From Gas Sensors to Biomimetic Artificial Noses

Austrian Institute of Technology GmbH, Biosensor Technologies, Konrad-Lorenzstraße 24, 3430 Tulln, Austria
*
Author to whom correspondence should be addressed.
Chemosensors 2018, 6(3), 32; https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors6030032
Submission received: 2 July 2018
/
Revised: 29 July 2018
/
Accepted: 3 August 2018
/
Published: 7 August 2018
(This article belongs to the Special Issue Electronic nose’s, Machine Olfaction and Electronic Tongue’s)
Abstract
:Since the first attempts to mimic the human nose with artificial devices, a variety of sensors have been developed, ranging from simple inorganic and organic gas detectors to biosensing elements incorporating proteins of the biological olfactory system. In order to design a device able to mimic the human nose, two major issues still need to be addressed regarding the complexity of olfactory coding and the extreme sensitivity of the biological system. So far, only 50 of the approximately 300–400 functioning olfactory receptors have been de-orphanized, still a long way from breaking the human olfactory code. On the other hand, the exceptional sensitivity of the human nose is based on amplification mechanisms difficult to reproduce with electronic circuits, and perhaps novel approaches are required to address this issue. Here, we review the recent literature on chemical sensing both in biological systems and artificial devices, and try to establish the state-of-the-art towards the design of an electronic nose.
1. Introduction
Parallel to our progress in understanding olfaction at the physiological level, attempts have been proposed to model our olfactory system with artificial devices. At about the same time as biochemical research was applied to olfaction with the discovery of odorant-binding proteins [1,2,3], the first attempt to mimic the human nose with an array of metal oxide-gas sensors was proposed [4].
Since then, a rapidly increasing number of publications have been exploring the possibility of measuring odours with artificial devices. The designed systems range from individual gas sensors to complex arrays, incorporating inorganic, organic and biological sensing elements into electronic, optical and other types of devices [5,6,7]. However, after nearly four decades, despite the large amount of information currently available on the biochemical mechanisms of odour perception, and the tremendous advances in electronics and computing, we are still very far from designing a general-purpose instrument able to analyse complex mixtures of gases in the way the human nose does. As a result, reliable instruments for general use are not yet available neither for industrial applications nor for research uses. Nevertheless, when we look at specific problems and focused approaches, a large variety of solutions are being proposed, tailored to selected applications in the fields of food quality, environmental monitoring and clinical diagnostics [8,9,10,11,12,13].
In this review, starting from our knowledge of the olfactory system, we shall try and identify which elements are most important for an artificial nose and what kind of strategies we can learn from physiology to meet our technological problems. Within such a perspective, we shall then analyse current solutions adopted so far for chemical analysis in the gas phase and summarise the state-of-the-art on gas sensing technologies. Finally, returning to the physiological olfactory system, we shall discuss what is still needed in terms of information and to what extent the biological system can be adapted to electronic devices, suggesting future trends in artificial smell detection and identification.
2. Information from the Olfactory System
When we plan to assemble an instrument for odour discrimination and recognition, using the approach of the human nose—what we would call an artificial nose—we are confronted with at least three major types of challenges: complexity of the olfactory code, limited knowledge of the biological system and the high sensitivity of the human nose. In the following sections, we shall appreciate the exceptional variety of natural odours and the complexity of the physiological olfactory system; we shall discover how largely ignorant we are of the functioning of our nose and the complexity of the olfactory code; we shall realise that in terms of analytical instruments we are still very far from attaining the sensitivity of the human nose, let alone that of an insect antenna.
2.1. Complexity of the Olfactory Code
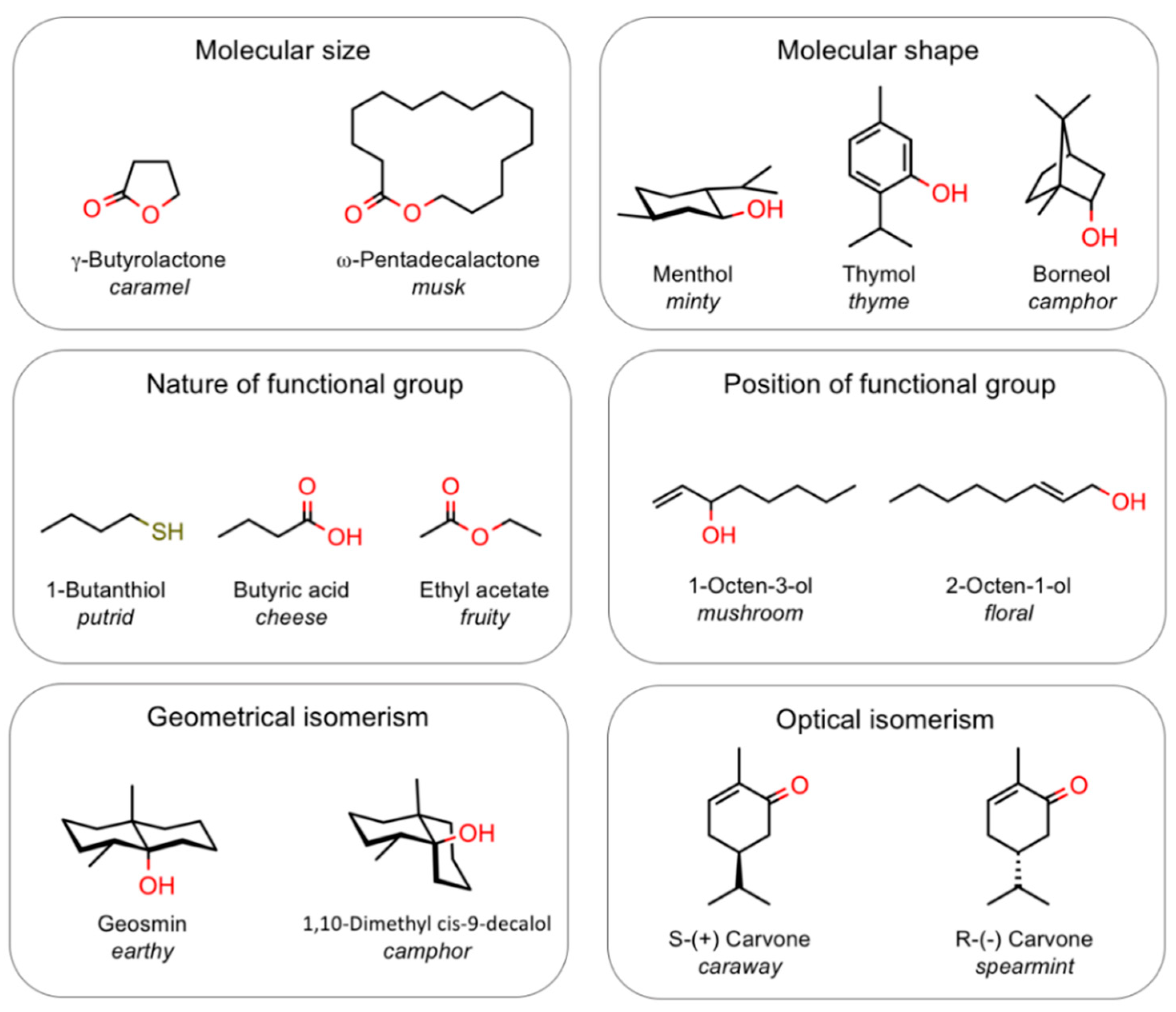
Perhaps the characteristic best describing the olfactory system is its complexity. This complexity, which prevented its study until recently, can be appreciated at different levels of information and processing. At the chemical level, the molecules in the gas phase, representing the olfactory stimuli, come in a very large variety of structures; moreover, natural odours are almost always produced by complex mixtures of chemicals, where both composition and relative concentrations are important to define the right odour character, but even simple stimuli, composed of a single type of molecules, are difficult to describe and quantify [14,15]. When we try to establish relationships between chemical structure and odour, we discover that several different parameters contribute to the odour character. Unlike colours or sounds, both related to a single variable (the wavelength), odours depend on nature and position of functional groups, but also on size and shape of the molecule. This last property, in particular, is difficult to define in quantitative terms. In fact, molecules can be linear or branched chains, rings from flat to almost spherical, and the occurrence of groups and side chains can greatly affect the overall shape of the structure. Moreover, often geometrical isomers and sometimes even enantiomers present different odours. The examples reported in Figure 1 help to illustrate such concepts.
