
Kinetics of Odorant Recognition with a Graphene-Based Olfactory Receptor Mimicry


1
Department of Nanobiotechnology, University of Natural Resources and Life Sciences (BOKU), Muthgasse 18, 1190 Vienna, Austria
2
Children’s Hospital Boston, Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, Boston, MA 02115, USA
*
Author to whom correspondence should be addressed.
†
Former name: Caroline Kotlowski.
Chemosensors 2022, 10(6), 203; https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors10060203
Submission received: 25 February 2022
/
Revised: 13 May 2022
/
Accepted: 20 May 2022
/
Published: 27 May 2022
(This article belongs to the Special Issue Nanomaterials Based on Bio/Chemical Sensors)
Abstract
:Malaria vector mosquito species rely on a handful of specific pheromones for mating; one of them, sulcatone (6-methyl-5-hepten-2-one), is also found in human exudation. Therefore, a complete understanding of the insect’s olfaction, and rapid real-time methods for odorant detection, are required. Here, we mimic the odorant recognition of the nerve cells of an insect’s antenna with a synthetic graphene-based bio-electro-interfacial odorant receptor. By this means, we obtain the kinetics of the genuine odorant recognition reaction and compare them to electro-antennogram data that represent the more complex scenario of a living insect. The odorant-binding proteins OBP 9A and 9B only associate with their ligands weakly, showing KDs of between 2.1 mM and 3 mM, while the binding kinetics of OBP proteins depend on the structural feature of a cystine knot and are modulated by the local milieu within a protein-aided enhancement zone.
1. Introduction
The small-molecule pheromone 6-methyl-5-hepten-2-one (sulcatone) merits attention for its two-fold role in one of the critical safety issues of our time, being linked to climate change [1,2]. It is associated with the spread of malaria, being one of the five main aggregation pheromones involved in swarming initiation and also mating success in malaria vector mosquito species [3,4,5]. The pheromone attracts the mosquito species, Anopheles arabiensis and An. gambiae, and increases mating for another five malaria vectors; moreover, it is a degradation product of terpenes and, thus, indicates the breakdown of plant defense [6]. In comparison, the structurally similar 3-octanol is a degradation product of lipids and indicates the activity of fungi [7] and, thus, the potential presence of mycotoxins [8]. For both C-8 marker compounds, rapid, real-time monitoring is highly desirable.
Here we used a graphene-based field-effect transistor (FET), which we linked communicatively to the odor binding proteins, TcasOBP9A and TcasOBP9B, of the beetle Tribolium castaneum to mimic the nerve cell receptor (Figure 1). The nerve cell receptor TcOr1 is highly conserved (ortholog of Drosophila Or83b) and prevails in most insects [9,10,11,12]. Beyond detecting two critical pheromone-volatiles in real time, this approach allows us to compare the reaction kinetics of odorant recognition in the insect’s antenna of our synthetic receptor, and so discover previously unknown regulatory pathways. We had previously established our bio-transistor, which detected the binding of various small molecules to Apis mellifera OBP14 [13], and revealed that the binding kinetics of OBP proteins depend on their cysteine bridging pattern [14]. While OBP14 contains 2 S-S-bonds, with cysteines linked according to the pattern 1–3, 4–5 (with no cystine knot, 2 is an additional free cysteine), OBP9A/B has 3 S-S-bonds that form a cystine knot with the (rotaxane-) pattern 1–3, 2–5, 4–6 [15]. A comparison of wild-type OBP14 and Amel SS-OBP14, a Gln44Cys, His 97Cys S-S-OBP14-double-mutant (containing an additional disulfide bond between helices α3 and α6, to achieve a 1–3, 2–5, 4–6 pattern) showed that the restriction of molecular dynamics and the stability increase in the rotaxane substructure, which results from cystine knot formation [16], while leaving the crystal structure largely congruent to the non-rotaxane ones, accelerates the on-rate in small-molecule binding and, ultimately, leads to tighter binding [14]. Therefore, it seems a logical step to link such an OBP to our sensor platform that shows a double cystine knot crosslinking pattern, especially considering future field sensor applications where stable proteins are required. We chose Tribolium castaneum OBP9A and OBP9B, according to their high expression levels in the antenna, which suggested an important role in olfactory processes; moreover, RNA-interference experiments in which both proteins were knocked out demonstrated a massive loss of sensitivity of the insect antennae [17].
On the dendrites of the living insect’s sensilium, pheromone volatiles are absorbed by a lymphatic liquid that can be viewed as a protein-aided enhancement zone, in which OBPs bind, protect, enrich and transport the pheromone [18] (Figure 1). However, the stability/dynamics-dependent ligand-binding kinetics leave room for speculation that smell-sensing may be controlled by a specific local milieu in the vicinity of the nerve cell membrane that would modulate the OBPs’ stability/dynamics. There are plenty of reports of nerve cells signaling via the release of ions or small molecules [19], thus changing, very locally, the milieu of the surrounding medium. Usually, such experiments are conducted with advanced microscopic techniques and the results are viewed in the light of cell-cell communication [20,21]; however, here, a case of cell-OBP communication becomes a matter for investigation. It is beyond the scope of this paper to investigate the local milieu at the smell receptor’s surface; however, we show a comparison between our synthetic receptor (i.e., a FET-transistor, linked to OBP9A/B) and the electroantennograms (EAG) of living Tribolium castaneum antenna. The binding of sulcatone and 3-octanol appears to be energetically equal (with comparable KD) but the binding kinetics are strongly accelerated in the living antenna.
2. Materials and Methods
Protein production. OBP9A and OBP9B were expressed in E.coli as described previously [13,14] and purified with standard chromatographical methods, including a final-size exclusion step with a Sepharose 200 column. The chromatograms showed a sharp, symmetric peak with no shoulders and a reducing SDS-Page of different fractions within this peak confirmed one sharp band. The non-reducing SDS-Page showed the same molecular weight; thus, the S-S-bonds have formed a cystine-knot pattern.
