
Mechanism of the Effect of High-Intensity Training on Urinary Metabolism in Female Water Polo Players Based on UHPLC-MS Non-Targeted Metabolomics Technique


Abstract
:1. Introduction
2. Materials and Methods
Subjects and Grouping
3. Training Program
3.1. Experimental Grouping and Sampling
3.2. Detection Method
Reagents and Instrument
3.3. Chromatography-Mass Spectrometry Analysis
3.3.1. Chromatographic Conditions
3.3.2. Q-TOF Mass Spectrometry Conditions
3.3.3. LC-MS Data Analysis
4. Results
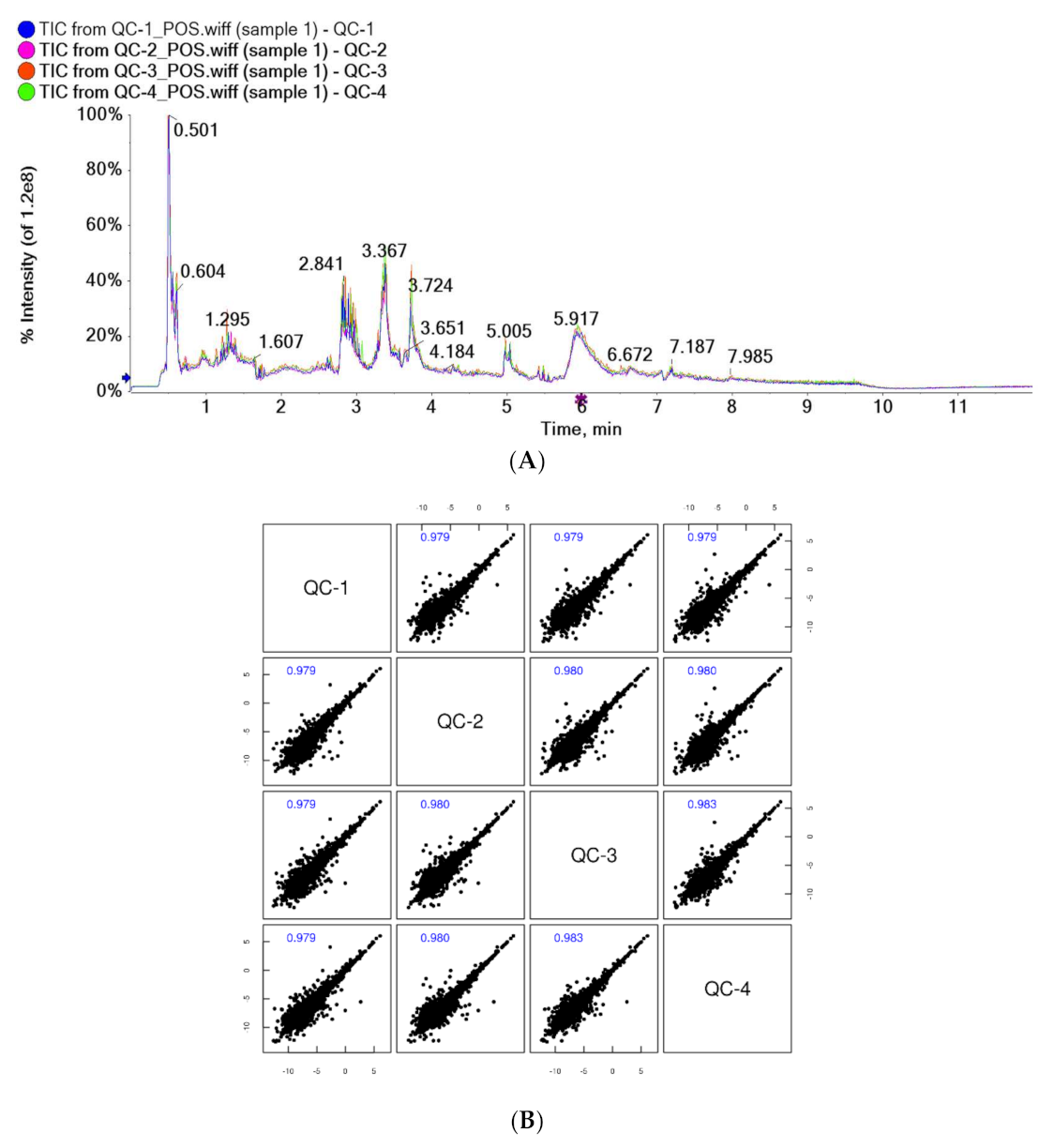
4.1. Stability Evaluation of Subjects’ Urine LC-MS Assay System
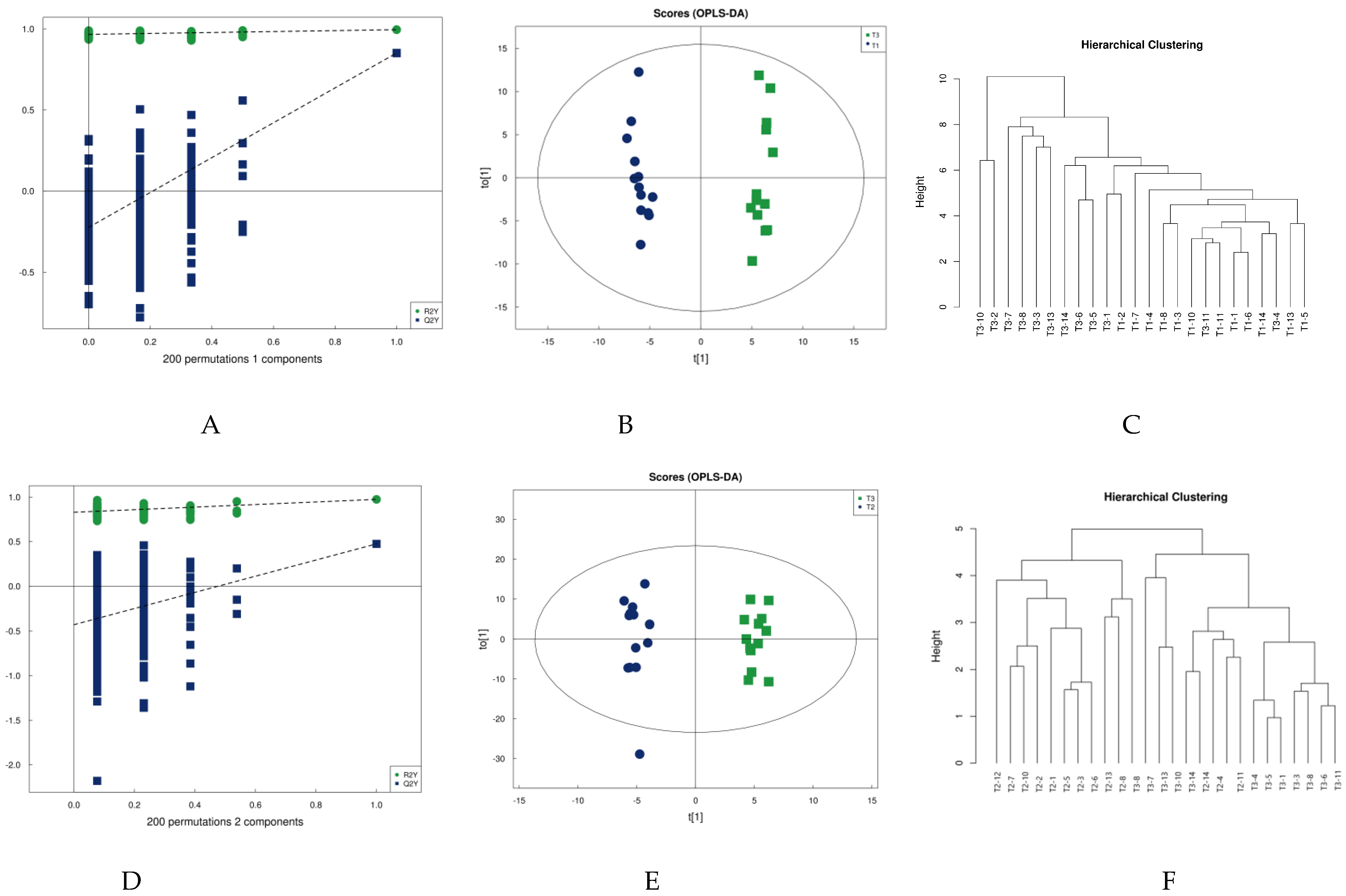
4.2. Multivariate Statistical Analysis
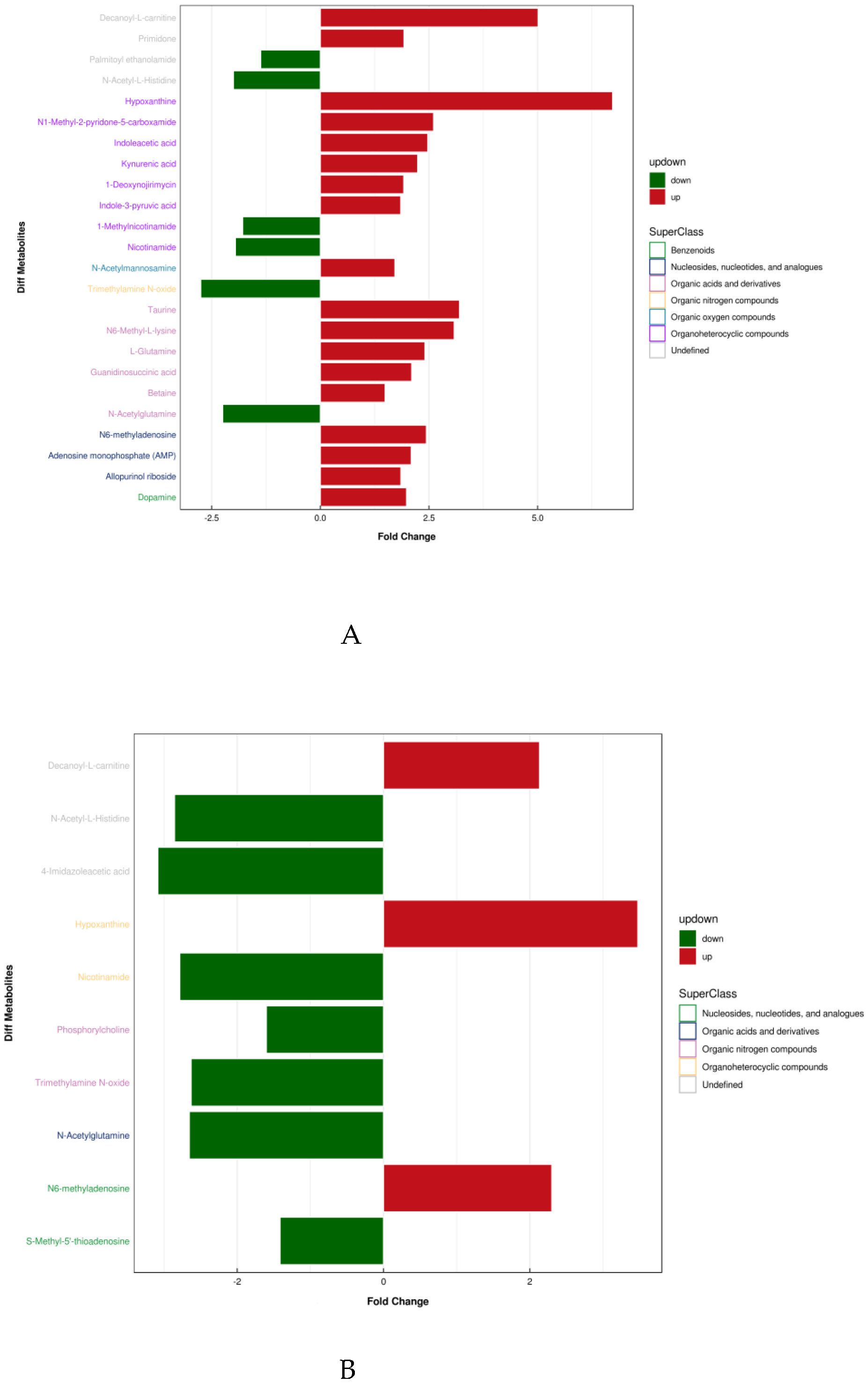
4.3. Screening of Differential Metabolites in Urine by LC-MS
4.4. Analysis of Differential Metabolites and Screening of Potential Biomarkers in Urine after Water Polo Games
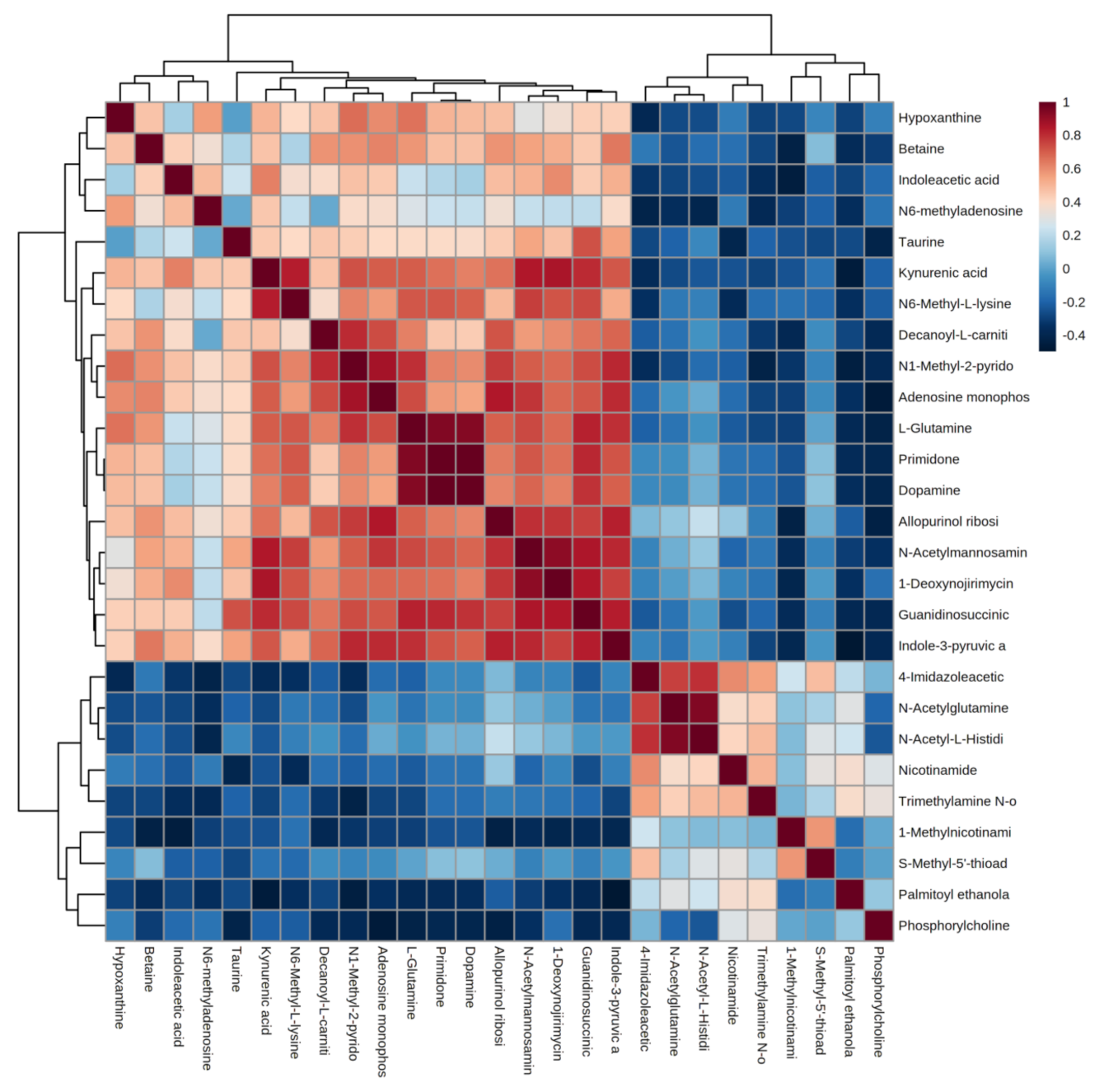
4.4.1. Correlation Analysis of Differential Metabolites
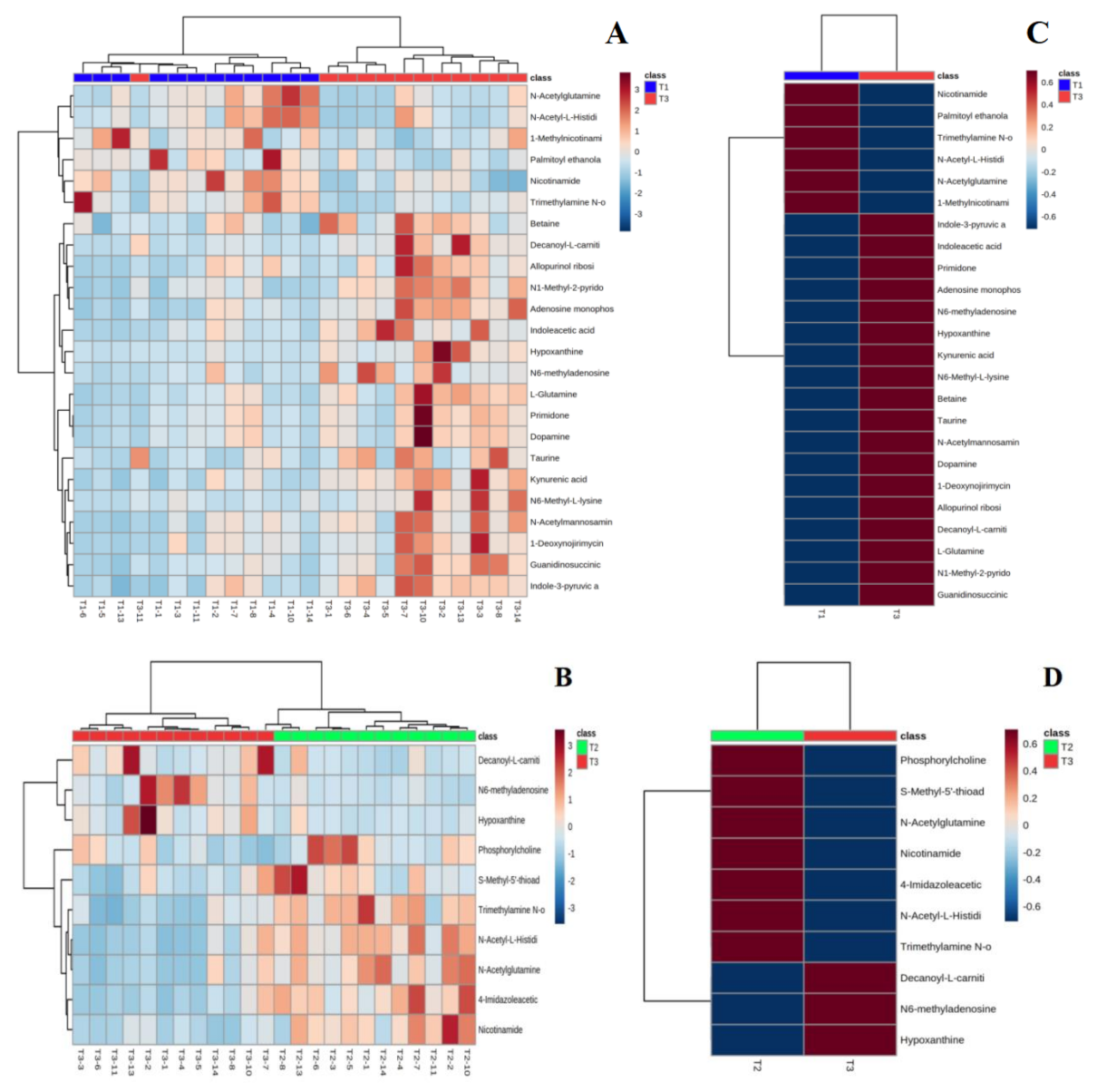
4.4.2. Hierarchical Clustering of Differential Metabolites
4.5. Analysis of the Metabolic Pathway of Urinary Differentials by Water Polo Exercise
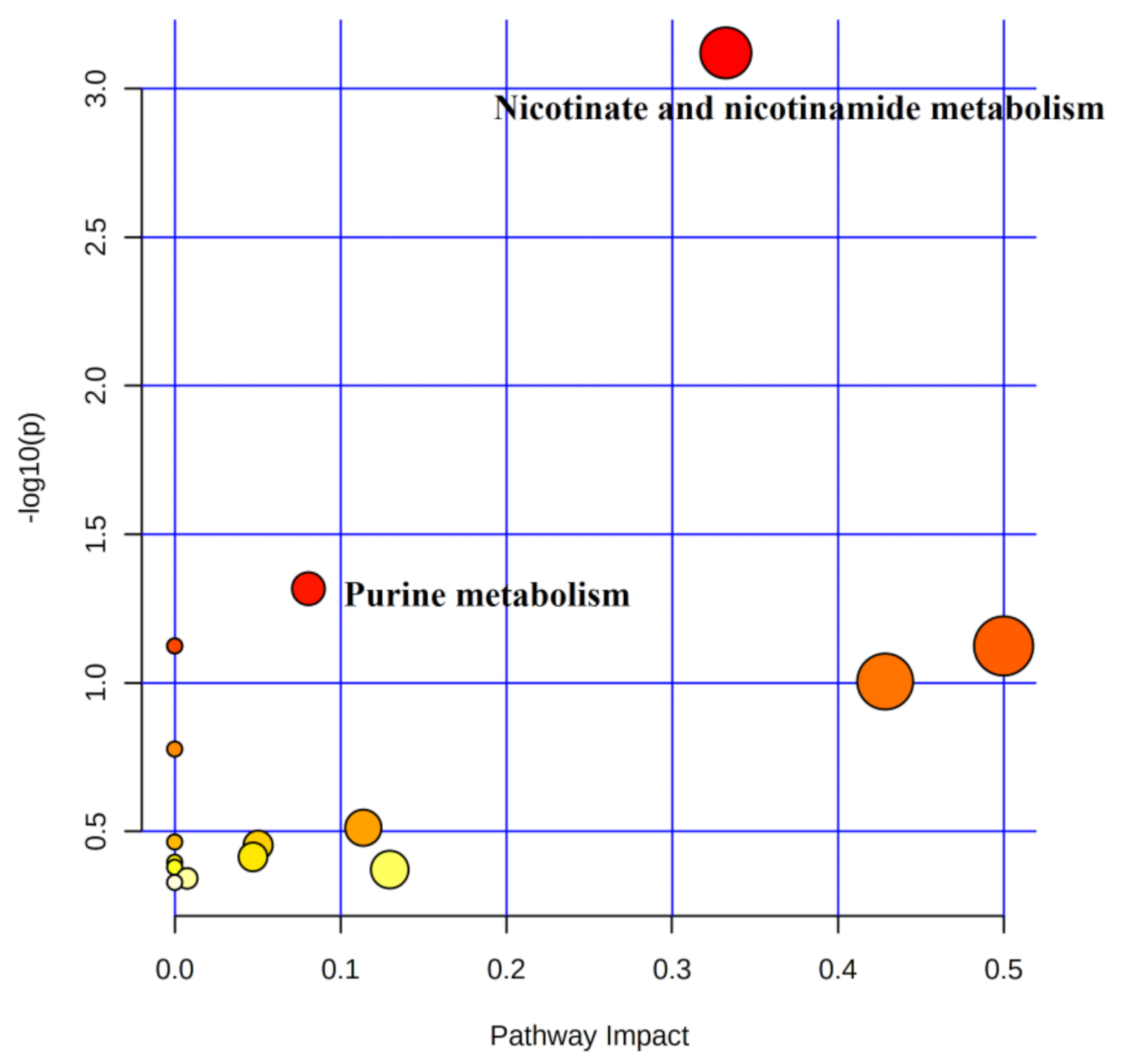
4.5.1. Metabolic Pathway Analysis of Urinary Differential Metabolites between Pre-Training and Immediate Post-Competition
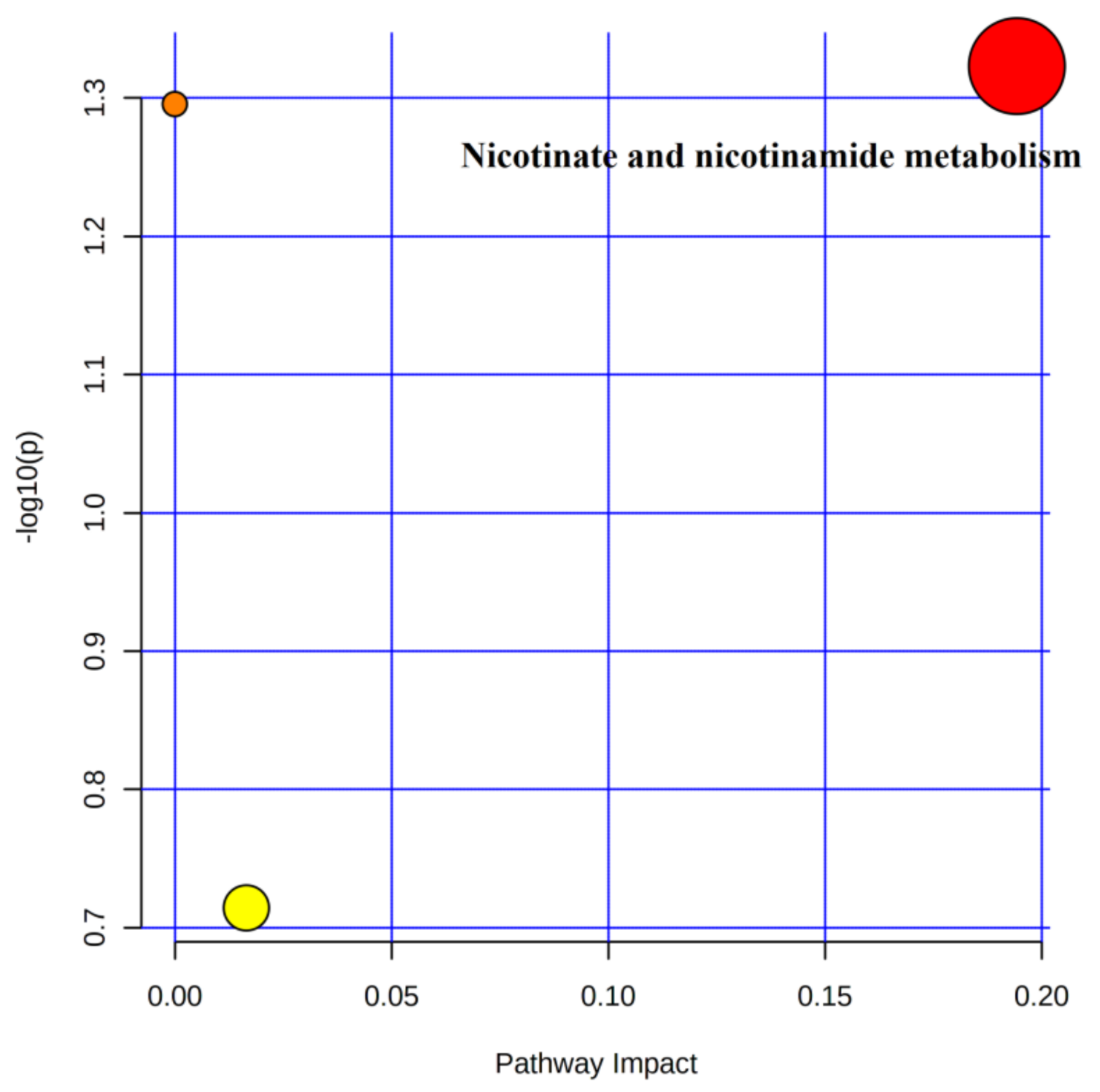
4.5.2. Metabolic Pathway Analysis of Urinary Differential Metabolites in the Immediate Post-Training and Post-Competition Periods
5. Discussion
5.1. Pathway Analysis of Purine Metabolism
5.2. Analysis of Niacin and Nicotinamide Metabolism
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. Metabonomics: Understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Azad, R.K.; Shulaev, V. Metabolomics technology and bioinformatics for precision medicine. Brief. Bioinform. 2019, 20, 1957–1971. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.H.; Chang, C.C.; Wang, L.J.; Chen, W.C.; Fan, S.Y.; Zang, C.Z.; Wang, D.Y. A Multi-analyte Lc-ms/ms Method for Screening and Quantification of Nitrosamines in Sartans. J. Food Drug Anal. 2020, 28, 98–107. [Google Scholar] [CrossRef]
- Kuwada, K.; Kawase, S.; Nakata, K.; Shinya, N.; Narukawa, Y.; Fuchino, H.; Kiuchi, F. LC-MS analysis of saponins of Achyranthes root in the Japanese market. J. Technol. Sci. 2020, 74, 135–141. [Google Scholar] [CrossRef]
