
Detecting the Bitterness of Milk-Protein-Derived Peptides Using an Electronic Tongue

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Skim Milk
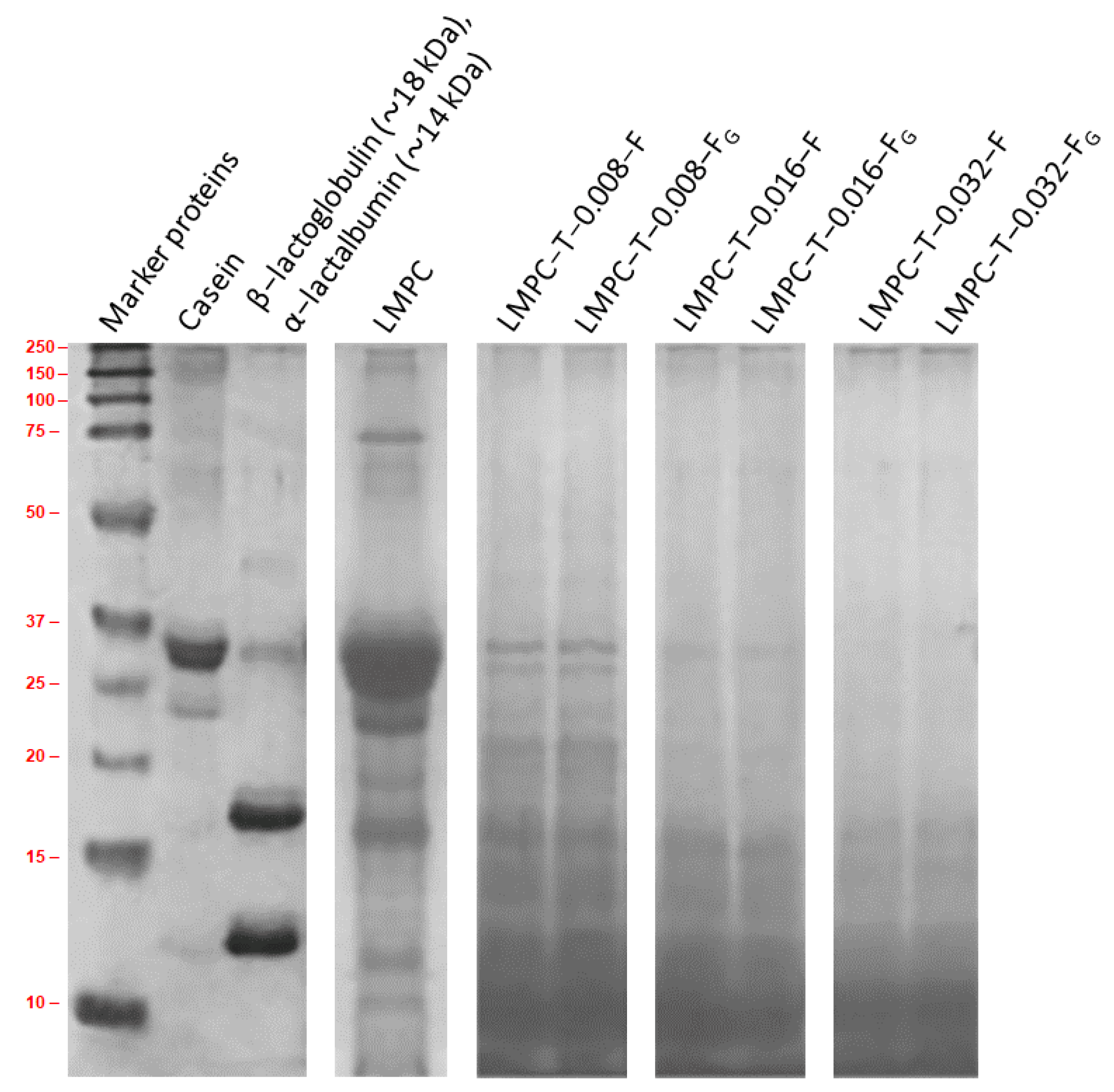
2.2. LMPC via a Membrane Filtration Process
2.3. Tryptic Digestion of Proteins in LMPC
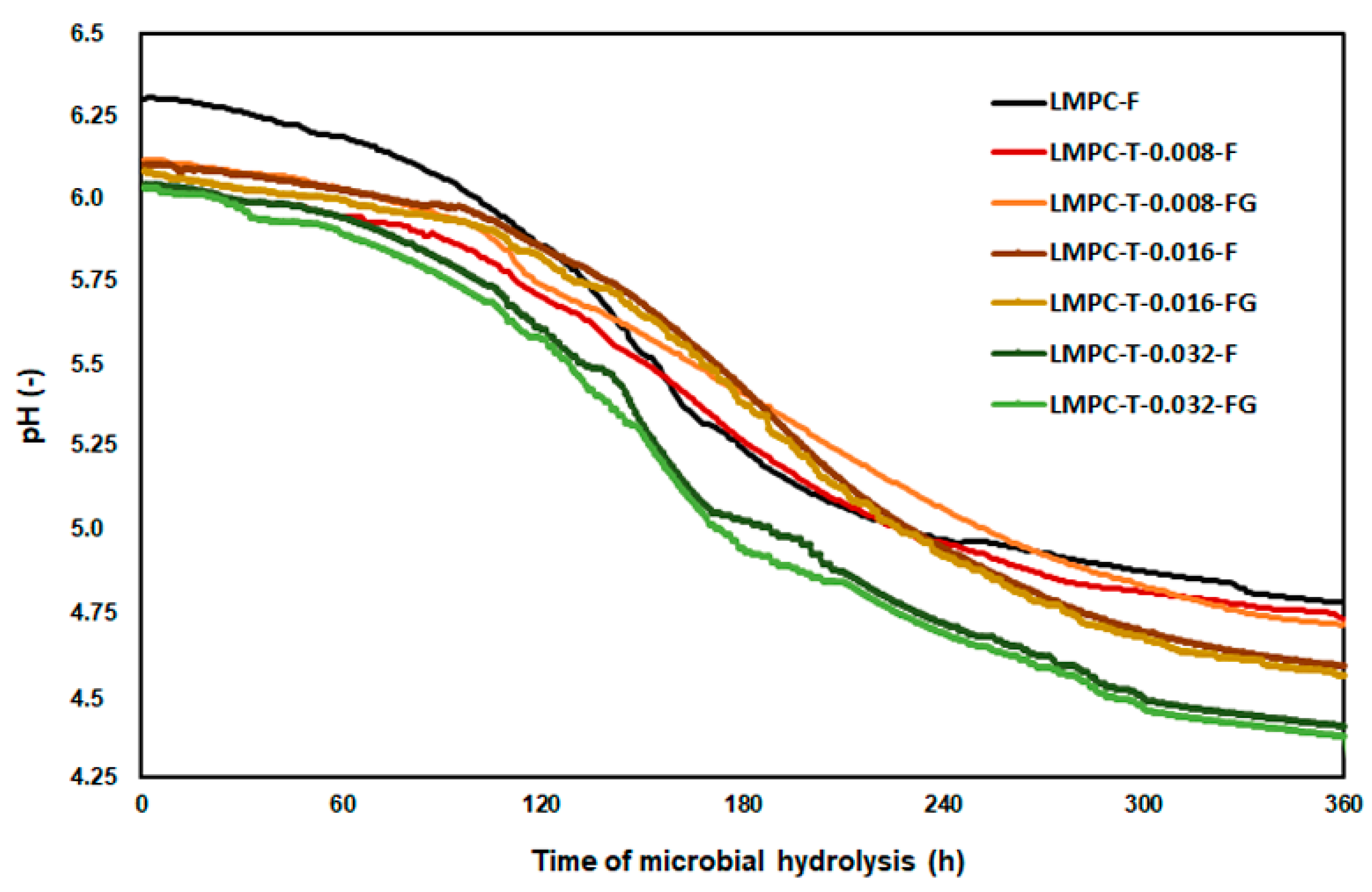
2.4. Microbial Hydrolysis
2.5. Sensory Analysis with Electronic Tongue
2.6. Statistical Analysis
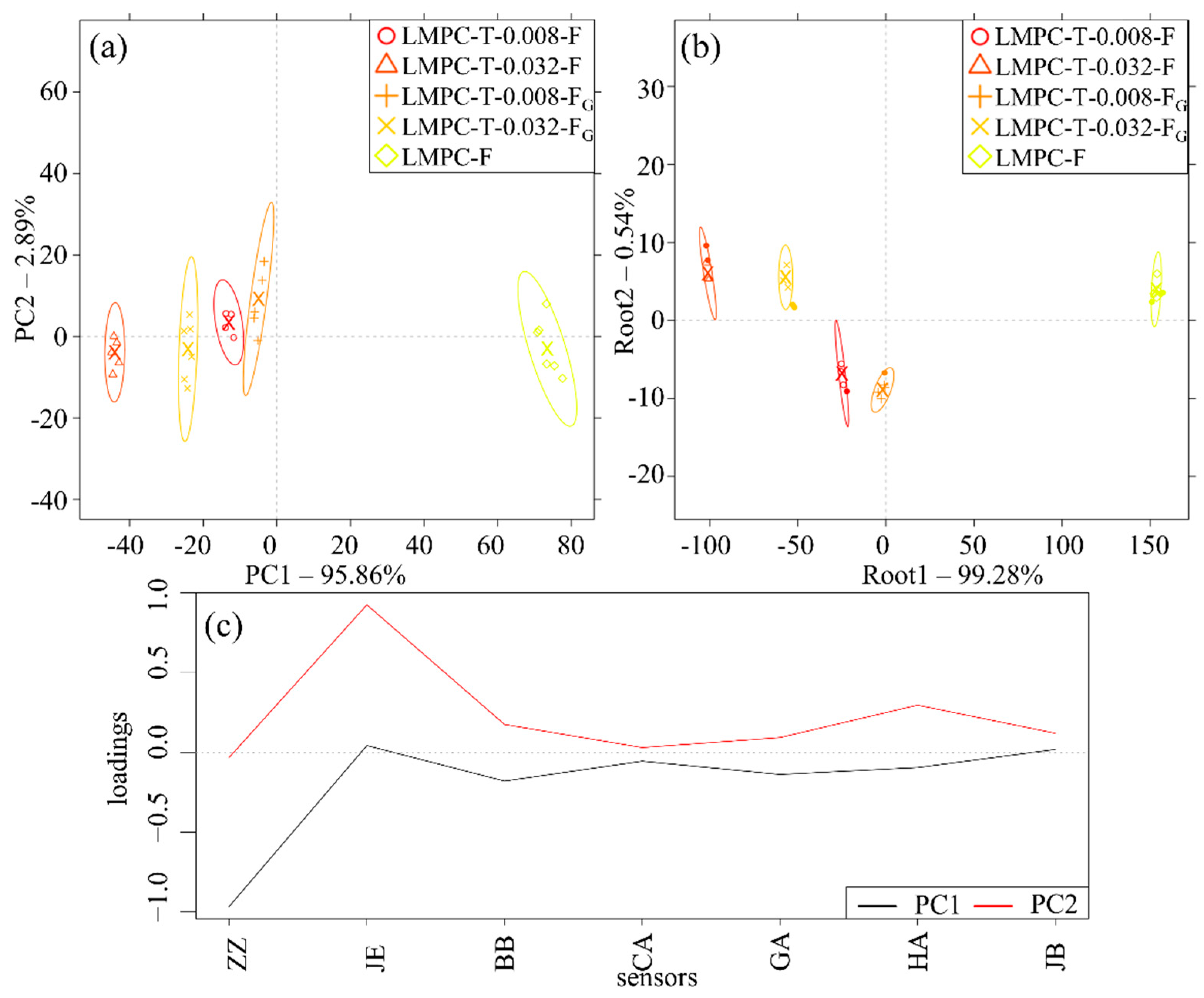
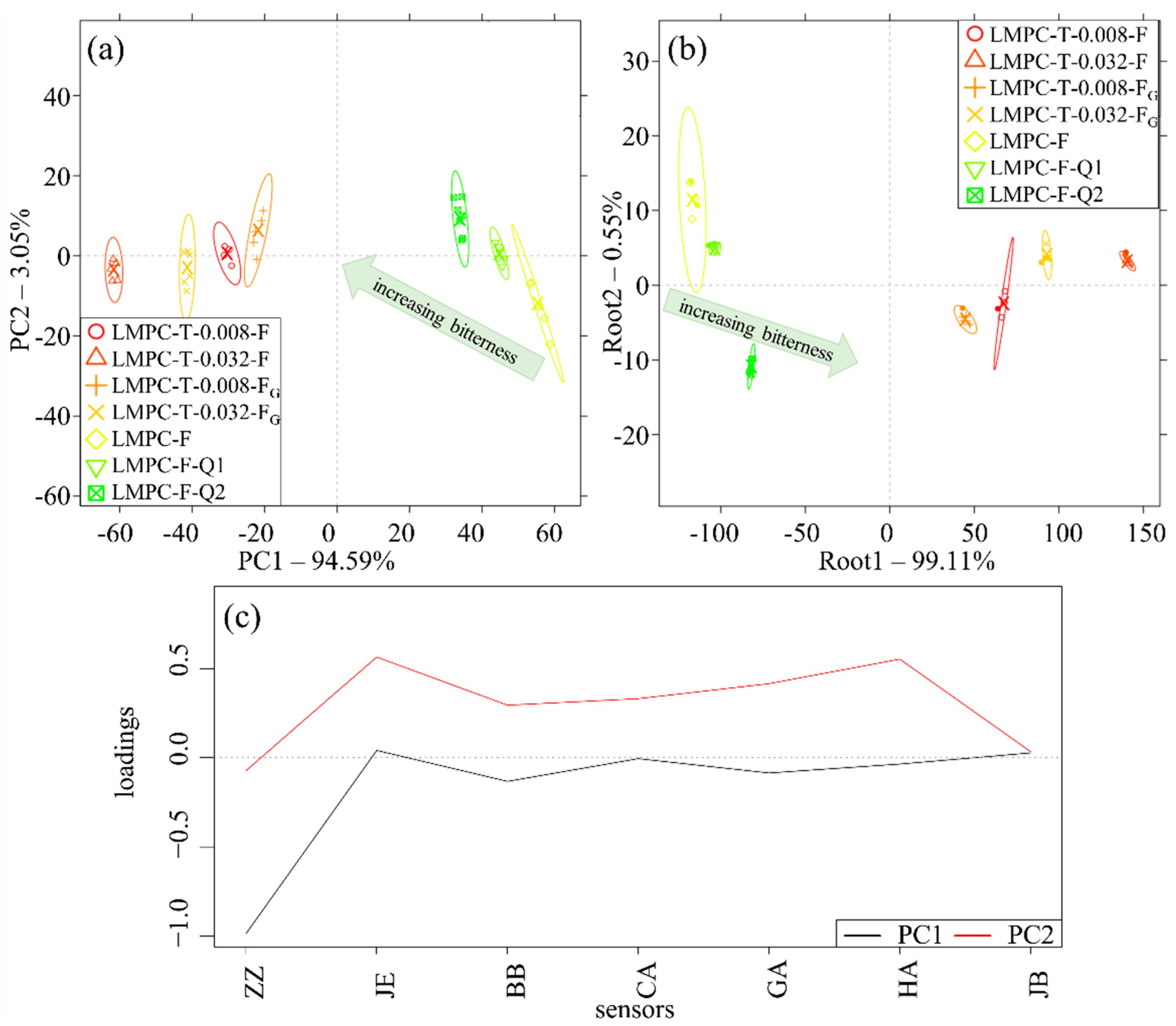
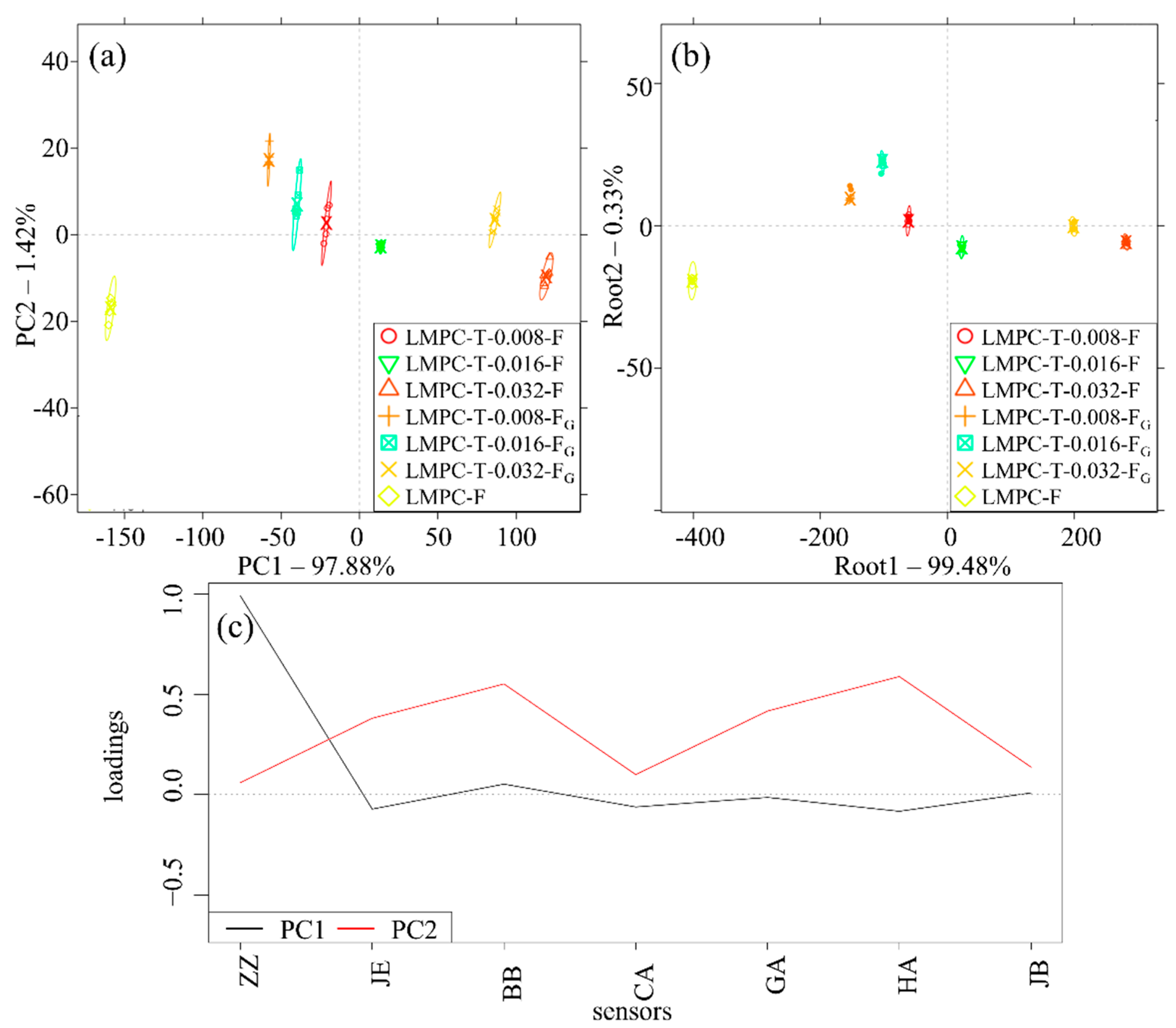
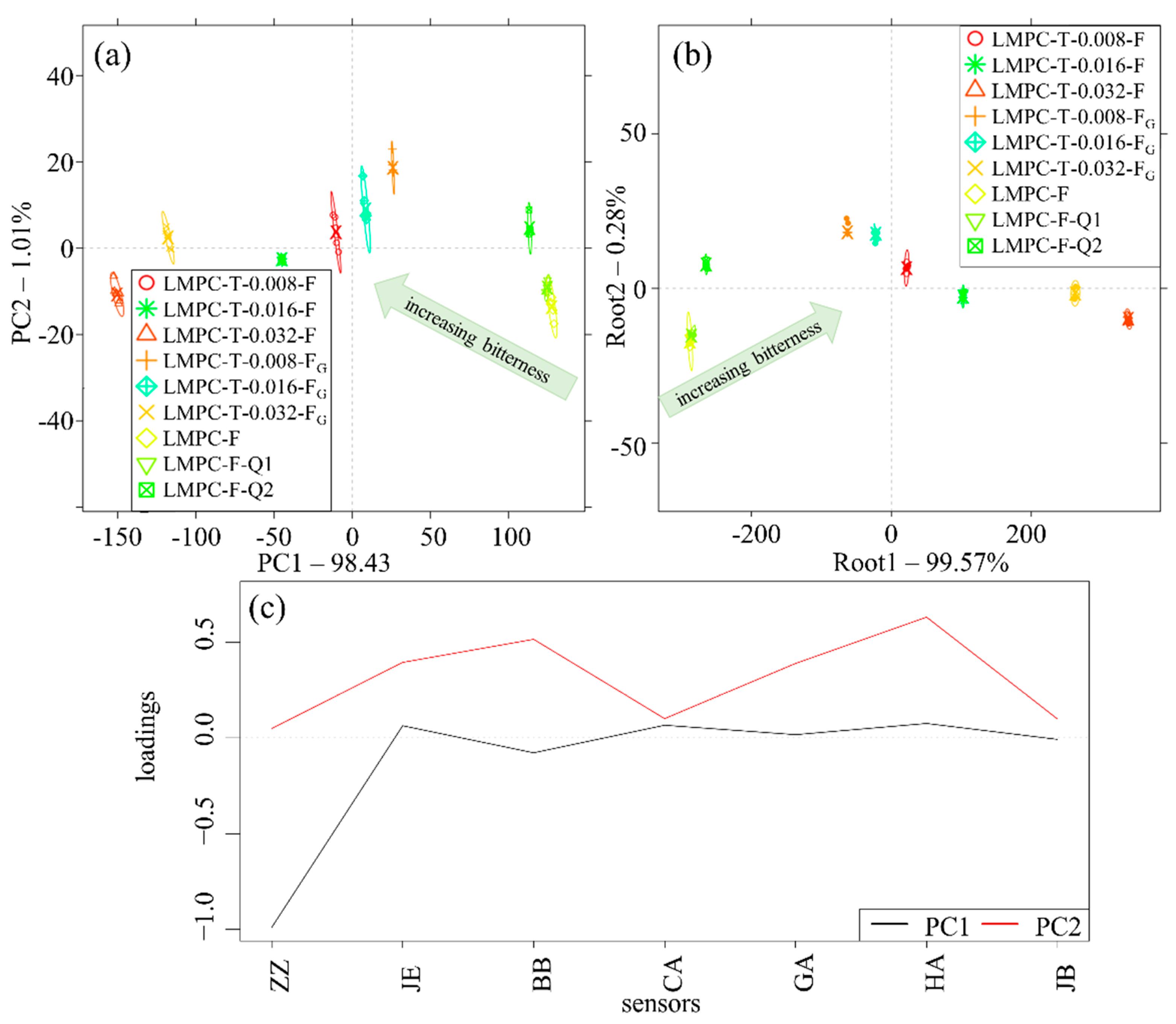
3. Results
4. Discussion
Justification of the Application of the E-Tongue for Evaluating Bitterness
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGregor, R.A.; Poppitt, S.D. Milk Protein for Improved Metabolic Health: A Review of the Evidence. Nutr. Metab. 2013, 10, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davoodi, S.H.; Shahbazi, R.; Esmaeili, S.; Sohrabvandi, S.; Mortazavian, A.; Jazayeri, S.; Taslimi, A. Health-Related Aspects of Milk Proteins. Iran. J. Pharm. Res. IJPR 2016, 15, 573–591. [Google Scholar] [PubMed]
- Patel, A.S.; Yeung, C.K.; Brennan, C.; Zheng, H. Ingredients and Formulation. Encycl. Dairy Sci. 2022, 663–673. [Google Scholar] [CrossRef]
- Meena, G.S.; Singh, A.K.; Panjagari, N.R.; Arora, S. Milk Protein Concentrates: Opportunities and Challenges. J. Food Sci. Technol. 2017, 54, 3010–3024. [Google Scholar] [CrossRef]
- Nath, A.; Eren, B.A.; Csighy, A.; Pastorne-Huszar, K.; Kisko, G.; Abranko, L.; Toth, A.; Szerdahelyi, E.; Kovacs, Z.; Koris, A.; et al. Production of Liquid Milk Protein Concentrate with Antioxidant Capacity, Angiotensin Converting Enzyme Inhibitory Activity, Antibacterial Activity, and Hypoallergenic Property by Membrane Filtration and Enzymatic Modification of Proteins. Processes 2020, 8, 871. [Google Scholar] [CrossRef]