The amino acid sequences of OBP9A and OBP9B are the following:
- TcasOBP9A: AMSEAQLKAAVKLVRNMCQPKSKATNEDIEKMHHGDWNIDRTAMCYMHCALNSNKLITKENV
- TcasOBP9B: AMSEAQMKAALKLVRNVCQPKTKATNEQIEAMHTGNWDLDKNGKCYMWCILNMYKLIGKDNS
- TcasOBP9A: FNRDYAITLAEKNLPTALKTASIEAANLCKDSAKTLDDKCVAAYEISKCLYESNPEKYFLP
- TcasOBP9B: FDWEAGIATLKAQAPESVRDPAIASVNNCKDAVKTTSDKCEAAYEIAHCMYLDNPEKYFLP
Preparation of large graphene oxide (GO) sheets. Graphene oxide (GO) was prepared by a modified Hummers/Offeman method, as follows [23]. First, 2 g of 125–250 μm-sized graphite flakes (NGS Naturgraphit GmbH, Leinburg, Germany) were stirred in 12 mL of concentrated sulfuric acid (98%) for 4 h at 80 °C. After cooling down to room temperature, the suspension was sonicated for 30 min at 300 W and diluted with 500 mL deionized H2O, then the flakes were filtrated at 0.2 μm pore size and dried in an exicator overnight. The graphite flakes were again suspended in concentrated sulfuric acid (120 mL) and stirred for 30 min at −20 °C. Subsequently, 15 g of potassium permanganate was interspersed over a period of 2 h while the exothermal reaction was kept at −20 °C, and further stirred at RT for another 2 h. For oxidative exfoliation, the suspension was diluted stepwise into 1 L of deionized H2O and a 30% solution of H2O2 was added. Subsequently, the graphene oxide was skimmed from the water surface and stirred in fresh deionized H2O.
Preparation of rGO-FETs. The employed protocol followed the procedure described elsewhere in the literature [13,14]. Briefly, silicon substrates with a 300 nm oxide layer were chosen as a basal layer for the FETs. The SiO2 substrates were cleaned with a standard RCA cleaning procedure. To obtain an adhesive self-assembled monolayer, the substrates were submerged in a 2% aminopropyltriethoxysilane (APTES) solution in ethanol for 1 h. After rinsing with ethanol, the substrates were heated to 120 °C for two hours and afterward cooled to room temperature. The graphene oxide flakes were applied to the APTES-modified Si-Wafer via drop-casting of the graphene-oxide suspension. The deposited graphene oxide was reduced in hydrazine vapor using hermetically sealed glass Petri dishes that contain the substrates and a droplet of hydrazine at 70 °C overnight. Flake distribution was first checked with an optical microscope and selected devices, then characterized using SEM (see Supplementary Materials S1). T-shaped source and drain electrodes were applied by the evaporation of gold onto the graphene surface, as shown in the sketch in Figure 1 and further described by the authors of [24].
Electrical characterization of synthetic receptors. The resulting FET chips were sorted, based on conductivity, and only chips that showed a resistance smaller than ~800 Ohms were used. A silver–silver chloride reference electrode (Flex Ref, World Precision Instruments, Sarasota, USA) was used to operate the FET device in a liquid gate configuration. Source-drain-currents (ISD) were plotted against the different gate voltages (VG) to optimize the measuring range (Figure 2A). A VG of −0.6 V was finally chosen for all subsequent measurements. The source-drain bias VSD = 0.05 V.
Synthetic receptors odorant binding. For the detection of odorants, the rGO-based biosensors were functionalized with odorant-binding proteins TcasOBP9A or TcasOBP9B from T. castaneum. In order to anchor the recombinant proteins to the sensing area, the inorganic sp2-carbon surface was π-π-stacked with 1-pyrenebutanoic acid succinimidyl ester (PBSE), which served as a bi-functional linker by covalently binding the proteins. Therefore, 20 μL of a 5 μM PBSE solution in THF was placed onto the detection area of the rGO-FET, followed by 10 μL of a 10 μM protein solution in PBS buffer (10 mM; pH 8.0); the chip was incubated for 2 h at 4 °C. The same buffer was also used to equilibrate the system until baseline stability was obtained, and to prepare odorant solutions of sulcatone and 3-octanol (Sigma-Aldrich) at concentrations of between 300 μM and 5 mM.
The synthetic receptors were contacted in a polymer flow cell and the source-drain currents were monitored in real time, using a Keithley 4200 semiconductor characterization system.
3. Results
Proteins. Expression and purification of the proteins OBP9A and OBP9B were controlled by SDS-Page and resulted in sharp single bands, both at the same MW. The presence of the disulfide bonds was checked by comparing reducing and non-reducing gels.
Plain synthetic transistor. The plain rGO-FETs were only loaded with the PBSE-linker (which, in the functional receptor, would link an OBP to the graphene surface). The linker also serves as a passivation layer, thus preventing all non-protein-covered surfaces from generating non-specific signals [13]. The transistor/receptor setup is depicted in Figure 1 and in panels G and H in Figure 2.
Feeding the odorants sulcatone, 3-octanol, and 2-octanone, at concentrations in the range of the maximum used in the following experiments (up to 5 mM), into these plain receptors resulted in no significant response, as shown in Figure 2B.
Synthetic ligand-binding receptor. Further controls were performed with the full functional receptor, consisting of an rGO-FET coupled to an OBP9B. A gate voltage swipe (Figure 2A) revealed that the synthetic receptor can be tuned and has a preferred operating range of between −0.6 and 0 V. Repeated exposure of the receptor to sulcatone stimuli of 0.5 mM or 1 mM proved it to have excellent baseline stability and reproducible signal amplification (Figure 2D). Detailed analysis of the single concentration steps yielded a high signal-to-noise ratio. The green dash-dotted and the blue dotted lines in Figure 2C are the simulated boundaries of a 20% error interval of the observed reaction rate. Fitting with established models [13], shown as a red line, yields an R2 of 0.99.