- Häkkinen, M.R.; Murtola, T.; Voutilainen, R.; Poutanen, M.; Linnanen, T.; Koskivuori, J.; Auriola, S. Simultaneous analysis by LC–MS/MS of 22 ketosteroids with hydroxylamine derivatization and underivatized estradiol from human plasma, serum and prostate tissue. J. Pharm. Biomed. Anal. 2018, 164, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Berton, R.; Conceição, M.S.; Libardi, C.A.; Canevarolo, R.R.; Gáspari, A.F.; Chacon-Mikahil, M.P.T.; Cavaglieri, C.R. Metabolic time-course response after resistance exercise: A metabolomics approach. J. Sports Sci. 2017, 35, 1211–1218. [Google Scholar] [CrossRef]
- Santone, C.; Dinallo, V.; Paci, M.; D’Ottavio, S.; Barbato, G.; Bernardini, S. Saliva metabolomics by NMR for the evaluation of sport performance. J. Pharm. Biomed. Anal. 2014, 88, 441–446. [Google Scholar] [CrossRef]
- Iñigo, S. Blood biomarkers in sports medicine and performance and the future of metabolomics. In High-Throughput Metabolomics Humana; Springer: New York, NY, USA, 2019; pp. 431–446. [Google Scholar]
- Rundqvist, H.; Veliça, P.; Barbieri, L.; Gameiro, P.A.; Bargiela, D.; Gojkovic, M.; Johnson, R.S. Cytotoxic T-cells mediate exercise-induced reductions in tumor growth. eLife 2020, 9, e59996. [Google Scholar] [CrossRef]
- Oertzen-Hagemann, V.; Kirmse, M.; Eggers, B.; Pfeiffer, K.; Marcus, K.; de Marées, M.; Platen, P. Effects of 12 Weeks of Hypertrophy Resistance Exercise Training Combined with Collagen Peptide Supplementation on the Skeletal Muscle Proteome in Recreationally Active Men. Nutrients 2019, 11, 1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu Lijun, W.T. Metabolomics of serum in rats by fatigue exercise and astaxanthin supplementation. J. Shandong Sport Univ. 2020, 36, 62–70. [Google Scholar]
- Li, J.H.; Wang, Z.H.; Zhu, X.J.; GE, Y.J. A comparison of metabolomical and traditional biochemical methods used for athletic talent identification. J. Phys. Educ. 2013, 20, 123–128. [Google Scholar]
- Xue, H.Y. Mechanism of Aerobic Exercise Intervention on Insulin Resistance in Rats Based on Pancreatic Mechanism of Aerobic Exercise Intervention on Insulin Resistance in Rats Based on Pancreatic Metabonomics. J. Beijing Sport Univ. 2018, 41, 63–69. [Google Scholar]
- Xinming, G. Study on the Characteristics of Human Serum Metabolites before and after Intermittent High-intensity Exercise. Chin. J. Sports Med. 2019, 38, 755–761. [Google Scholar]
- Zhang, X. The Influence of Exercise Modes on DEN-induced Hepatocellular Carcinoma and Differential Metabonomic Profiling in Liver. China Sport Sci. Technol. 2019, 55, 68–73. [Google Scholar]
- Li, B. Effects of Aerobic Exercise on Plasma Metabolites in Prediabetes Subjects. Chin. J. Sports Med. 2018, 37, 301–308. [Google Scholar] [CrossRef]
