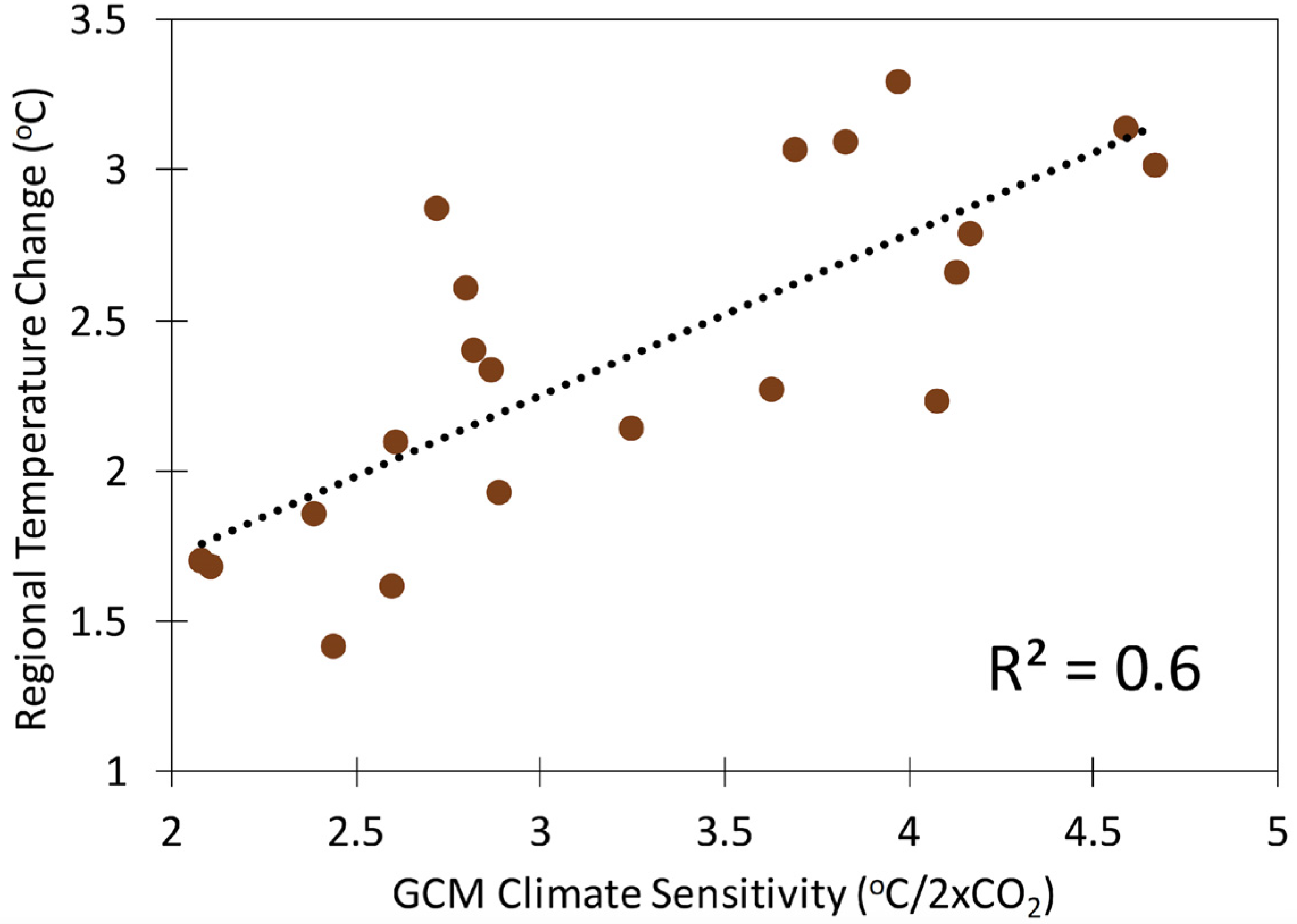
2.2. Uncertainty from Different Climate Models
2.3. Uncertainty from Natural Climate Variability
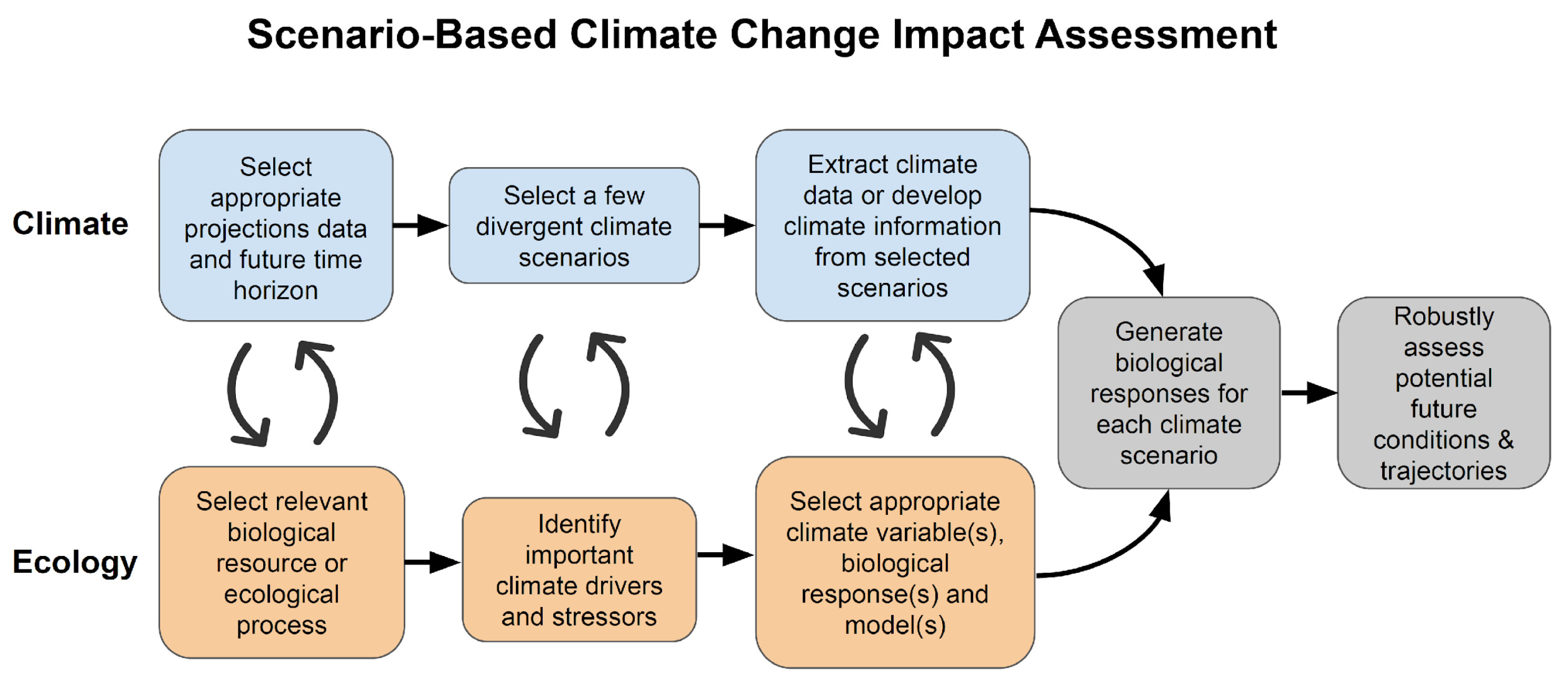
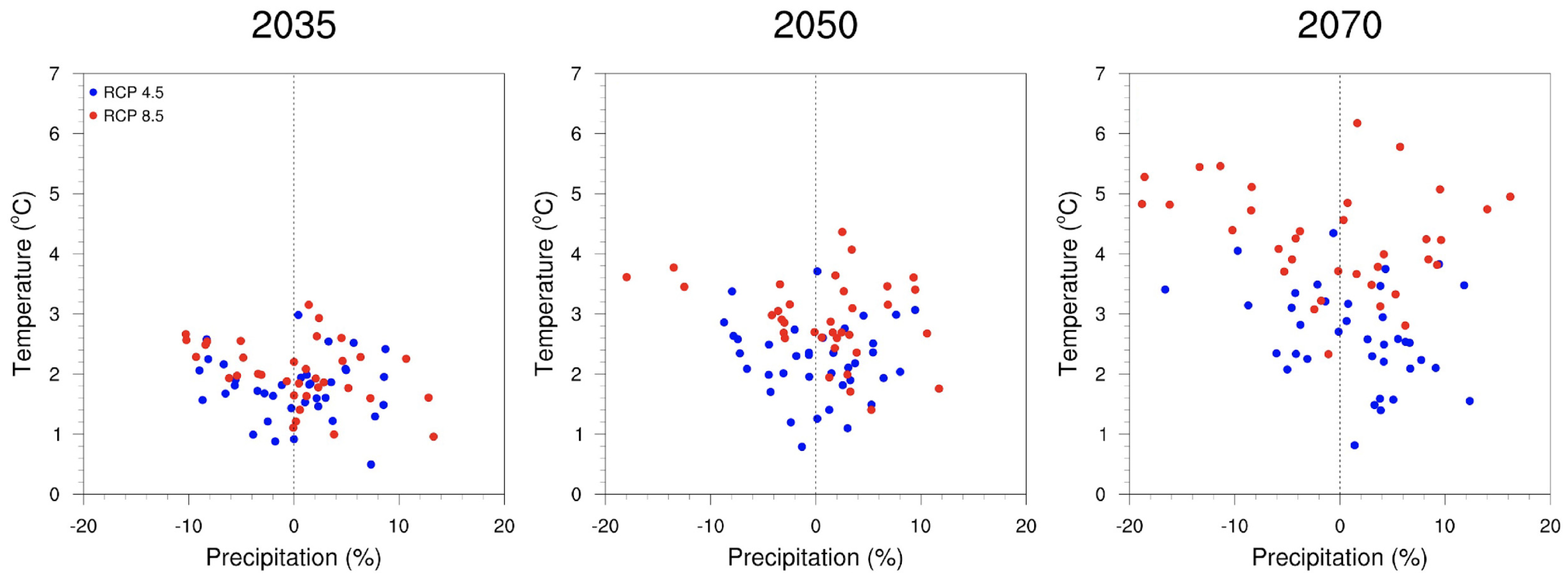
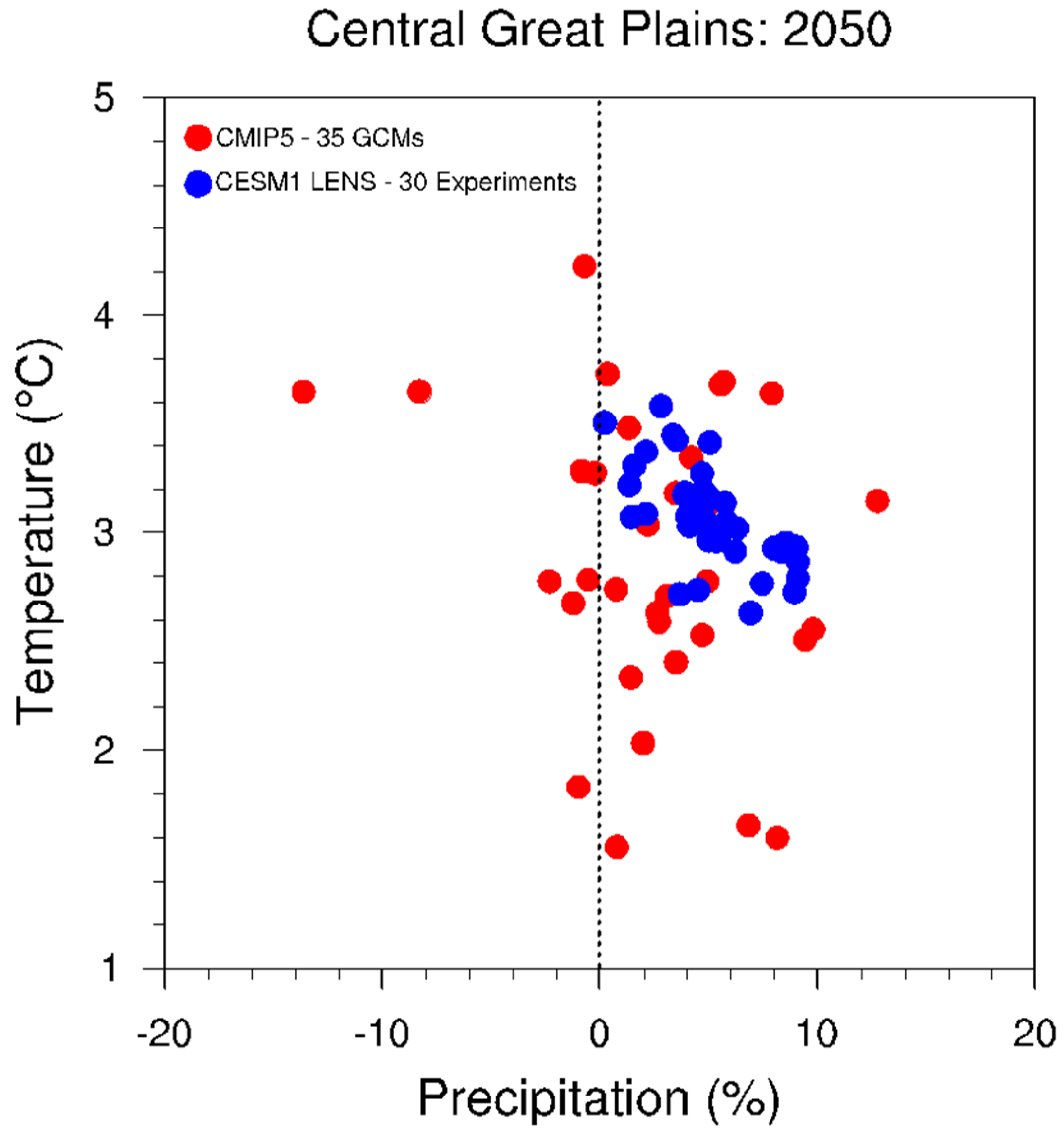
2.4. Approaches That Work across Climate Projections-Related Uncertainties
3. Constraints Associated with Data Availability at Relevant Spatiotemporal Scales
4. Assessment of Relationships between Climate and Biological Processes
5. Relevance of Extreme Climate Events in Driving Biological Impacts
6. Building Robustness into Our Understanding of Future Biological Responses
6.1. Choosing Ecological Response Variables and Prediction Methods
6.2. Linking Climate Variables to Ecological Responses
6.3. Challenges and Limitations of Modeling Biological Responses to Climate
7. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| Southwest Colorado Social-Ecological Climate Resilience (SECR) Project: Pinyon-Juniper Landscape The Southwest Colorado Social-Ecological Climate Resilience (SECR) Project utilized an interdisciplinary team of land managers, ecological and climate scientists, ranchers, and conservation planners to develop an innovative climate planning framework for four targeted landscapes. The Pinyon-Juniper landscape in the San Juan Basin described below was one of four landscapes evaluated throughout the 3-year duration of the SECR Project [17]. One of the management targets was the Pinyon-Juniper landscape. This 376,358-hectare landscape occurs on warm and dry foothills, mesas, canyons, and plateaus and consists of a mosaic of vegetation types; dominant tree species include pinyon pine (Pinus edulis), Utah juniper (Juniperus osteosperma), and Rocky Mountain juniper (J. scopulorum). Elevations are generally between 1,600–2,300 meters. The bi-modal precipitation pattern is important for cone crop production; winter precipitation primarily replenishes deep soil moisture, while summer precipitation provides shallow rooted plants with additional growing season moisture. Any change to the historic precipitation pattern is likely to influence mortality and recruitment of the dominant trees. These evergreen woodlands withstand very cold winter nights and hot summer days. The landscape is exceedingly rich with historic cultural resources, primarily from the ancestral Puebloans (500–1200 AD), as well as biological and land-use diversity. The assessment of ecological responses was evaluated using three climate scenarios: Hot and Dry, Feast or Famine and Warm and Wet to describe the potential future mid-century (2035) scenarios (described in Table S1 in Supplementary Materials). Important climate metrics were compared across all three scenarios. Droughts similar to the 2002 severe drought year were projected to become more frequent (once every 5-years) in the Hot and Dry scenario compared to no change in drought frequency in the Warm and Wet scenario. The wildfire season was projected to increase in duration by 1- month per year in the Hot and Dry scenario, whereas, only a slight increase in the fire season was projected for the Warm and Wet scenario. The fire season was also projected to lengthen in the Feast or Famine scenario. Both qualitative and quantitative ecological response models were developed. The important drivers were determined to be precipitation, temperature and drought. Winter moisture is critical for replenishing deep soil moisture and can help mitigate hot and dry summers. Summer monsoon moisture is required for pinyon pine cone production. Drought impacts are amplified by insects; Ips spp. bark beetles generally increase with drought and large old trees are more susceptible, leading to large die-off of mature trees. Fires are also more likely to increase in severity during severe droughts. Crown fires that kill the dominant pinyon and juniper trees are also more prevalent during drought years. For each climate scenario we delineated areas that were likely to be lost, threatened, persistent, and emergent for pinyon and juniper. Random Forest models were used to depict the change for each future scenario. Expert elicitation was used to evaluate the plausible socio-ecological impacts from the qualitative and quantitative response models. The process and plausible socio-ecological impacts are summarized below.  |