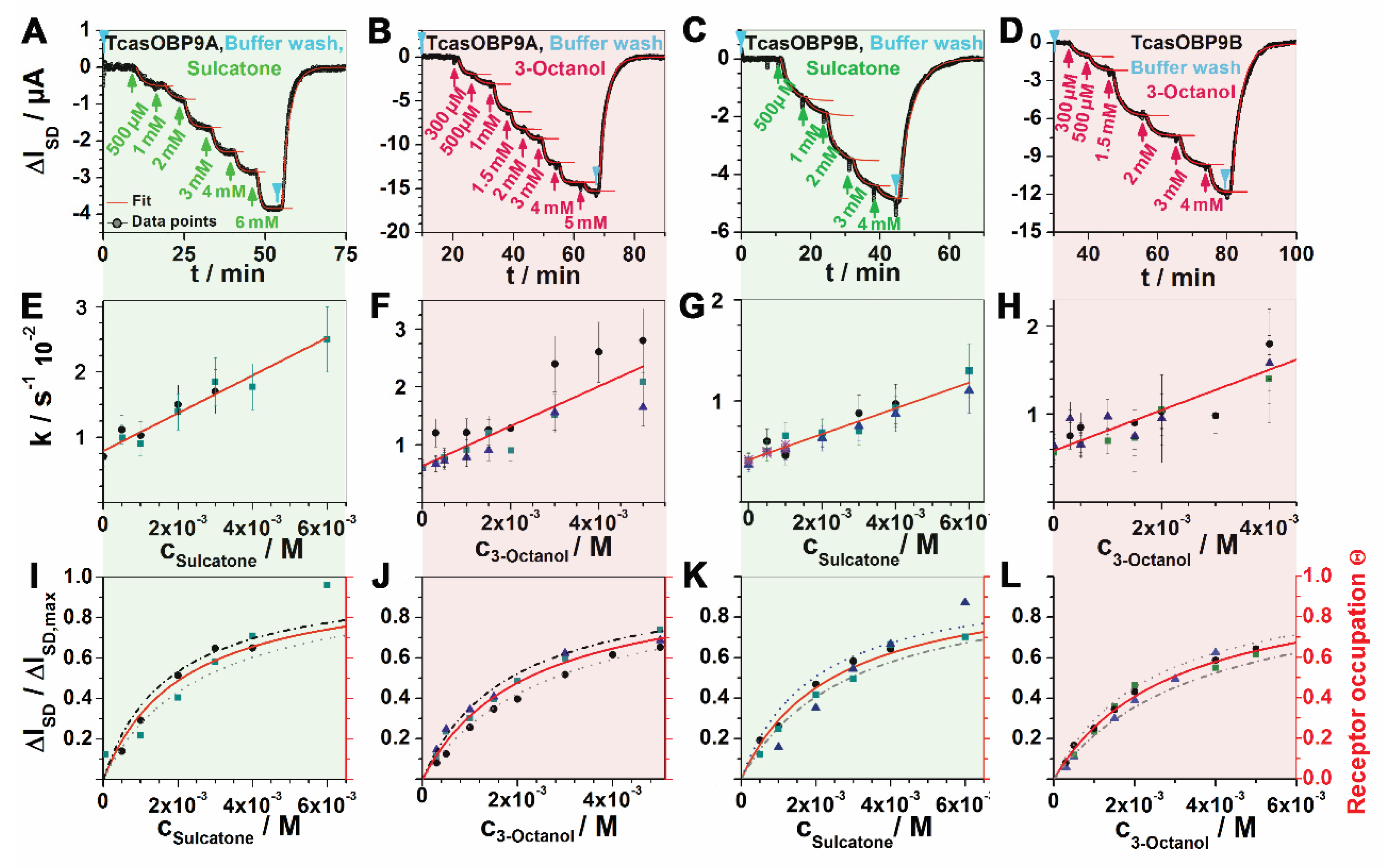
Synthetic odorant receptors were set up with OBP9A and OBP9B and stimulated with the odorants sulcatone and 3-octanol at stepwise-increasing concentrations from 300 μM to 6 mM as the source-drain current, ΔISD, was monitored on-line in real time (shown in Figure 3A–D). The subsequent rinsing step with pure buffer re-set the system to the pristine baseline level, demonstrating the full reversibility of the affinity reaction between the receptor and the odorant. These raw data were subjected to a global analysis that proved the internal consistency of each data set (each OBP–odorant combination) as described previously by the authors of [13]. Dissociation constants (KD) were obtained in two ways. For the kinetic approach, see the k/codorant plots in panels E–H, in which the intercept and slopes indicate koff and kon; thus, a kinetic KD is given by their quotient. For thermodynamic KDs, which more directly represent the equilibrium state, the relative response was plotted against the stimulus concentration (Figure 3, panels I–L), which also yielded the receptor occupation. Fitting in a Langmuir isotherm yielded the KDs summarized in Table 1. The kinetic KDs corresponded to the thermodynamic ones within an error margin of 20%; thus, proving the internal consistency of the data.
Compared to OBP9B, OBP9A yielded slightly higher but still similar on-rates and affinities, while wild-type OBP14 also yielded similar affinities to sulcatone. The same holds true for the OBP9A-binding 3-octanol. Overall, both odorants are comparable OBP-binders (Table 1). However, a comparison between wtOBP14 and the more disulfide-crosslinked Gln44Cys, His 97Cys mutant in terms of binding eugenol reveals a 10-fold affinity gain (on-rate-driven). In contrast, BmorPBP1 binds bombykol with high affinity, despite a loose S-S-pattern similar to OBP14 at pH 8, and differs from the S-S-mutant OBP14- or OBP14-binding higher-affinity ligands by a 100-fold lowered exchange rate (lower kon compensated for by lower koff). Binding shows a strong pH dependence, with no detectable binding at pH 4 [26].
The synthetic receptors with TcasOBP9A/B were closely comparable to the living insect’s antennae of the T. castaneum in terms of binding equilibria but differed in the exchange rate, which was 10 times higher in the insect.
