
Does Local Adaptation Impact on the Distribution of Competing Aedes Disease Vectors?


1
Smithsonian Tropical Research Institute, P.O. Box 0843-03092, Balboa Ancon, Panama
2
Instituto de Investigaciones Científicas y Servicios de Alta Tecnología, P.O. Box 0843-01103, Panama City, Panama
3
Programa Centroamericano de Maestría en Entomología, Universidad de Panamá, P.O. Box 0843-01103, Panama City, Panama
*
Authors to whom correspondence should be addressed.
Climate 2021, 9(2), 36; https://0-doi-org.brum.beds.ac.uk/10.3390/cli9020036
Submission received: 16 January 2021
/
Revised: 7 February 2021
/
Accepted: 8 February 2021
/
Published: 22 February 2021
(This article belongs to the Special Issue Impacts of Climate Change on Species)
{kind=link}
{kind=link}
Abstract
:Ae. (Stegomyia) aegypti L. and Aedes (Stegomyia) albopictus Skuse mosquitoes are major arboviral disease vectors in human populations. Interspecific competition between these species shapes their distribution and hence the incidence of disease. While Ae. albopictus is considered a superior competitor for ecological resources and displaces its contender Ae. aegypti from most environments, the latter is able to persist with Ae. albopictus under particular environmental conditions, suggesting species occurrence cannot be explained by resource competition alone. The environment is an important determinant of species displacement or coexistence, although the factors underpinning its role remain little understood. In addition, it has been found that Ae. aegypti can be adapted to the environment across a local scale. Based on data from the Neotropical country of Panama, we present the hypothesis that local adaptation to the environment is critical in determining the persistence of Ae. aegypti in the face of its direct competitor Ae. albopictus. We show that although Ae. albopictus has displaced Ae. aegypti in some areas of Panama, both species coexist across many areas, including regions where Ae. aegypti appear to be locally adapted to dry climate conditions and less vegetated environments. Based on these findings, we describe a reciprocal transplant experiment to test our hypothesis, with findings expected to provide fundamental insights into the role of environmental variation in shaping the landscape of emerging arboviral disease.
1. Introduction
The mosquitoes Ae. aegypti and Ae. albopictus are the primary vectors of arboviruses to humans, including the most common viruses, dengue (DENV), chikungunya (CHIKV) and Zika (ZIKV), but also Mayaro (MYA) and yellow fever (YF). These diseases are collectively responsible for over 50 million cases and 50,000 deaths every year globally [1,2]. Although there is a vaccine for YF, human cases are on the rise [3] and despite attempts to reduce the burden of disease [4,5], there is currently no effective prevention scheme for other Aedes-borne diseases. As a result, the reduction of arboviral disease transmission relies on population control including the larval habitat destruction, adult insecticide spraying and the genetic or biological manipulation of vector populations [6]. The successful implementation of any control method depends on a good understanding of mosquito ecology and evidence-based decisions of when and where this should be implemented [4]. To this end, disease prediction models are an important resource, although their accuracy depends on the specificity of the biological parameters which are input into the model [7]. An important consideration for disease prediction is how Ae. mosquitoes may shift their geographical distributions as a result of both biological competition and environmental variation, and how this may influence the disease transmission landscape [4]. Although shifts in Aedes distributions due to climate change have been explored, such models generally disregard the impact of the adaptive potential of local populations on the outcome of inter-specific interactions, which could be a critical driver of biological parameters shaping Aedes geographical distributions [8,9]. Furthermore, locally adapted populations can severely impact vector control efforts that are based on a gene drive system, either by promoting or limiting the spread of the target characteristic [10].
2. The Displacement of Ae. aegypti by Ae. albopictus Is Context-Dependent
The global distribution of Ae. aegypti has recently been shifting due to the invasion of the ecologically similar Asian tiger mosquito, Ae. albopictus. While Ae. aegypti has been widespread since its expansion from Africa during the 17th century, Ae. albopictus has been expanding from Asia to be globally distributed within the last ~40 years [11]. As a result, Ae. albopictus has displaced resident Ae. aegypti from many locations including Panama, the South Eastern USA, Madagascar, La Reunion, Mayotte and Bermuda [12,13,14,15,16,17,18,19,20,21]. Displacement of Ae. aegypti is the expected outcome given that Ae. albopictus is a superior competitor under semi-field conditions [14,15]. However, both mosquitoes also often persist together under particular environmental conditions, with this co-existence being stable over time [12,21], which suggests that local adaptation to the environment could play a role in determining displacement or coexistence. Although such studies have found differences in the distribution or abundance of Aedes species across geographical regions, few studies have looked at adaptive differences across these populations [17,22,23], and none address the impact of local adaptation on the Aedes competitive interaction.
The main mechanisms allowing Ae. albopictus to outcompete and displace Ae. aegypti are cited as satyrization, where interspecific mating reduces Ae. aegypti fitness because hybrids of the latter are not produced, and the biological competition for ecological resources at the larval stage [15,24]. A clear understanding of the factors underpinning the condition dependent displacement of Ae. aegypti remain elusive, but environmental conditions appear to play a key role. Previous studies have suggested that the ability of Ae. aegypti to persist, despite invasion by Ae. albopictus, is associated with dry climate conditions and/or urban environments [9]. These findings are largely based on studies from Florida, which show that Ae. aegypti are found in higher abundances in urbanised areas than Ae. albopictus, which are more abundant in rural or vegetated settings [9,25]. Differences in species distributions are also associated with changes in rainfall, humidity and temperature across the sampled range [26,27]. Moreover, we [21] recently described the geographical distributions of Ae. aegypti and Ae. albopictus across the heterogeneous landscape of Panama, including across the Azuero Peninsula, an isolated region subject to a sharp west to east environmental gradient (i.e., >150 km) (Figure 1). In this study, Ae. aegypti was shown to persist with Ae. albopictus only in regions with a dry and seasonal climate, while displacement was observed in wet tropical areas with a higher humidity and more vegetation. This condition dependent displacement was seen despite both species sharing a similar ecological niche and was hypothesized to result from environmental variation shaping the nature of the species interaction, including the outcome of biological competition.