- Feuille, E.; Nowak-Węgrzyn, A. Food Protein-Induced Enterocolitis Syndrome, Allergic Proctocolitis, and Enteropathy. Curr. Allergy Asthma Rep. 2015, 15, 50. [Google Scholar] [CrossRef]
- Walsh, J.; Meyer, R.; Shah, N.; Quekett, J.; Fox, A.T. Differentiating Milk Allergy (IgE and Non-IgE Mediated) from Lactose Intolerance: Understanding the Underlying Mechanisms and Presentations. Br. J. Gen. Pract. 2016, 66, e609–e611. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Qian, G.; Sun, J.; Yang, M.; Shi, X.; Yang, H.; Wu, J.; Wang, Z.; Zheng, Y.; Yue, X. Evaluation of Antigenicity and Nutritional Properties of Enzymatically Hydrolyzed Cow Milk. Sci. Rep. 2021, 11, 18623. [Google Scholar] [CrossRef]
- Abd El-Salam, M.H.; El-Shibiny, S. Preparation, Properties, and Uses of Enzymatic Milk Protein Hydrolysates. Crit. Rev. Food Sci. Nutr. 2017, 57, 1119–1132. [Google Scholar] [CrossRef]
- Patel, S. Functional Food Relevance of Whey Protein: A Review of Recent Findings and Scopes Ahead. J. Funct. Foods 2015, 19, 308–319. [Google Scholar] [CrossRef]
- Medeiros, V.; Rainha, N.; Paiva, L.; Lima, E.; Baptista, J. Bovine Milk Formula Based on Partial Hydrolysis of Caseins by Bromelain Enzyme: Better Digestibility and Angiotensin-Converting Enzyme-Inhibitory Properties. Int. J. Food Prop. 2014, 17, 806–817. [Google Scholar] [CrossRef]
- Li-Chan, E.C.Y. Bioactive Peptides and Protein Hydrolysates: Research Trends and Challenges for Application as Nutraceuticals and Functional Food Ingredients. Curr. Opin. Food Sci. 2015, 1, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Aouadi, B.; Zaukuu, J.L.Z.; Vitális, F.; Bodor, Z.; Fehér, O.; Gillay, Z.; Bazar, G.; Kovacs, Z. Historical Evolution and Food Control Achievements of near Infrared Spectroscopy, Electronic Nose, and Electronic Tongue—Critical Overview. Sensors 2020, 20, 5479. [Google Scholar] [CrossRef]