To cope with such complex and diverse variety of chemical structures, the human olfactory nose is equipped with more than 300 receptors [16,17,18,19,20]. Such a large number of sensors represent an additional level of complexity, although still inadequate when compared with the thousands of different molecules present in the environment and the millions of possible combinations. To discriminate between millions of odours with only few hundred sensors, the strategy used by the nose is that of the combinatorial code, where each receptor is sensitive at different levels to a variety of chemicals, while each odorant can stimulate to different extents several receptors [21,22]. Reproducing such complex system in an artificial device poses high challenges. The colour vision works on the same principle, but being much simpler (three basic elements instead of 300), its reproduction in artificial devices did not pose major problems, and cameras and optical detectors able to analyse colours with high accuracy have been in use for long time. On the other hand, the auditory system uses thousands of receptors, each tuned to a single wavelength, but such apparent complexity is balanced by the fact that sounds are recognised on the basis of a single variable, the wavelength, just like colours. As a result, microphones can accurately reproduce any type of sound and have been in common use for many decades. By contrast, chemical recognition is based on multidimensional measurements, whose parameters are difficult to define [14,15].
There is also complexity at the processing level. The responses of the 300 olfactory receptors generate in the olfactory bulbs unique odour pictures, that can be actually visualised using fluorescence dyes and imaging systems [23]. Such pictures, however, are further processed in the brain and integrated with inputs from other sensory modalities, as well as compared with memories of past experience [24,25]. Finally, the smell produces a sensation that becomes expressed through verbal description, behavioural responses and emotions. Although now we know a great deal on how peripheral olfactory signals are integrated at the level of the olfactory bulbs, we are still very ignorant on the complex neural network through which these messages reach other regions of the brain and what kind of processing they undergo before being consciously perceived.
2.2. Knowledge of the Biological System
The basic type of knowledge essential for modelling the human nose can be regarded as the olfactory code. In analogy with the colour code, we can define the olfactory code as the list of elementary odours that can be combined to produce the innumerable different sensations we experience every day. These odours could be represented by chemical structures, each one being the best ligand for each olfactory receptor. Therefore, breaking the olfactory code corresponds to deciphering the specificity of each of our 300 olfactory receptors.
The first attempt to systematically break the olfactory code was reported by John Amoore, who applied the study of specific anosmias to this task [26,27,28]. The phenomenon of specific anosmia, first discovered by Guillot [29] and extensively described by Le Magnen [30] is the inability of some subjects of the human population to detect one or more odours. It is equivalent to colour blindness (or daltonism) and is related to the absence or the malfunctioning of one or more olfactory receptors. However, unlike daltonism, specific anosmia is much more frequent in the human population. Moreover, the occurrence of such defects can be very high, depending on the type of smell. As an example, anosmia to the urinous odour 5α-androst-16-en-3-one affects almost half of the human population [31]. Based on a compilation of specific anosmias identified in the human population, Amoore proposed a list of about 30 basic odorants, to be regarded as “primary odours” [27].
Much later, Linda Buck and Richard Axel, using the tools of molecular biology, discovered a very large gene family coding for several hundreds of olfactory receptors in genomes of mammals [32]. In the human genome a total of 857 OR genes have been found. More than half of them are pseudogenes, leaving 391 potentially functional receptors [20,33], thus increasing by more than one order of magnitude the first prediction of Amoore on the number of primary odours.
A parallel type of information comes from recent transcriptome projects aimed at identifying RNA sequences encoding olfactory receptors and actually present in the nose [33,34]. From such studies we learn two important facts: (1) the expression of olfactory receptors at the RNA level can be highly variable between genes; (2) the repertoire of olfactory genes in the human nose is different between individuals, providing a molecular evidence to the phenomenon of specific anosmias observed earlier in the human population.
A profiling study [34], performed on 26 human subjects, revealed that only 90 OR genes were expressed in the noses of all 26 individuals, while 140 additional genes were found in half of the subjects, the total number of genes identified in all 26 subjects being 343. For a first simplified model of the human nose, we can assume an olfactory code based on 90 ORs, while a better approach could include the additional genes detected in half of the subject analysed reaching a total of 230 ORs. The high percentage of OR genes present only in a part of the human population provides an idea on the wide occurrence of specific anosmias. At the same time, it is difficult to define a set of genes typically expressed in the average human individual, in order to list the basic odours that an artificial nose should be able to smell.