Appendix B
| Species Status Assessment for the Southern White-tailed Ptarmigan (Ptarmigan SSA) The U.S. Fish and Wildlife Service (USFWS) uses a species status assessment (SSA) framework to inform listing decisions under the Endangered Species Act (ESA; [165]). The SSA gathers available information on a species’ biology and ecological needs, assesses current conditions of the species’ habitats and populations, and forecasts future conditions for the species under a range of plausible scenarios. An SSA for the southern white-tailed ptarmigan (Lagopus leucura altipetens) was initiated in 2018 and involved an interdisciplinary collaboration of species experts, climate scientists, conservation geneticists, and population modelers. In 2020, the USFWS completed the SSA for the southern white-tailed ptarmigan [55], which was used to inform the determination that the subspecies was not warranted for listing. The management target was the viability of the southern white-tailed ptarmigan. The subspecies inhabits alpine ecosystems at or above treeline year-round; in Colorado they occupy alpine habitats between 3353–4114 meters [166]. A conceptual model of habitat and demographic factors required by southern white-tailed ptarmigan to maintain population resiliency and support species viability was developed that included: (a) winter snow conditions that support well-distributed areas of soft snow suitable for snow roosting; (b) late-lying snowfields that help maintain brood-rearing habitats and provide thermal refugia; (c) summer precipitation/monsoonal moisture that provide regular cooling and late-season moisture; (d) brood-rearing habitat that provides young chicks with invertebrates and adults with chicks desirable forage forbs; and (e) abundant willow needed by all life stages that influences the distribution of ptarmigan. Ptarmigan populations are maintained through immigration [167,168,169], emphasizing the importance of demographic connectivity among suitable alpine habitats. The assessment of biological responses was evaluated using three climate scenarios: Very Hot and Dry, Hot, and Hot and Very Wet to describe the plausible future scenarios (described in Table S2 in Supplementary Materials) out to mid-century (2050). Several climate metrics important to ptarmigan habitat were considered under these scenarios, including frequency of severe drought, spring snowpack, seasonal temperatures, frequency of intense rainfall events, and monsoonal precipitation. This SSA considered both qualitative and quantitative ecological response models. Expert input was used to evaluate the impacts of future climate scenarios on ptarmigan habitat and demography, specifically in terms of projections for temperature and precipitation. Dynamic global vegetation models (DGVMs) were also considered as a tool to inform the evaluation of future conditions for the southern white-tailed ptarmigan. However, the results of the DGVM that were evaluated conflicted with the understanding of the potential future habitat conditions for the southern white-tailed ptarmigan. Instead, the future condition of the southern white-tailed ptarmigan was evaluated under the three climate scenarios using several decades of demographic and ecological data in combination with expert input. The SSA process for the southern white-tailed ptarmigan and plausible impacts to habitat and demographic factors are summarized in the figure below.  |