The electroantennograms (EAG) in Figure 2E,F are courtesy of Karthi Balakrishnan [25]. The kinetic constants, KD and kon, were converted from pressure units (μbar and μbar−1) to molar units using Henry’s law constants, at 0.14 mol m−3 pa−1 for sulcatone [27] and 0.31 mol m−3 pa−1 for 3-octanol [28]. The results are shown in Table 1.
4. Discussion
In the living insect’s antenna, an odorant is captured, accumulated, protected, and transported by an odorant-binding protein, which then hands the odorant over to the odorant receptor on the nerve cell membrane [18]. Speaking in technological terms, a membrane receptor can be described as a transistor. While a synthetic field-effect transistor transduces and enhances an electrical signal (electric field) that is applied to its gate into a source-drain current, initiating a “signal cascade” that ends with a graph on our screens, the receptor, as a biological transistor, transduces the biological signal (the presence of an odorant) into its own form of nerve cell signal cascade, which, in turn, ends up with the insect sensing a smell. Much like the synthetic FET, the signal cascade of the receptor is largely electrical, although the charge carriers are ions. The latest generation of synthetic FET transistors, which we introduced in a previous work [13] and which will be compared to a membrane receptor in the light of this new data, comes even closer to the principle of a biological transistor because it is biologically, not electrically, gated (triggered by an odorant). Briefly, the signal generation is based on the high sensitivity of graphene–SiO2 sandwiches to local electric fields. At the nanometer scale, the semiconducting graphene layer is somewhat hilly in terms of topography and surface potential [29]; thus, it behaves like the 2-dimensional equivalent of a bulk semiconductor with p- and n-doped patches. Applying a bias in a lateral direction between the source and drain electrodes yields an electrical circuit consisting of x serial n-p-n-p junctions (a transistor array). The OBPs, which are communicatively close when linked to the graphene, exhibit a substantial change of dipole moment upon binding their ligands [30,31]; the signals displayed in the ID-VG curves, as well as the ID vs time plots, prove that the transistor array reacts very sensitively to this field effect.
It has always been puzzling that the olfactory receptors in an insect’s antennae, as such, lack specificity and that the OBPs also fail to compensate for this missing selectivity, all of which still leaves the highly selective smell perception unexplained [31]. Therefore, it seems plausible that nerve cells may auto-regulate their sensing of a specific odorant by altering their local milieu (i.e., the milieu within their protein-aided enhancement zone), irrespective of the precise type of milieu regulation. We have shown previously that pH is part of the affinity modulating milieu [26]; furthermore, pH-dependent NMR structures of the pheromone-binding protein 1 [18] also point to a pH-modulated mechanism. However, it is premature to rule out other compounds of a modulating milieu at this stage. This milieu-regulation in insects’ antennae is likely to pose additional challenges to the development of bio-inspired sensing applications.
However, when used as a sensor, our system was capable of detecting even low-affinity OBP ligands and provided a certain level of progress in terms of selectivity and the ability to finely discriminate between small-molecule structures via the mutagenetic tailoring of the binding pocket [14,32,33].
Our synthetic smell receptor provides an opportunity to study smell-sensing in the absence of any presumed autoregulation (in a stable milieu, i.e., a buffer solution) and compare it to the living insect´s antennae. Remarkably, both systems—the FET-receptor and the living antenna—showed almost equal affinities to sulcatone and 3-octanol (this confirms that in both systems, the same binding reaction is predominant) but differed in the exchange rate, which is given by approx. 10-fold higher on- and off-rates in the antenna. This acceleration in the real antennae is crucial for the insect´s ultrafast smell-sensing, which allows for in-flight navigation.
A comparison between recombinant Apis mellifera OBP14 constructs, containing (mutant) or not containing (wt) an additional disulfide bond [14]; therefore, differing in a rotaxane-like intertwinement of their polypeptide chains, reveals the importance of a stabilizing cystine knot for odorant binding. We hypothesize that the wild-type OBP14 (as in other similar OBPs that lack one of the 3 S-S bonds) exhibits enhanced conformational dynamics, which slows down odorant-binding due to a tweezer-like restrictive oscillation of the binding pocket. In fact, we found that OBPs were very stable already, with a vacant binding pocket, but this stability further increased when the binding pocket was occupied with an odorant, characterized by a melting-point increase from 57 °C to 64 °C [14]. This link between binding kinetics (especially the exchange rate) and the dynamics of the construct, indicates that any milieu that affected stability would result in differential on- and off-rates for odorants. In this context, the increased (but compensating for each other) on- and off-rates in living antennae, when compared to those we measured with our synthetic olfactory receptor, may result from a stabilizing milieu within the protein-aided enhancement zone.
Finally, our data show no significant differences between the OBP9 variants A and B. It has been reported that OBP9B is naturally expressed at high levels in the antenna, but if OBP9B is knocked out, OBP9A is ready as a substitute.
5. Conclusions
This work may open up a path for explaining how the insect´s olfactory system discriminates between odorants, despite the lack of specific receptors, and to a future understanding of ultrafast smell-sensing. It is only meant as a starting point and should encourage others to shed further light on the molecular dynamics of OBPs, as well as the local milieu (and milieu changes) in which OBPs work in the antenna. This FET-transistor, sensitive to the malaria vectors of mosquito pheromones, is an opportunity for future developments in malaria risk monitoring; this work should also raise awareness of the subtle regulatory mechanisms that might make it more difficult to mimic insects for odor-recognition applications.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/chemosensors10060203/s1.