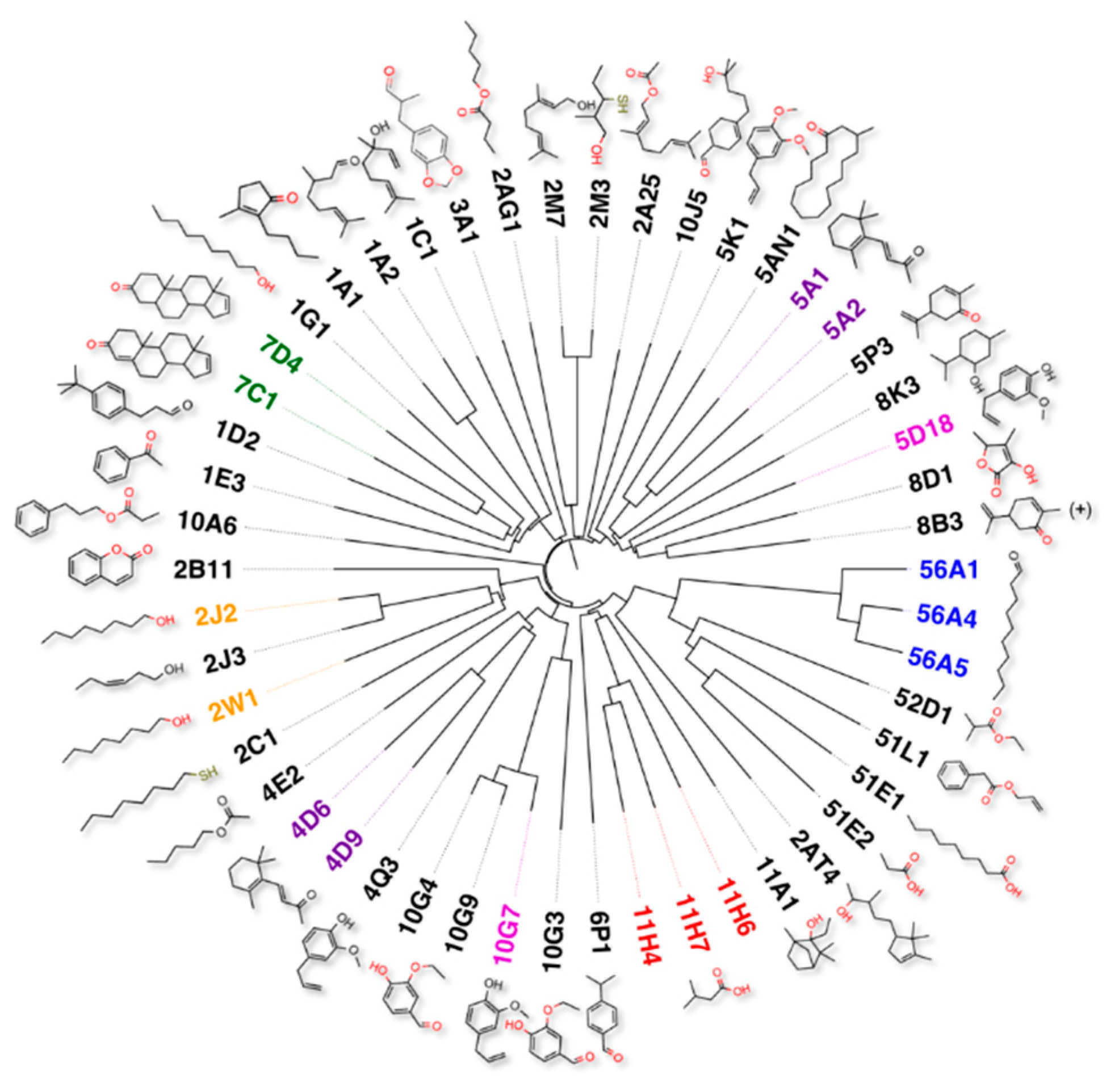
These figures give us a first idea on the complexity of the olfactory code. However, we might find in the end that the number of primary odours can be further reduced, when we account for the redundancy of the olfactory code. In fact, looking at the 50 human ORs which have been “deorphanised”, i.e., for which ligands have been identified, we often find two or three ORs tuned to the same molecular structures. This can be easily appreciated from the phylogenetic tree of Figure 2, where these 50 ORs are reported together with the structures of their best ligands. The redundancy observed for several of them probably represents a measure to ensure that in spite of random mutations that might switch off some receptors, we would still be able to detect some important odours. The lesson to learn, in this case, is that redundancy of sensors tuned to the same smells can be useful in an artificial nose to cope with the expected deterioration and poisoning of sensing elements exposed all the time to the environment.
2.3. Sensitivity
One of the great challenges in reproducing the functioning of biological noses is to match their exceptional sensitivity. Even the human nose, whose performance if far below that of an insect antenna, can detect volatile molecules at concentrations lower than any analytical instrument. The sensitivity to odours is usually reported as “olfactory threshold”, that is the minimum concentration that an average individual is able to detect. Often olfactory thresholds are referred to concentrations of the odorant in water and span several orders of magnitude across different volatile compounds. Excluding very weak odorants, such as hydrocarbons, lower alcohols and long-chain fatty acids, odours can be perceived by humans in solutions containing from some ppm (parts per million or mg/L) to few ppt (parts per trillion or ng/L), spanning six orders of magnitude.