References
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Nolan, C.; Overpeck, J.T.; Allen, J.R.M.; Anderson, P.M.; Betancourt, J.L.; Binney, H.A.; Brewer, S.; Bush, M.B.; Chase, B.M.; Cheddadi, R. Past and future global transformation of terrestrial ecosystems under climate change. Science 2018, 361, 920–923. [Google Scholar] [CrossRef] [Green Version]
- Jackson, S.T. Transformational ecology and climate change. Science 2021, 373, 1085–1086. [Google Scholar] [CrossRef]
- Millar, C.I.; Stephenson, N.L. Temperate forest health in an era of emerging megadisturbance. Science 2015, 349, 823–826. [Google Scholar] [CrossRef]
- Díaz, S.; Settele, J.; Brondízio, E.S.; Ngo, H.T.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.A.; Butchart, S.H.M.; Chan, K.M.A. Pervasive human-driven decline of life on Earth points to the need for transformative change. Science 2019, 366, eaax3100. [Google Scholar] [CrossRef] [Green Version]
- Bridle, J.; van Rensburg, A. Discovering the limits of ecological resilience. Science 2020, 367, 626–627. [Google Scholar] [CrossRef] [PubMed]
- Crausbay, S.D.; Betancourt, J.; Bradford, J.; Cartwright, J.; Dennison, W.C.; Dunham, J.; Enquist, C.A.F.; Frazier, A.G.; Hall, K.R.; Littell, J.S. Unfamiliar territory: Emerging themes for ecological drought research and management. One Earth 2020, 3, 337–353. [Google Scholar] [CrossRef]
- Heino, J.; Alahuhta, J.; Bini, L.M.; Cai, Y.; Heiskanen, A.; Hellsten, S.; Kortelainen, P.; Kotamäki, N.; Tolonen, K.T.; Vihervaara, P. Lakes in the era of global change: Moving beyond single-lake thinking in maintaining biodiversity and ecosystem services. Biol. Rev. 2021, 96, 89–106. [Google Scholar] [CrossRef]
- Hughes, T.P.; Anderson, K.D.; Connolly, S.R.; Heron, S.F.; Kerry, J.T.; Lough, J.M.; Baird, A.H.; Baum, J.K.; Berumen, M.L.; Bridge, T.C. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 2018, 359, 80–83. [Google Scholar] [CrossRef] [Green Version]
- Sully, S.; Burkepile, D.E.; Donovan, M.K.; Hodgson, G.; Van Woesik, R. A global analysis of coral bleaching over the past two decades. Nat. Commun. 2019, 10, 1264. [Google Scholar] [CrossRef] [Green Version]
- Grosse, G.; Goetz, S.; McGuire, A.D.; Romanovsky, V.E.; Schuur, E.A.G. Changing permafrost in a warming world and feedbacks to the Earth system. Environ. Res. Lett. 2016, 11, 40201. [Google Scholar] [CrossRef]
- Berner, L.T.; Massey, R.; Jantz, P.; Forbes, B.C.; Macias-Fauria, M.; Myers-Smith, I.; Kumpula, T.; Gauthier, G.; Andreu-Hayles, L.; Gaglioti, B.V. Summer warming explains widespread but not uniform greening in the Arctic tundra biome. Nat. Commun. 2020, 11, 4621. [Google Scholar] [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobre, C.A.; Sampaio, G.; Borma, L.S.; Castilla-Rubio, J.C.; Silva, J.S.; Cardoso, M. Land-use and climate change risks in the Amazon and the need of a novel sustainable development paradigm. Proc. Natl. Acad. Sci. USA 2016, 113, 10759–10768. [Google Scholar] [CrossRef] [Green Version]
- Macias-Fauria, M.; Post, E. Effects of sea ice on Arctic biota: An emerging crisis discipline. Biol. Lett. 2018, 14, 20170702. [Google Scholar] [CrossRef] [Green Version]
- Flynn, M.; Ford, J.D.; Pearce, T.; Harper, S.L.; Team, I.R. Participatory scenario planning and climate change impacts, adaptation and vulnerability research in the Arctic. Environ. Sci. Policy 2018, 79, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Rondeau, R.; Bidwell, M.; Neely, B.; Rangwala, I.; Yung, L.; Wyborn, C. Pinyon-juniper landscape: San Juan Basin, Colorado Social-Ecological Climate Resilience Project Report. North Central Climate Science Center: Ft. Collins, CO, USA, 2017. Available online: https://www.sciencebase.gov/catalog/item/5d10f7e4e4b0941bde5502c4 (accessed on 10 September 2021).