What permits Ae. aegypti to persist with Ae. albopictus under some environmental conditions is still unknown. It is thought Ae. aegypti could perform better under dry climate conditions because they are better adapted [9]. For example, the eggs of Ae. aegypti are more tolerant to higher temperatures and can withstand desiccation for longer periods of time in comparison to the eggs of Ae. albopictus, which are able to survive lower temperatures through diapause [9,28]. Innate between-species differences in biological parameters are likely to play a role, but may not provide a full explanation, particularly across tropical environments where temporal variation in environmental variables is limited. In addition to such differences, there is also the possibility that local adaptive differences are seen within populations across a regional scale. Whether mosquitoes are locally adapted is an important consideration when predicting the outcome of species competition since locally adapted species may have a competitive edge and reduce the successful establishment of invasive competitors [29,30,31,32], particularly when confronted by an environmental gradient [33]. Studies using genomic technology provide an invaluable opportunity to link the underlying genomic variation to associated biological parameters. Such an approach provides information on population fitness, providing a comprehensive method to the study of Aedes persistence.
3. The Genomic Signal of Local Environmental Adaptation in Ae. aegypti
There is evidence that populations of Ae. mosquitoes are locally adapted to their environments. Previous experimental work has shown that populations of Ae. aegypti from divergent climates are locally adapted at the larval stage, because they develop at different rates when reared under the same temperature conditions [23]. In addition, the ability of Ae. aegypti to compete with Ae. albopictus under laboratory conditions differs among populations [17]. More recently, studies of genomics have revealed that Aedes populations are divergent at loci correlated with various environmental parameters [22,34]. This includes across broad spatial scales for Ae. aegypti, associated with human population density [35]. Moreover, a recent study by our research group [36] used a landscape genomics approach to reveal that different populations of Ae. aegypti across Panama, have differences in genomic variation associated with the environment across a small spatial scale and despite low population structure, i.e., high gene flow. This variation is likely to reflect differential adaption to heterogeneous environments across the country. In particular, we found that genomic loci with a signal of local adaptation differed between wet tropical conditions such as those experienced along the Caribbean coast and the dry tropical conditions typical of the Pacific coast of Panama. Interestingly, the eastern Azuero Peninsula, which is the driest region in Panama (Figure 1), had the least variation in the composition of its putatively adaptive alleles, suggesting these alleles could be fixed in the population. Furthermore, differences in this putatively adaptive genomic variation were largely associated with temperature and vegetation, which are important for the development and survival of Ae. aegypti eggs and larvae [37,38,39]. Of all the loci potentially under natural selection, 128 were consistently identified as outliers and likely to be true positives. These loci provide potential targets for future studies into the local adaptation of Ae. aegypti.
4. Local Adaptation in Ae. aegypti Determines Coexistence with Ae. albopictus
Given the findings of our two recent studies from Panama that there is condition-dependent displacement, and that some populations of Ae. aegypti have putatively adaptive loci in areas of coexistence, we hypothesise that local environmental adaptation of Ae. aegypti might allow them to persist in competition with invading Ae. albopictus. Since Ae. aegypti has been present in Panama for the last ~100 years [40], it is not surprising that they would be locally adapted. However, since Ae. albopictus only invaded Panama in 2002 and are still expanding [40], we do not expect their populations to be adapted to the local conditions. Therefore, Ae. aegypti could be better able to survive and exploit local habitats than Ae. albopictus. If local adaptation plays a role in population persistence, we would expect Ae. aegypti to harbour genomic loci with a signal of selection and correlated to the local environmental conditions in regions where both species occur.
We previously identified areas within Panama which are likely to have populations of Ae. aegypti with adaptive genomic variation based on Generalised Dissimilarity Modelling analysis. This method compares the turnover in the composition of loci which are not expected to be under selection, i.e., neutral loci, to the compositional turnover in the candidate adaptive loci over geographical space and takes into account differences in the environment [36]. We overlaid our Aedes co-occurrence data [21] from Panama onto the graphical representation of adaptive genomic variation of Ae. aegypti across the country. Interestingly, this comparison reveals that both Ae. aegypti and Ae. albopictus tend to coexist in dry tropical regions where Ae. aegypti have divergent candidate loci, i.e., where Ae. aegypti are potentially adapted to the local environment (Figure 1). This finding is consistent with the prediction that local environmental adaptation contributes to Ae. aegypti persistence. For example, Ae. aegypti with putatively adaptive loci were found within the dry tropical Pacific regions of Chiriquí (David), Coclé, the eastern Azuero Peninsula and provincial Panamá where both species co-occur [21,41]. In comparison, Ae. aegypti were no longer found in many areas where these particular set of candidate adaptive alleles were not detected (See Figure 2 for reference to the regions within Panama). Although there was also genetic evidence for local adaptation in the isolated wet tropical region of Bocas del Toro and Costa Abajo near Colon, it is unknown whether this variation will allow Ae. aegypti to resist invasion by Ae. albopictus, given that Ae. albopictus was only recorded in Costa Abajo in 2018 and has not yet reached Bocas del Toro. Although tantalising, the patterns observed here are correlational and based on genomic information alone without the measurement of biological parameters. The hypothesis that local adaptation to the environment in Ae. aegypti influences their persistence on invasion by Ae. albopictus requires further confirmation within an experimental framework.