Author Contributions
Conceptualization, C.B.; methodology, C.B.; software, C.B.; validation, C.B.; formal analysis, C.B.; investigation, C.B.; writing—original draft preparation, C.B. and K.B.; writing—review and editing, C.B.; visualization C.B. and K.B.; supervision, C.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Rosa Mastrogiacomo and Paolo Pelosi for the proteins. We are grateful to a number of colleagues who contributed their advice: Karthi Balakrishnan, Stefan Schütz, and Wolfgang Knoll.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Grünig, M.; Mazzi, D.; Calanca, P.; Karger, D.N.; Pellissier, L. Crop and forest pest metawebs shift towards increased linkage and suitability overlap under climate change. Commun. Biol. 2020, 3, 233. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mozūraitis, R.; Hajkazemian, M.; Zawada, J.W.; Szymczak, J.; Pålsson, K.; Sekar, V.; Biryukova, I.; Friedländer, M.R.; Koekemoer, L.L.; Baird, J.K.; et al. Male swarming aggregation pheromones increase female attraction and mating success among multiple African malaria vector mosquito species. Nat. Ecol. Evol. 2020, 4, 1395–1401. [Google Scholar] [CrossRef] [PubMed]
- McBride, C.S.; Baier, F.; Omondi, A.B.; Spitzer, S.A.; Lutomiah, J.; Sang, R.; Ignell, R.; Vosshall, L.B. Evolution of mosquito preference for humans linked to an odorant receptor. Nature 2014, 515, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Raji, J.I.; Melo, N.; Castillo, J.S.; Gonzalez, S.; Saldana, V.; Stensmyr, M.C.; DeGennaro, M. Aedes aegypti mosquitoes detect acidic volatiles found in human odor using the IR8a pathway. Curr. Biol. 2019, 29, 1253–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francke, W.; Schulz, S. 4.04-Pheromones of terrestrial invertebrates. In Comprehensive Natural Products II; Elsevier: Amsterdam, The Netherlands, 2010; pp. 153–223. [Google Scholar]
- Quiroz, A.; Pettersson, J.; Pickett, J.A.; Wadhams, L.J.; Niemeyer, H.M. Semiochemicals Mediating Spacing Behavior of Bird Cherry-Oat Aphid, Rhopalosiphum padi Feeding on Cereals. J. Chem. Ecol. 1997, 23, 2599–2607. [Google Scholar] [CrossRef]
- Aphids as Crop Pest; Van Emden, H.F.; Harrington, R. (Eds.) Cabi: Wallingford, UK, 2014. [Google Scholar]
- Jones, W.D.; Nguyen, T.A.T.; Kloss, B.; Lee, K.J.; Vosshall, L.B. Functional conservation of an insect odorant receptor gene across 250 million years of evolution. Curr. Biol. 2005, 15, R119–R121. [Google Scholar] [CrossRef] [Green Version]
- Trebels, B.; Dippel, S.; Schaaf, M.; Balakrishnan, K.; Wimmer, E.A.; Schachtner, J. Adult neurogenesis in the mushroom bodies of red flour beetles (Tribolium castaneum, Herbst) is influenced by the olfactory environment. Sci. Rep. 2020, 10, 1090. [Google Scholar] [CrossRef]
- Engsontia, P.; Sanderson, A.P.; Cobb, M.; Walden, K.K.; Robertson, H.M.; Brown, S. The red flour beetle’s large nose: An expanded odorant receptor gene family in Tribolium castaneum. Insect Biochem. Mol. Biol. 2008, 38, 387–397. [Google Scholar] [CrossRef]
- Dippel, S.; Kollmann, M.; Oberhofer, G.; Montino, A.; Knoll, C.; Krala, M.; Wimmer, E.A. Morphological and transcriptomic analysis of a beetle chemosensory system reveals a gnathal olfactory center. BMC Biol. 2016, 14, 90. [Google Scholar] [CrossRef] [Green Version]
- Larisika, M.; Kotlowski, C.; Steininger, C.; Mastrogiacomo, R.; Pelosi, P.; Schütz, S.; Peteu, S.; Kleber, C.; Reiner-Rozman, C.; Nowak, C.; et al. Electronic olfactory sensor based on A. mellifera odorant-binding protein 14 on a reduced graphene oxide field-effect transistor. Angew. Chem. Int. Ed. 2015, 54, 13245–13248. [Google Scholar] [CrossRef] [PubMed]
- Kotlowski, C.; Larisika, M.; Guerin, P.M.; Kleber, C.; Kröber, T.; Mastrogiacomo, R.; Nowak, C.; Pelosi, P.; Schütz, S.; Schwaighofer, A.; et al. Fine discrimination of volatile compounds by graphene-immobilized odorant-binding proteins. Sens. Actuators B Chem. 2018, 256, 564–572. [Google Scholar] [CrossRef]
- Daly, N.L.; Craik, D.J. Bioactive cystine knot proteins. Curr. Opin. Chem. Biol. 2011, 15, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Lustbader, J.W.; Liu, Y.; Canfield, R.E.; Hendrickson, W.A. Structure of human chorionic gonadotropin at 2.6 Å resolution from MAD analysis of the selenomethionyl protein. Structure 1994, 2, 545–558. [Google Scholar] [CrossRef] [Green Version]