- Symstad, A.J.; Fisichelli, N.A.; Miller, B.W.; Rowland, E.; Schuurman, G.W. Multiple methods for multiple futures: Integrating qualitative scenario planning and quantitative simulation modeling for natural resource decision making. Clim. Risk Manag. 2017, 17, 78–91. [Google Scholar] [CrossRef]
- Thorne, J.H.; Gogol-Prokurat, M.; Hill, S.; Walsh, D.; Boynton, R.M.; Choe, H. Vegetation refugia can inform climate-adaptive land management under global warming. Front. Ecol. Environ. 2020, 18, 281–287. [Google Scholar] [CrossRef]
- Raffini, F.; Bertorelle, G.; Biello, R.; D’Urso, G.; Russo, D.; Bosso, L. From nucleotides to satellite imagery: Approaches to identify and manage the invasive pathogen Xylella fastidiosa and its insect vectors in Europe. Sustainability 2020, 12, 4508. [Google Scholar] [CrossRef]
- Peterson, G.D.; Cumming, G.S.; Carpenter, S.R. Scenario planning: A tool for conservation in an uncertain world. Conserv. Biol. 2003, 17, 358–366. [Google Scholar] [CrossRef] [Green Version]
- Rowland, E.R.; Cross, M.S.; Hartmann, H. Considering Multiple Futures: Scenario Planning to Address Uncertainty in Natural Resource Conservation; Fish and Wildlife Service: Washington, DC, USA, 2014. [Google Scholar]
- Star, J.; Rowland, E.L.; Black, M.E.; Enquist, C.A.F.; Garfin, G.; Hoffman, C.H.; Hartmann, H.; Jacobs, K.L.; Moss, R.H.; Waple, A.M. Supporting adaptation decisions through scenario planning: Enabling the effective use of multiple methods. Clim. Risk Manag. 2016, 13, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Ogden, A.E.; Innes, J.L. Application of structured decision making to an assessment of climate change vulnerabilities and adaptation options for sustainable forest management. Ecol. Soc. 2009, 14, 11. [Google Scholar] [CrossRef]
- Stein, B.A.; Glick, P.; Edelson, N.; Staudt, A. Climate-Smart Conservation: Putting Adaptation Principles into Practice [Online]; National Wildlife Federation: Washington, DC, USA, 2014. [Google Scholar]
- Cross, M.S.; Zavaleta, E.S.; Bachelet, D.; Brooks, M.L.; Enquist, C.A.F.; Fleishman, E.; Graumlich, L.J.; Groves, C.R.; Hannah, L.; Hansen, L. The Adaptation for Conservation Targets (ACT) framework: A tool for incorporating climate change into natural resource management. Environ. Manag. 2012, 50, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Schuurman, G.W.; Hoffman, C.H.; Cole, D.N.; Lawrence, D.J.; Morton, J.M.; Magness, D.R.; Cravens, A.E.; Covington, S.; O’Malley, R.; Fisichelli, N.A. Resist-Accept-Direct (RAD)—A Framework for the 21st-Century Natural Resource Manager; Report no. NPS/NRSS/CCRP/NRR—2020/2213; National Park Service: Washington, DC, USA, 2020. [Google Scholar]
- Hausfather, Z.; Peters, G.P. Emissions—The ‘business as usual’story is misleading 2020. Nature 2020, 577, 618–620. [Google Scholar] [CrossRef]
- Burgess, M.G.; Ritchie, J.; Shapland, J.; Pielke, R. IPCC baseline scenarios have over-projected CO2 emissions and economic growth. Environ. Res. Lett. 2020, 16, 14016. [Google Scholar] [CrossRef]
- Held, H.; Aykut, S.; Hedemann, C.; Li, C.; Marotzke, J.; Petzold, J.; Schneider, U. Plausibility of model-based emissions scenarios. In Hamburg Climate Futures Outlook 2021: Assessing the Plausibility of Deep Decarbonization by 2050; Cluster of Excellence Climate, Climatic Change, and Society (CLICCS): Hamburg, Germany, 2021; pp. 21–26. [Google Scholar]
- Fowler, H.J.; Blenkinsop, S.; Tebaldi, C. Linking climate change modelling to impacts studies: Recent advances in downscaling techniques for hydrological modelling. Int. J. Climatol. A J. R. Meteorol. Soc. 2007, 27, 1547–1578. [Google Scholar] [CrossRef]
- Daniels, A.E.; Morrison, J.F.; Joyce, L.A.; Crookston, N.L.; Chen, S.-C.; McNulty, S.G. Climate Projections FAQ. In Gen. Tech. Rep. RMRS-GTR-277WWW; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2012; Volume 277, 32 p. [Google Scholar]
- Kotamarthi, R.; Mearns, L.; Hayhoe, K.; Castro, C.L.; Wuebbles, D. Use of Climate Information for Decision-Making and Impacts Research: State of Our Understanding; Argonne National Laboratory: Lemont, IL, USA, 2016. [Google Scholar]
- Chakraborty, D.; Dobor, L.; Zolles, A.; Hlásny, T.; Schueler, S. High-resolution gridded climate data for Europe based on bias-corrected EURO-CORDEX: The ECLIPS dataset. Geosci. Data J. 2020, 8, 121–131. [Google Scholar] [CrossRef]
- Platts, P.J.; Omeny, P.; Marchant, R. AFRICLIM: High-resolution climate projections for ecological applications in Africa. Afr. J. Ecol. 2015, 53, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Lukas, J.; Barsugli, J.; Doesken, N.; Rangwala, I.; Wolter, K. Climate change in Colorado: A synthesis to support water resources management and adaptation. Univ. Color. Boulder Color. 2014. [Google Scholar] [CrossRef]
- Roe, G.H.; Baker, M.B. Why is climate sensitivity so unpredictable? Science 2007, 318, 629–632. [Google Scholar] [CrossRef] [Green Version]
- Knutti, R. Should we believe model predictions of future climate change? Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2008, 366, 4647–4664. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, E.; Sutton, R. The potential to narrow uncertainty in regional climate predictions. Bull. Am. Meteorol. Soc. 2009, 90, 1095–1108. [Google Scholar] [CrossRef] [Green Version]
- Andrews, T.; Gregory, J.M.; Webb, M.J.; Taylor, K.E. Forcing, feedbacks and climate sensitivity in CMIP5 coupled atmosphere-ocean climate models. Geophys. Res. Lett. 2012, 39, L09712. [Google Scholar] [CrossRef] [Green Version]
- Vial, J.; Dufresne, J.-L.; Bony, S. On the interpretation of inter-model spread in CMIP5 climate sensitivity estimates. Clim. Dyn. 2013, 41, 3339–3362. [Google Scholar] [CrossRef]
- Ulbrich, U.; Leckebusch, G.C.; Pinto, J.G. Extra-tropical cyclones in the present and future climate: A review. Theor. Appl. Climatol. 2009, 96, 117–131. [Google Scholar] [CrossRef] [Green Version]
- Rangwala, I.; Pepin, N.; Vuille, M.; Miller, J. Influence of climate variability and large-scale circulation on the mountain cryosphere. In The High-Mountain Cryosphere: Environmental Changes and Human Risks; Cambridge University Press: Cambridge, UK, 2015; pp. 9–27. [Google Scholar]
- Kirtman, B.; Power, S.B.; Adedoyin, A.J.; Boer, G.J.; Bojariu, R.; Camilloni, I.; Doblas-Reyes, F.; Fiore, A.M.; Kimoto, M.; Meehl, G. Near-Term Climate Change: Projections and Predictability. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Doblas-Reyes, F.J.; Sorensson, A.A.; Almazroui, M.; Dosio, A.; Gutowski, W.J.; Haarsma, R.; Hamdi, R.; Hewitson, B.; Kwon, W.-T.; Lamptey, B.L. Linking Global to Regional Climate Change. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Lempert, R. Scenarios that illuminate vulnerabilities and robust responses. Clim. Chang. 2013, 117, 627–646. [Google Scholar] [CrossRef]
- Shepherd, T.G. Storyline approach to the construction of regional climate change information. Proc. R. Soc. A 2019, 475, 20190013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joyce, L.A.; Coulson, D. Climate scenarios and projections: A Technical Document Supporting the USDA Forest Service 2020 RPA Assessment. In Gen. Tech. Rep. RMRS-GTR-413; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2020; Volume 413, 85p. [Google Scholar] [CrossRef]
- Lawrence, D.J.; Runyon, A.N.; Gross, J.E.; Schuurman, G.W.; Miller, B.W. Divergent, plausible, and relevant climate futures for near-and long-term resource planning. Clim. Chang. 2021, 167, 38. [Google Scholar] [CrossRef]
