
Plant Species Richness in Multiyear Wet and Dry Periods in the Chihuahuan Desert

 , , , and
, , , and
Abstract
:1. Introduction
1.1. Species Richness Responses
1.2. Precipitation Characteristics
1.3. Legacies
1.4. Dryland Landscapes
2. Methods
2.1. Study Area
2.2. Precipitation Data
2.3. Species Richness Data
2.4. Analysis
2.4.1. Objective 1: Identifying Precipitation Periods and Species Richness Responses
2.4.2. Objective 2: Comparing Daily Precipitation Characteristics and Richness for Periods with Similar Precipitation
2.4.3. Objective 3: Comparing the Sequence of Wet-Dry Periods and Richness over the Record
3. Results
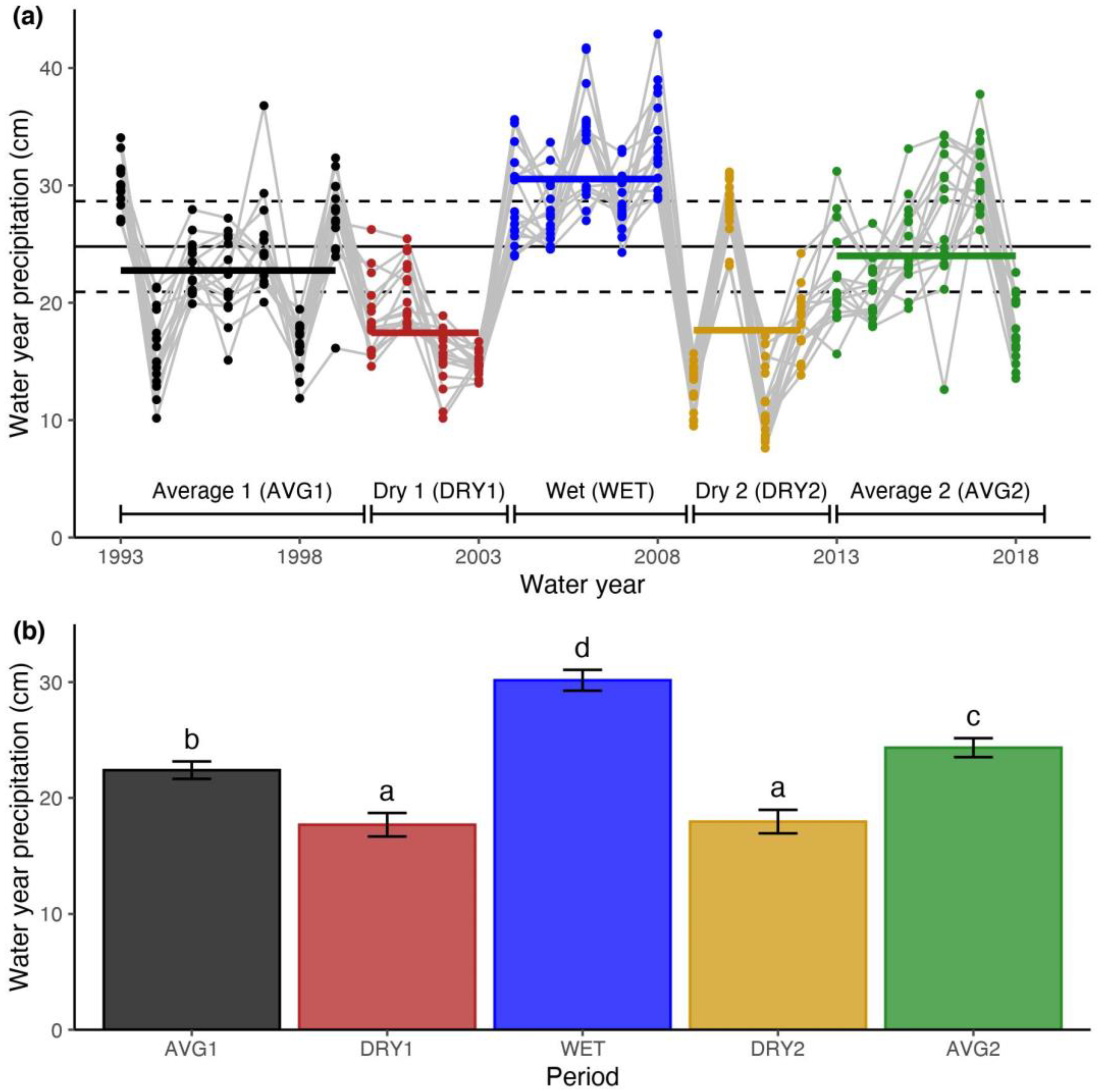
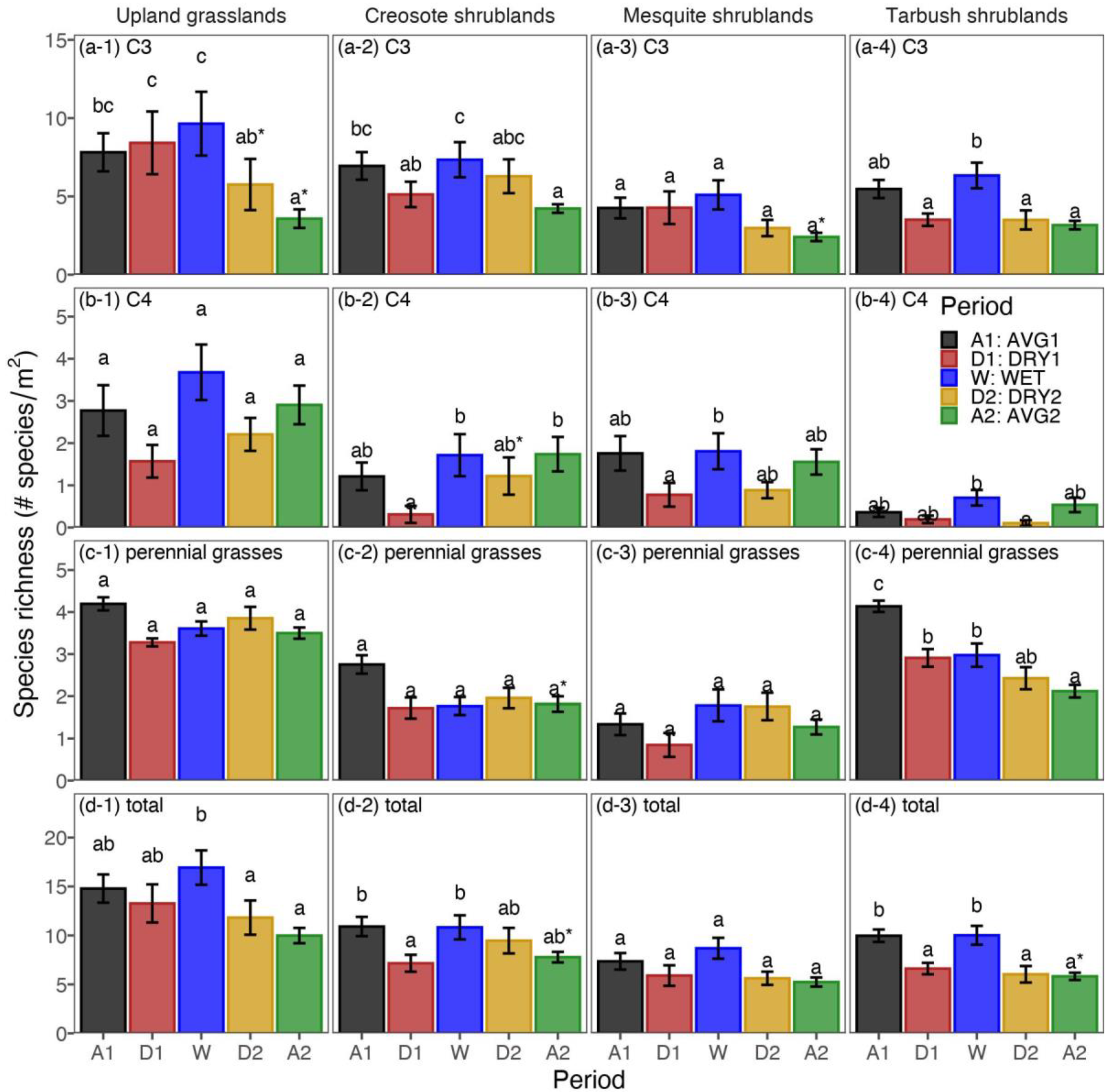
3.1. Objective 1: Identifying Precipitation Periods and Species Richness Responses
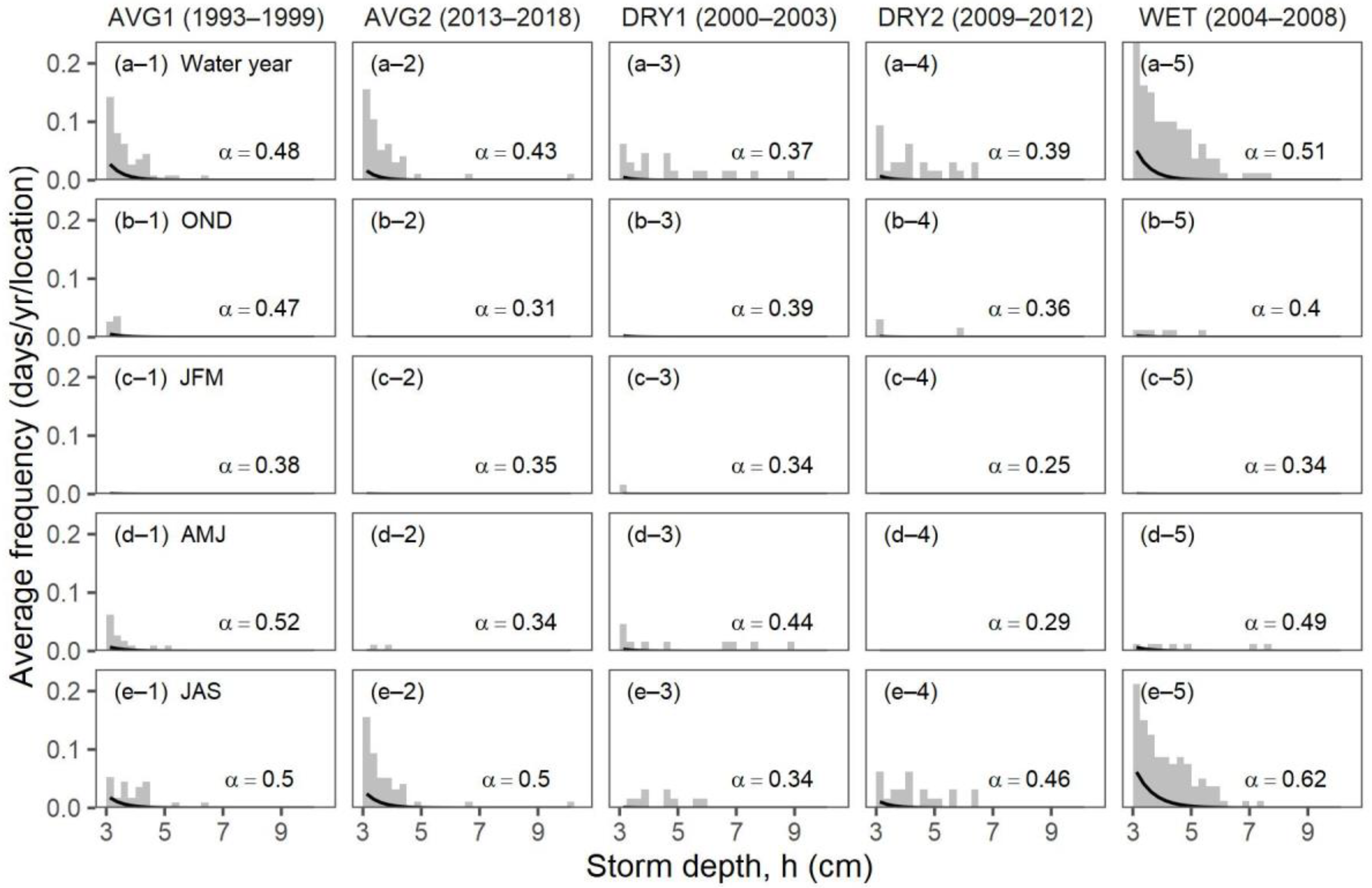
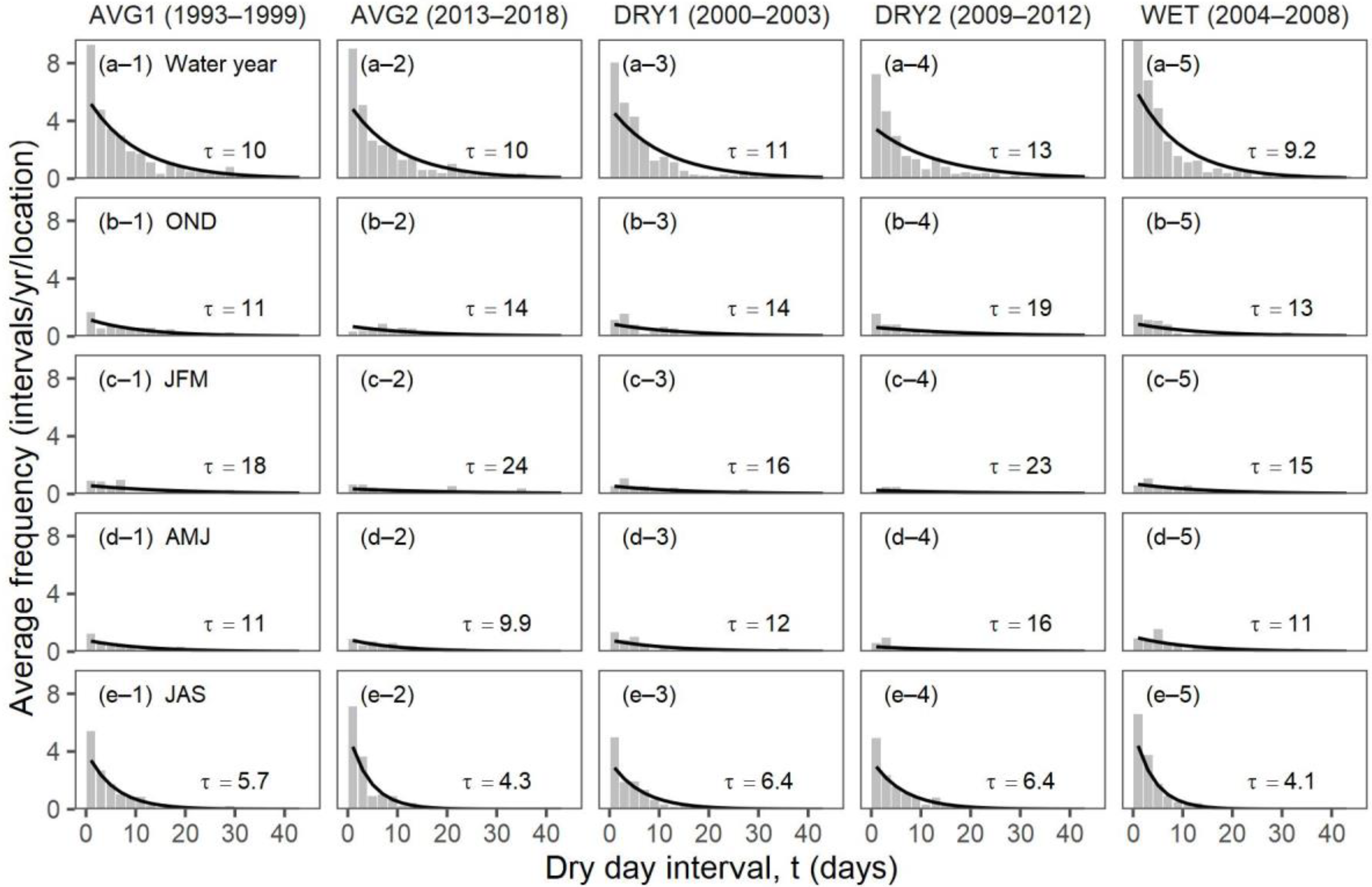
3.2. Objective 2: Comparing Daily Precipitation Characteristics and Richness for Periods with Similar Precipitation
3.3. Objective 3: Comparing the Sequence of Wet-Dry Periods and Richness over the Record
4. Discussion
4.1. Richness and Rainfall Comparisons between Periods
4.2. Importance of Legacies
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Donat, M.G.; Lowry, A.L.; Alexander, L.V.; O’Gorman, P.A.; Maher, N. More Extreme Precipitation in the World’s Dry and Wet Regions. Nat. Clim. Chang. 2016, 6, 508–513. [Google Scholar] [CrossRef]
- Pendergrass, A.G.; Knutti, R.; Lehner, F.; Deser, C.; Sanderson, B.M. Precipitation Variability Increases in a Warmer Climate. Sci. Rep. 2017, 7, 17966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, L.J.; Epstein, H.E.; Li, J.; Okin, G.S. Aeolian Process Effects on Vegetation Communities in an Arid Grassland Ecosystem. Ecol. Evol. 2012, 2, 809–821. [Google Scholar] [CrossRef] [PubMed]
- Bestelmeyer, B.T.; Ellison, A.M.; Fraser, W.R.; Gorman, K.B.; Holbrook, S.J.; Laney, C.M.; Ohman, M.D.; Peters, D.P.C.; Pillsbury, F.C.; Rassweiler, A.; et al. Analysis of Abrupt Transitions in Ecological Systems. Ecosphere 2011, 2, 129. [Google Scholar] [CrossRef]
- Hoover, D.L.; Knapp, A.K.; Smith, M.D. Resistance and Resilience of a Grassland Ecosystem to Climate Extremes. Ecology 2014, 95, 2646–2656. [Google Scholar] [CrossRef] [Green Version]
- Okin, G.S.; Sala, O.E.; Vivoni, E.R.; Zhang, J.; Bhattachan, A. The Interactive Role of Wind and Water in Functioning of Drylands: What Does the Future Hold? Bioscience 2018, 68, 670–677. [Google Scholar] [CrossRef]
- Peters, D.P.C.; Bestelmeyer, B.T.; Herrick, J.E.; Fredrickson, E.L.; Monger, H.C.; Havstad, K.M. Disentangling Complex Landscapes: New Insights into Arid and Semiarid System Dynamics. Bioscience 2006, 56, 491–501. [Google Scholar] [CrossRef] [Green Version]
- Petrie, M.D.; Peters, D.P.C.; Yao, J.; Blair, J.M.; Burruss, N.D.; Collins, S.L.; Derner, J.D.; Gherardi, L.A.; Hendrickson, J.R.; Sala, O.E.; et al. Regional Grassland Productivity Responses to Precipitation during Multiyear Above- and below-Average Rainfall Periods. Glob. Chang. Biol. 2018, 24, 1935–1951. [Google Scholar] [CrossRef]
- Petrie, M.D.; Peters, D.P.C.; Burruss, N.D.; Ji, W.; Savoy, H.M. Differing Climate and Landscape Effects on Regional Dryland Vegetation Responses during Wet Periods Allude to Future Patterns. Glob. Chang. Biol. 2019, 25, 3305–3318. [Google Scholar] [CrossRef]
- Petrie, M.D.; Peters, D.P.C.; Burruss, N.D.; Ji, W.; Savoy, H.M. Local-regional similarity in drylands increases during multiyear wet and dry periods and in response to extreme events. Ecosphere 2019, 10, e02939. [Google Scholar] [CrossRef] [Green Version]
- Schreiner-McGraw, A.; Ajami, H.; Vivoni, E.R. Extreme Weather Events and Transmission Losses in Arid Streams. Environ. Res. Lett. 2019, 14, 084002. [Google Scholar] [CrossRef]
- Smith, M.D. An Ecological Perspective on Extreme Climatic Events: A Synthetic Definition and Framework to Guide Future Research. J. Ecol. 2011, 99, 656–663. [Google Scholar] [CrossRef]
- Estiarte, M.; Vicca, S.; Peñuelas, J.; Bahn, M.; Beier, C.; Emmett, B.A.; Fay, P.A.; Hanson, P.J.; Hasibeder, R.; Kigel, J.; et al. Few Multiyear Precipitation-Reduction Experiments Find a Shift in the Productivity-Precipitation Relationship. Glob. Chang. Biol. 2016, 22, 2570–2581. [Google Scholar] [CrossRef] [Green Version]