- Fang, C. Influence of training models on water polo athletes blood serumenzyme and carbamide azote. Sichuan Sports Sci. 2016, 35, 31–34. [Google Scholar]
- Chen, A. Catalytic electrolysis of the cyanuric acid in swimming pool water. Appl. Chem. Ind. 2020, 49, 1970–1973. [Google Scholar]
- Plumb, R.S.; Granger, J.H.; Stumpf, C.L.; Johnson, K.A.; Smith, B.W.; Gaulitz, S.; Castro-Perez, J. A rapid screening approach to metabonomics using UPLC and oa-TOF mass spectrometry: Application to age, gender and diurnal variation in normal/Zucker obese rats and black, white and nude mice. Analyst 2005, 130, 844–849. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, L.; Lan, X.; Cohen, D.; Zhang, Y.; Ravindran, A.V.; Xie, P. Polyunsaturated fatty acids metabolism, purine metabolism and inosine as potential independent diagnostic biomarkers for major depressive disorder in children and adolescents. Mol. Psychiatry 2018, 24, 1478–1488. [Google Scholar] [CrossRef]
- Benton, H.P.; Ivanisevic, J.; Mahieu, N.G.; Kurczy, M.E.; Johnson, C.H.; Franco, L.; Siuzdak, G. Autonomous Metabolomics for Rapid Metabolite Identification in Global Profiling. Anal. Chem. 2015, 87, 884–891. [Google Scholar] [CrossRef] [Green Version]
- Bouatra, S.; Aziat, F.; Mandal, R.; Guo, A.C.; Wilson, M.R.; Knox, C.; Wishart, D.S. The human urine metabolome. PLoS ONE 2013, 8, e73076. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Morató, J.; Pozo, Ó.J.; Marcos, J. Targeting human urinary metabolome by LC-MS/MS: A review. Bioanalysis 2018, 10, 489–516. [Google Scholar] [CrossRef]
- Cruzat, V.F.; Pantaleão, L.C.; Donato, J., Jr.; de Bittencourt, P.I.H., Jr.; Tirapegui, J. Oral supplementations with free and dipeptide forms of l-glutamine in endotoxemic mice: Effects on muscle glutamine-glutathione axis and heat shock proteins. J. Nutr. Biochem. 2014, 25, 345–352. [Google Scholar] [CrossRef]
- Fernandes, C.V.; Julio, T. Effects of oral supplementation with glutamine and alanyl-glutamine on glutamine, glutamate, and glutathione status in trained rats and subjected to long-duration exercise. Nutrition 2009, 25, 428–435. [Google Scholar]
- Curi, R.; Newsholme, P.; Marzuca-Nassr, G.N.; Takahashi, H.K.; Hirabara, S.M.; Cruzat, V.; de Bittencourt, P.I.H., Jr. Regulatory principles in metabolism-then and now. Biochem. J. 2016, 473, 1845–1857. [Google Scholar] [CrossRef]
- Afzal, R.; Dowling, J.K.; McCoy, C.E. Impact of Exercise on Immunometabolism in Multiple Sclerosis. J. Clin. Med. 2020, 9, 3038. [Google Scholar] [CrossRef] [PubMed]