- Zaukuu, J.-L.Z.; Bazar, G.; Gillay, Z.; Kovacs, Z. Emerging Trends of Advanced Sensor Based Instruments for Meat, Poultry and Fish Quality– a Review. Crit. Rev. Food Sci. Nutr. 2019, 60, 3443–3460. [Google Scholar] [CrossRef] [PubMed]
- Escuder-gilabert, L.; Peris, M. Review: Highlights in Recent Applications of Electronic Tongues in Food Analysis. Anal. Chim. Acta 2010, 665, 15–25. [Google Scholar] [CrossRef]
- Wei, Z.; Zhang, W.; Wang, Y.; Wang, J. Monitoring the Fermentation, Post-Ripeness and Storage Processes of Set Yogurt Using Voltammetric Electronic Tongue. J. Food Eng. 2017, 203, 41–52. [Google Scholar] [CrossRef]
- Wei, Z.; Wang, J. Detection of Antibiotic Residues in Bovine Milk by a Voltammetric Electronic Tongue System. Anal. Chim. Acta 2011, 694, 46–56. [Google Scholar] [CrossRef]
- Bougrini, M.; Tahri, K.; Saidi, T.; El, N.; El, A.; Bouchikhi, B.; Bari, N. El Classification of Honey According to Geographical and Botanical Origins and Detection of Its Adulteration Using Voltammetric Electronic Tongue. Food Anal. Methods 2016, 9, 2161–2173. [Google Scholar] [CrossRef]
- de Morais, T.C.B.; Rodrigues, D.R.; de Carvalho Polari Souto, U.T.; Lemos, S.G. A Simple Voltammetric Electronic Tongue for the Analysis of Coffee Adulterations. Food Chem. 2019, 273, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Q.; Yang, Y.; Wu, J.; Liu, Z.; Chen, X.H.; Dong, C.; Chen, Q.; Zhang, Z.; Guo, Z. Rapid Sensing of Total Theaflavins Content in Black Tea Using a Portable Electronic Tongue System Coupled to Efficient Variables Selection Algorithms. J. Food Compos. Anal. 2019, 75, 43–48. [Google Scholar] [CrossRef]
- Martina, V.; Ionescu, K.; Pigani, L.; Terzi, F.; Ulrici, A.; Zanardi, C.; Seeber, R. Development of an Electronic Tongue Based on a PEDOT-Modified Voltammetric Sensor. Anal. Bioanal. Chem. 2007, 387, 2101–2110. [Google Scholar] [CrossRef] [PubMed]
- Apetrei, I.M.; Apetrei, C. Application of Voltammetric E-Tongue for the Detection of Ammonia and Putrescine in Beef Products. Sens. Actuators B Chem. 2016, 234, 371–379. [Google Scholar] [CrossRef]