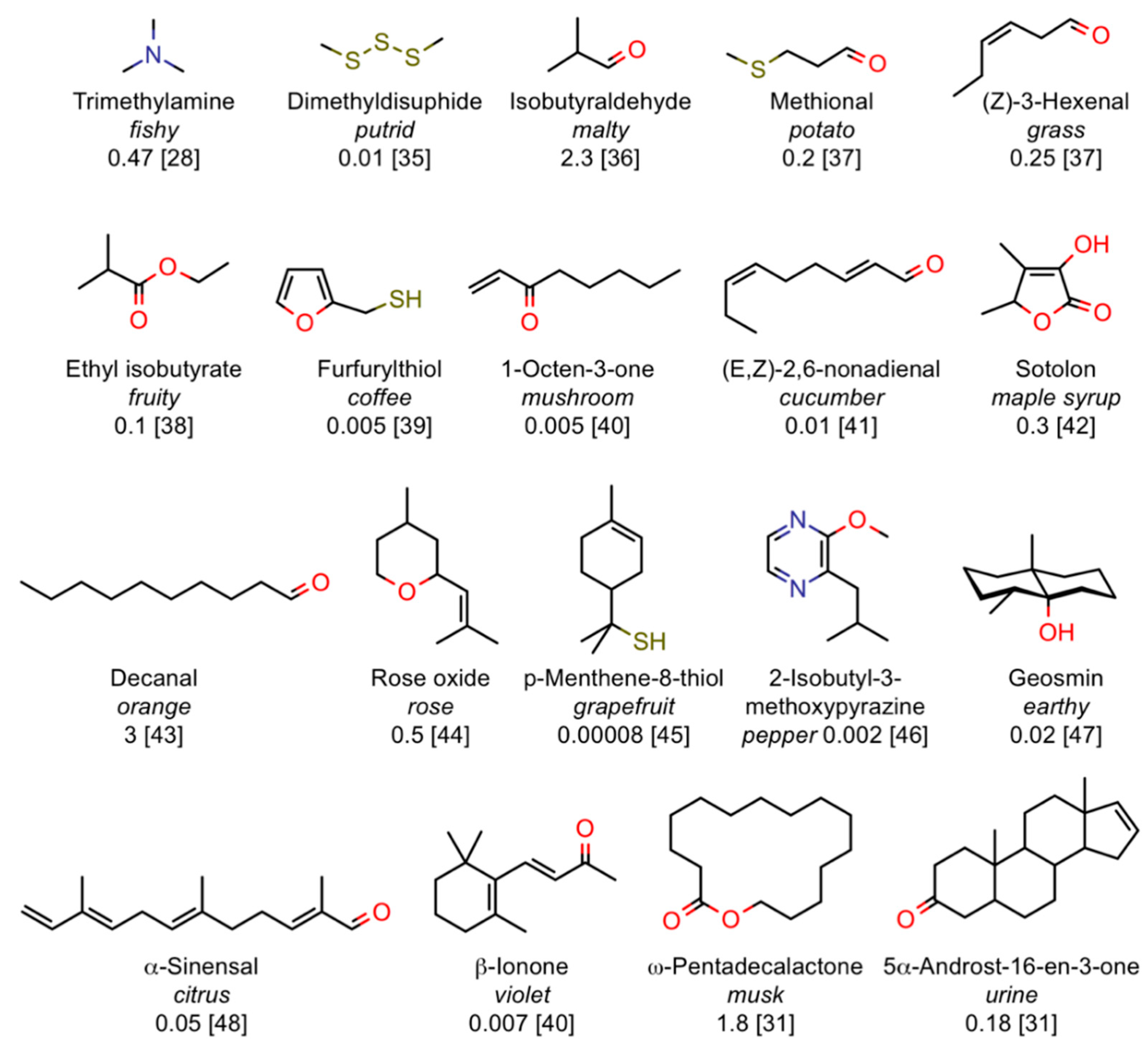
Such a wide range of potencies across different molecular structures suggests that only few volatile compounds are able to fully and efficiently activate their corresponding olfactory receptors, while the majority of odorants are moderate to poor ligands for the available set of ORs and therefore require higher concentrations to switch-on a receptor. This observation may indicate a back door to the breaking of the olfactory code. As a first crude approach, we can take the strongest odorants as those best fitting single ORs and suggest them as candidates for “primary odours”. We cannot exclude, however, that for some types of odours higher thresholds may depend on receptors, which are not finely tuned to one or few molecular structures, but rather respond to a class of chemicals. Figure 3 reports some typical examples of odorants with very low olfactory thresholds [28,31,35,36,37,38,39,40,41,42,43,44,45,46,47,48].
The actual number of molecules reaching the nose is difficult to evaluate, depending on the air/water partition coefficient, the temperature, the air flow and other parameters, but certainly is a very small fraction of those contained in the sniff bottle. However, for the strongest odorants, such as 2-isobutyl-3-methoxypyrazine or geosmin (detected in water solution at concentrations around 10 pM), we can estimate that few picograms are enough to be perceived by the human nose. Incidentally, we can observe that insects are much more sensitive to odours, in particular to pheromones. It has been estimated that about 1000 molecules hitting the antenna of the silkmoth Bombyx mori are enough to elicit a behavioural response to bombykol, the specific sex pheromone [49,50,51]. For most odorants, however, human olfactory thresholds are much higher with detection limits in the order of nanograms. Such values compare well with the performance of most mass spectrometers, which are the most sensitive analytical instruments. However, mass spectrometers are bulky, expensive and complex equipment, and, unlike the olfactory system, need a separation step before performing the analysis.
Therefore, we should better compare the sensitivity of the biological nose with those of electronic instruments currently used as gas detectors. These will be described in the next section, but we can anticipate that their detection limits are orders of magnitude higher than that of the human nose. Unless we improve the sensitivity by at least 3–5 orders of magnitude, we cannot adopt currently available devices for the analysis in real time of odours in the food industry or in the environment. This problem has represented so far a major bottleneck and is going to be one of the main concerns in building an artificial nose. Electronic amplifiers can improve the signals, but up to a certain extent, because the noise, intrinsic to any type of measurement, will be amplified by the same factor.
A new approach therefore is needed, and again we can get inspiration form nature, although at present technical difficulties need also to be overcome. In the biological olfactory system amplification is obtained by the use of an enzymatic cascade through a G-protein and the action of adenylate cyclase [52,53]. This mechanism is equivalent to an electronic amplifier and represent only part of the solution. To amplify the signal and at the same time reduce the noise, in the physiological system the output from several hundreds peripheral olfactory neurons converge to a single glomerulus in the olfactory bulb [16,54]. It would be tempting to connect a large number of identical gas sensors in parallel to achieve a similar result, but the current technology does not enable us to fabricate identical sensors, as the neurons in our nose.
3. An Artificial Nose
To model the olfactory system with an artificial device, we can identify three steps in the process of translating chemical information encoded in volatile molecules into measurable parameters describing odour quality and concentration.
- An array of gas sensors, able to interact with volatile molecules and produce some sort of signal (electrical, optical, etc.) suitable to be amplified and processed.
- An amplifier to cope with the very low concentrations associated with odours.
- A pattern recognition software for recognizing specific response profiles associated with different odours.
In the following sections we shall discuss current technologies applied to the fabrication of gas sensors, focusing on biosensors for odours based on binding proteins. Instead, aspects related to signal amplification and pattern recognition are outside the scope of the present review.