5. An Experimental Framework to Test for Local Adaptation in the Presence of a Competitor
Given that both Aedes species coexist in the dry tropical eastern Azuero Peninsula of Panama, where Ae. aegypti appear to be locally adapted, but not in the wet tropical conditions of the southwestern Azuero Peninsula, the hypothesis could be tested with a reciprocal transplant experiment (Figure 2). First, to confirm whether Ae. aegypti are truly adapted to the local environmental conditions, the impact of different climate conditions on a range of fitness phenotypes should be determined. The expectation is that Ae. aegypti adapted to the local environmental conditions will have a higher fitness within their source environment and experience a lower fitness on translocation to divergent environmental conditions. The experimental design requires at least F1 Ae. aegypti acquired from the controlled mating of both putatively ‘dry adapted’ (i.e., East Azuero) and ‘wet adapted’ (i.e., Southwest Azuero) laboratory colonies sourced from wild populations. This is both to ensure parentage and so that all samples are subjected to the same ambient conditions before their placement within the experiment, i.e., to ensure that the observed response to the environment results from genetic heritage and not environmental plasticity. Eggs obtained from the matings will form the basis of the experiment, whereby replicate batches of mosquitoes are allowed to develop under semi-field conditions in both the local environment and alternative environment. Differences in observed fitness phenotypes such as egg hatch rate, development time, larval survival and female wing size as a proxy for fecundity can be determined through comparative statistical analysis. Here, since both populations are tested within their source environment, any natural differences between populations will be apparent. However, it is important to note that care must be taken when extrapolating to other regions, especially where the invasion history of Ae. aegypti is unknown. In this circumstance, innate differences in the biological traits of populations, i.e., desiccation resistance, should be tested and/or controlled for.
Second, to test whether the local adaptation of Ae. aegypti impacts the outcome of biological competition, these same phenotypic traits should be measured with and without the addition of Ae. albopictus within the experimental set up. If the fitness of Ae. aegypti is uniformly reduced in the presence of Ae. albopictus in both wet and dry environments, then species replacement of Ae. aegypti is expected to occur across Panama regardless of climate conditions. However, if Ae. aegypti is able to maintain high fitness in the presence of Ae. albopictus in dry but not in wet environments, this mechanism could explain the condition-dependent nature of species co-existence. Third, genomic technology targeting the genomic loci we previously identified as putatively under selection should be used to link phenotype to genotype. For example, laboratory experiments using mosquitoes have been used to link quantitative trait loci to phenotypes relating to diapause, viral infection or resistance to insecticides and filarial parasites [42,43,44,45]. Informative quantitative analyses [46,47] can be performed to identify which genomic regions are truly impacted by local adaptation and later targeted for functional studies.
6. Implications
An understanding of both the function and distribution of adaptive genes will broaden our knowledge of the biological requirements of Aedes species important for disease prediction. Improving the accuracy of biological parameters will aid models predicting the outcome of population control, i.e., through the release of Wolbachia bacterium, which relies on an increased fitness to the mosquito to promote its spread [10]. Importantly, an understanding of how fitness and adaptive traits are distributed in relation to the environment allows us to consider the adaptive potential when modelling future population responses [48]. With a rise in gene editing strategies and CRISPR technology [49], adaptive genes could be potential targets for population control, either as targets for modification or as genes conferring increased fitness and/or traits that aid the gene drive of disease resistance. Determining whether local adaptation impacts on Aedes competitive interactions is important because species co-occurrence could facilitate the emergence of sylvatic arboviral disease. Ae. albopictus is an opportunistic feeder, able to utilise a wide range of peri-domestic habitats outside of its native range [50,51] and the species could act as an efficient bridge vector for emergent zoonotic diseases from the forest [51]. The addition of the specialised human commensal Ae. aegypti provides the opportunity for any emergent epidemic to spread and be maintained within the urban population [52,53,54,55]. Untangling the factors enabling cooccurrence of species will allow us to identify locations with a higher risk of emergent sylvatic disease and act towards preventing new urban outbreaks. In areas where Ae. albopictus has displaced Ae. aegypti, control strategies currently aimed at targeting Ae. aegypti will require modification to account for their wider use of peridomestic habitats and wider range of animal hosts. In addition, gene drive systems which are effective in reducing disease transmission by Ae. aegypti [56] will require development for the targeted control of Ae. albopictus.
Author Contributions
K.L.B., W.O.M. and J.R.L. designed the study. K.L.B. and J.R.L. performed the data analysis and figure preparation. K.L.B. and J.R.L. wrote the manuscript with contributions from W.O.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was sponsored in part by the Government of Panama through MINSA’s grant Zika Project (Objective #2) to J.R.L. Support for KLB comes from the Smithsonian’s George E. Burch Fellowship and Smithsonian Fellowship Program, the Edward and Jeanne Kashian Family Foundation and Mr Nicholas Logothetis of Chartwell Consulting Group Inc. The National System of Investigation (SNI) of SENACYT currently supports research activities by J.R.L. (157-2017; 16-2020).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are previously available in the provided references of Bennett et al. (2021) and Bennett et al. (2021).
Acknowledgments
We are thankful to the Panama’s Ministry of Environment (Mi Ambiente) and MINSA, the Smithsonian Tropical Research Institute (STRI) and the Instituto de Investigaciones Científicas y Servicios de Alta Tecnología (INDICASAT) for supporting our research and project logistics.
Conflicts of Interest
The authors received funding from The Edward M. and Jeanne C. Kashian Family Foundation Inc., and Nicholas Logothetis of Chartwell Consulting. There are no patents, products in development or marketed products associated with this research to declare.