- Rennie, M.J.; Edwards, R.H.T.; Krywawych, S.; Davies, C.T.M.; Halliday, D.; Waterlow, J.C.; Millward, D.J. Effect of exercise on protein turnover in man. Clin. Sci. 1981, 61, 627–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jie, W. 1H-NMR Study of the Effect of Astaxanthin Supplementation on Urine Metabolism in Fatigue Rats; ShanXi University: Taiyuan, China, 2018. [Google Scholar]
- Eriksson, L.S.; Broberg, S.; Björkman, O.; Wahren, J. Ammonia metabolism during exercise in man. Clin. Physiol. 1985, 5, 325–336. [Google Scholar] [CrossRef]
- Kingsbury, K.J.; Kay, L.; Hjelm, M. Contrasting plasma free amino acid patterns in elite athletes: Association with fatigue and infection. Br. J. Sports Med. 1998, 32, 25. [Google Scholar] [CrossRef] [Green Version]
- Kargotich, S.; Keast, D.; Goodman, C.; Bhagat, C.I.; Joske, D.J.L.; Dawson, B.; Morton, A.R. Monitoring 6 weeks of progressive endurance training with plasma glutamine. Int. J. Sports Med. 2007, 28, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Gibala, M.J.; MacLean, D.A.; Graham, T.E.; Saltin, B. Tricarboxylic acid cycle intermediate pool size and estimated cycle flux in human muscle during exercise. Am. J. Physiol. 1998, 275 Pt 1, E235–E242. [Google Scholar] [CrossRef]
- Tan, F.; Polglaze, T.; Clark, S. Protocols for the Physiological Assessment of Water Polo Players; Human Kinetics: Windsor, ON, Canada, 2012. [Google Scholar]
- Josef, F. Biomarkers of peripheral muscle fatigue during exercise. BMC Musculoskelet. Disord. 2012, 13, 1–13. [Google Scholar]
- Beato, M.; Coratella, G.; Schena, F.; Hulton, A.T. Evaluation of the external and internal workload in female futsal players. Biol. Sport 2017, 34, 227. [Google Scholar] [CrossRef] [PubMed]
- Sewell, D.A.; Gleeson, M.; Blannin, A.K. Hyperammonaemia in relation to high-intensity exercise duration in man. Eur. J. Appl. Physiol. Occup. Physiol. 1994, 69, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Bernard, K. AMP deamination delays muscle acidification during heavy exercise and hypoxia. J. Biol. Chem. 2006, 281, 3057–3066. [Google Scholar]
- Murray, W.A. The biological significance of purine salvage. Annu. Rev. Biochem. 2003, 40, 811–826. [Google Scholar] [CrossRef]
- Sahlin, K.; Ekberg, K.; Cizinsky, S. Changes in plasma hypoxanthine and free radical markers during exercise in man. Acta Physiol. Scand. 1991, 142, 275–281. [Google Scholar] [CrossRef]