3.1. Detecting the Signals
Chemical sensors to be used in an artificial nose should respond to odours using the same strategy adopted by the human nose. To clarify this concept, we can break it into a number of more specific and focused questions:
- Type of sensing elements. Obviously we can adopt sensors very different in their nature from olfactory receptors (they do not even have to be proteins), as far as they discriminate between different molecules on the same basis adopted by the biological systems; we know that, apart from few exceptions, stereochemical parameters are more important than functional groups, therefore our sensors should discriminate on the basis of size and shape; as an example, it is important that 1-octanol and 3-octanol, which smell floral and mushroom, respectively, should be clearly discriminated, while the latter should be rated as similar to 3-octanone, which also smells mushroom.
- Number of sensing elements. Mammals use several hundreds of different receptors, and humans in particular just over 300; are they really required, when with only three optical sensors we can discriminate millions of colours? The answer lies in the different performances of colour vision and olfaction. While with our nose we can pick-up single specific olfactory notes in a complex odour (just think of how we can detect with our nose the ingredients present in foods by inhaling their volatiles without separating the components), the three colours mix in our brain and we only perceive the final result. Such feature of olfaction is very important when it comes to smell some off-odour in our food among a variety of different good flavours, but is much more important in other animals, whose life depends on a correct functioning of their sense of smell: the gazelle can detect the smell of the lion, and vice versa, among a large number of environmental odours; likewise, an insect can recognise the sex pheromone released by the female of its own species among dozen of similar odours produced by other insects. On the other hand, the auditory system is equipped with thousands of sensors, each tuned to a very narrow part of the spectrum, thus enabling us to understand a conversation against an intense background of different sounds.
- The question of the code. Physiological noses detect and discriminate between a very large number of odorants using a combinatorial approach, which is based on a certain code. Just like the letters of the alphabet, used to form thousands of different words, elementary signals generated by each olfactory receptor combine into a complex odour picture. Now, the question is whether we are bound to use the same code as the natural one when designing an electronic nose. The answer is complex and depends on what sort of an instrument we aim to achieve. For a faithful of reproduction of the human nose, probably we have to use chemical sensors with more or less the selectivity of the natural olfactory receptors. In other words, we can better use the same code, but not necessarily. Our olfactory receptors are rather broadly tuned and cover with their sensitivity most of the spectrum of chemical structures within certain size limits: therefore, we could also adopt sensing elements whose individual response spectra are different from those of the natural receptors, provided that the all set of sensors covers the same area of chemical structures detected by the human nose. To make a simple analogy with colour vision, an instrument for measuring the colour could be made with three sensors not necessarily tuned to the same wavelength of our three rhodopsins, as long as the set of the three sensors covers all the interval from 400 to 800 nm.
We can conclude that we need a large number of sensors, likely of the order of 100 or more, with broad overlapping specificities, and discriminating odorants on the same strategies adopted by the human nose; but we are not bound to adopt the same code. However, a knowledge of the human olfactory code is still essential when choosing the sensing elements. Table 1 summarizes the main characteristics of biological noses and artificial chemosensors mimicking the coding and discrimination of the human olfactory system.
Once agreed upon the fact that we do not need to incorporate our olfactory receptors into artificial sensors for odours, we can discuss the criteria for selecting the most appropriate materials. Once again, the choice depends on the applications of our electronic nose.
3.2. The Question of the Environment
A basic distinction can be made on the environment in which our sensors should operate. Both in natural system and in artificial devices, temperature and humidity are of great importance, particularly when using proteins as sensing elements. Strange enough, to smell air-borne molecules we use a system whose components work in a water environment and would become inactivated if dried. When animals emerged from an aqueous to a terrestrial life, this was probably one of the greatest challenges they had to face. Olfaction is vital to most animal species and the first terrestrial species needed to smell the new environment to survive. Vertebrates keep their olfactory receptors within a moist environment with the help of a mucus made of large glycoproteins [55]. This solution works well in a living organism, where the elements of the system are continuously replaced, but would pose technical difficulties in an artificial device. Moreover, a layer of a hydrogel to keep the right humidity around the proteins of a biosensor, mimicking the biological mucus, could slow down the diffusion of odorant molecules towards the sensing elements, and, even worse, away from them, thus creating serious problems for the regeneration of the device. In nature, apart from a flow of mucus continuously washing the olfactory epithelium, there are active mechanisms of interrupting the signal and regenerating the receptors [56,57].
The choice of the materials for our sensing elements is related to that of the environment. Proteins, such as receptors and soluble binding proteins, would provide the best selectivity for odorants, but require an aqueous medium. On the other hand, solid state sensors, such as those based on semiconducting organic materials do not provide, at the present state-of-the-art, a satisfactory capacity of discriminating between volatiles of similar structure but different odours. In the next section we shall compare the performance of different types of gas sensors and evaluate advantages and problems related to each kind of material.
4. Types and Performance of Gas Sensors
4.1. Metal Oxides
We focus here on metal oxides and conducting polymers as chemical sensors, representing the most frequently used platforms in gas sensing devices. The first materials adopted for gas detection were the so-called MOS (metal oxide sensor) constituted of pellets of metal oxides, such as zinc, tin, nickel, but also other transition metals, whose electrical resistance changes when in contact with organic compounds in the gas phase. More recently, graphene-based materials have been added to this family of gas detectors [58]. These sensors are simple, cheap and robust. However, they present two main problems: (a) they have very limited selectivity and respond with similar intensities to most, although not to all, volatiles; still, a rudimentary version of an “artificial nose” was built with three such sensors [4], although the main value of that first work was to prove the feasibility of detecting and discriminating smells; (b) the other disadvantage is more of technical nature and is their very slow regeneration, as they have to be heated at 300 °C to bring the response back to base line. These sensors are still used as smoke alarms or to detect leaking of cooking gas in homes.
4.2. Conducting Polymers
Organic conducting polymers represented a great step forward to improve selectivity. As in the case of MOS, electrical resistance is the parameter measured in response to odours. Large variations have been observed, depending on the polymer and the odorants. Perhaps the highest sensitivity has been recorded using polypyrrole and ammonia [59,60,61,62]. But the great advantage with these materials is the possibility of changing their spectra of response by modifying their chemical structure. The skeleton can be represented by chains of identical units of pyrrole, thiophene, aniline and other aromatic units, but could also contain more than one basic ring connected in different ways along the chain. Moreover, and most interestingly, the monomer can be chemically modified in a large number of possibilities, each giving rise to a different polymer. For instance, polymers analogous to polypyrrole have been prepared starting from derivatives of pyrrole bearing different groups attached to the nitrogen atoms, form alkyl chains of different lengths, aromatic rings and functional groups, such as a hydroxy, a carbonyl, a carboxylic acid or an amine [63]. These groups are similar to those found in proteins, although the linear structure of conducting polymers does not allow the formation of a binding cavities, as in proteins. Consequently, the selectivity of conducting polymers and other organic materials, although being much better than that of MOS, is still very broad.