- Runyon, A.N.; Schuurman, G.W.; Miller, B.W.; Symstad, A.; Hardy, A. Climate Change Scenario Planning for Resource Stewardship at Wind Cave National Park; National Park Service: Ft. Collins, CO, USA, 2021. [Google Scholar]
- Weigel, A.P.; Knutti, R.; Liniger, M.A.; Appenzeller, C. Risks of model weighting in multimodel climate projections. J. Clim. 2010, 23, 4175–4191. [Google Scholar] [CrossRef]
- Mote, P.; Brekke, L.; Duffy, P.B.; Maurer, E. Guidelines for constructing climate scenarios. Eos. Trans. Am. Geophys. Union 2011, 92, 257–258. [Google Scholar] [CrossRef]
- Rupp, D.E.; Abatzoglou, J.T.; Hegewisch, K.C.; Mote, P.W. Evaluation of CMIP5 20th century climate simulations for the Pacific Northwest USA. J. Geophys. Res. Atmos. 2013, 118, 10–884. [Google Scholar] [CrossRef]
- Rupp, D.E. An Evaluation of 20th Century Climate for the Southeastern United States as Simulated by Coupled Model Intercomparison Project Phase 5 (CMIP5) Global Climate Models; U.S. Geological Survey Open-File Report 2016-1047; USGS: Raleigh, NC, USA, 2016; 32 p. [CrossRef] [Green Version]
- USFWS. Species Status Assessment Report for the Southern White-Tailed Ptarmigan (Lagopus leucura altipetens); US Fish and Wildlife Service: Lakewood, CO, USA, 2020. [Google Scholar]
- USFWS. Species Status Assessment Report for Colorado Hookless Cactus (Sclerocactus glaucus and Sclerocactus dawsonii); US Fish and Wildlife Service: Lakewood, CO, USA, 2021. [Google Scholar]
- Runyon, A.N.; Carlson, A.R.; Gross, J.; Lawrence, D.J.; Schuurman, G.W. Repeatable Approaches to Work with Scientific Uncertainty and Advance Climate Change Adaptation in US National Parks. Proc. Parks Steward. Forum 2020, 36. [Google Scholar] [CrossRef] [Green Version]
- Barsugli, J.J.; Guentchev, G.; Horton, R.M.; Wood, A.; Mearns, L.O.; Liang, X.; Winkler, J.A.; Dixon, K.; Hayhoe, K.; Rood, R.B. The practitioner’s dilemma: How to assess the credibility of downscaled climate projections. Eos. Trans. Am. Geophys. Union 2013, 94, 424–425. [Google Scholar] [CrossRef] [Green Version]
- Jennings, K.S.; Winchell, T.S.; Livneh, B.; Molotch, N.P. Spatial variation of the rain–snow temperature threshold across the Northern Hemisphere. Nat. Commun. 2018, 9, 1148. [Google Scholar] [CrossRef] [Green Version]
- Barsugli, J.J.; Ray, A.J.; Livneh, B.; Dewes, C.F.; Heldmyer, A.; Rangwala, I.; Guinotte, J.M.; Torbit, S. Projections of mountain snowpack loss for wolverine denning elevations in the Rocky Mountains. Earth’s Future 2020, 8, e2020EF001537. [Google Scholar] [CrossRef]
- Easterling, D.R.; Kunkel, K.E.; Arnold, J.R.; Knutson, T.; LeGrande, A.N.; Leung, L.R.; Vose, R.S.; Waliser, D.E.; Wehner, M.F. Precipitation Change in the United States BT-Climate Science Special Report: Fourth National Climate Assessment; Wuebbles, D.J., Fahey, D.W., Hibbard, K.A., Dokken, D.J., Stewart, B.C., Maycock, T.K., Eds.; U.S. Global Change Research Program: Washington, DC, USA, 2017; pp. 207–230.
- Kendon, E.J.; Roberts, N.M.; Senior, C.A.; Roberts, M.J. Realism of rainfall in a very high-resolution regional climate model. J. Clim. 2012, 25, 5791–5806. [Google Scholar] [CrossRef]
- Chan, S.C.; Kendon, E.J.; Roberts, N.M.; Fowler, H.J.; Blenkinsop, S. The characteristics of summer sub-hourly rainfall over the southern UK in a high-resolution convective permitting model. Environ. Res. Lett. 2016, 11, 94024. [Google Scholar] [CrossRef]
- Liu, C.; Ikeda, K.; Rasmussen, R.; Barlage, M.; Newman, A.J.; Prein, A.F.; Chen, F.; Chen, L.; Clark, M.; Dai, A. Continental-scale convection-permitting modeling of the current and future climate of North America. Clim. Dyn. 2017, 49, 71–95. [Google Scholar] [CrossRef]
- Fowler, H.J.; Lenderink, G.; Prein, A.F.; Westra, S.; Allan, R.P.; Ban, N.; Barbero, R.; Berg, P.; Blenkinsop, S.; Do, H.X. Anthropogenic intensification of short-duration rainfall extremes. Nat. Rev. Earth Environ. 2021, 2, 107–122. [Google Scholar] [CrossRef]
- Redmond, K.T. The depiction of drought: A commentary. Bull. Am. Meteorol. Soc. 2002, 83, 1143–1147. [Google Scholar] [CrossRef] [Green Version]
- Hobbins, M.; Rangwala, I.; Barsugli, J.; Dewes, C. Extremes in evaporative demand and their implications for droughts and drought monitoring in the 21st century. In Extreme Hydrology and Climate Variability; Elsevier: Amsterdam, The Netherlands, 2019; pp. 325–341. [Google Scholar]
- Ficklin, D.L.; Novick, K.A. Historic and projected changes in vapor pressure deficit suggest a continental-scale drying of the United States atmosphere. J. Geophys. Res. Atmos. 2017, 122, 2061–2079. [Google Scholar] [CrossRef]
- Williams, A.P.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang. 2013, 3, 292–297. [Google Scholar] [CrossRef]
- Urban, M.C.; Bocedi, G.; Hendry, A.P.; Mihoub, J.-B.; Pe’er, G.; Singer, A.; Bridle, J.R.; Crozier, L.G.; De Meester, L.; Godsoe, W. Improving the forecast for biodiversity under climate change. Science 2016, 353, aad8466. [Google Scholar] [CrossRef] [Green Version]
- Tewksbury, J.J.; Anderson, J.G.T.; Bakker, J.D.; Billo, T.J.; Dunwiddie, P.W.; Groom, M.J.; Hampton, S.E.; Herman, S.G.; Levey, D.J.; Machnicki, N.J. Natural history’s place in science and society. Bioscience 2014, 64, 300–310. [Google Scholar] [CrossRef] [Green Version]
- Dawson, T.P.; Jackson, S.T.; House, J.I.; Prentice, I.C.; Mace, G.M. Beyond predictions: Biodiversity conservation in a changing climate. Science 2011, 332, 53–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepetz, V.; Massot, M.; Schmeller, D.S.; Clobert, J. Biodiversity monitoring: Some proposals to adequately study species’ responses to climate change. Biodivers. Conserv. 2009, 18, 3185–3203. [Google Scholar] [CrossRef]
- Pacifici, M.; Foden, W.B.; Visconti, P.; Watson, J.E.M.; Butchart, S.H.M.; Kovacs, K.M.; Scheffers, B.R.; Hole, D.G.; Martin, T.G.; Akçakaya, H.R. Assessing species vulnerability to climate change. Nat. Clim. Chang. 2015, 5, 215–224. [Google Scholar] [CrossRef]
- Buckley, L.B. Linking traits to energetics and population dynamics to predict lizard ranges in changing environments. Am. Nat. 2008, 171, E1–E19. [Google Scholar] [CrossRef] [Green Version]
- Coops, N.C.; Waring, R.H.; Schroeder, T.A. Combining a generic process-based productivity model and a statistical classification method to predict the presence and absence of tree species in the Pacific Northwest, USA. Ecol. Modell. 2009, 220, 1787–1796. [Google Scholar] [CrossRef]
- Angilletta, M.J., Jr.; Angilletta, M.J. Thermal Adaptation: A Theoretical and Empirical Synthesis; Oxford University Press: Oxford, UK, 2009. [Google Scholar] [CrossRef]
- Fisichelli, N.; Wright, A.; Rice, K.; Mau, A.; Buschena, C.; Reich, P.B. First-year seedlings and climate change: Species-specific responses of 15 North American tree species. Oikos 2014, 123, 1331–1340. [Google Scholar] [CrossRef]
- Chapin III, F.S.; Shaver, G.R. Physiological and growth responses of arctic plants to a field experiment simulating climatic change. Ecology 1996, 77, 822–840. [Google Scholar] [CrossRef]
- Liu, H.; Mi, Z.; Lin, L.I.; Wang, Y.; Zhang, Z.; Zhang, F.; Wang, H.; Liu, L.; Zhu, B.; Cao, G. Shifting plant species composition in response to climate change stabilizes grassland primary production. Proc. Natl. Acad. Sci. USA 2018, 115, 4051–4056. [Google Scholar] [CrossRef] [Green Version]
- Stewart, R.I.A.; Dossena, M.; Bohan, D.A.; Jeppesen, E.; Kordas, R.L.; Ledger, M.E.; Meerhoff, M.; Moss, B.; Mulder, C.; Shurin, J.B. Mesocosm experiments as a tool for ecological climate-change research. Adv. Ecol. Res. 2013, 48, 71–181. [Google Scholar]