- Ruiz-rico, M.; Fuentes, A.; Masot, R.; Alcañiz, M.; Fernández-segovia, I.; Manuel, J. Use of the Voltammetric Tongue in Fresh Cod (Gadus Morhua) Quality Assessment. Innov. Food Sci. Emerg. Technol. 2013, 18, 256–263. [Google Scholar] [CrossRef]
- Dias, L.A.; Peres, A.M.; Veloso, A.C.A.; Reis, F.S.; Vilas-Boas, M.; Machado, A.A.S.C. An Electronic Tongue Taste Evaluation: Identification of Goat Milk Adulteration with Bovine Milk. Sens. Actuators B Chem. 2009, 136, 209–217. [Google Scholar] [CrossRef]
- Li, D.; Peng, J.; Kwok, L.; Zhang, W.; Sun, T. Metabolomic Analysis of Streptococcus Thermophilus S10-Fermented Milk. LWT 2022, 161, 113368. [Google Scholar] [CrossRef]
- Lipkowitz, J.B.; Ross, C.F.; Diako, C.; Smith, D.M. Discriminating Aging and Protein-to-Fat Ratio in Cheddar Cheese Using Sensory Analysis and a Potentiometric Electronic Tongue. J. Dairy Sci. 2018, 101, 1990–2004. [Google Scholar] [CrossRef]
- Xu, M.; Wang, J.; Zhu, L. The Qualitative and Quantitative Assessment of Tea Quality Based on E-Nose, E-Tongue and E-Eye Combined with Chemometrics. Food Chem. 2019, 289, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Dias, L.G.; Veloso, A.C.A.; Sousa, M.E.B.C.; Estevinho, L.; Machado, A.A.S.C.; Peres, A.M. A Novel Approach for Honey Pollen Profile Assessment Using an Electronic Tongue and Chemometric Tools. Anal. Chim. Acta 2015, 900, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Hong, X.; Wang, J. Use of Electronic Nose and Tongue to Track Freshness of Cherry Tomatoes Squeezed for Juice Consumption: Comparison of Different Sensor Fusion Approaches. Food Bioprocess Technol. 2015, 8, 158–170. [Google Scholar] [CrossRef]