In some cases, however, discrimination between homologous compounds of the same chemical class, such as the series of linear alcohols, has been achieved with such materials, but the mechanisms behind are not well understood [61]. Basically, we assume that a ligand with a functional group very likely should establish the strongest interaction between its functional group and the charges located along the backbone of the polymer. In polypyrrole, there is on the average a positive charge every four pyrrole units. Such interaction can be modulated by groups attached to the backbone of the polymer, which could make the accession to the charged atoms easier or more difficult, according to the structures of these groups and those of the odorants. For example, in some cases there could be steric hindrance preventing the odorant molecule from reaching the polymer backbone. In other cases, the groups attached to the polymer main chain might facilitate the approaching of the odorant by establishing weak types of interaction.
At present a large number of semiconducting materials have been synthesised and characterised, and more than 100 are commercially available [62]. However, we are still unable to design a conducting polymer with a sort of binding pocket, as in a protein, exhibiting a certain specificity to molecular structures. This does not mean that a better focused research and perhaps new strategies could lead to semiconducting materials with designed and specific binding pockets for target volatiles.
4.3. Olfactory Receptors
If OBPs can be regarded as the ideal proteins to use in electronic sensing devices, thanks to their high stability combined with easy and inexpensive production and the possibility of modifying their specificity of binding, still they do not bear close relationships to the olfactory system, at least to the human nose. In fact, OBPs of vertebrates are dedicated to pheromone detection and linked to the vomeronasal organ, a function and an organ that have disappeared in higher primates [64]. OBPs of insects, on the other hand, although many of them are tuned to plant volatiles, chemicals that we can also smell as humans, do not mirror the specificities of the human olfactory receptors.
Consequently, the ideal choice for mimicking the human nose, falls on olfactory receptors [65,66,67,68,69,70]. This choice, however, carries serious difficulties that at present state of the technology cannot be addressed. As most kinds of receptors, those of our nose are transmembrane proteins, therefore need a membrane to sit across, and are very delicate as far as their three-dimensional structure is concerned. Artificial membranes can be fabricated and are rather stable in a laboratory environment but represent a weak element when part of a commercial instrument. The expression and reconstitution of olfactory receptors is far from easy, although few of them have been successfully expressed with very low yields, in heterologous systems.
There is another problem of different nature when dealing with membrane-bound receptors. Once the odorant has entered the binding site and a signal has been produced, it is almost impossible to regenerate the receptor. The strategy that is often followed is that of having what seems like disposable sensors: the entire complex is engulfed inside the cell and its components recycled. A company claims to provide an array with all the human olfactory receptors to be used as an artificial nose. This array is disposable and can be used for a single measurement, leaving open all the questions on the reliability of such measurements.
4.4. Soluble Binding Proteins
Proteins would represent the ideal sensing elements for an electronic nose, providing the required selectivity and offering the possibility of borrowing the same proteins actually used in the olfactory system. However, as already discussed, proteins require an aqueous environment, a situation difficult to reproduce in an electronic device aimed at detecting air-borne molecules. Nevertheless, they can be adopted for sensing devices in solution, instruments that we could more appropriately call electronic tongues rather than electronic noses.
Thanks to their simple structure and high stability, odorant-binding proteins (OBPs) represent good candidates to be incorporated into sensing devices and have been successfully used to detect odorants. OBP is the common name given to two distinct families of proteins of vertebrates and insects endowed with the role of carrying hydrophobic odorants across an aqueous environment, the vertebrate nasal mucus and the insect sensillar lymph, to the membrane-bound olfactory receptors. Vertebrate OBPs contain 150–180 amino acids folded in a compact structure known as β-barrel and constituted of eight β-strands and a small α-helix [71,72,73]. Those of insects are smaller (120–140 amino acids) and folded into six α-helices domains [74,75]. Proteins of both classes present a binding cavity where molecules of odorants and pheromones can be accommodated with relatively good affinity (dissociation constants in the micromolar order). Their selectivity to hydrophobic ligands is not as high as known for olfactory receptors, but fine discrimination has been reported in several cases, such as the two enantiomers of carvone bound with different affinities by the pig OBP [76] or the panda OBP5 able to distinguish between stearic, oleic and linoleic acid, as well as isomers of oleic acid differing for the position of the double bond [77].
OBPs present several advantages if used in biosensing elements for odours. First, they can be produced in bacteria in relatively large quantities (20–40 mg/L) and easily purified by simple column chromatography. More interestingly, their compact structure grants them an exceptional stability to temperature, withstanding boiling for several minutes [78,79,80,81], to proteolytic digestion [82] and organic solvents. This characteristic is certainly useful in proteins being exposed to harsh environmental conditions.
The structure of OBPs can also be modified with simple protocols of site-directed mutagenesis to change their affinity and selectivity to ligands and generate tailor made sensing elements for specific requirements. For example, replacing phenylalanine 88 in the pig OBP with a tryptophan obtained the effect of inserting a fluorescent probe in the binding cavity to monitor the entrance of ligands. At the same time, such replacement increased the affinity of the protein to polycyclic hydrocarbons [83], but also unexpectedly endowed the pig OBP with the capacity of discriminating between the two enantiomers of carvone [76]. Another example is given by one of the OBPs of the moth Helicoverpa armigera, GOBP2, which is tuned to the species-specific sex pheromone (Z)-11-hexadecenal and structurally related aldehydes. Replacing two or three amino acid residues in the binding pocket of the protein was enough to drastically reduce affinities to long-chain aldehydes and shift the specificity of GOBP2 to terpenoids [84]. In a similar work, a single amino acid mutation in the OBP3 of the giant panda, which binds long-chain aldehydes and several plant-derived terpenoids, narrowed the specificity of this protein to the second class of compounds [77]. Based on the wide and detailed structural information on OBPs and on the role of single amino acid residues, as learned from studies of site–directed mutagenesis, we can easily conceive the possibility of designing completely new proteins tuned to desired ligands.