- Knapp, A.K.; Chen, A.; Griffin-Nolan, R.J.; Baur, L.E.; Carroll, C.J.W.; Gray, J.E.; Hoffman, A.M.; Li, X.; Post, A.K.; Slette, I.J.; et al. Resolving the Dust Bowl Paradox of Grassland Responses to Extreme Drought. Proc. Natl. Acad. Sci. USA 2020, 117, 22249–22255. [Google Scholar] [CrossRef]
- Peters, D.P.C.; Yao, J.; Sala, O.E.; Anderson, J.P. Directional Climate Change and Potential Reversal of Desertification in Arid and Semiarid Ecosystems. Glob. Chang. Biol. 2012, 18, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Peters, D.P.C.; Yao, J.; Browning, D.; Rango, A. Mechanisms of Grass Response in Grasslands and Shrublands during Dry or Wet Periods. Oecologia 2014, 174, 1323–1334. [Google Scholar] [CrossRef]
- Carroll, C.J.W.; Slette, I.J.; Griffin-Nolan, R.J.; Baur, L.E.; Hoffman, A.M.; Denton, E.M.; Gray, J.E.; Post, A.K.; Johnston, M.K.; Yu, Q.; et al. Is a Drought a Drought in Grasslands? Productivity Responses to Different Types of Drought. Oecologia 2021, 1–10. [Google Scholar] [CrossRef]
- Collins, S.L.; Koerner, S.E.; Plaut, J.A.; Okie, J.G.; Brese, D.; Calabrese, L.B.; Carvajal, A.; Evansen, R.J.; Nonaka, E. Stability of Tallgrass Prairie during a 19-Year Increase in Growing Season Precipitation. Funct. Ecol. 2012, 26, 1450–1459. [Google Scholar] [CrossRef]
- Peters, D.P.C.; Burruss, N.D.; Okin, G.S.; Hatfield, J.L.; Scroggs, S.L.P.; Huang, H.; Brungard, C.W.; Yao, J. Deciphering the Past to Inform the Future: Preparing for the next (“Really Big”) Extreme Event. Front. Ecol. Environ. 2020, 18, 401–408. [Google Scholar] [CrossRef] [Green Version]
- Albertson, F.W.; Weaver, J.E. History of the Native Vegetation of Western Kansas during Seven Years of Continuous Drought. Ecol. Monogr. 1942, 12, 23–51. [Google Scholar] [CrossRef] [Green Version]
- Knapp, A.K.; Beier, C.; Briske, D.D.; Classen, A.T.; Luo, Y.; Reichstein, M.; Smith, M.D.; Smith, S.D.; Bell, J.E.; Fay, P.A.; et al. Consequences of More Extreme Precipitation Regimes for Terrestrial Ecosystems. Bioscience 2008, 58, 811–821. [Google Scholar] [CrossRef]
- Seneviratne, S.I.; Nicholls, N.; Easterling, D.; Goodess, C.M.; Kanae, S.; Kossin, J.; Luo, Y.; Marengo, J.; McInnes, K.; Rahimi, M.; et al. Changes in Climate Extremes and their Impacts on the Natural Physical Environment. In Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation; Field, C.B., Barros, V., Stocker, T.F., Dahe, Q., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 109–230. ISBN 9781139177245. [Google Scholar]
- Schwinning, S.; Sala, O.E. Hierarchy of Responses to Resource Pulses in Arid and Semi-Arid Ecosystems. Oecologia 2004, 141, 211–220. [Google Scholar] [CrossRef]
- Reynolds, J.F.; Kemp, P.R.; Ogle, K.; Fernández, R.J. Modifying the ‘Pulse–Reserve’ Paradigm for Deserts of North America: Precipitation Pulses, Soil Water, and Plant Responses. Oecologia 2004, 141, 194–210. [Google Scholar] [CrossRef]
- Noy-Meir, I. Desert Ecosystems: Environment and Producers. Annu. Rev. Ecol. Syst. 1973, 4, 25–52. [Google Scholar] [CrossRef]
- Sala, O.E.; Gherardi, L.A.; Reichmann, L.; Jobbágy, E.; Peters, D. Legacies of Precipitation Fluctuations on Primary Production: Theory and Data Synthesis. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3135–3144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichmann, L.G.; Sala, O.E.; Peters, D.P.C. Precipitation Legacies in Desert Grassland Primary Production Occur through Previous-Year Tiller Density. Ecology 2013, 94, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Knapp, A.K. Rainfall Variability, Carbon Cycling, and Plant Species Diversity in a Mesic Grassland. Science 2002, 298, 2202–2205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maestre, F.T.; Quero, J.L.; Gotelli, N.J.; Escudero, A.; Ochoa, V.; Delgado-Baquerizo, M.; Garcia-Gomez, M.; Bowker, M.A.; Soliveres, S.; Escolar, C.; et al. Plant Species Richness and Ecosystem Multifunctionality in Global Drylands. Science 2012, 335, 214–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hector, A.; Hautier, Y.; Saner, P.; Wacker, L.; Bagchi, R.; Joshi, J.; Scherer-Lorenzen, M.; Spehn, E.M.; Bazeley-White, E.; Weilenmann, M.; et al. General Stabilizing Effects of Plant Diversity on Grassland Productivity through Population Asynchrony and Overyielding. Ecology 2010, 91, 2213–2220. [Google Scholar] [CrossRef] [PubMed]