- Harkness, R.A.; Simmonds, R.J.; Coade, S.B. Purine transport and metabolism in man: The effect of exercise on concentrations of purine bases, nucleosides and nucleotides in plasma, urine, leucocytes and erythrocytes. Clin. Sci. 1983, 64, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Huang, W. Determination of nicotinic acid, isonicotinic acid, nicotinamide, isonicotinamide, 1-methyl nicotinamide chloride and nicotinic acid precursor, (tryptophan) in fortified functional beverages. Food Sci. Technol. 2019, 44, 336–340. [Google Scholar]
- Romani, M.; Hofer, D.C.; Katsyuba, E.; Auwerx, J. Niacin: An old lipid drug in a new NAD+ dress. J. Lipid Res. 2019, 60, 741–746. [Google Scholar] [CrossRef] [Green Version]
- Pajk, M.; Cselko, A.; Varga, C.; Posa, A.; Tokodi, M.; Boldogh, I.; Radak, Z. Exogenous nicotinamide supplementation and moderate physical exercise can attenuate the aging process in skeletal muscle of rats. Biogerontology 2017, 18, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Cantó, C.; Gerhart-Hines, Z.; Feige, J.N.; Lagouge, M.; Noriega, L.; Milne, J.C.; Auwerx, J. AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity. Nature 2009, 458, 1056–1060. [Google Scholar] [CrossRef] [PubMed]
- Ponugoti, B.; Kim, D.H.; Xiao, Z.; Smith, Z.; Miao, J.; Zang, M.; Kemper, J.K. SIRT1 deacetylates and inhibits SREBP-1C activity in regulation of hepatic lipid metabolism. J. Biol. Chem. 2010, 285, 33959–33970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirschey, M.D.; Shimazu, T.; Goetzman, E.; Jing, E.; Schwer, B.; Lombard, D.B.; Verdin, E. SIRT3 regulates mitochondrial fatty-acid oxidation by reversible enzyme deacetylation. Nat. Int. Wkly. J. Sci. 2010, 464, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Cox, G.R.; Mujika, I.; Van den Hoogenband, C.R. Nutritional recommendations for water polo. Int. J. Sport Nutr. Exerc. Metab. 2014, 24, 382–391. [Google Scholar] [CrossRef] [Green Version]
- Shasha, J. Study on the characteristics of energy supply and energy supply training of swimming sports. Sci. Technol. Inf. 2013, 35, 121–123. [Google Scholar]
- Yanchao, Z. Systematic Physiology and Biochemistry Training Monitoring of Long Distance Swimming Event; Shanghai University of Sport: Shanghai, China, 2012. [Google Scholar]
- Zhao, Y.; Pan, X.; Zhao, J.; Wang, Y.; Peng, Y.; Zhong, C. Decreased transketolase activity contributes to impaired hippocampal neurogenesis induced by thiamine deficiency. J. Neurochem. 2010, 111, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Li, J. Matebolic Study on Sprint Swimmer Participation in the 15th Doha Asian Games. China Sport Sci. 2008, 28, 42. [Google Scholar]