- Montino, A.; Balakrishnan, K.; Dippel, S.; Trebels, B.; Neumann, P.; Wimmer, E.A. Mutually Exclusive Expression of Closely Related Odorant-Binding Proteins 9A and 9B in the Antenna of the Red Flour Beetle Tribolium castaneum. Biomolecules 2021, 11, 1502. [Google Scholar] [CrossRef]
- Damberger, F.F.; Michel, E.; Ishida, Y.; Leal, W.S.; Wüthrich, K. Pheromone discrimination by a pH-tuned polymorphism of the Bombyx mori pheromone-binding protein. Proc. Natl. Acad. Sci. USA 2013, 110, 18680–18685. [Google Scholar] [CrossRef] [Green Version]
- Kaissling, K.E. Responses of insect olfactory neurons to single pheromone molecules. In Olfactory Concepts of Insect Control-Alternative to Insecticides; Springer: Cham, Switzerland, 2019; pp. 1–27. [Google Scholar]
- Meyer, S.A.; Ozbay, B.N.; Potcoava, M.; Salcedo, E.; Restrepo, D.; Gibson, E.A. Super-resolution imaging of ciliary microdomains in isolated olfactory sensory neurons using a custom two-color stimulated emission depletion microscope. J. Biomed. Opt. 2016, 21, 066017. [Google Scholar] [CrossRef] [Green Version]
- Wiltgen, S.M.; Smith, I.F.; Parker, I. Superresolution localization of single functional IP3R channels utilizing Ca2+ flux as a readout. Biophys. J. 2010, 99, 437–446. [Google Scholar] [CrossRef] [Green Version]
- Paczkowski, S. Insect Olfaction as an Information Filter for Chemo-Analytical Applications. Doctoral Dissertation, Georg-August-University Göttingen, Göttingen, Germany, 2014. [Google Scholar]
- Hummers, W.S.; Offeman, R.E. Preparation of Graphitic Oxide. J. Am. Chem. Soc. 1958, 80, 1339. [Google Scholar] [CrossRef]
- Reiner-Rozman, C.; Kotlowski, C.; Knoll, W. Electronic biosensing with functionalized rGO FETs. Biosensors 2016, 6, 17. [Google Scholar] [CrossRef] [Green Version]
- Predator, F.P. Olfactory Responses of Two Coleopteran Species: The Stored Product Pest Tribolium castaneum and the Forest Pest Predator Dastarcus helophoroides. Doctoral Dissertation, Georg-August-University Göttingen, Göttingen, Germany, 2019. [Google Scholar]
- Bonazza, C.; Zhu, J.; Hasler, R.; Mastrogiacomo, R.; Pelosi, P.; Knoll, W. Responses of the Pheromone-Binding Protein of the Silk Moth Bombyx mori on a Graphene Biosensor Match Binding Constants in Solution. Sensors 2021, 21, 499. [Google Scholar] [CrossRef] [PubMed]
- Dykyj, J.; Svoboda, J.; Wilhoit, R.C.; Frenkel, M.; Hall, K.R. Vapor Pressure and Antoine Constants for Oxygen Containing Organic Compounds; Springer: Berlin, Germany, 2000. [Google Scholar]
- Sander, R. Compilation of Henry’s law constants (version 4.0) for water as solvent. Atmos. Chem. Phys. 2015, 15, 4399–4981. [Google Scholar] [CrossRef] [Green Version]
- Rutter, G.M.; Jung, S.; Klimov, N.N.; Newell, D.B.; Zhitenev, N.B.; Stroscio, J.A. Microscopic polarization in bilayer graphene. Nat. Phys. 2011, 7, 649–655. [Google Scholar] [CrossRef]
- Macchia, E.; Alberga, D.; Manoli, K.; Mangiatordi, G.F.; Magliulo, M.; Palazzo, G.; Giordano, F.; Lattanzi, G.; Torsi, L. Organic bioelectronics probing conformational changes in surface confined proteins. Sci. Rep. 2016, 6, 28085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Kazzy, M.; Hurot, C.; Weerakkody, J.S.; Buhot, A.; Hou, Y. Biomimetic Olfactory Biosensors and Bioelectronic Noses; IFSA Publishing: Barcelona, Spain, 2020; pp. 15–63. [Google Scholar]
- Ricatti, J.; Acquasaliente, L.; Ribaudo, G.; De Filippis, V.; Bellini, M.; Llovera, R.E.; Barollo, S.; Pezzani, R.; Zagotto, G.; Persaud, K.C.; et al. Effects of point mutations in the binding pocket of the mouse major urinary protein MUP20 on ligand affinity and specificity. Sci. Rep. 2019, 9, 300. [Google Scholar] [CrossRef] [Green Version]
- Katada, S.; Hirokawa, T.; Oka, Y.; Suwa, M.; Touhara, K. Structural basis for a broad but selective ligand spectrum of a mouse olfactory receptor: Mapping the odorant-binding site. J. Neurosci. 2005, 25, 1806–1815. [Google Scholar] [CrossRef]
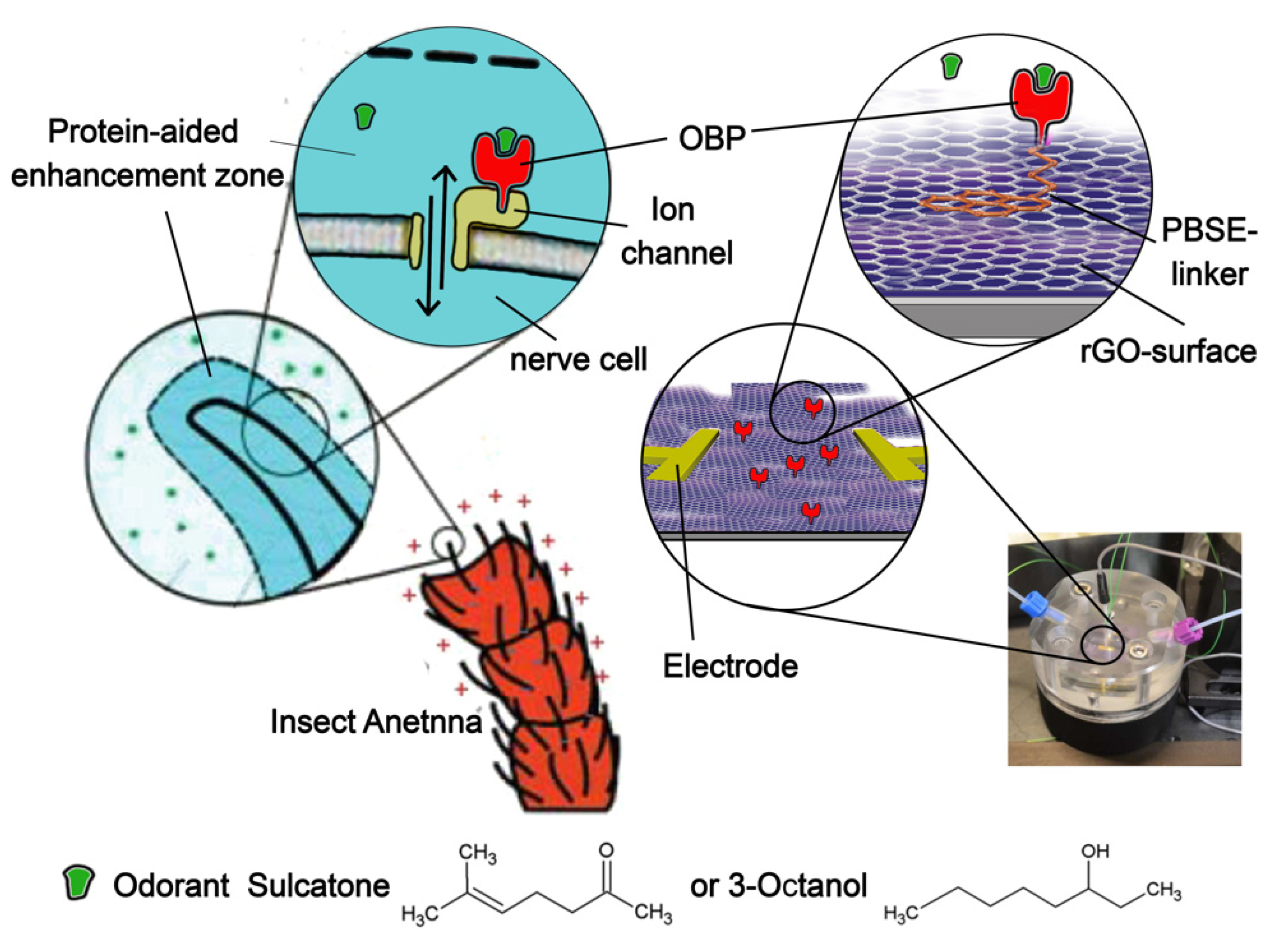
Figure 1.