- Isbell, F.; Craven, D.; Connolly, J.; Loreau, M.; Schmid, B.; Beierkuhnlein, C.; Bezemer, T.M.; Bonin, C.; Bruelheide, H.; de Luca, E.; et al. Biodiversity Increases the Resistance of Ecosystem Productivity to Climate Extremes. Nature 2015, 526, 574–577. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Downing, J.A. Biodiversity and Stability in Grasslands. Nature 1994, 367, 363–365. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Knops, J.M.H. Biodiversity and Ecosystem Stability in a Decade-Long Grassland Experiment. Nature 2006, 441, 629–632. [Google Scholar] [CrossRef]
- Hector, A.; Schmid, B.; Beierkuhnlein, C.; Caldeira, M.C.; Diemer, M.; Dimitrakopoulos, P.G.; Finn, J.A.; Freitas, H.; Giller, P.S.; Good, J.; et al. Plant Diversity and Productivity Experiments in European Grasslands. Science 1999, 286, 1123–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, K.; Cardinale, B.J.; Fox, J.W.; Gonzalez, A.; Loreau, M.; Wayne Polley, H.; Reich, P.B.; van Ruijven, J. Species Richness and the Temporal Stability of Biomass Production: A New Analysis of Recent Biodiversity Experiments. Am. Nat. 2014, 183, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hautier, Y.; Seabloom, E.W.; Borer, E.T.; Adler, P.B.; Harpole, W.S.; Hillebrand, H.; Lind, E.M.; MacDougall, A.S.; Stevens, C.J.; Bakker, J.D.; et al. Eutrophication Weakens Stabilizing Effects of Diversity in Natural Grasslands. Nature 2014, 508, 521–525. [Google Scholar] [CrossRef]
- Hautier, Y.; Tilman, D.; Isbell, F.; Seabloom, E.W.; Borer, E.T.; Reich, P.B. Anthropogenic Environmental Changes Affect Ecosystem Stability via Biodiversity. Science 2015, 348, 336–340. [Google Scholar] [CrossRef] [Green Version]
- Gherardi, L.A.; Sala, O.E. Enhanced Interannual Precipitation Variability Increases Plant Functional Diversity That in Turn Ameliorates Negative Impact on Productivity. Ecol. Lett. 2015, 18, 1293–1300. [Google Scholar] [CrossRef]
- Griffin-Nolan, R.J.; Carroll, C.J.W.; Denton, E.M.; Johnston, M.K.; Collins, S.L.; Smith, M.D.; Knapp, A.K. Legacy Effects of a Regional Drought on Aboveground Net Primary Production in Six Central US Grasslands. Plant Ecol. 2018, 219, 505–515. [Google Scholar] [CrossRef]
- Frenette-Dussault, C.; Shipley, B.; Léger, J.-F.; Meziane, D.; Hingrat, Y. Functional Structure of an Arid Steppe Plant Community Reveals Similarities with Grime’s C-S-R Theory. J. Veg. Sci. 2012, 23, 208–222. [Google Scholar] [CrossRef]
- Kooyers, N.J. The Evolution of Drought Escape and Avoidance in Natural Herbaceous Populations. Plant Sci. 2015, 234, 155–162. [Google Scholar] [CrossRef]
- Arnone, J.A.; Jasoni, R.L.; Lucchesi, A.J.; Larsen, J.D.; Leger, E.A.; Sherry, R.A.; Luo, Y.; Schimel, D.S.; Verburg, P.S.J. A Climatically Extreme Year Has Large Impacts on C4 Species in Tallgrass Prairie Ecosystems but Only Minor Effects on Species Richness and Other Plant Functional Groups. J. Ecol. 2011, 99, 678–688. [Google Scholar] [CrossRef]
- Gherardi, L.A.; Sala, O.E. Enhanced Precipitation Variability Decreases Grass- and Increases Shrub-Productivity. Proc. Natl. Acad. Sci. USA 2015, 112, 12735–12740. [Google Scholar] [CrossRef] [Green Version]
- Suttle, K.B.; Thomsen, M.A.; Power, M.E. Species Interactions Reverse Grassland Responses to Changing Climate. Science 2007, 315, 640–642. [Google Scholar] [CrossRef] [Green Version]
- Duniway, M.C.; Petrie, M.D.; Peters, D.P.C.; Anderson, J.P.; Crossland, K.; Herrick, J.E. Soil Water Dynamics at 15 Locations Distributed across a Desert Landscape: Insights from a 27-Yr Dataset. Ecosphere 2018, 9, e02335. [Google Scholar] [CrossRef] [Green Version]
- Loik, M.E.; Breshears, D.D.; Lauenroth, W.K.; Belnap, J. A Multi-Scale Perspective of Water Pulses in Dryland Ecosystems: Climatology and Ecohydrology of the Western USA. Oecologia 2004, 141, 269–281. [Google Scholar] [CrossRef]
- Small, E.E. Climatic Controls on Diffuse Groundwater Recharge in Semiarid Environments of the Southwestern United States. Water Resour. Res. 2005, 41, 1–17. [Google Scholar] [CrossRef]
- Grant, K.; Kreyling, J.; Dienstbach, L.F.H.; Beierkuhnlein, C.; Jentsch, A. Water Stress Due to Increased Intra-Annual Precipitation Variability Reduced Forage Yield but Raised Forage Quality of a Temperate Grassland. Agric. Ecosyst. Environ. 2014, 186, 11–22. [Google Scholar] [CrossRef]
- Grant, K.; Kreyling, J.; Beierkuhnlein, C.; Jentsch, A. Importance of Seasonality for the Response of a Mesic Temperate Grassland to Increased Precipitation Variability and Warming. Ecosystems 2017, 20, 1454–1467. [Google Scholar] [CrossRef]