- Henry, G.H.R.; Molau, U. Tundra plants and climate change: The International Tundra Experiment (ITEX). Glob. Chang. Biol. 1997, 3, 1–9. [Google Scholar] [CrossRef]
- Fraser, L.H.; Henry, H.A.L.; Carlyle, C.N.; White, S.R.; Beierkuhnlein, C.; Cahill, J.F., Jr.; Casper, B.B.; Cleland, E.; Collins, S.L.; Dukes, J.S. Coordinated distributed experiments: An emerging tool for testing global hypotheses in ecology and environmental science. Front. Ecol. Environ. 2013, 11, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef] [PubMed]
- Mouquet, N.; Lagadeuc, Y.; Devictor, V.; Doyen, L.; Duputié, A.; Eveillard, D.; Faure, D.; Garnier, E.; Gimenez, O.; Huneman, P. Predictive ecology in a changing world. J. Appl. Ecol. 2015, 52, 1293–1310. [Google Scholar] [CrossRef]
- Wadgymar, S.M.; Ogilvie, J.E.; Inouye, D.W.; Weis, A.E.; Anderson, J.T. Phenological responses to multiple environmental drivers under climate change: Insights from a long-term observational study and a manipulative field experiment. New Phytol. 2018, 218, 517–529. [Google Scholar] [CrossRef] [Green Version]
- Nogués-Bravo, D.; Rodríguez-Sánchez, F.; Orsini, L.; de Boer, E.; Jansson, R.; Morlon, H.; Fordham, D.A.; Jackson, S.T. Cracking the code of biodiversity responses to past climate change. Trends Ecol. Evol. 2018, 33, 765–776. [Google Scholar] [CrossRef] [Green Version]
- Fordham, D.A.; Jackson, S.T.; Brown, S.C.; Huntley, B.; Brook, B.W.; Dahl-Jensen, D.; Gilbert, M.T.P.; Otto-Bliesner, B.L.; Svensson, A.; Theodoridis, S. Using paleo-archives to safeguard biodiversity under climate change. Science 2020, 369, eabc5654. [Google Scholar] [CrossRef]
- Case, M.J.; Lawler, J.J.; Tomasevic, J.A. Relative sensitivity to climate change of species in northwestern North America. Biol. Conserv. 2015, 187, 127–133. [Google Scholar] [CrossRef]
- Martin, T.G.; Burgman, M.A.; Fidler, F.; Kuhnert, P.M.; Low-Choy, S.; McBride, M.; Mengersen, K. Eliciting expert knowledge in conservation science. Conserv. Biol. 2012, 26, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Nabhan, G.P. Perspectives in Ethnobiology: Ethnophenology and climate change. J. Ethnobiol. 2010, 30, 1–4. [Google Scholar] [CrossRef]
- Vinyeta, K.; Lynn, K. Exploring the role of traditional ecological knowledge in climate change initiatives. In Gen. Tech. Rep. PNW-GTR-879; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2013; Volume 879, 37 p. [Google Scholar]
- Smith, S.D.P.; Bunnell, D.B.; Burton, G.A., Jr.; Ciborowski, J.J.H.; Davidson, A.D.; Dickinson, C.E.; Eaton, L.A.; Esselman, P.C.; Evans, M.A.; Kashian, D.R. Evidence for interactions among environmental stressors in the Laurentian Great Lakes. Ecol. Indic. 2019, 101, 203–211. [Google Scholar] [CrossRef]
- Krivtsov, V. Investigations of indirect relationships in ecology and environmental sciences: A review and the implications for comparative theoretical ecosystem analysis. Ecol. Modell. 2004, 174, 37–54. [Google Scholar] [CrossRef]
- Tylianakis, J.M.; Didham, R.K.; Bascompte, J.; Wardle, D.A. Global change and species interactions in terrestrial ecosystems. Ecol. Lett. 2008, 11, 1351–1363. [Google Scholar] [CrossRef]
- Suttle, K.B.; Thomsen, M.A.; Power, M.E. Species interactions reverse grassland responses to changing climate. Science 2007, 315, 640–642. [Google Scholar] [CrossRef] [Green Version]
- Santini, L.; Cornulier, T.; Bullock, J.M.; Palmer, S.C.F.; White, S.M.; Hodgson, J.A.; Bocedi, G.; Travis, J.M.J. A trait-based approach for predicting species responses to environmental change from sparse data: How well might terrestrial mammals track climate change? Glob. Chang. Biol. 2016, 22, 2415–2424. [Google Scholar] [CrossRef] [Green Version]
- Bozinovic, F.; Pörtner, H. Physiological ecology meets climate change. Ecol. Evol. 2015, 5, 1025–1030. [Google Scholar] [CrossRef] [PubMed]
- Knapp, A.K.; Hoover, D.L.; Wilcox, K.R.; Avolio, M.L.; Koerner, S.E.; La Pierre, K.J.; Loik, M.E.; Luo, Y.; Sala, O.E.; Smith, M.D. Characterizing differences in precipitation regimes of extreme wet and dry years: Implications for climate change experiments. Glob. Chang. Biol. 2015, 21, 2624–2633. [Google Scholar] [CrossRef] [Green Version]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef] [Green Version]
- Ummenhofer, C.C.; Meehl, G.A. Extreme weather and climate events with ecological relevance: A review. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160135. [Google Scholar] [CrossRef]
- Garcia, R.A.; Cabeza, M.; Rahbek, C.; Araújo, M.B. Multiple dimensions of climate change and their implications for biodiversity. Science 2014, 344, 1247579. [Google Scholar] [CrossRef] [Green Version]
- Helmuth, B.; Russell, B.D.; Connell, S.D.; Dong, Y.; Harley, C.D.G.; Lima, F.P.; Sará, G.; Williams, G.A.; Mieszkowska, N. Beyond long-term averages: Making biological sense of a rapidly changing world. Clim. Chang. Responses 2014, 1, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Navarro, M.Á.P.; Sapes, G.; Batllori, E.; Serra-Diaz, J.M.; Esteve, M.A.; Lloret, F. Climatic suitability derived from species distribution models captures community responses to an extreme drought episode. Ecosystems 2019, 22, 77–90. [Google Scholar] [CrossRef] [Green Version]
- Germain, S.J.; Lutz, J.A. Climate extremes may be more important than climate means when predicting species range shifts. Clim. Chang. 2020, 163, 579–598. [Google Scholar] [CrossRef]
- Cuena-Lombraña, A.; Fois, M.; Fenu, G.; Cogoni, D.; Bacchetta, G. The impact of climatic variations on the reproductive success of Gentiana lutea L. in a Mediterranean mountain area. Int. J. Biometeorol. 2018, 62, 1283–1295. [Google Scholar] [CrossRef] [PubMed]
- Jentsch, A.; Beierkuhnlein, C. Research frontiers in climate change: Effects of extreme meteorological events on ecosystems. Comptes Rendus Geosci. 2008, 340, 621–628. [Google Scholar] [CrossRef]
- Wethey, D.S.; Woodin, S.A.; Hilbish, T.J.; Jones, S.J.; Lima, F.P.; Brannock, P.M. Response of intertidal populations to climate: Effects of extreme events versus long term change. J. Exp. Mar. Bio. Ecol. 2011, 400, 132–144. [Google Scholar] [CrossRef]
- Harris, R.M.B.; Beaumont, L.J.; Vance, T.R.; Tozer, C.R.; Remenyi, T.A.; Perkins-Kirkpatrick, S.E.; Mitchell, P.J.; Nicotra, A.B.; McGregor, S.; Andrew, N.R. Biological responses to the press and pulse of climate trends and extreme events. Nat. Clim. Chang. 2018, 8, 579–587. [Google Scholar] [CrossRef]
- Maxwell, S.L.; Butt, N.; Maron, M.; McAlpine, C.A.; Chapman, S.; Ullmann, A.; Segan, D.B.; Watson, J.E.M. Conservation implications of ecological responses to extreme weather and climate events. Divers. Distrib. 2019, 25, 613–625. [Google Scholar] [CrossRef]
- Weiskopf, S.R.; Rubenstein, M.A.; Crozier, L.G.; Gaichas, S.; Griffis, R.; Halofsky, J.E.; Hyde, K.J.W.; Morelli, T.L.; Morisette, J.T.; Muñoz, R.C. Climate change effects on biodiversity, ecosystems, ecosystem services, and natural resource management in the United States. Sci. Total Environ. 2020, 733, 137782. [Google Scholar] [CrossRef]
- Turner, M.G.; Calder, W.J.; Cumming, G.S.; Hughes, T.P.; Jentsch, A.; LaDeau, S.L.; Lenton, T.M.; Shuman, B.N.; Turetsky, M.R.; Ratajczak, Z. Climate change, ecosystems and abrupt change: Science priorities. Philos. Trans. R. Soc. B 2020, 375, 20190105. [Google Scholar] [CrossRef] [Green Version]
- Batllori, E.; Lloret, F.; Aakala, T.; Anderegg, W.R.L.; Aynekulu, E.; Bendixsen, D.P.; Bentouati, A.; Bigler, C.; Burk, C.J.; Camarero, J.J. Forest and woodland replacement patterns following drought-related mortality. Proc. Natl. Acad. Sci. USA 2020, 117, 29720–29729. [Google Scholar] [CrossRef] [PubMed]