Nerve cell receptor in the insect’s antenna, mimicked by our synthetic receptor. Left side: nerve cells in the insect’s sensilium are equipped with the ion-channel receptor TcOr1, which binds OBPs (which, in turn, bind odorants) and responds with ion signaling [22]. TcOr1 faces into a liquid lymphatic solution that constitutes a protein-aided enhancement (or enrichment) zone [11,12]. Right side: synthetic receptor. Instead of a nerve cell membrane containing ion channels, the signal is generated by a highly surface charge-sensitive graphene semiconductor surface. OBPs remain communicatively linked to the graphene, which is sustained by a Si/SiO substrate. In both cases, the response is read out electrically, with electrodes placed at the antenna (for electroantennograms) or on the graphene surface (for rGO-FET measurements). The structures of both odorants are depicted at the bottom.
Figure 1.
Nerve cell receptor in the insect’s antenna, mimicked by our synthetic receptor. Left side: nerve cells in the insect’s sensilium are equipped with the ion-channel receptor TcOr1, which binds OBPs (which, in turn, bind odorants) and responds with ion signaling [22]. TcOr1 faces into a liquid lymphatic solution that constitutes a protein-aided enhancement (or enrichment) zone [11,12]. Right side: synthetic receptor. Instead of a nerve cell membrane containing ion channels, the signal is generated by a highly surface charge-sensitive graphene semiconductor surface. OBPs remain communicatively linked to the graphene, which is sustained by a Si/SiO substrate. In both cases, the response is read out electrically, with electrodes placed at the antenna (for electroantennograms) or on the graphene surface (for rGO-FET measurements). The structures of both odorants are depicted at the bottom.

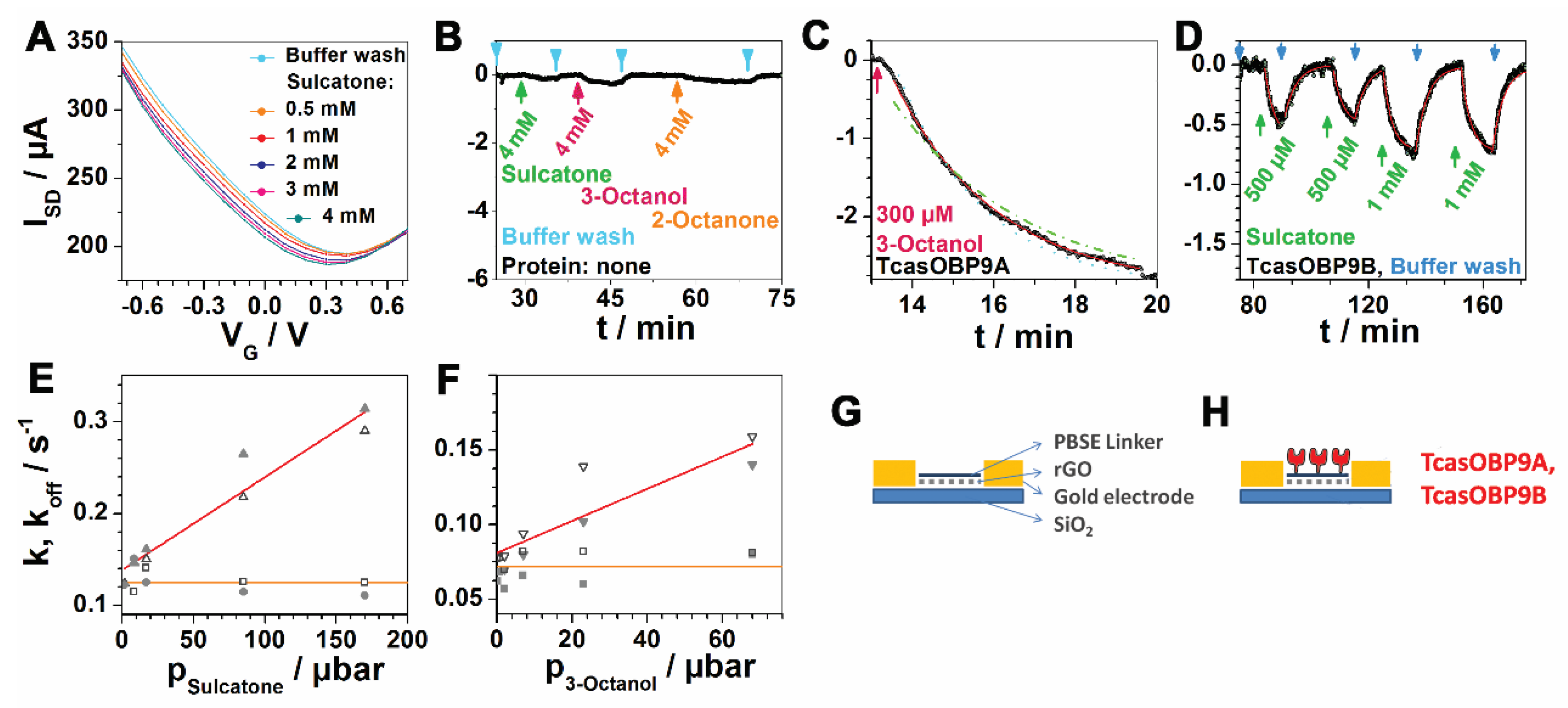
Figure 2.