- Gibbens, R.P.; Lenz, J.M. Root Systems of Some Chihuahuan Desert Plants. J. Arid Environ. 2001, 49, 221–263. [Google Scholar] [CrossRef]
- Kurc, S.A.; Small, E.E. Dynamics of Evapotranspiration in Semiarid Grassland and Shrubland Ecosystems during the Summer Monsoon Season, Central New Mexico. Water Resour. Res. 2004, 40. [Google Scholar] [CrossRef]
- Felton, A.J.; Slette, I.J.; Smith, M.D.; Knapp, A.K. Precipitation Amount and Event Size Interact to Reduce Ecosystem Functioning during Dry Years in a Mesic Grassland. Glob. Chang. Biol. 2020, 26, 658–668. [Google Scholar] [CrossRef]
- Porporato, A.; Laio, F.; Ridolfi, L.; Caylor, K.K.; Rodriguez-Iturbe, I. Soil Moisture and Plant Stress Dynamics along the Kalahari Precipitation Gradient. J. Geophys. Res. Atmos. 2003, 108, 1–8. [Google Scholar] [CrossRef]
- Rodriguez-Iturbe, I.; Porporato, A.; Laio, F.; Ridolfi, L. Plants in Water-Controlled Ecosystems: Active Role in Hydrologic Processes and Response to Water Stress. Adv. Water Resour. 2001, 24, 695–705. [Google Scholar] [CrossRef]
- Fernandez-Illescas, C.P.; Porporato, A.; Laio, F.; Rodriguez-Iturbe, I. The Ecohydrological Role of Soil Texture in a Water-Limited Ecosystem. Water Resour. Res. 2001, 37, 2863–2872. [Google Scholar] [CrossRef]
- Monger, C.; Sala, O.E.; Duniway, M.C.; Goldfus, H.; Meir, I.A.; Poch, R.M.; Throop, H.L.; Vivoni, E.R. Legacy Effects in Linked Ecological–Soil–Geomorphic Systems of Drylands. Front. Ecol. Environ. 2015, 13, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Reichmann, L.G.; Sala, O.E. Differential Sensitivities of Grassland Structural Components to Changes in Precipitation Mediate Productivity Response in a Desert Ecosystem. Funct. Ecol. 2014, 28, 1292–1298. [Google Scholar] [CrossRef] [Green Version]
- Thomey, M.L.; Collins, S.L.; Vargas, R.; Johnson, J.E.; Brown, R.F.; Natvig, D.O.; Friggens, M.T. Effect of Precipitation Variability on Net Primary Production and Soil Respiration in a Chihuahuan Desert Grassland. Glob. Chang. Biol. 2011, 17, 1505–1515. [Google Scholar] [CrossRef]
- Gherardi, L.A.; Sala, O.E. Effect of Interannual Precipitation Variability on Dryland Productivity: A Global Synthesis. Glob. Chang. Biol. 2019, 25, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Heisler-White, J.L.; Blairr, J.M.; Kelly, E.F.; Harmoney, K.; Knapp, A.K. Contingent Productivity Responses to More Extreme Rainfall Regimes across a Grassland Biome. Glob. Chang. Biol. 2009, 15, 2894–2904. [Google Scholar] [CrossRef]
- IPCC. Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change; Masson-Delmotte, V., Zhai, P., Pörtner, H.-O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Eds.; World Meteorological Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Gibbens, R.P.; McNeely, R.P.; Havstad, K.M.; Beck, R.F.; Nolen, B. Vegetation Changes in the Jornada Basin from 1858 to 1998. J. Arid Environ. 2005, 61, 651–668. [Google Scholar] [CrossRef]
- Huenneke, L.F.; Anderson, J.P.; Remmenga, M.; Schlesinger, W.H. Desertification Alters Patterns of Aboveground Net Primary Production in Chihuahuan Ecosystems. Glob. Chang. Biol. 2002, 8, 247–264. [Google Scholar] [CrossRef]
- Li, J.; Okin, G.S.; Alvarez, L.; Epstein, H. Effects of Wind Erosion on the Spatial Heterogeneity of Soil Nutrients in Two Desert Grassland Communities. Biogeochemistry 2008, 88, 73–88. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Reynolds, J.F.; Cunningham, G.L.; Huenneke, L.F.; Jarrell, W.M.; Virginia, R.A.; Whitford, W.G. Biological Feedbacks in Global Desertification. Science 1990, 247, 1043–1048. [Google Scholar] [CrossRef] [Green Version]
- Pierce, N.A.; Archer, S.R.; Bestelmeyer, B.T.; James, D.K. Grass-Shrub Competition in Arid Lands: An Overlooked Driver in Grassland–Shrubland State Transition? Ecosystems 2019, 22, 619–628. [Google Scholar] [CrossRef]
- Manning, S.J.; Barbour, M.G. Root Systems, Spatial Patterns, and Competition for Soil Moisture between Two Desert Subshrubs. Am. J. Bot. 1988, 75, 885–893. [Google Scholar] [CrossRef]
- Fredrickson, E.; Havstad, K.M.; Estell, R.; Hyder, P. Perspectives on Desertification: South-Western United States. J. Arid Environ. 1998, 39, 191–207. [Google Scholar] [CrossRef]
- Anderson, J.P. LTER Weather Station Daily Summary Climate Data Ver 113. Available online: https://0-doi-org.brum.beds.ac.uk/10.6073/pasta/c13106744e57bae0f626bba25b635e0a (accessed on 12 August 2020).