References
- World Health Organization. Yellow Fever Fact Sheet No. 100. 2014. Available online: www.who.int/mediacentre/factsheets/fs100/en (accessed on 7 July 2020).
- World Health Organization. World Health Report. Insect-Borne Diseases. Available online: www.who.int/whr/1996/media_centre/executive_summary1/en/index9.html (accessed on 7 July 2020).
- Barrett, A.D.T. The reemergence of yellow fever. Science 2018, 361, 847–848. [Google Scholar] [CrossRef]
- Roiz, D.; Wilson, A.L.; Scott, T.W.; Fonseca, D.M.; Jourdain, F.; Müller, P.; Velayudhan, R.; Corbel, V. Integrated Aedes management for the control of Aedes-borne diseases. PLoS Negl. Trop. Dis. 2018, 12, e0006845. [Google Scholar] [CrossRef] [Green Version]
- López-Camacho, C.; Kim, Y.C.; Abbink, P.; Larocca, R.A.; Huiskonen, J.T.; Barouch, D.H.; Reyes-Sandoval, A. Assessment of immunogenicity and efficacy of a Zika vaccine using modified vaccinia ankara virus as carriers. Pathogens 2019, 8, 216. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Dhama, K.; Khandia, R.; Munjal, A.; Karthik, K.; Tiwari, R.; Chakraborty, S.; Malik, Y.S.; Bueno-Marí, R. Prevention and control strategies to counter Zika virus, a special focus on intervention approaches against vector mosquitoes-current updates. Front. Microbiol. 2018, 9, 87. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Legros, M.; Gould, F.; Lloyd, A.L. Understanding uncertainties in model-based predictions of Ae. aegypti population dynamics. PLoS Negl. Trop. Dis. 2010, 4, e830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kearney, M.; Porter, W.P.; Williams, C.; Ritchie, S.; Hoffmann, A.A. Integrating biophysical models and evolutionary theory to predict climatic impacts on species’ ranges: The dengue mosquito Ae. aegypti in Australia. Funct. Ecol. 2009, 23, 528–538. [Google Scholar] [CrossRef]
- Lounibos, L.P.; Juliano, S.A. Where vectors collide: The importance of mechanisms shaping the realized niche for modeling ranges of invasive Ae. mosquitoes. Biol. Invasions 2018, 20, 1913–1929. [Google Scholar] [CrossRef] [PubMed]
- Hancock, P.A.; White, V.L.; Ritchie, S.A.; Hoffmann, A.A.; Godfray, H.C.J. Predicting Wolbachia invasion dynamics in Ae. aegypti populations using models of density-dependent demographic traits. BMC Biol. 2016, 14, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.N.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Ae. aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef]
- Bargielowski, I.; Carrasquilla, M.C.; Nishimura, N.; Lounibos, L.P. Coexistence of Ae. aegypti and Ae. albopictus (Diptera: Culicidae) in Peninsular Florida two decades after competitive displacements. J. Med. Entomol. 2016, 53, 1385–1390. [Google Scholar] [CrossRef]
- O’Meara, G.F.; Evans Leonard, F.J.; Gettman, A.D.; Cuda, J.P. Spread of Ae. albopictus and decline of Ae. aegypti (Diptera: Culicidae) in Florida. J. Med. Entomol. 1995, 32, 554–562. [Google Scholar] [CrossRef]
- Braks, M.A.H.; Honório, N.A.; Lounibos, L.P.; Lourenço-de-Oliveira, R.; Juliano, S.A. Interspecific competition between two invasive species of container mosquitoes, Ae. aegypti and Ae. albopictus (Diptera: Culicidae), in Brazil. Ann. Entomol. Soc. Am. 2004, 97, 130–139. [Google Scholar] [CrossRef]
- Juliano, S.A. Species Introduction and replacement among mosquitoes: Interspecific resource competition or apparent competition? Ecology 1998, 79, 255–268. [Google Scholar] [CrossRef]
- Kaplan, L.; Kendell, D.; Robertson, D.; Livdahl, T.; Khatchikian, C. Ae. aegypti and Ae. albopictus in Bermuda: Extinction, invasion, invasion and extinction. Biol. Invasions 2010, 12, 3277–3288. [Google Scholar] [CrossRef]
- Leisnham, P.; Juliano, S. Interpopulation differences in competitive effect and response of the mosquito Ae. aegypti and resistance to invasion by a superior competitor. Oecologia 2010, 164, 221–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raharimalala, F.N.; Ravaomanarivo, L.H.; Ravelonandro, P.; Rafarasoa, L.S.; Zouache, K.; Tran-Van, V.; Mousson, L.; Failloux, A.-B.; Hellard, E.; Moro, C.V.; et al. Biogeography of the two major arbovirus mosquito vectors, Ae. aegypti and Ae. albopictus (Diptera, Culicidae), in Madagascar. Parasit. Vectors 2012, 5, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagny Beilhe, L.; Delatte, H.; Quilici, S.; Fontenille, D. Progressive decrease in Ae. aegypti distribution in Reunion Island since the 1900s. J. Med. Entomol. 2009, 46, 1541–1545. [Google Scholar] [CrossRef]