- Sofaer, H.R.; Barsugli, J.J.; Jarnevich, C.S.; Abatzoglou, J.T.; Talbert, M.K.; Miller, B.W.; Morisette, J.T. Designing ecological climate change impact assessments to reflect key climatic drivers. Glob. Chang. Biol. 2017, 23, 2537–2553. [Google Scholar] [CrossRef] [PubMed]
- Albano, C.M.; McCarthy, M.I.; Dettinger, M.D.; McAfee, S.A. Techniques for constructing climate scenarios for stress test applications. Clim. Chang. 2021, 164, 1–25. [Google Scholar] [CrossRef]
- Williams, J.W.; Jackson, S.T. Novel climates, no-analog communities, and ecological surprises. Front. Ecol. Environ. 2007, 5, 475–482. [Google Scholar] [CrossRef]
- Van de Pol, M.; Jenouvrier, S.; Cornelissen, J.H.C.; Visser, M.E. Behavioural, ecological and evolutionary responses to extreme climatic events: Challenges and directions. Phil. Trans. R. Soc. B 2017, 372, 20160134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.D. An ecological perspective on extreme climatic events: A synthetic definition and framework to guide future research. J. Ecol. 2011, 99, 656–663. [Google Scholar] [CrossRef]
- Kayler, Z.E.; De Boeck, H.J.; Fatichi, S.; Grünzweig, J.M.; Merbold, L.; Beier, C.; McDowell, N.; Dukes, J.S. Experiments to confront the environmental extremes of climate change. Front. Ecol. Environ. 2015, 13, 219–225. [Google Scholar] [CrossRef]
- Fuentes, M.; Maynard, J.A.; Guinea, M.; Bell, I.P.; Werdell, P.J.; Hamann, M. Proxy indicators of sand temperature help project impacts of global warming on sea turtles in northern Australia. Endanger. Species Res. 2009, 9, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Mair, L.; Jönsson, M.; Räty, M.; Bärring, L.; Strandberg, G.; Lämås, T.; Snäll, T. Land use changes could modify future negative effects of climate change on old-growth forest indicator species. Divers. Distrib. 2018, 24, 1416–1425. [Google Scholar] [CrossRef] [Green Version]
- Moraitis, M.L.; Valavanis, V.D.; Karakassis, I. Modelling the effects of climate change on the distribution of benthic indicator species in the Eastern Mediterranean Sea. Sci. Total Environ. 2019, 667, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Cramer, W.; Bondeau, A.; Woodward, F.I.; Prentice, I.C.; Betts, R.A.; Brovkin, V.; Cox, P.M.; Fisher, V.; Foley, J.A.; Friend, A.D. Global response of terrestrial ecosystem structure and function to CO2 and climate change: Results from six dynamic global vegetation models. Glob. Chang. Biol. 2001, 7, 357–373. [Google Scholar] [CrossRef] [Green Version]
- Daniel, C.J.; Frid, L.; Sleeter, B.M.; Fortin, M. State-and-transition simulation models: A framework for forecasting landscape change. Methods Ecol. Evol. 2016, 7, 1413–1423. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Graham, C.H. The ability of climate envelope models to predict the effect of climate change on species distributions. Glob. Chang. Biol. 2006, 12, 2272–2281. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Jenouvrier, S.; Caswell, H.; Barbraud, C.; Holland, M.; Strœve, J.; Weimerskirch, H. Demographic models and IPCC climate projections predict the decline of an emperor penguin population. Proc. Natl. Acad. Sci. USA 2009, 106, 1844–1847. [Google Scholar] [CrossRef] [Green Version]
- Morgan, M.G.; Pitelka, L.F.; Shevliakova, E. Elicitation of expert judgments of climate change impacts on forest ecosystems. Clim. Chang. 2001, 49, 279–307. [Google Scholar] [CrossRef]
- O’Neill, S.J.; Osborn, T.J.; Hulme, M.; Lorenzoni, I.; Watkinson, A.R. Using expert knowledge to assess uncertainties in future polar bear populations under climate change. J. Appl. Ecol. 2008, 45, 1649–1659. [Google Scholar] [CrossRef]
- Herr, A.; Dambacher, J.M.; Pinkard, E.; Glen, M.; Mohammed, C.; Wardlaw, T. The uncertain impact of climate change on forest ecosystems–How qualitative modelling can guide future research for quantitative model development. Environ. Model. Softw. 2016, 76, 95–107. [Google Scholar] [CrossRef]
- Weeks, D.; Malone, P.; Welling, L. Climate change scenario planning: A tool for managing parks into uncertain futures. Park Sci. 2011, 28, 26–33. [Google Scholar]
- Beeton, T.A.; McNeeley, S.M.; Miller, B.W.; Ojima, D.S. Grounding simulation models with qualitative case studies: Toward a holistic framework to make climate science usable for US public land management. Clim. Risk Manag. 2019, 23, 50–66. [Google Scholar] [CrossRef]
- Marcot, B.G.; Holthausen, R.S.; Raphael, M.G.; Rowland, M.M.; Wisdom, M.J. Using Bayesian belief networks to evaluate fish and wildlife population viability under land management alternatives from an environmental impact statement. For. Ecol. Manage. 2001, 153, 29–42. [Google Scholar] [CrossRef]
- Bode, M.; Baker, C.M.; Benshemesh, J.; Burnard, T.; Rumpff, L.; Hauser, C.E.; Lahoz-Monfort, J.J.; Wintle, B.A. Revealing beliefs: Using ensemble ecosystem modelling to extrapolate expert beliefs to novel ecological scenarios. Methods Ecol. Evol. 2017, 8, 1012–1021. [Google Scholar] [CrossRef] [Green Version]
- Miller, B.W.; Symstad, A.J.; Frid, L.; Fisichelli, N.A.; Schuurman, G.W. Co-producing simulation models to inform resource management: A case study from southwest South Dakota. Ecosphere 2017, 8, e02020. [Google Scholar] [CrossRef] [Green Version]
- Austin, M.P.; Van Niel, K.P. Improving species distribution models for climate change studies: Variable selection and scale. J. Biogeogr. 2011, 38, 1–8. [Google Scholar] [CrossRef]
- Franklin, J.; Davis, F.W.; Ikegami, M.; Syphard, A.D.; Flint, L.E.; Flint, A.L.; Hannah, L. Modeling plant species distributions under future climates: How fine scale do climate projections need to be? Glob. Chang. Biol. 2013, 19, 473–483. [Google Scholar] [CrossRef] [Green Version]
- Beaumont, L.J.; Hughes, L.; Pitman, A.J. Why is the choice of future climate scenarios for species distribution modelling important? Ecol. Lett. 2008, 11, 1135–1146. [Google Scholar] [CrossRef]
- Urban, M.C.; Tewksbury, J.J.; Sheldon, K.S. On a collision course: Competition and dispersal differences create no-analogue communities and cause extinctions during climate change. Proc. R. Soc. B Biol. Sci. 2012, 279, 2072–2080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavergne, S.; Mouquet, N.; Thuiller, W.; Ronce, O. Biodiversity and climate change: Integrating evolutionary and ecological responses of species and communities. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 321–350. [Google Scholar] [CrossRef] [Green Version]
- Ockendon, N.; Baker, D.J.; Carr, J.A.; White, E.C.; Almond, R.E.A.; Amano, T.; Bertram, E.; Bradbury, R.B.; Bradley, C.; Butchart, S.H.M. Mechanisms underpinning climatic impacts on natural populations: Altered species interactions are more important than direct effects. Glob. Chang. Biol. 2014, 20, 2221–2229. [Google Scholar] [CrossRef] [PubMed]
- D’Amen, M.; Rahbek, C.; Zimmermann, N.E.; Guisan, A. Spatial predictions at the community level: From current approaches to future frameworks. Biol. Rev. 2017, 92, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Heikkinen, R.K.; Luoto, M.; Virkkala, R.; Pearson, R.G.; Körber, J. Biotic interactions improve prediction of boreal bird distributions at macro-scales. Glob. Ecol. Biogeogr. 2007, 16, 754–763. [Google Scholar] [CrossRef]