Controls, EAGs, and setup. (A) ID-VG curves of sulcatone, as used to determine the VG measuring range. (B) Negative control of the plain FET (without protein), as shown in panel S. (C) Close-up of one titration step, showing the fit (red line) in a k ± 20% simulated error range, as described previously [14] (dotted and dashed-dotted lines). (D) Quantitative repeatability. The first and second, as well as the third and fourth, injections had the same odorant concentration and led to the same stimulus of the synthetic receptor. (E,F) Electroantennogram (EAG) data are shown as k/p-plots (red lines) and are reproduced courtesy of the authors of [25]. Orange lines show the non-concentration-dependent off-rates obtained from these EAGs. (G,H) Sketches of the setup.
Figure 2.
Controls, EAGs, and setup. (A) ID-VG curves of sulcatone, as used to determine the VG measuring range. (B) Negative control of the plain FET (without protein), as shown in panel S. (C) Close-up of one titration step, showing the fit (red line) in a k ± 20% simulated error range, as described previously [14] (dotted and dashed-dotted lines). (D) Quantitative repeatability. The first and second, as well as the third and fourth, injections had the same odorant concentration and led to the same stimulus of the synthetic receptor. (E,F) Electroantennogram (EAG) data are shown as k/p-plots (red lines) and are reproduced courtesy of the authors of [25]. Orange lines show the non-concentration-dependent off-rates obtained from these EAGs. (G,H) Sketches of the setup.

Figure 3.
Synthetic receptor kinetics. (A–D) The real-time response of synthetic receptors, set up with OBP9A or OBP9B (as shown in panel T) when binding stepwise increasing concentrations of sulcatone or 3-octanol. (E–H) Observed rate constants (k) of each concentration step, plotted against the concentration. Intersepts and slopes of the linear regressions yield kinetic off-rates koff and on-rates kon, respectively. (I–L) Binding equilibria (thermodynamics) of each OBP/odorant pair above. Responses to each concentration at saturation are plotted against the concentration. Langmuir fits (red curves) yielded the dissociation constants KD. Dotted and dashed-dotted lines indicate ± 20% simulated error ranges. The second (red) ordinate shows receptor occupation. All synthetic receptor responses were measured in triplicate.
Figure 3.
Synthetic receptor kinetics. (A–D) The real-time response of synthetic receptors, set up with OBP9A or OBP9B (as shown in panel T) when binding stepwise increasing concentrations of sulcatone or 3-octanol. (E–H) Observed rate constants (k) of each concentration step, plotted against the concentration. Intersepts and slopes of the linear regressions yield kinetic off-rates koff and on-rates kon, respectively. (I–L) Binding equilibria (thermodynamics) of each OBP/odorant pair above. Responses to each concentration at saturation are plotted against the concentration. Langmuir fits (red curves) yielded the dissociation constants KD. Dotted and dashed-dotted lines indicate ± 20% simulated error ranges. The second (red) ordinate shows receptor occupation. All synthetic receptor responses were measured in triplicate.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Odorant-binding kinetics of OBPs.
| Protein (Species) | Ligand | KD/μM | kon /M−1 s−1 | koff /s−1 |
|---|---|---|---|---|
| TcasOBP9A | 6-Methyl-5- hepten-2-one (Sulcatone) | 2100 | 2.9 | 0.008 |
| (Tribolium castaneum) | 3-Octanol | 2300 | 3.4 | 0.006 |
| TcasOBP9B | 6-Methyl-5- hepten-2-one (Sulcatone) | 3000 | 1.3 | 0.004 |
| (Tribolium castaneum) | 3-Octanol | 2900 | 2.4 | 0.006 |
| AmelOBP14 [13] | Homovanillic acid | 4 | 1130 | 0.008 |
| (Apis mellifera) | Methyl vanillate | 20 | 230 | 0.010 |
| Eugenol | 40 | 170 | 0.006 | |
| Citral | 800 | 9 | 0.003 | |
| Methyl eugenol | 1400 | 6 | 0.006 | |
| Geraniol | 3300 | 3 | 0.003 | |
| 6-Methyl-5- hepten-2-one (Sulcatone) | 3500 | 5 | 0.008 | |
| Amel SS-OBP14 [13] (Apis mellifera) | Eugenol | 4 | 750 | 0.006 |
| BmorPBP1 [26] | Bombykol (pH 8) | 5 | 20 | 0.000094 |
| (Bombyx mori) | Bombykol (pH 4) | - | no binding | no binding |
| Antenna EAG [25] | 6-Methyl-5- hepten-2-one (Sulcatone) | 1300 | 73 | 0.095 |
| (Tribolium castaenum) | 3-Octanol | 2500 | 32 | 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bonazza, C.; Bonazza, K. Kinetics of Odorant Recognition with a Graphene-Based Olfactory Receptor Mimicry. Chemosensors 2022, 10, 203. https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors10060203
AMA Style
Bonazza C, Bonazza K. Kinetics of Odorant Recognition with a Graphene-Based Olfactory Receptor Mimicry. Chemosensors. 2022; 10(6):203. https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors10060203
Chicago/Turabian StyleBonazza, Caroline, and Klaus Bonazza. 2022. "Kinetics of Odorant Recognition with a Graphene-Based Olfactory Receptor Mimicry" Chemosensors 10, no. 6: 203. https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors10060203
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.