- Bo, G. Research on the Metabolic Features of Middle and Long Distance Runners in Heavy Load Training Stage and the Possible Mechanism of Acupoint Stimulation Based on Metabolomics; Shanghai University of Sport: Shanghai, China, 2019. [Google Scholar]
- Wei, X. Similarities and Differences of Niacin and Niacinamide in Study on Metabolism and Production of Ruminant. Chin. J. Anim. Nutr. 2020, 32, 5509–5515. [Google Scholar]
- Da Li, W.P.S.; Zhou, Y.M.; Liu, Q.G.; Zhou, S.S.; Luo, N.; Bian, F.N.; Guo, M. Chronic niacin overload may be involved in the increased prevalence of obesity in US children. World J. Gastroenterol. 2010, 16, 2378–2387. [Google Scholar]
- Rutkowski, B.; Rutkowski, P.; Słomińska, E.; Smolenski, R.T.; Świerczyński, J. Cellular toxicity of nicotinamide metabolites. J. Ren. Nutr. 2012, 22, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M.; Hawley, J.A.; Wong, S.H.; Jeukendrup, A.E. Carbohydrates for training and competition. J. Sports Sci. 2011, 29 (Suppl. 1), S17–S27. [Google Scholar] [CrossRef]
- Costill, D.L.; Flynn, M.G.; Kirwan, J.P.; Houmard, J.A.; Mitchell, J.B.; Thomas, R.; Park, S.H. Effects of repeated days of intensified training on muscle glycogen and swimming performance. Med. Sci. Sports Exerc. 1988, 20, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Kerksick, C.; Harvey, T.; Stout, J.; Campbell, B.; Wilborn, C.; Kreider, R.; Antonio, J. International society of sports nutrition position stand: Nutrient timing. J. Int. Soc. Sports Nutr. 2017, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shaw, G.; Koivisto, A.; Gerrard, D.; Burke, L.M. Nutrition considerations for open-water swimming. Int. J. Sport Nutr. Exerc. Metab. 2014, 24, 373–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Number | Age (Years) | Height (cm) | Weight (kg) | BMI | Athlete Level |
|---|---|---|---|---|---|---|
| Female | 12 | 16.46 ± 1.56 | 172.69 ± 5.14 | 65.23 ± 7.97 | 21.84 ± 2.30 | Level 1 |
| NO. | R2X (cum) | R2Y (cum) | Q2 (cum) | Model Source | Validation Results |
|---|---|---|---|---|---|
| 1 | 0.218 | 0.95 | 0.633 | T3 vs. T1 | Established |
| 2 | 0.287 | 0.987 | 0.752 | T3 vs. T2 | Established |
| No. | Name | Ion | m/z | rt(s) | FC | Trend T2 vs. T1 (p) | FC | Trend T3 vs. T2 (p) |
|---|---|---|---|---|---|---|---|---|
| 1 | N1-methyl-2-pyridone-5-carboxamide (2-pyridone) | (M + H) + | 153.065 | 85.348 | 2.61 | ↑ (0.000042) | ||
| 2 | Indoleacetic acid | (M + H) + | 176.070 | 224.632 | 2.47 | ↑ (0.0045) | ||