4.5. Transducing Strategies
When using metal oxides or semiconducting polymers as gas sensing elements, a signal is directly produced as a change in electric resistance upon interactions with odours. But a protein, such as a receptor or a soluble binding protein, only undergoes a conformational change, that has to be converted into a signal that can be measured, amplified and processed. There are a number of alternatives to perform such step, basically falling into three categories, where changes of mass, optical signals or electric properties are measured.
Using piezoelectric quartz crystals, a change of mass can be accurately measured by monitoring its oscillation frequency. The method, called surface acoustic wave (SAW), involves immobilization of the protein the protein on the crystal and monitoring the change of frequency occurring when a ligand binds to the protein, thus increasing the mass of the sensor [85,86]. The method has been applied to odour sensing, using the bovine and the pig OBPs to monitor the presence of common volatile compounds [87]. A problem related to such technique when utilised to detect chemicals in the gas phase is caused by changes in the environmental humidity, that can be difficult to control. In some cases, a reference sensor can reduce the inaccuracy involved [87].
Optical sensors can record changes in refractive index consequent to binding of a ligand to a protein or a DNA fragment. The technique, called surface plasmon resonance (SPR) was adopted in the Biacore commercial instruments. This approach has been used to monitor binding of odorants to olfactory receptors [88,89] immobilised on a prism, with detection limits in the micromolar range. The method, however, requires the use of bulky and expensive instruments. Moreover, it provides reliable results when the ligand is a large molecule, such as a protein or DNA fragment, but is not very accurate in the case of a small organic compounds, as an odorant. A simplified system has been described, with an OBP immobilised on an optical fibre, that could lead to the fabrication of portable biosensors [90].
Electronic sensors modified with OBPs have been used in voltage, current and impedance detectors. In some recently assembled devices, field effect transistors have been modified by attaching OBPs to the gate electrode and measuring the variations in current produced in the presence of various ligands. A study of this type, using the OBP14 of the honey bee linked to reduced graphene oxide, successfully detected eugenol, the best volatile found for the same protein in solution fluorescent assays, but was insensitive to methyleugenol, thus exhibiting a finely tuned specificity [91,92]. High selectivity was also reported for the pigOBP1 immobilised on the gold gate electrode of a transistor, that was able to discriminate between the two enantiomers of carvone, again producing responses in agreement with data obtained by fluorescent binding assays [76].
Several recent papers describe the fabrication of biosensors using interdigitated electrodes coated with OBPs embedded in nitrocellulose. Such devices respond to the presence of ligands with changes in impedance [93,94,95,96].
Several terpenoids and honey bee pheromones were detected with a biosensor using Apis mellifera OBP2 [88], while the OBP2 of Bactrocera dorsalis immobilised in a similar type of device was found to respond isoamyl acetate, β-ionone and benzaldehyde [95]. The honey bee CSP3 was also used in a parallel study and reported to detect isoamyl acetate, geraniol and phenylacetaldehyde [93].
At present, there are many reports in the literature describing biosensors where an olfactory soluble protein is used as a detector element, but each one is focused on a different protein and different ligands, making a comparison of the published data almost impossible. Moreover, studies comparing the performance of a protein when immobilised in a biosensor with that of the same protein in solution are still lacking. Systematic investigations need to be performed on large numbers of ligands for each protein and dissociation constants calculated from the biosensor responses should be compared with values measured in classical ligand-binding assays.
5. Conclusions
After reviewing the information available on the physiological olfactory systems and the state-of-the-art on gas sensor technologies, we can return to our first question and ask what we still need to design an instrument capable of discriminating odours with performance similar to that of the human nose. The conclusion is that we are still far from building an artificial nose, although this represents a feasible goal in the future.
The main difficulties still come from our poor knowledge of the olfactory code, having deciphered only 50 letters of about 300 that make our smell alphabet. Such basic set of information is necessary even if it might seem as if we do not necessarily need to always use all our biological olfactory receptors as sensing elements. After having identified the best ligands for each of our 300 olfactory receptors, we can certainly reduce the total number of sensors needed, based on the apparent redundancy already observed within the 50 deorphanized ORs. It is likely that a first rudimentary version of our electronic nose could be based on less than 100 different sensors. However, it is worth noticing that redundancy, that makes the biological system more robust and reliable, also improves the performance of electronic noses [97].
Concerning the detecting elements, only proteins have so far shown a selectivity comparable to that of our ORs. However, proteins present some technical disadvantages, due to their poor stability and easy degradation in common environmental conditions. Besides, they need an aqueous environment to keep their functionality, and therefore an appropriate solution for providing an air-water interface. As proteins, OBPs present several advantages over ORs for their better stability and easy of synthesis, but we cannot exclude that our knowledge on the structure and binding modes of OBPs might enable us to design and synthesize completely artificial proteins with improved properties with respect to the biological ones.
Finally, sensitivity still represents a major issue. Currently available gas sensors work at concentrations of ligands several orders of magnitude higher than those capable of producing an odour in our nose. Classical amplifiers, on the other hand, have limits due to the intrinsic noise of electronic circuits. A novel approach to solve this problem is needed, but so far it is difficult to formulate any suggestion on the basis of available technologies.