- Dong, W.; Hu, R.; Long, Y.; Li, H.; Zhang, Y.; Zhu, K.; Chu, Z. Comparative Evaluation of the Volatile Profiles and Taste Properties of Roasted Coffee Beans as Affected by Drying Method and Detected by Electronic Nose, Electronic Tongue, and HS-SPME-GC-MS. Food Chem. 2019, 272, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Gil, L.; Barat, J.M.; Baigts, D.; Martínez-máñez, R.; Soto, J.; Garcia-breijo, E.; Aristoy, M.; Toldrá, F.; Llobet, E. Monitoring of Physical—Chemical and Microbiological Changes in Fresh Pork Meat under Cold Storage by Means of a Potentiometric Electronic Tongue. Food Chem. 2011, 126, 1261–1268. [Google Scholar] [CrossRef]
- Han, F.; Huang, X.; Teye, E.; Haiyang, G. Quantitative Analysis of Fish Microbiological Quality Using Electronic Tongue Coupled with Nonlinear Pattern Recognition Algorithms. J. Food Saf. 2015, 35, 336–344. [Google Scholar] [CrossRef]
- Nath, A.; Csighy, A.; Eren, B.A.; Tjandra Nugraha, D.; Pásztorné-Huszár, K.; Tóth, A.; Takács, K.; Szerdahelyi, E.; Kiskó, G.; Kovács, Z.; et al. Bioactive Peptides from Liquid Milk Protein Concentrate by Sequential Tryptic and Microbial Hydrolysis. Processes 2021, 9, 1688. [Google Scholar] [CrossRef]
- Lorenzen, P.; MEISEL, H. Influence of Trypsin Action in Yoghurt Milk on the Release of Caseinophosphopeptide-rich Fractions and Physical Properties of the Fermented Products. Int. J. Dairy Technol. 2005, 58, 119–124. [Google Scholar] [CrossRef]
- Beshkova, D.; Simova, E.; Frengova, G.; Simov, Z. Production of Flavour Compounds by Yogurt Starter Cultures. J. Ind. Microbiol. Biotechnol. 1998, 20, 180–186. [Google Scholar] [CrossRef]
- Li, S.; Tang, S.; He, Q.; Hu, J.; Zheng, J. Changes in Proteolysis in Fermented Milk Produced by Streptococcus Thermophilus in Co-Culture with Lactobacillus Plantarum or Bifidobacterium Animalis Subsp. Lactis during Refrigerated Storage. Molecules 2019, 24, 3699. [Google Scholar] [CrossRef] [Green Version]
- Alpha MOS. Technical Note T-SAS-04 Astree Electrochemical Sensor Technology. 2009. Available online: www.alpha-mos.com (accessed on 1 June 2022).