- Anderson, J.P. Precipitation Data from a Standard Can Rain Gauge at the LTER Weather Station, Jornada Basin, Southern New Mexico, USA, 1992-Ongoing Ver 100. Available online: https://0-doi-org.brum.beds.ac.uk/10.6073/pasta/559d8d1fa1c542b8ad8088b65b48081b (accessed on 12 August 2020).
- Yao, J.; Anderson, J.P.; Savoy, H.; Peters, D.P.C. Gap-Filled Daily Precipitation at the 15 Long-Term NPP Sites at Jornada Basin LTER, 1980-Ongoing Ver 75. Available online: https://0-doi-org.brum.beds.ac.uk/10.6073/pasta/cf3c45e5480551453f1f9041d664a28f (accessed on 12 August 2020).
- Huenneke, L.F.; Clason, D.; Muldavin, E. Spatial Heterogeneity in Chihuahuan Desert Vegetation: Implications for Sampling Methods in Semi-Arid Ecosystems. J. Arid Environ. 2001, 47, 257–270. [Google Scholar] [CrossRef]
- Peters, D.P.C.; Huenneke., L.F. Seasonal Non-Destructive Vegetation Measurements at 15 Net Primary Production (NPP) Study Sites at Jornada Basin LTER, 1989-Ongoing Ver 106. Available online: https://0-doi-org.brum.beds.ac.uk/10.6073/pasta/18dad6748af96c98b72cea3436bf7fe4 (accessed on 16 December 2020).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. Linear and nonlinear mixed effects models. R Package Version 2019, 3, 1–89. [Google Scholar]
- Lenth, R. Emmeans: Estimated marginal means, aka least-squares means. R Package Version 1.4 2018, 1, 3. [Google Scholar]
- Saros, J.E.; Anderson, N.J.; Juggins, S.; McGowan, S.; Yde, J.C.; Telling, J.; Bullard, J.E.; Yallop, M.L.; Heathcote, A.J.; Burpee, B.T.; et al. Arctic Climate Shifts Drive Rapid Ecosystem Responses across the West Greenland Landscape. Environ. Res. Lett. 2019, 14, 074027. [Google Scholar] [CrossRef]