| 3 | Adenosine monophosphate (AMP) | (M + H − H2O) + | 330.058 | 289.773 | 2.091 | ↑ (0.00046) | ||
| 4 | N-acetylglutamine | (M + H) + | 189.086 | 313.151 | 0.44 | ↓ (0.00412) | 0.38 | ↓ (0.000114) |
| 5 | Guanidinosuccinic acid | (M + H) + | 176.066 | 403.111 | 2.10 | ↑ (0.000969) | ||
| 6 | Decanoyl-L-carnitine | M+ | 316.247 | 189.977 | 5.01 | ↑ (0.00076) | 2.13 | ↑ (0.02635) |
| 7 | Trimethylamine N-oxide (TMAO) | (M + H) + | 76.075 | 328.576 | 0.36 | ↓ (0.00172) | 0.38 | ↓ (0.0000139) |
| 8 | N6-methyl-L-lysine | (M + H − H2O) + | 143.116 | 369.352 | 3.08 | ↑ (0.0091) | ||
| 9 | L-Glutamine (L-Gln) | (M + H) + | 147.075 | 225.261 | 2.41 | ↑ (0.00288) | ||
| 10 | 1-Deoxynojirimycin | (M + CH3COO + 2H) + | 224.112 | 261.669 | 1.92 | ↑ (0.0241) | ||
| 11 | Allopurinol riboside | (M + H − H2O) + | 251.077 | 131.79 | 1.85 | ↑ (0.0370) | ||
| 12 | Hypoxanthine (Hx) | (M + H) + | 137.044 | 201.653 | 6.73 | ↑ (0.0035) | 3.47 | ↑ (0.0134) |
| 13 | Taurine | (M + H) + | 126.021 | 299.017 | 3.20 | ↑ (0.00096) | ||
| 14 | Indole-3-pyruvic acid | (M + NH4) + | 221.090 | 289.189 | 1.85 | ↑ (0.00267) | ||
| 15 | Nicotinamide (NAM) | (M + H) + | 123.054 | 64.824 | 0.49 | ↓ (0.00005) | 0.36 | ↓ (0.0000062) |
| 16 | N-Acetyl-L-Histidine | (M + H) + | 198.086 | 315.946 | 0.50 | ↓ (0.023) | 0.35 | ↓ (0.0000089) |
| 17 | N6-methyladenosine | (M + H) + | 181.071 | 65.181 | 2.44 | ↑ (0.00338) | 2.30 | ↑ (0.00480) |
| 18 | Betaine (Bet) | (M + H) + | 189.086 | 287.639 | 1.49 | ↑ (0.025) | ||
| 19 | Palmitoyl ethanolamide | (M + H) + | 300.289 | 36.74 | 0.72 | ↓ (0.018) | ||
| 20 | Primidone | (M + H) + | 219.111 | 225.554 | 1.92 | ↑ (0.046) | ||
| 21 | Dopamine | (M + H − H2O) + | 136.074 | 225.573 | 1.98 | ↑ (0.0532) | ||
| 22 | L-methylnicotinamide | M+ | 137.069 | 260.072 | 0.56 | ↓ (0.054) | ||
| 23 | Kynurenic acid | (M + H) + | 190.048 | 197.539 | 2.24 | ↑ (0.00071) | ||
| 24 | N-acetylmannosamine | (M + H − H2O) + | 204.085 | 257.449 | 1.72 | ↑ (0.00828) | ||
| 25 | 4-imidazoleacetic acid | (M + H) + | 127.049 | 341.473 | 0.32 | ↓ (0.000028) | ||
| 26 | S-Methyl-5′-thioadenosine | (M + H) + | 298.09 | 99.834 | 0.70 | ↓ (0.01918) | ||
| 27 | Phosphorylcholine | (M + H) + | 184.072 | 487.868 | 0.62 | ↓ (0.02277) |
| NO. | Pathway Name | Match Statue | p | −log(p) | Holm p | FDR | Impact |
|---|---|---|---|---|---|---|---|
| 1 | Nicotinate and nicotinamide metabolism | 3/15 | 0.00075822 | 3.1202 | 0.06369 | 0.06369 | 0.33246 |
| 2 | Purine metabolism | 3/65 | 0.048181 | 1.3171 | 1.0 | 1.0 | 0.08068 |