- Bagny Beilhe, L.; Arnoux, S.; Delatte, H.; Lajoie, G.; Fontenille, D. Spread of invasive Ae. albopictus and decline of resident Ae. aegypti in urban areas of Mayotte 2007–2010. Biol. Invasions 2012, 14, 1623–1633. [Google Scholar] [CrossRef]
- Bennett, K.; McMillan, W.; Enríquez, V.; Barraza, E.; Díaz, M.; Baca, B.; Whiteman, A.; Medina, J.; Ducasa, M.; Gómez Martínez, C.; et al. The role of heterogeneous environmental conditions in shaping the spatiotemporal distribution of competing Ae. mosquitoes in Panama: Implications for the landscape of arboviral disease. Biol. Invasions 2021, in press. [Google Scholar]
- Sherpa, S.; Guéguen, M.; Renaud, J.; Blum, M.G.B.; Gaude, T.; Laporte, F.; Akiner, M.; Alten, B.; Aranda, C.; Barre-Cardi, H.; et al. Predicting the success of an invader: Niche shift versus niche conservatism. Ecol. Evol. 2019, 9, 12658–12675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tun-Lin, W.; Burkot, T.R.; Kay, B.H. Effects of temperature and larval diet on development rates and survival of the dengue vector Ae. aegypti in north Queensland, Australia. Med. Vet. Entomol. 2000, 14, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Tripet, F.; Lounibos, L.P.; Robbins, D.; Moran, J.; Nishimura, N.; Blosser, E.M. Competitive reduction by satyrization? Evidence for interspecific mating in nature and asymmetric reproductive competition between invasive mosquito vectors. Am. J. Trop. Med. Hyg. 2011, 85, 265–270. [Google Scholar] [CrossRef]
- Lounibos, L.P.; Kramer, L.D. Invasiveness of Ae. aegypti and Ae. albopictus and vectorial capacity for chikungunya virus. J. Infect. Dis. 2016, 214, S453–S458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopperstad, K.A.; Reiskind, M.H. Recent changes in the local distribution of Ae. aegypti (Diptera: Culicidae) in South Florida, USA. J. Med. Entomol. 2016, 53, 836–842. [Google Scholar] [CrossRef]
- Reiskind, M.H.; Lounibos, L.P. Spatial and temporal patterns of abundance of Ae. aegypti L. (Stegomyia aegypti) and Ae. albopictus (Stegomyia albopictus) (Skuse) in southern Florida. Med. Vet. Entomol. 2013, 27, 421–429. [Google Scholar] [CrossRef]
- Juliano, S.A.; O’Meara, G.F.; Morrill, J.R.; Cutwa, M.M. Desiccation and thermal tolerance of eggs and the coexistence of competing mosquitoes. Oecologia 2002, 130, 458–469. [Google Scholar] [CrossRef] [Green Version]
- Carroll, S.P. Facing change: Forms and foundations of contemporary adaptation to biotic invasions. Mol. Ecol. 2008, 17, 361–372. [Google Scholar] [CrossRef]
- Leibold, M.A.; Urban, M.C.; De Meester, L.; Klausmeier, C.A.; Vanoverbeke, J. Regional neutrality evolves through local adaptive niche evolution. Proc. Natl. Acad. Sci. USA 2019, 116, 2612–2617. [Google Scholar] [CrossRef] [Green Version]
- Pinsky, M.L. Species coexistence through competition and rapid evolution. Proc. Natl. Acad. Sci. USA 2019, 116, 2407–2409. [Google Scholar] [CrossRef] [Green Version]
- Urban, M.C.; De Meester, L. Community monopolization: Local adaptation enhances priority effects in an evolving metacommunity. Proc. R. Soc. Biol. Sci. 2009, 276, 4129–4138. [Google Scholar] [CrossRef] [PubMed]
- Case, T.J.; Taper, M.L. Interspecific competition, environmental gradients, gene flow, and the coevolution of species’ borders. Am. Nat. 2000, 155, 583–605. [Google Scholar] [CrossRef]
- Sherpa, S.; Blum, M.G.B.; Després, L. Cold adaptation in the Asian tiger mosquito’s native range precedes its invasion success in temperate regions. Evolution 2019, 73, 1793–1808. [Google Scholar] [CrossRef] [PubMed]
- Sherpa, S.; Rioux, D.; Goindin, D.; Fouque, F.; François, O.; Després, L. At the origin of a worldwide invasion: Unraveling the genetic makeup of the caribbean bridgehead populations of the dengue vector Ae. aegypti. Genome Biol. Evol. 2018, 10, 56–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, K.L.; McMillan, W.O.; Loaiza, J.R. The genomic signal of local adaptation in Ae. aegypti mosquitoes. Evol. Appl. 2021, in press. [Google Scholar] [CrossRef]