- Kovács, Z.; Dalmadi, I.; Lukács, L.; László, S.; Szántai-Kohegyi, K.; Kókai, Z.; Fekete, A. Geographical Origin Identification of Pure Sri Lanka Tea Infusions with Electronic Nose, Electronic Tongue and Sensory Profile Analysis. J. Chemom. 2010, 24, 121–130. [Google Scholar] [CrossRef]
- Rachid, O.; Simons, F.E.R.; Rawas-Qalaji, M.; Simons, K.J. An Electronic Tongue: Evaluation of the Masking Efficacy of Sweetening and/or Flavoring Agents on the Bitter Taste of Epinephrine. AAPS PharmSciTech 2010, 11, 550–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemieux, L.; Simard, R.E. Bitter Flavour in Dairy Products. II. A Review of Bitter Peptides from Caseins: Their Formation, Isolation and Identification, Structure Masking and Inhibition. Lait 1992, 72, 335–385. [Google Scholar] [CrossRef]
- Breslin, P.A.S. Human Gustation and Flavour. Flavour Fragr. J. 2001, 16, 439–456. [Google Scholar] [CrossRef]
- Keast, R.S.J.; Roper, J. A Complex Relationship among Chemical Concentration, Detection Threshold, and Suprathreshold Intensity of Bitter Compounds. Chem. Senses 2007, 32, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Brereton, R.G. Exploratory Data Analysis. In Chemometrics for Pattern Recognition; John Wiley & Sons, Ltd.: Chichester, UK, 2009; pp. 47–106. ISBN 9780470746462. [Google Scholar]
- Berrueta, L.A.; Alonso-Salces, R.M.; Héberger, K. Supervised Pattern Recognition in Food Analysis. J. Chromatogr. A 2007, 1158, 196–214. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing 2017; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Matoba, T.; Hayashi, R.; Hata, T. Isolation of Bitter Peptides from Tryptic Hydrolysate of Casein and Their Chemical Structure. Agric. Biol. Chem. 1970, 34, 1235–1243. [Google Scholar] [CrossRef] [Green Version]
- PÉLISSIER, J.P.; Manchon, P. Comparative study of the bitter taste of enzymic hydrolysates from cow, ewe and goat caseins. J. Food Sci. 1976, 41, 231–233. [Google Scholar] [CrossRef]
- Abraham, A.G.; De Antoni, G.L.; Añon, M.C. Proteolytic Activity of Lactobacillus Bulgaricus Grown in Milk. J. Dairy Sci. 1993, 76, 1498–1505. [Google Scholar] [CrossRef]
- Liu, E.; Zheng, H.; Shi, T.; Ye, L.; Konno, T.; Oda, M.; Shen, H.; Ji, Z.-S. Relationship between Lactobacillus Bulgaricus and Streptococcus Thermophilus under Whey Conditions: Focus on Amino Acid Formation. Int. Dairy J. 2016, 56, 141–150. [Google Scholar] [CrossRef]
- Tzvetkova, I.; Dalgalarrondo, M.; Danova, S.; Iliev, I.; Ivanova, I.; Chobert, J.M.; Haertlé, T. Hydrolysis of Major Dairy Proteins by Lactic Acid Bacteria from Bulgarian Yogurts. J. Food Biochem. 2007, 31, 680–702. [Google Scholar] [CrossRef]
- Relkin, P.; Mulvihill, D.M. Thermal Unfolding of Β-lactoglobulin, A-lactalbumin, and Bovine Serum Albumin. A Thermodynamic Approach. Crit. Rev. Food Sci. Nutr. 1996, 36, 565–601. [Google Scholar] [CrossRef]
- Anema, S.G.; McKenna, A.B. Reaction Kinetics of Thermal Denaturation of Whey Proteins in Heated Reconstituted Whole Milk. J. Agric. Food Chem. 1996, 44, 422–428. [Google Scholar] [CrossRef]
- Anema, S.G. Heat-Induced Changes in Caseins and Casein Micelles, Including Interactions with Denatured Whey Proteins. Int. Dairy J. 2021, 122, 105136. [Google Scholar] [CrossRef]
- Dalgleish, D.G. Denaturation and Aggregation of Serum Proteins and Caseins in Heated Milk. J. Agric. Food Chem. 1990, 38, 1995–1999. [Google Scholar] [CrossRef]
- Gezimati, J.; Singh, H.; Creamer, L.K. Heat-Induced Interactions and Gelation of Mixtures of Bovine β-Lactoglobulin and Serum Albumin. J. Agric. Food Chem. 1996, 44, 804–810. [Google Scholar] [CrossRef]
- Manderson, G.A.; Hardman, M.J.; Creamer, L.K. Effect of Heat Treatment on the Conformation and Aggregation of β-Lactoglobulin A, B, and C. J. Agric. Food Chem. 1998, 46, 5052–5061. [Google Scholar] [CrossRef]
- Jang, H.D.; Swaisgood, H.E. Disulfide Bond Formation Between Thermally Denatured β-Lactoglobulin and κ-Casein in Casein Micelles. J. Dairy Sci. 1990, 73, 900–904. [Google Scholar] [CrossRef]
- Hong, Y.-H.; Creamer, L.K. Changed Protein Structures of Bovine β-Lactoglobulin B and α-Lactalbumin as a Consequence of Heat Treatment. Int. Dairy J. 2002, 12, 345–359. [Google Scholar] [CrossRef]
- Cheema, M.; Mohan, M.S.; Campagna, S.R.; Jurat-Fuentes, J.L.; Harte, F.M. The Association of Low-Molecular-Weight Hydrophobic Compounds with Native Casein Micelles in Bovine Milk. J. Dairy Sci. 2015, 98, 5155–5163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, W.G.; Groves, M.L. Primary Sequence of Beta, Gamma, and Minor Caseins. J. Dairy Sci. 1975, 58, 574–582. [Google Scholar] [CrossRef]
- Minamiura, N.; Matsumura, Y.; Fukumoto, J.; Yamamoto, T. Bitter Peptides in Cow Milk Casein Digests with Bacterial Proteinase Part I. Isolation and Determination of Amino Acid Sequence of a Bitter Peptide. Agric. Biol. Chem. 1972, 36, 588–595. [Google Scholar] [CrossRef]