- Xu, L.; Yu, G.; Zhang, W.; Tu, Z.; Tan, W. Change Features of Time-Series Climate Variables from 1962 to 2016 in Inner Mongolia, China. J. Arid Land 2020, 12, 58–72. [Google Scholar] [CrossRef] [Green Version]
- Browning, D.M.; Maynard, J.J.; Karl, J.W.; Peters, D.C. Breaks in MODIS Time Series Portend Vegetation Change: Verification Using Long-Term Data in an Arid Grassland Ecosystem. Ecol. Appl. 2017, 27, 1677–1693. [Google Scholar] [CrossRef]
- Zeileis, A.; Leisch, F.; Hornik, K.; Kleiber, C. Strucchange: An R Package for Testing for Structural Change in Linear Regression Models. J. Stat. Softw. 2002, 7, 1–38. [Google Scholar] [CrossRef] [Green Version]
- Zeileis, A.; Kleiber, C.; Krämer, W.; Hornik, K. Testing and Dating of Structural Changes in Practice. Comput. Stat. Data Anal. 2003, 44, 109–123. [Google Scholar] [CrossRef] [Green Version]
- Porporato, A.; Daly, E.; Rodriguez-Iturbe, I. Soil Water Balance and Ecosystem Response to Climate Change. Am. Nat. 2004, 164, 625. [Google Scholar] [CrossRef]
- Delignette-Muller, M.L.; Dutang, C. fitdistrplus: An R Package for Fitting Distributions. J. Stat. Softw. 2015, 64, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Sala, O.E.; Lauenroth, W.K. Small Rainfall Events: An Ecological Role in Semiarid Regions. Oecologia 1982, 53, 301–304. [Google Scholar] [CrossRef]
- Sala, O.E.; Lauenroth, W.K.; Parton, W.J. Long-Term Soil Water Dynamics in the Shortgrass Steppe. Ecology 1992, 73, 1175–1181. [Google Scholar] [CrossRef]
- Vivoni, E.R.; Moreno, H.A.; Mascaro, G.; Rodriguez, J.C.; Watts, C.J.; Garatuza-Payan, J.; Scott, R.L. Observed Relation between Evapotranspiration and Soil Moisture in the North American Monsoon Region. Geophys. Res. Lett. 2008, 35, L22403. [Google Scholar] [CrossRef] [Green Version]