- Brady, O.J.; Johansson, M.A.; Guerra, C.A.; Bhatt, S.; Golding, N.; Pigott, D.M.; Delatte, H.; Grech, M.G.; Leisnham, P.T.; Maciel-de-Freitas, R.; et al. Modelling adult Ae. aegypti and Ae. albopictus survival at different temperatures in laboratory and field settings. Parasit. Vectors 2013, 6, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lounibos, L.P.; O’Meara, G.F.; Juliano, S.A.; Nishimura, N.; Escher, R.L.; Reiskind, M.H.; Cutwa, M.; Greene, K. Differential survivorship of invasive mosquito species in South Florida cemeteries: Do site-specific microclimates explain patterns of coexistence and exclusion? Ann. Entomol. Soc. Am. 2010, 103, 757–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vezzani, D.; Rubio, A.; Velázquez, S.M.; Schweigmann, N.; Wiegand, T. Detailed assessment of microhabitat suitability for Ae. aegypti (Diptera: Culicidae) in Buenos Aires, Argentina. Acta Trop. 2005, 95, 123–131. [Google Scholar] [CrossRef]
- Eskildsen, G.A.; Rovira, J.R.; Dutari, L.C.; Smith, O.; Miller, M.J.; Bennett, K.L.; McMillan, W.O.; Loaiza, J.R. Maternal invasion history of Ae. aegypti and Ae. albopictus into the Isthmus of Panama: Implications for the control of emergent viral disease agents. PLoS ONE 2018, 13, e0194874. [Google Scholar] [CrossRef] [Green Version]
- Bennett, K.L.; Martínez Gómez, C.; Almanza, A.; Rovira, J.R.; McMillan, W.O.; Enriquez, V.; Barraza, E.; Diaz, M.; Galan, J.S.; Whiteman, A.; et al. High infestation of invasive Ae. mosquitoes along the highways of Panama: Impact on vector ecology and disease control. Parasit. Vectors 2019, 12, 264. [Google Scholar] [CrossRef] [Green Version]
- Juneja, P.; Ariani, C.V.; Ho, Y.S.; Akorli, J.; Palmer, W.J.; Pain, A.; Jiggins, F.M. Exome and transcriptome sequencing of Ae. aegypti identifies a locus that confers resistance to Brugia malayi and alters the immune response. PLoS Pathog. 2015, 11, e1004765. [Google Scholar] [CrossRef] [Green Version]
- Faucon, F.; Dusfour, I.; Gaude, T.; Navratil, V.; Boyer, F.; Chandre, F.; Sirisopa, P.; Thanispong, K.; Juntarajumnong, W.; Poupardin, R.; et al. Identifying genomic changes associated with insecticide resistance in the dengue mosquito Ae. aegypti by deep targeted sequencing. Genome Res. 2015, 25, 1347–1359. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, W.E.; Emerson, K.J.; Catchen, J.M.; Cresko, W.A.; Holzapfel, C.M. Footprints in time: Comparative quantitative trait loci mapping of the pitcher-plant mosquito, Wyeomyia smithii. Proc. R. Soc. Biol. Sci. 2012, 279, 4551–4558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, K.E.; Flick, D.; Fleming, K.H.; Jochim, R.; Beaty, B.J.; Black, W.C., IV. Quantitative Trait Loci that control dengue-2 virus dissemination in the mosquito Ae. aegypti. Genetics 2005, 170, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Josephs, E.B.; Berg, J.J.; Ross-Ibarra, J.; Coop, G. Detecting Adaptive Differentiation in Structured Populations with Genomic Data and Common Gardens. Genetics 2019, 211, 989–1004. [Google Scholar] [CrossRef] [Green Version]
- Montesinos-López, O.A.; Martín-Vallejo, J.; Crossa, J.; Gianola, D.; Hernández-Suárez, C.M.; Montesinos-López, A.; Juliana, P.; Singh, R. New Deep Learning Genomic-Based Prediction Model for Multiple Traits with Binary, Ordinal, and Continuous Phenotypes. G3 Genes Genomes Genet. 2019, 9, 1545–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldvogel, A.-M.; Feldmeyer, B.; Rolshausen, G.; Exposito-Alonso, M.; Rellstab, C.; Kofler, R.; Mock, T.; Schmid, K.; Schmitt, I.; Bataillon, T.; et al. Evolutionary genomics can improve prediction of species’ responses to climate change. Evol. Lett. 2020, 4, 4–18. [Google Scholar] [CrossRef]
- Li, M.; Yang, T.; Kandul, N.P.; Bui, M.; Gamez, S.; Raban, R.; Bennett, J.; Sánchez, C.H.M.; Lanzaro, G.C.; Schmidt, H.; et al. Development of a confinable gene drive system in the human disease vector Ae. aegypti. eLife 2020, 9, e51701. [Google Scholar] [CrossRef] [PubMed]