- Davis, A.J.; Jenkinson, L.S.; Lawton, J.H.; Shorrocks, B.; Wood, S. Making mistakes when predicting shifts in species range in response to global warming. Nature 1998, 391, 783–786. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; De Siqueira, M.F.; Grainger, A.; Hannah, L. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Buma, B. Disturbance interactions: Characterization, prediction, and the potential for cascading effects. Ecosphere 2015, 6, 1–15. [Google Scholar] [CrossRef]
- Yates, K.L.; Bouchet, P.J.; Caley, M.J.; Mengersen, K.; Randin, C.F.; Parnell, S.; Fielding, A.H.; Bamford, A.J.; Ban, S.; Barbosa, A.M. Outstanding challenges in the transferability of ecological models. Trends Ecol. Evol. 2018, 33, 790–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkett, V.R.; Wilcox, D.A.; Stottlemyer, R.; Barrow, W.; Fagre, D.; Baron, J.; Price, J.; Nielsen, J.L.; Allen, C.D.; Peterson, D.L. Nonlinear dynamics in ecosystem response to climatic change: Case studies and policy implications. Ecol. Complex. 2005, 2, 357–394. [Google Scholar] [CrossRef] [Green Version]
- Perret, D.L.; Leslie, A.B.; Sax, D.F. Naturalized distributions show that climatic disequilibrium is structured by niche size in pines (Pinus L.). Glob. Ecol. Biogeogr. 2019, 28, 429–441. [Google Scholar] [CrossRef]
- Bonan, G.B.; Doney, S.C. Climate, ecosystems, and planetary futures: The challenge to predict life in Earth system models. Science 2018, 359, 6375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, J.M.; Chalmandrier, L.; Lenoir, J.; Burgess, T.I.; Essl, F.; Haider, S.; Kueffer, C.; McDougall, K.; Milbau, A.; Nuñez, M.A. Lags in the response of mountain plant communities to climate change. Glob. Chang. Biol. 2018, 24, 563–579. [Google Scholar] [CrossRef]
- Bertrand, R.; Lenoir, J.; Piedallu, C.; Riofrio-Dillon, G.; de Ruffray, P.; Vidal, C.; Pierrat, J.-C.; Gégout, J.-C. Changes in plant community composition lag behind climate warming in lowland forests. Nature 2011, 479, 517–520. [Google Scholar] [CrossRef]
- Svenning, J.; Sandel, B. Disequilibrium vegetation dynamics under future climate change. Am. J. Bot. 2013, 100, 1266–1286. [Google Scholar] [CrossRef]
- McLaughlin, B.C.; Zavaleta, E.S. Predicting species responses to climate change: Demography and climate microrefugia in C alifornia valley oak (Quercus lobata). Glob. Chang. Biol. 2012, 18, 2301–2312. [Google Scholar] [CrossRef]
- Williams, J.W.; Ordonez, A.; Svenning, J.-C. A unifying framework for studying and managing climate-driven rates of ecological change. Nat. Ecol. Evol. 2021, 5, 17–26. [Google Scholar] [CrossRef]
- Comte, L.; Grenouillet, G. Do stream fish track climate change? Assessing distribution shifts in recent decades. Ecography 2013, 36, 1236–1246. [Google Scholar]
- Bachelet, D.; Ferschweiler, K.; Sheehan, T.J.; Sleeter, B.M.; Zhu, Z. Projected carbon stocks in the conterminous USA with land use and variable fire regimes. Glob. Chang. Biol. 2015, 21, 4548–4560. [Google Scholar] [CrossRef] [PubMed]
- Slavich, E.; Warton, D.I.; Ashcroft, M.B.; Gollan, J.R.; Ramp, D. Topoclimate versus macroclimate: How does climate mapping methodology affect species distribution models and climate change projections? Divers. Distrib. 2014, 20, 952–963. [Google Scholar] [CrossRef]
- Dullinger, S.; Dirnböck, T.; Grabherr, G. Modelling climate change-driven treeline shifts: Relative effects of temperature increase, dispersal and invasibility. J. Ecol. 2004, 92, 241–252. [Google Scholar] [CrossRef]
- Harsch, M.A.; Bader, M.Y. Treeline form–A potential key to understanding treeline dynamics. Glob. Ecol. Biogeogr. 2011, 20, 582–596. [Google Scholar] [CrossRef]
- Rondeau, R.; Fink, M.; Rodda, G.; Kummel, M. Treeline Monitoring in the San Juan Basin Tundra: A Pilot Project. Ph.D. Thesis, Colorado State University, Fort Collins, CO, USA, 2012. [Google Scholar]
- Fink, M.; Rondeau, R.; Decker, K. Treeline Monitoring in the San Juan Mountains; Unpublished Report, Colorado Natural Heritage Program; Colorado State University: Ft. Collins, CO, USA, 2014; Available online: https://cnhp.colostate.edu/wp-content/uploads/download/documents/2014/Final_Treeline_report_2014.pdf (accessed on 10 September 2021).
- Davis, E.L.; Brown, R.; Daniels, L.; Kavanagh, T.; Gedalof, Z. Regional variability in the response of alpine treelines to climate change. Clim. Chang. 2020, 162, 1365–1384. [Google Scholar] [CrossRef]
- Miller, B.W.; Morisette, J.T. Integrating research tools to support the management of social-ecological systems under climate change. Ecol. Soc. 2014, 19, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.R.; Allan, N.L.; McGowan, C.P.; Szymanski, J.A.; Oetker, S.R.; Bell, H.M. Development of a species status assessment process for decisions under the US Endangered Species Act. J. Fish Wildl. Manag. 2018, 9, 302–320. [Google Scholar] [CrossRef] [Green Version]
- Seglund, A.; Street, P.A.; Aagaard, K.; Runge, J.; Flenner, M. Southern White-Tailed Ptarmigan (Lagopus leucura altipetens) Population Assessment and Conservation Considerations in Colorado, Unpublished Report; Colorado Parks and Wildlife: Denver, CO, USA, 2018.
- Martin, K.; Stacey, P.B.; Braun, C.E. Recruitment, dispersal, and demographic rescue in spatially-structured white-tailed ptarmigan populations. Condor 2000, 102, 503–516. [Google Scholar] [CrossRef]
- Sandercock, B.K.; Martin, K.; Hannon, S.J. Life history strategies in extreme environments: Comparative demography of arctic and alpine ptarmigan. Ecology 2005, 86, 2176–2186. [Google Scholar] [CrossRef]
- Wann, G.T. Reproductive Ecology and Population Viability of Alpine-Endemic Ptarmigan Populations in Colorado. Ph.D. Thesis, Colorado State University, Fort Collins, CO, USA, 2017. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Climate | Ecology | |
|---|---|---|
| Uncertainty Differences in projected climate change or ecological impacts arising from differences across climate models, emission scenarios, and choice and structure of ecological methods and models |
|
|
| Complexity Complex interactions between climate and ecological process and their relevant spatiotemporal scales |
|
|
| Constraints Availability of suitable observed and modeled data at appropriate spatiotemporal scales |
|
|
| Tool | Features | Website |
|---|---|---|
| Climate Toolbox |
| https://climatetoolbox.org/ |
| USGS National Climate Change Viewer |
| https://www2.usgs.gov/landresources/lcs/nccv/maca2/maca2_counties.html |
| US Reclamation’s Downscaled CMIP3 and CMIP5 Climate Projections |
| https://gdo-dcp.ucllnl.org/ |
| KNMI Climate Explorer |
| https://climexp.knmi.nl/start.cgi |
| Climate Futures Toolbox |
| https://www.earthdatascience.org/cft/index.html |
| WorldClim |
| https://www.worldclim.org/data/cmip6/cmip6climate.html |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rangwala, I.; Moss, W.; Wolken, J.; Rondeau, R.; Newlon, K.; Guinotte, J.; Travis, W.R. Uncertainty, Complexity and Constraints: How Do We Robustly Assess Biological Responses under a Rapidly Changing Climate? Climate 2021, 9, 177. https://0-doi-org.brum.beds.ac.uk/10.3390/cli9120177
Rangwala I, Moss W, Wolken J, Rondeau R, Newlon K, Guinotte J, Travis WR. Uncertainty, Complexity and Constraints: How Do We Robustly Assess Biological Responses under a Rapidly Changing Climate? Climate. 2021; 9(12):177. https://0-doi-org.brum.beds.ac.uk/10.3390/cli9120177
Chicago/Turabian StyleRangwala, Imtiaz, Wynne Moss, Jane Wolken, Renee Rondeau, Karen Newlon, John Guinotte, and William Riebsame Travis. 2021. "Uncertainty, Complexity and Constraints: How Do We Robustly Assess Biological Responses under a Rapidly Changing Climate?" Climate 9, no. 12: 177. https://0-doi-org.brum.beds.ac.uk/10.3390/cli9120177