- Sala, O.E.; Parton, W.J.; Joyce, L.A.; Lauenroth, W.K. Primary Production of the Central Grassland Region of the United States. Ecology 1988, 69, 40–45. [Google Scholar] [CrossRef]
- Mulhouse, J.M.; Hallett, L.M.; Collins, S.L. The Influence of Seasonal Precipitation and Grass Competition on 20 Years of Forb Dynamics in Northern Chihuahuan Desert Grassland. J. Veg. Sci. 2017, 28, 250–259. [Google Scholar] [CrossRef]
- Notaro, M.; Liu, Z.; Gallimore, R.G.; Williams, J.W.; Gutzler, D.S.; Collins, S. Complex Seasonal Cycle of Ecohydrology in the Southwest United States. J. Geophys. Res. 2010, 115, G04034. [Google Scholar] [CrossRef]
- Hottenstein, J.D.; Ponce-Campos, G.E.; Moguel-Yanes, J.; Moran, M.S. Impact of Varying Storm Intensity and Consecutive Dry Days on Grassland Soil Moisture. J. Hydrometeorol. 2015, 16, 106–117. [Google Scholar] [CrossRef] [Green Version]
- Sala, O.E.; Gherardi, L.A.; Peters, D.P.C. Enhanced Precipitation Variability Effects on Water Losses and Ecosystem Functioning: Differential Response of Arid and Mesic Regions. Clim. Chang. 2015, 131, 213–227. [Google Scholar] [CrossRef] [Green Version]
- Coffin, D.P.; Lauenroth, W.K. A Gap Dynamics Simulation Model of Succession in a Semiarid Grassland. Ecol. Modell. 1990, 49, 229–266. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
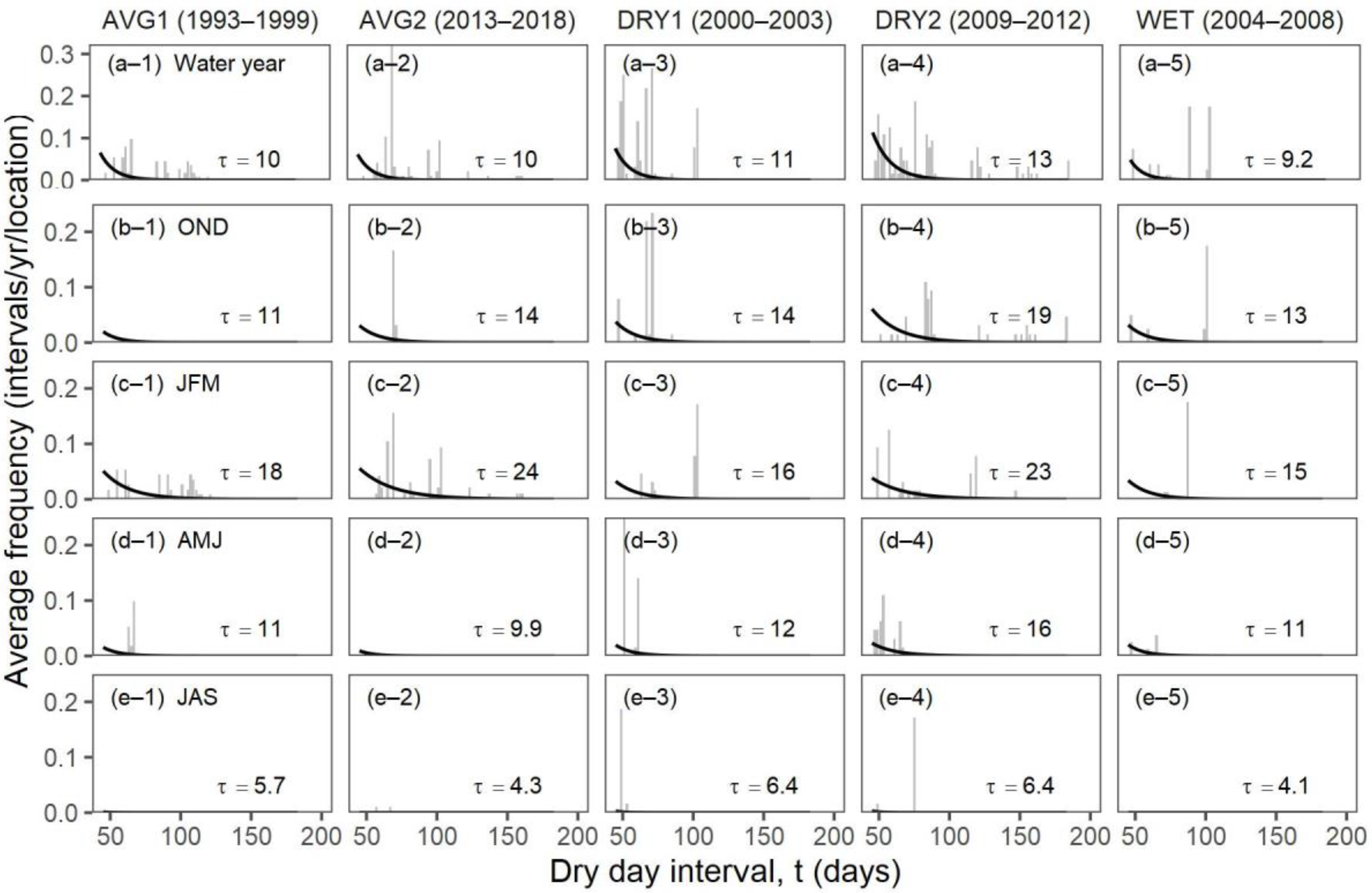
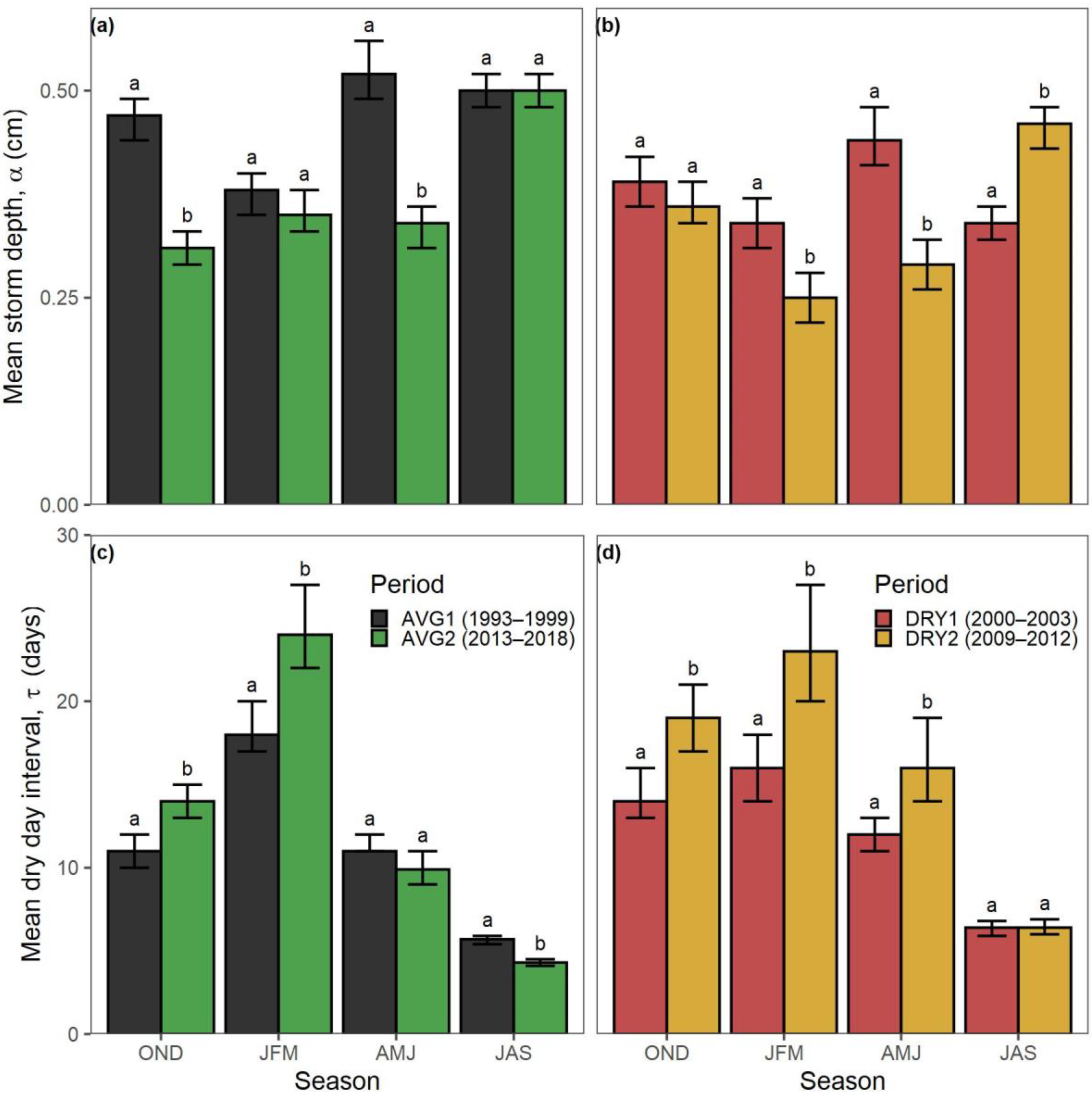
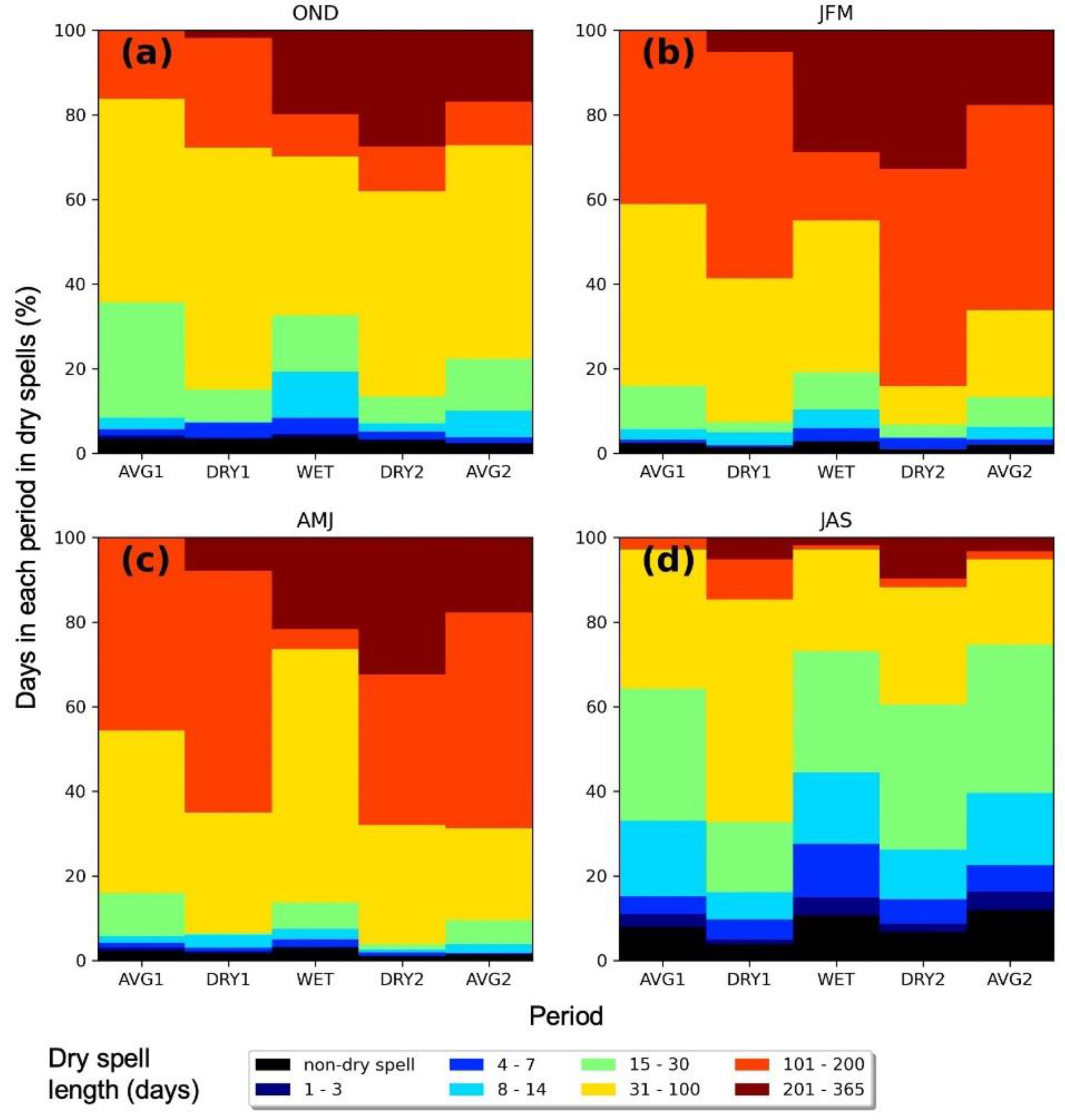
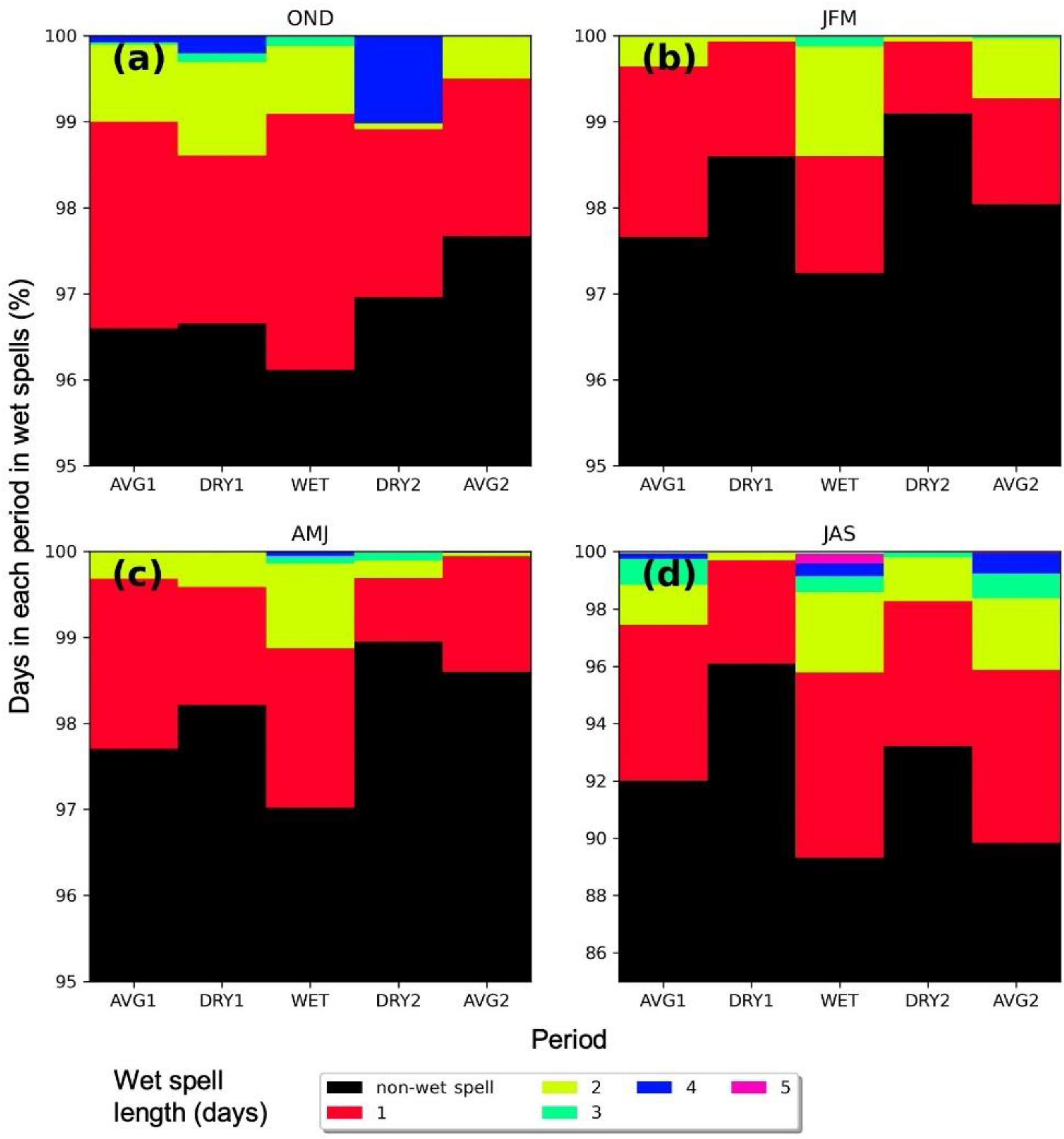
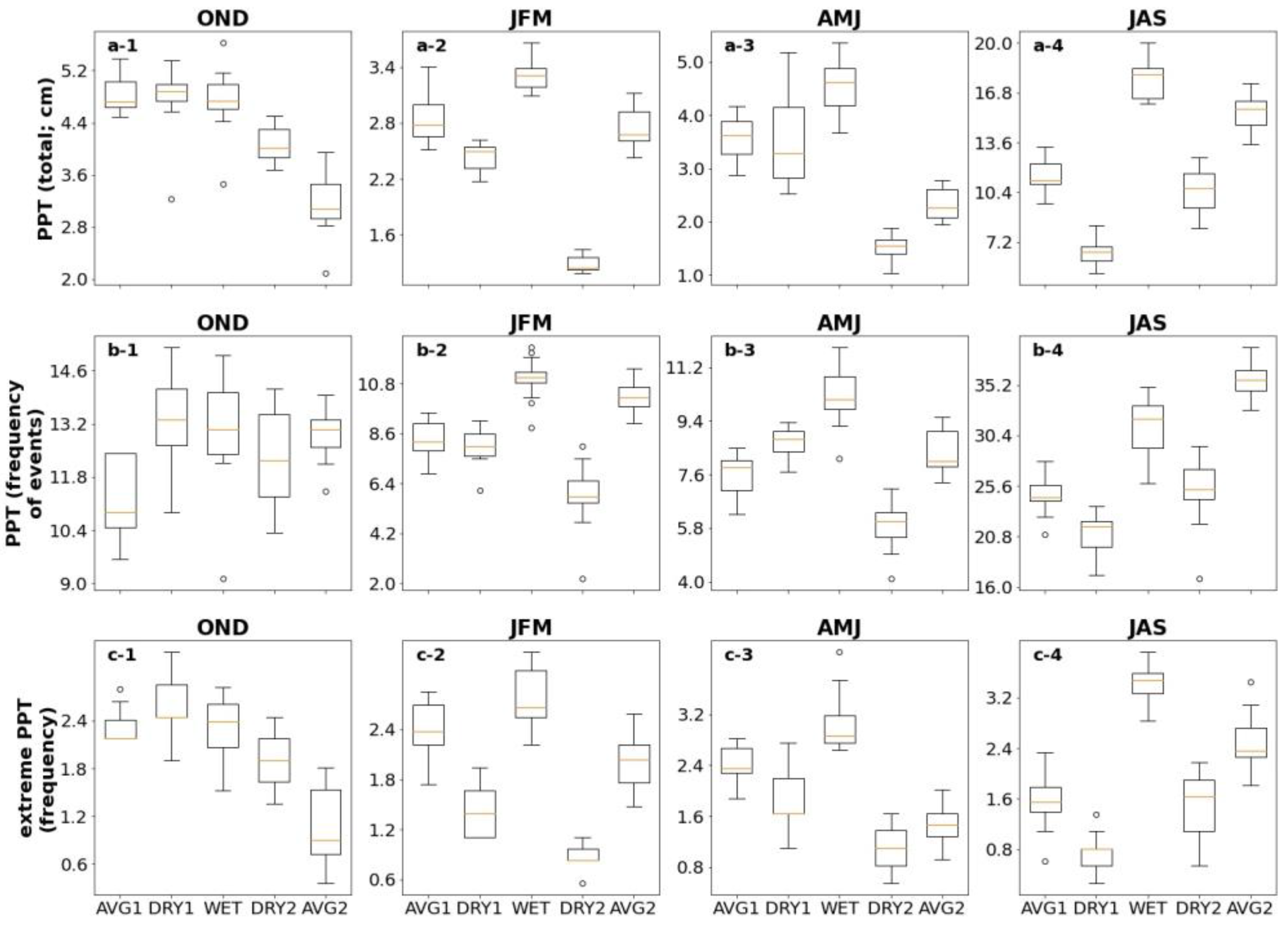
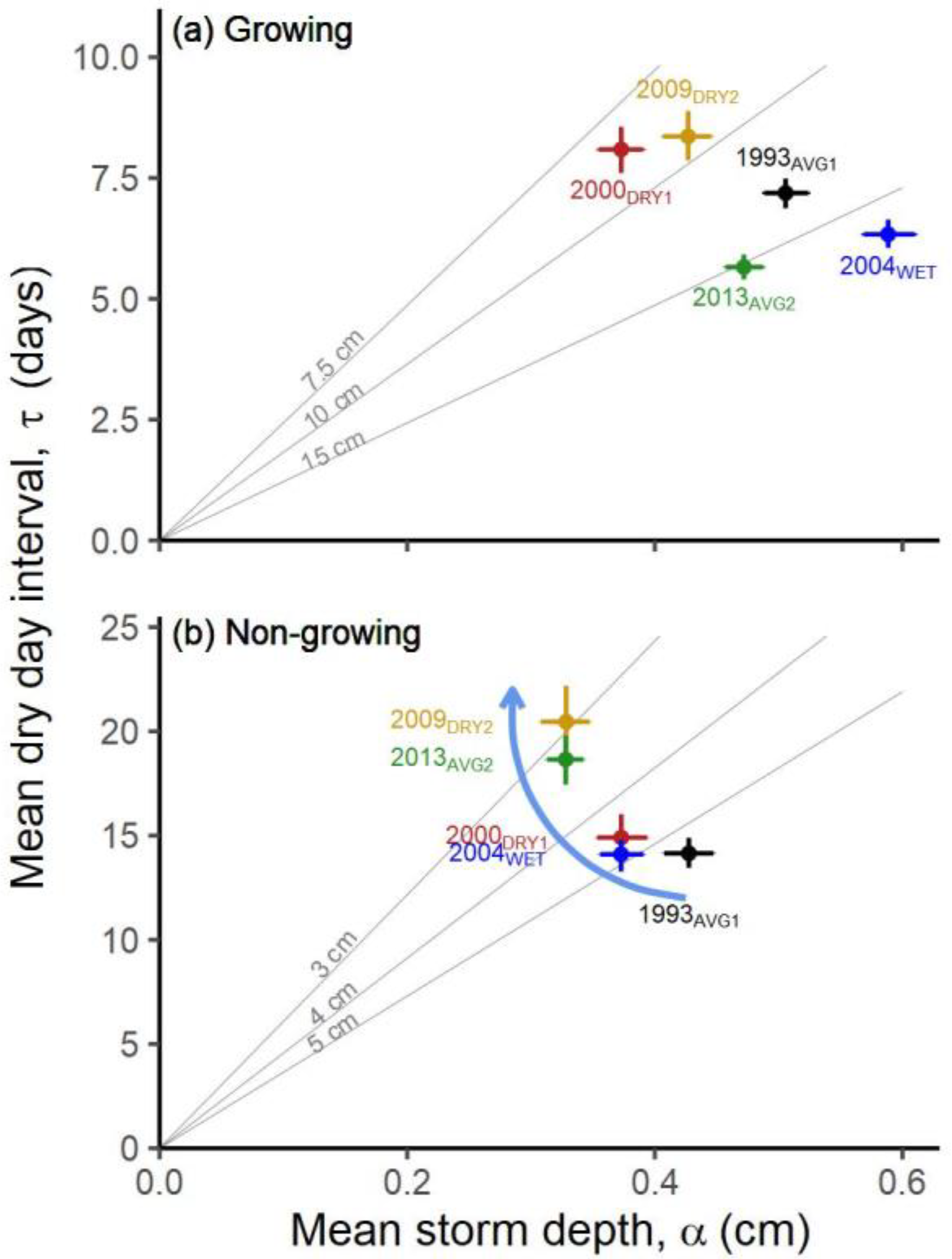
| Ecosystem Type | Functional Group (Relevant Season) | Richness 1 [Figure 2] | Mean Storm Depth (Alpha) [Figure 7a,b] | Mean Dry-Day Interval (Tau) [Figure 7c,d] | Dry Spell Length (31–200 Days) [Figure 8] | Wet Spell Length (>2 Days) [Figure 9] | Total PPT 2 [Figure 10a] | PPT 2 Frequency [Figure 10b] | Extreme PPT 2 Frequency [Figure 10c] |