- Faraji, A.; Egizi, A.; Fonseca, D.M.; Unlu, I.; Crepeau, T.; Healy, S.P.; Gaugler, R. Comparative host feeding patterns of the Asian tiger mosquito, Ae. albopictus, in urban and suburban Northeastern USA and implications for disease transmission. PLoS Negl. Trop. Dis. 2014, 8, e3037. [Google Scholar] [CrossRef] [Green Version]
- Pereira Dos Santos, T.; Roiz, D.; Santos de Abreu, F.V.; Luz, S.L.B.; Santalucia, M.; Jiolle, D.; Santos Neves, M.S.A.; Simard, F.; Lourenço-de-Oliveira, R.; Paupy, C. Potential of Ae. albopictus as a bridge vector for enzootic pathogens at the urban-forest interface in Brazil. Emerg. Microbes Infect. 2018, 7, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.E.; Evans, B.R.; Zheng, W.; Obas, V.; Barrera-Martinez, L.; Egizi, A.; Zhao, H.; Caccone, A.; Powell, J.R. Human impacts have shaped historical and recent evolution in Ae. aegypti, the dengue and yellow fever mosquito. Evolution 2014, 68, 514–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trpis, M.; Hausermann, W. Genetics of house-entering behaviour in East African populations of Ae. aegypti (L.) (Diptera: Culicidae) and its relevance to speciation. Bull. Entomol. Res. 1978, 68, 521–532. [Google Scholar] [CrossRef]
- McBride, C.S.; Baier, F.; Omondi, A.B.; Spitzer, S.A.; Lutomiah, J.; Sang, R.; Ignell, R.; Vosshall, L.B. Evolution of mosquito preference for humans linked to an odorant receptor. Nature 2014, 515, 222. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.R.; Tabachnick, W.J. History of domestication and spread of Ae. aegypti—A Review. Mem. Inst. Oswaldo Cruz. 2013, 108, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.E.; Franz, A.W.E.; Reid, W.R.; Olson, K.E. Antiviral effectors and gene drive strategies for mosquito population suppression or replacement to mitigate arbovirus transmission by Ae. aegypti. Insects 2020, 11, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
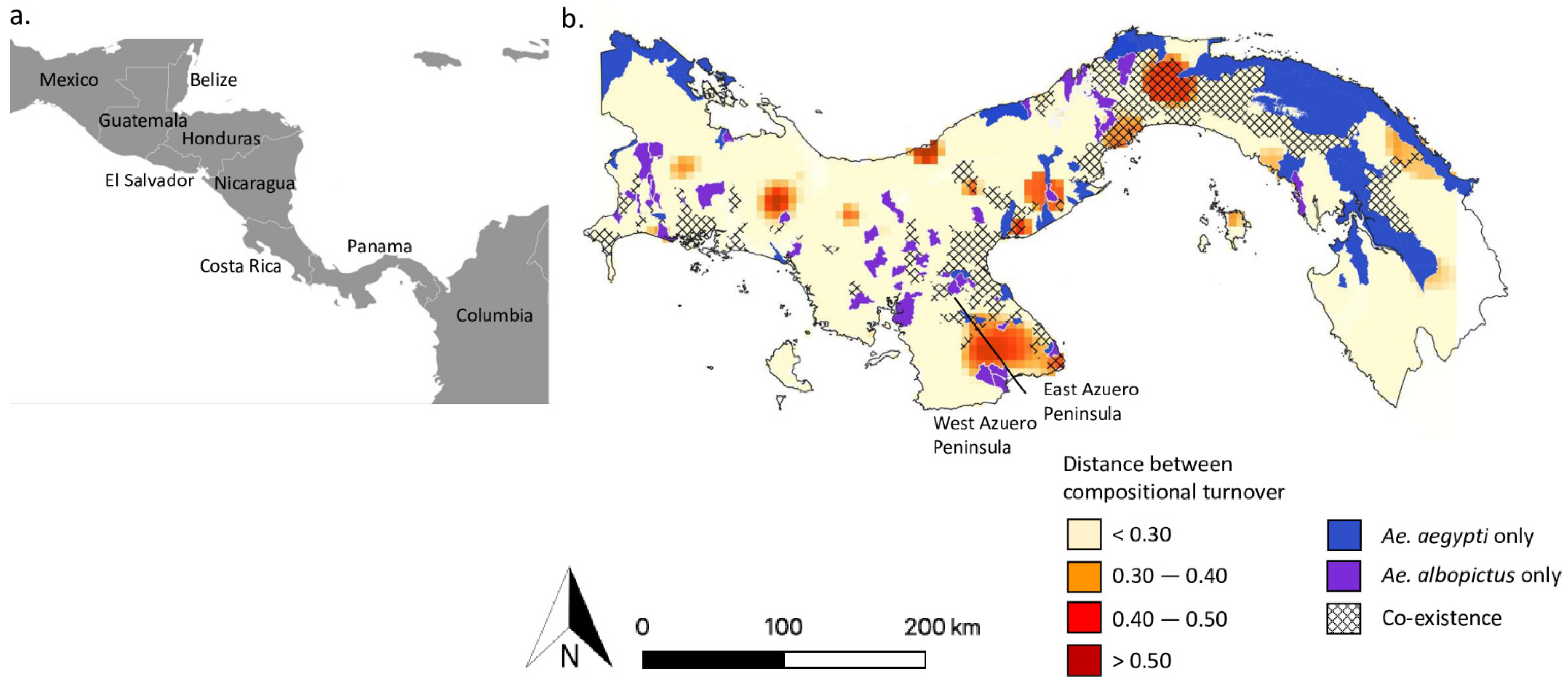
Figure 1.
Putatively adaptive loci are predicted in Ae. aegypti populations within areas of species co-existence. (a) The location of Panama within Central America. (b) Counties with Aedes co-occurrence (dashed areas) are shown based on the recent species distributions as reported by Bennett et al. from 2018 and the Panamanian Ministry of Health (MINSA) in 2017 [21]. The co-occurrence data are overlaid onto a map with red coloured areas representing the potential environmental adaptation of Ae. aegypti from Bennett et al. [36]. These areas were identified through a Generalised Dissimilarity Modelling analysis and represent the difference in allele compositional turnover between a putatively neutral reference loci dataset and a putatively adaptive candidate dataset, using a Procrustes superimposition on their PCA ordinations. A greater predicted distance between the allele composition of the reference and candidate datasets provides a greater indication of populations with adaptive alleles. Purple and blue counties indicate Aedes sampling without co-existence.
Figure 1.
Putatively adaptive loci are predicted in Ae. aegypti populations within areas of species co-existence. (a) The location of Panama within Central America. (b) Counties with Aedes co-occurrence (dashed areas) are shown based on the recent species distributions as reported by Bennett et al. from 2018 and the Panamanian Ministry of Health (MINSA) in 2017 [21]. The co-occurrence data are overlaid onto a map with red coloured areas representing the potential environmental adaptation of Ae. aegypti from Bennett et al. [36]. These areas were identified through a Generalised Dissimilarity Modelling analysis and represent the difference in allele compositional turnover between a putatively neutral reference loci dataset and a putatively adaptive candidate dataset, using a Procrustes superimposition on their PCA ordinations. A greater predicted distance between the allele composition of the reference and candidate datasets provides a greater indication of populations with adaptive alleles. Purple and blue counties indicate Aedes sampling without co-existence.