| 3 | D-Glutamine and D-glutamate metabolism | 1/6 | 0.075082 | 1.1245 | 1.0 | 1.0 | 0.0 |
| 4 | Nitrogen metabolism | 1/6 | 0.075082 | 1.1245 | 1.0 | 1.0 | 0.0 |
| 5 | Phosphonate and phosphinate metabolism | 1/6 | 0.075082 | 1.1245 | 1.0 | 1.0 | 0.5 |
| 6 | Taurine and hypotaurine metabolism | 1/8 | 0.098896 | 1.0048 | 1.0 | 1.0 | 0.42857 |
| 7 | Arginine biosynthesis | 1/14 | 0.16689 | 0.77756 | 1.0 | 1.0 | 0.0 |
| 8 | Alanine, aspartate and glutamate metabolism | 1/28 | 0.3071 | 0.51272 | 1.0 | 1.0 | 0.11378 |
| 9 | Glyoxylate and dicarboxylate metabolism | 1/32 | 0.34284 | 0.46491 | 1.0 | 1.0 | 0.0 |
| 10 | Glycine, serine and threonine metabolism | 1/33 | 0.3515 | 0.45407 | 1.0 | 1.0 | 0.05034 |
| 11 | Amino sugar and nucleotide sugar metabolism | 1/37 | 0.38506 | 0.41447 | 1.0 | 1.0 | 0.04723 |
| 12 | Pyrimidine metabolism | 1/39 | 0.40122 | 0.39662 | 1.0 | 1.0 | 0.0 |
| 13 | Tryptophan metabolism | 1/41 | 0.41697 | 0.3799 | 1.0 | 1.0 | 0.0 |
| 14 | Tyrosine metabolism | 1/42 | 0.4247 | 0.37192 | 1.0 | 1.0 | 0.12972 |
| 15 | Primary bile acid biosynthesis | 1/46 | 0.45464 | 0.34233 | 1.0 | 1.0 | 0.00758 |
| 16 | Aminoacyl-tRNA biosynthesis | 1/48 | 0.46906 | 0.32878 | 1.0 | 1.0 | 0.0 |
| No. | Pathway Name | Match Statue | P | −log(p) | Holm p | FDR | Impact |
|---|---|---|---|---|---|---|---|
| 1 | Nicotinate and nicotinamide metabolism | 1/15 | 0.04752 | 1.3231 | 1.0 | 1.0 | 0.1943 |
| 2 | Histidine metabolism | 1/16 | 0.050622 | 1.2957 | 1.0 | 1.0 | 0 |
| 3 | Purine metabolism | 1/65 | 0.19304 | 0.714435 | 1.0 | 1.0 | 0.01651 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.-l.; Chen, A.-p.; Li, J.-y.; Sun, Z.; Yan, S.-l.; Xu, K.-y. Mechanism of the Effect of High-Intensity Training on Urinary Metabolism in Female Water Polo Players Based on UHPLC-MS Non-Targeted Metabolomics Technique. Healthcare 2021, 9, 381. https://0-doi-org.brum.beds.ac.uk/10.3390/healthcare9040381
Wang L-l, Chen A-p, Li J-y, Sun Z, Yan S-l, Xu K-y. Mechanism of the Effect of High-Intensity Training on Urinary Metabolism in Female Water Polo Players Based on UHPLC-MS Non-Targeted Metabolomics Technique. Healthcare. 2021; 9(4):381. https://0-doi-org.brum.beds.ac.uk/10.3390/healthcare9040381
Chicago/Turabian StyleWang, Lei-lei, An-ping Chen, Jian-ying Li, Zhuo Sun, Shi-liang Yan, and Kai-yuan Xu. 2021. "Mechanism of the Effect of High-Intensity Training on Urinary Metabolism in Female Water Polo Players Based on UHPLC-MS Non-Targeted Metabolomics Technique" Healthcare 9, no. 4: 381. https://0-doi-org.brum.beds.ac.uk/10.3390/healthcare9040381