- Bumberger, E.; Belitz, H.D. Bitter Taste of Enzymic Hydrolysates of Casein. I. Isolation, Structural and Sensorial Analysis of Peptides from Tryptic Hydrolysates of Beta-Casein. Z. Lebensm. Unters. Forsch. 1993, 197, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Kieliszek, M.; Pobiega, K.; Piwowarek, K.; Kot, A.M. Characteristics of the Proteolytic Enzymes Produced by Lactic Acid Bacteria. Molecules 2021, 26, 1858. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.W.; Nam, M.S. Bioactive Peptides in Milk and Dairy Products: A Review. Korean J. Food Sci. Anim. Resour. 2015, 35, 831–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courtin, P.; Monnet, V.; Rul, F. Cell-Wall Proteinases PrtS and PrtB Have a Different Role in Streptococcus Thermophilus/Lactobacillus Bulgaricus Mixed Cultures in Milk. Microbiology 2002, 148, 3413–3421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagliazucchi, D.; Martini, S.; Solieri, L. Bioprospecting for Bioactive Peptide Production by Lactic Acid Bacteria Isolated from Fermented Dairy Food. Fermentation 2019, 5, 96. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhang, Y.; Li, H.; Liu, X. The Potential of Proteins{,} Hydrolysates and Peptides as Growth Factors for Lactobacillus and Bifidobacterium: Current Research and Future Perspectives. Food Funct. 2020, 11, 1946–1957. [Google Scholar] [CrossRef]
- Shinoda, I.; Tada, M.; Okai, H.; Fukui, S. Bitter Taste of H-Pro-Phe-Pro-Gly-Pro-Ile-Pro-OH Corresponding to the Partial Sequence (Positions 61 ~ 67) of Bovine β -Casein, and Related Peptides. Agric. Biol. Chem. 1986, 50, 1247–1254. [Google Scholar] [CrossRef]
- Sullivan, J.J.; Jago, G.R. A Model for Bitter Peptide Formation and Degradation in Cultured Dairy Products. Aust. J. Dairy Technol. 1970, 25, 111. [Google Scholar]
- Sullivan, J.J.; Jago, G.R. Pyrroli-Donecarboxylyl Peptidase Activity in Streptococcus Cremoris ML1. Aust. J. Dairy Technol. 1970, 25, 141. [Google Scholar]
- Chen, C.; Zhao, S.; Hao, G.; Yu, H.; Tian, H.; Zhao, G. Role of Lactic Acid Bacteria on the Yogurt Flavour: A Review. Int. J. Food Prop. 2017, 20, S316–S330. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Zhang, C.; Li, S.; Zhang, Y.; Yang, Z. Growth and Exopolysaccharide Production by Streptococcus Thermophilus ST1 in Skim Milk. Brazilian J. Microbiol. 2011, 42, 1470–1478. [Google Scholar] [CrossRef] [Green Version]
- Malaka, R.; Maruddin, F.; Dwyana, Z.; Vargas, M.V. Assessment of Exopolysaccharide Production by Lactobacillus Delbrueckii Subsp. Bulgaricus Ropy Strain in Different Substrate Media. Food Sci. Nutr. 2020, 8, 1657–1664. [Google Scholar] [CrossRef]
- Kim, M.-R.; Choi, S.-Y.; Lee, C.-H. Molecular Characterization and Bitter Taste Formation of Tryptic Hydrolysis of 11S Glycinin. J. Microbiol. Biotechnol. 1999, 9, 509–513. [Google Scholar]
- Bouchier, P.J.; O’Cuinn, G.; Harrington, D.; FitzGerald, R.J. Debittering and Hydrolysis of a Tryptic Hydrolysate of β-Casein with Purified General and Proline Specific Aminopeptidases from Lactococcus Lactis Ssp. Cremoris AM2. J. Food Sci. 2001, 66, 816–820. [Google Scholar] [CrossRef]
- Shimamura, T.; Nishimura, T.; Iwasaki, A.; Odake, S.; Akuzawa, R. Degradation of a Bitter Peptide Derived from Casein by Lactic Acid Bacterial Peptidase. Food Sci. Technol. Res. 2009, 15, 191–194. [Google Scholar] [CrossRef] [Green Version]
- Savijoki, K.; Ingmer, H.; Varmanen, P. Proteolytic Systems of Lactic Acid Bacteria. Appl. Microbiol. Biotechnol. 2006, 71, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Espla, M.D.; Rul, F. PepS from Streptococcus Thermophilus. A New Member of the Aminopeptidase T Family of Thermophilic Bacteria. Eur. J. Biochem. 1999, 263, 502–510. [Google Scholar] [CrossRef] [Green Version]
- López-Fandiño, R.; Ardö, Y. Effect of Heat Treatment on the Proteolytic/Peptidolytic Enzyme System of a Lactobacillus Delbrueckii Subsp. Bulgaricus Strain. J. Dairy Res. 1991, 58, 469–475. [Google Scholar] [CrossRef]
- Rival, S.G.; Boeriu, C.G.; Wichers, H.J. Caseins and Casein Hydrolysates. 2. Antioxidative Properties and Relevance to Lipoxygenase Inhibition. J. Agric. Food Chem. 2001, 49, 295–302. [Google Scholar] [CrossRef]
- Power, O.; Fernández, A.; Norris, R.; Riera, F.A.; FitzGerald, R.J. Selective Enrichment of Bioactive Properties during Ultrafiltration of a Tryptic Digest of β-Lactoglobulin. J. Funct. Foods 2014, 9, 38–47. [Google Scholar] [CrossRef]
- Ballatore, M.B.; Bettiol, M. del R.; Vanden Braber, N.L.; Aminahuel, C.A.; Rossi, Y.E.; Petroselli, G.; Erra-Balsells, R.; Cavaglieri, L.R.; Montenegro, M.A. Antioxidant and Cytoprotective Effect of Peptides Produced by Hydrolysis of Whey Protein Concentrate with Trypsin. Food Chem. 2020, 319, 126472. [Google Scholar] [CrossRef]