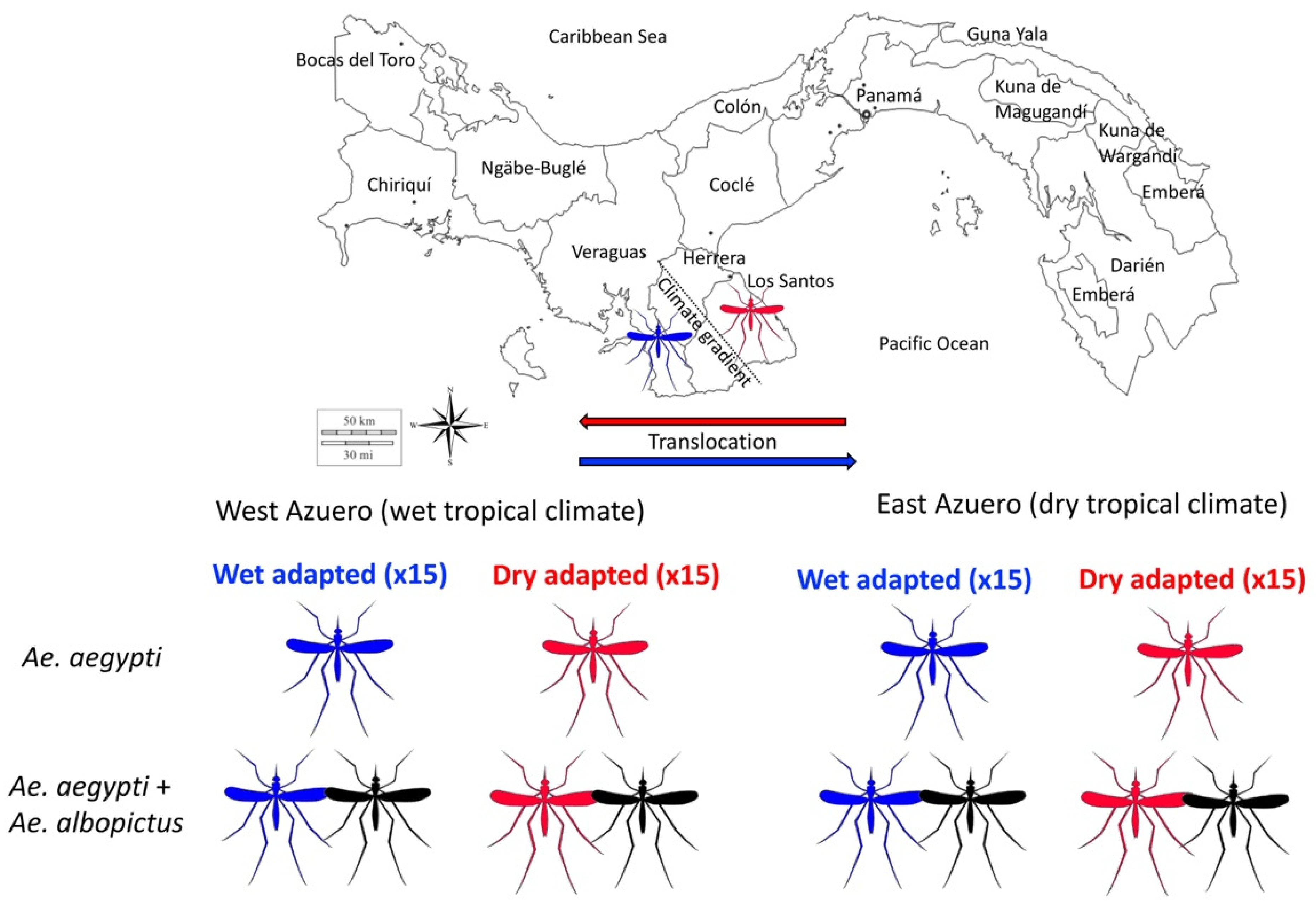
Figure 2.
Reciprocal transplant experiment to test the hypothesis that local adaptation promotes the persistence of Ae. aegypti in the presence of a competitor. Laboratory generated populations of “dry adapted” Ae. aegypti sourced from the dry and hot East Azuero Peninsula (red mosquito) could be tested for fitness phenotype and genotypes as they develop both in their local environment and when transplanted across a climate gradient to the wetter region of the Southwest Azuero Peninsula. Similarly, “wet adapted” Ae. aegypti from the West Azuero Peninsula (blue mosquito) are transplanted to the dry East Azuero Peninsula as well as being tested under local conditions. In addition, the fitness phenotypes and genomic variation of both wet and dry adapted Ae. aegypti should be characterised with and without its competitor, Ae. albopictus (black mosquito), under both local and contrasting climate conditions.
Figure 2.
Reciprocal transplant experiment to test the hypothesis that local adaptation promotes the persistence of Ae. aegypti in the presence of a competitor. Laboratory generated populations of “dry adapted” Ae. aegypti sourced from the dry and hot East Azuero Peninsula (red mosquito) could be tested for fitness phenotype and genotypes as they develop both in their local environment and when transplanted across a climate gradient to the wetter region of the Southwest Azuero Peninsula. Similarly, “wet adapted” Ae. aegypti from the West Azuero Peninsula (blue mosquito) are transplanted to the dry East Azuero Peninsula as well as being tested under local conditions. In addition, the fitness phenotypes and genomic variation of both wet and dry adapted Ae. aegypti should be characterised with and without its competitor, Ae. albopictus (black mosquito), under both local and contrasting climate conditions.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bennett, K.L.; McMillan, W.O.; Loaiza, J.R. Does Local Adaptation Impact on the Distribution of Competing Aedes Disease Vectors? Climate 2021, 9, 36. https://0-doi-org.brum.beds.ac.uk/10.3390/cli9020036
AMA Style
Bennett KL, McMillan WO, Loaiza JR. Does Local Adaptation Impact on the Distribution of Competing Aedes Disease Vectors? Climate. 2021; 9(2):36. https://0-doi-org.brum.beds.ac.uk/10.3390/cli9020036
Chicago/Turabian StyleBennett, Kelly L., William Owen McMillan, and Jose R. Loaiza. 2021. "Does Local Adaptation Impact on the Distribution of Competing Aedes Disease Vectors?" Climate 9, no. 2: 36. https://0-doi-org.brum.beds.ac.uk/10.3390/cli9020036
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.