|---|---|---|---|---|---|---|---|---|---|
| Upland grasslands | C3 (AMJ) | A1 > A2 D1 > D2 | A1 > A2 D1 > D2 | D1 < D2 | A1 < A2 D1 > D2 | A1 > A2 D1 > D2 | D1 > D2 | A1 > A2 D1 > D2 | |
| C4 (JAS) | |||||||||
| Perennial grasses (JAS) | |||||||||
| Creosote Shrublands | C3 (AMJ) | A1 > A2 | A1 > A2 | A1 < A2 | A1 > A2 | A1 > A2 | |||
| C4 (JAS) | D1 < D2 | D1 < D2 | D1 > D2 | D1< D2 | D1 < D2 | D1 < D2 | D1 < D2 | ||
| Perennial grasses (JAS) | A1 > A2 | A1 > A2 | A1 > A2 | A1 < A2 | A1 < A2 | A1 < A2 | A1 < A2 | ||
| Mesquite Shrublands | C3 (AMJ) | A1 > A2 | A1 > A2 | A1 < A2 | A1 > A2 | A1 > A2 | |||
| C4 (JAS) | |||||||||
| Perennial grasses (JAS) | |||||||||
| Tarbush Shrublands | C3 (AMJ) | ||||||||
| C4 (JAS) | |||||||||
| Perennial grasses (JAS) | A1 > A2 | A1 > A2 | A1 > A2 | A1 < A2 | A1 < A2 | A1 < A2 | A1 < A2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peters, D.P.C.; Savoy, H.M.; Stillman, S.; Huang, H.; Hudson, A.R.; Sala, O.E.; Vivoni, E.R. Plant Species Richness in Multiyear Wet and Dry Periods in the Chihuahuan Desert. Climate 2021, 9, 130. https://0-doi-org.brum.beds.ac.uk/10.3390/cli9080130
Peters DPC, Savoy HM, Stillman S, Huang H, Hudson AR, Sala OE, Vivoni ER. Plant Species Richness in Multiyear Wet and Dry Periods in the Chihuahuan Desert. Climate. 2021; 9(8):130. https://0-doi-org.brum.beds.ac.uk/10.3390/cli9080130
Chicago/Turabian StylePeters, Debra P. C., Heather M. Savoy, Susan Stillman, Haitao Huang, Amy R. Hudson, Osvaldo E. Sala, and Enrique R. Vivoni. 2021. "Plant Species Richness in Multiyear Wet and Dry Periods in the Chihuahuan Desert" Climate 9, no. 8: 130. https://0-doi-org.brum.beds.ac.uk/10.3390/cli9080130