- Tonolo, F.; Fiorese, F.; Moretto, L.; Folda, A.; Scalcon, V.; Grinzato, A.; Ferro, S.; Arrigoni, G.; Bindoli, A.; Feller, E.; et al. Identification of New Peptides from Fermented Milk Showing Antioxidant Properties: Mechanism of Action. Antioxidants 2020, 9, 117. [Google Scholar] [CrossRef] [Green Version]
- Matoba, T.; Hata, T. Relationship between Bitterness of Pepticles and Their Chemical Structures. Agric. Biol. Chem. 1972, 36, 1423–1431. [Google Scholar] [CrossRef]
- Apak, R.; Özyürek, M.; Güçlü, K.; Çapanoğlu, E. Antioxidant Activity/Capacity Measurement. 1. Classification, Physicochemical Principles, Mechanisms, and Electron Transfer (ET)-Based Assays. J. Agric. Food Chem. 2016, 64, 997–1027. [Google Scholar] [CrossRef] [PubMed]
- Sabeena Farvin, K.H.; Baron, C.P.; Nielsen, N.S.; Jacobsen, C. Antioxidant Activity of Yoghurt Peptides: Part 1-in Vitro Assays and Evaluation in ω-3 Enriched Milk. Food Chem. 2010, 123, 1081–1089. [Google Scholar] [CrossRef]
- Sabeena Farvin, K.H.; Baron, C.P.; Nielsen, N.S.; Otte, J.; Jacobsen, C. Antioxidant Activity of Yoghurt Peptides: Part 2—Characterisation of Peptide Fractions. Food Chem. 2010, 123, 1090–1097. [Google Scholar] [CrossRef]
- Tan, P.S.; van Kessel, T.A.; van de Veerdonk, F.L.; Zuurendonk, P.F.; Bruins, A.P.; Konings, W.N. Degradation and Debittering of a Tryptic Digest from Beta-Casein by Aminopeptidase N from Lactococcus Lactis Subsp. Cremoris Wg2. Appl. Environ. Microbiol. 1993, 59, 1430–1436. [Google Scholar] [CrossRef] [Green Version]
- Jaggupilli, A.; Howard, R.; Upadhyaya, J.D.; Bhullar, R.P.; Chelikani, P. Bitter Taste Receptors: Novel Insights into the Biochemistry and Pharmacology. Int. J. Biochem. Cell Biol. 2016, 77, 184–196. [Google Scholar] [CrossRef]
- Upadhyaya, J.; Pydi, S.P.; Singh, N.; Aluko, R.E.; Chelikani, P. Bitter Taste Receptor T2R1 Is Activated by Dipeptides and Tripeptides. Biochem. Biophys. Res. Commun. 2010, 398, 331–335. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Habara, M.; Ikezazki, H.; Chen, R.; Naito, Y.; Toko, K. Advanced Taste Sensors Based on Artificial Lipids with Global Selectivity to Basic Taste Qualities and High Correlation to Sensory Scores. Sensors 2010, 10, 3411–3443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyanaga, Y.; Tanigake, A.; Nakamura, T.; Kobayashi, Y.; Ikezaki, H.; Taniguchi, A.; Matsuyama, K.; Uchida, T. Prediction of the Bitterness of Single, Binary- and Multiple-Component Amino Acid Solutions Using a Taste Sensor. Int. J. Pharm. 2002, 248, 207–218. [Google Scholar] [CrossRef]
- Ley, J.P. Masking Bitter Taste by Molecules. Chemosens. Percept. 2008, 1, 58–77. [Google Scholar] [CrossRef]
- Liu, B.Y.; Zhu, K.X.; Peng, W.; Guo, X.N.; Zhou, H.M. Effect of Sequential Hydrolysis with Endo- and Exo-Peptidase on Bitterness Properties of Wheat Gluten Hydrolysates. RSC Adv. 2016, 6, 27659–27668. [Google Scholar] [CrossRef]
- Bleibaum, R.N.; Stone, H.; Tan, T.; Labreche, S.; Saint-Martin, E.; Isz, S. Comparison of Sensory and Consumer Results with Electronic Nose and Tongue Sensors for Apple Juices. Food Qual. Prefer. 2002, 13, 409–422. [Google Scholar] [CrossRef]
- Newman, J.; Egan, T.; Harbourne, N.; O’Riordan, D.; Jacquier, J.C.; O’Sullivan, M. Correlation of Sensory Bitterness in Dairy Protein Hydrolysates: Comparison of Prediction Models Built Using Sensory, Chromatographic and Electronic Tongue Data. Talanta 2014, 126, 46–53. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Antioxidant Capacity (mg eqv. Ascorbic Acid·L−1) | Samples | Antioxidant Capacity (mg eqv. Ascorbic Acid·L−1) |
|---|---|---|---|
| LMPC-T-0.008-F | 122.16 ± 20.25 A | LMPC-T-0.008-FG | 99.08 ± 18.61 a |
| LMPC-T-0.016-F | 131.75 ± 27.42 A | LMPC-T-0.016-FG | 108.22 ± 28.76 a |
| LMPC-T-0.032-F | 147.34 ± 26.68 a | LMPC-T-0.032-FG | 128.31 ± 32.04 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nath, A.; Eren, B.A.; Zinia Zaukuu, J.-L.; Koris, A.; Pásztorné-Huszár, K.; Szerdahelyi, E.; Kovacs, Z. Detecting the Bitterness of Milk-Protein-Derived Peptides Using an Electronic Tongue. Chemosensors 2022, 10, 215. https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors10060215
Nath A, Eren BA, Zinia Zaukuu J-L, Koris A, Pásztorné-Huszár K, Szerdahelyi E, Kovacs Z. Detecting the Bitterness of Milk-Protein-Derived Peptides Using an Electronic Tongue. Chemosensors. 2022; 10(6):215. https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors10060215
Chicago/Turabian StyleNath, Arijit, Burak Atilla Eren, John-Lewis Zinia Zaukuu, András Koris, Klára Pásztorné-Huszár, Emőke Szerdahelyi, and Zoltan Kovacs. 2022. "Detecting the Bitterness of Milk-Protein-Derived Peptides Using an Electronic Tongue" Chemosensors 10, no. 6: 215. https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors10060215