Author Contributions
P.P. prepared a first draft of the manuscript, to which J.Z. and W.K. added substantial contributions. The final version was revised and approved by all authors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pelosi, P.; Pisanelli, A.M.; Baldaccini, N.E.; Gagliardo, A. Binding of [3H]-2-isobutyl-3-methoxypyrazine to cow olfactory mucosa. Chem. Senses 1981, 6, 77–85. [Google Scholar] [CrossRef]
- Pelosi, P.; Baldaccini, N.E.; Pisanelli, A.M. Identification of a specific olfactory receptor for 2-isobutyl-3-methoxypyrazine. Biochem. J. 1982, 201, 245–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, R.G.; Riddiford, L.M. Pheromone binding and inactivation by moth antennae. Nature 1981, 293, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Persaud, K.; Dodd, G. Analysis of discrimination mechanisms in the mammalian olfactory system using a model nose. Nature 1982, 299, 352–355. [Google Scholar] [CrossRef] [PubMed]
- Persaud, K.C.; Marco, S.; Gutiérrez-Gálvez, A. Neuromorphic Olfaction; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2013. [Google Scholar]
- Stitzel, S.E.; Aernecke, M.J.; Walt, D.R. Artificial noses. Annu. Rev. Biomed. Eng. 2011, 13, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Wasilewski, T.; Gębicki, J.; Kamysz, W. Bioelectronic nose: Current status and perspectives. Biosens. Bioelectron. 2017, 87, 480–494. [Google Scholar] [CrossRef] [PubMed]
- Capelli, L.; Sironi, S.; Del Rosso, R. Electronic noses for environmental monitoring applications. Sensors 2014, 14, 19979–20007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bikov, A.; Lázár, Z.; Horvath, I. Established methodological issues in electronic nose research: How far are we from using these instruments in clinical settings of breath analysis? J. Breath Res. 2015, 9. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zhang, H.; Chen, Q.; Mei, C.; Liu, G. Recent advances in electronic nose techniques for monitoring of fermentation process. World J. Microbiol. Biotechnol. 2015, 31, 1845–1852. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Méndez, M.L.; De Saja, J.A.; González-Antón, R.; García-Hernández, C.; Medina-Plaza, C.; García-Cabezón, C.; Martín-Pedrosa, F. Electronic Noses and Tongues in Wine Industry. Front. Bioeng. Biotechnol. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Wojnowski, W.; Majchrzak, T.; Dymerski, T.; Gębicki, J.; Namieśnik, J. Electronic noses: Powerful tools in meat quality assessment. Meat Sci. 2017, 131, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Majchrzak, T.; Wojnowski, W.; Dymerski, T.; Gębicki, J.; Namieśnik, J. Electronic noses in classification and quality control of edible oils: A review. Food Chem. 2018, 246, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Rossiter, K.J. Structure-Odor Relationships. Chem. Rev. 1996, 96, 3201–3240. [Google Scholar] [CrossRef] [PubMed]
- Sell, C.S. On the unpredictability of odor. Angew. Chem. Int. Ed. Engl. 2006, 45, 6254–6261. [Google Scholar] [CrossRef] [PubMed]
- Mombaerts, P. Molecular biology of odorant receptors in vertebrates. Annu. Rev. Neurosci. 1999, 22, 487–509. [Google Scholar] [CrossRef] [PubMed]
- Mombaerts, P. Odorant receptor genes in humans. Curr. Opin. Genet. Dev. 1999, 9, 315–320. [Google Scholar] [CrossRef]
- Mombaerts, P. The human repertoire of odorant receptor genes and pseudogenes. Annu. Rev. Genom. Hum. Genet. 2001, 2, 493–510. [Google Scholar] [CrossRef] [PubMed]
- Crasto, C.; Singer, M.S.; Shepherd, G.M. The olfactory receptor family album. Genome Biol. 2001, 2. [Google Scholar] [CrossRef]
- Olender, T.; Nativ, N.; Lancet, D. HORDE: Comprehensive resource for olfactory receptor genomics. Methods Mol. Biol. 2013, 1003, 23–38. [Google Scholar] [PubMed]
- Malnic, B.; Hirono, J.; Sato, T.; Buck, L.B. Combinatorial receptor codes for odors. Cell 1999, 96, 713–723. [Google Scholar] [CrossRef]
- Korsching, S. Olfactory maps and odor images. Curr. Opin. Neurobiol. 2002, 12, 387–392. [Google Scholar] [CrossRef]
- Imai, T.; Sakano, H.; Vosshall, L.B. Topographic mapping—The olfactory system. Cold Spring Harb. Perspect. Biol. 2010, 2. [Google Scholar] [CrossRef] [PubMed]
- Murthy, V.N. Olfactory maps in the brain. Annu. Rev. Neurosci. 2011, 34, 233–258. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Sakano, H. How is the olfactory map formed and interpreted in the mammalian brain? Annu. Rev. Neurosci. 2011, 34, 467–499. [Google Scholar] [CrossRef] [PubMed]
- Amoore, J.E. Specific anosmia: A clue to the olfactory code. Nature 1967, 214, 1095–1098. [Google Scholar] [CrossRef] [PubMed]
- Amoore, J.E. A Plan to Identify Most of the Primary Odors. In Olfaction and Taste III; Pfaffmann, C., Ed.; Rockefeller University Press: New York, NY, USA, 1969; pp. 158–171. [Google Scholar]
- Amoore, J.E. Specific anosmia and the concept of primary odors. Chem. Senses 1977, 2, 267–281. [Google Scholar] [CrossRef]
- Guillot, M. Anosmies Partielles et Odeurs Fondamentales. CRH Acad. Sci. 1948, 226, 1307–1309. [Google Scholar]
- Le Magnen, J. Les Phenomenes Olfacto-sexuels chez l’Homme. Arch. Sci. Physiol. 1952, 6, 125–160. [Google Scholar]
- Amoore, J.E.; Pelosi, P.; Forrester, L.J. Specific anosmia to 5α-androst-16-en-3-one and ω pentadecalactone: The urinous and musky primary odors. Chem. Senses 1977, 2, 401–425. [Google Scholar] [CrossRef]
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- Olender, T.; Keydar, I.; Pinto, J.M.; Tatarskyy, P.; Alkelai, A.; Chien, M.S.; Fishilevich, S.; Restrepo, D.; Matsunami, H.; Gilad, Y.; et al. The human olfactory transcriptome. BMC Genom. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Verbeurgt, C.; Wilkin, F.; Tarabichi, M.; Gregoire, F.; Dumont, J.E.; Chatelain, P. Profiling of olfactory receptor gene expression in whole human olfactory mucosa. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Buttery, R.G.; Guadagni, D.G.; Ling, L.C.; Seifert, R.M.; Lipton, W. Additional volatile components of cabbage, broccoli, and cauliflower. J. Agric. Food Chem. 1976, 24, 829–832. [Google Scholar] [CrossRef]
- Amoore, J.E.; Firrester, L.J.; Pelosi, P. Specific anosmia to isobutyladehyde: The malty primary odor. Chem. Senses 1976, 2, 17–25. [Google Scholar]
- Buttery, R.G.; Teranishi, R.; Ling, L.C.; Turnbaugh, J.G. Quantitative and sensory studies on tomato paste volatiles. J. Agric. Food Chem. 1990, 38, 336–340. [Google Scholar] [CrossRef]
- Buttery, R.G.; Seifert, R.M.; Ling, L.C.; Soderstrom, E.L.; Ogawa, J.M.; Turnbaugh, J.G. Additional aroma components of honeydew melon. J. Agric. Food Chem. 1982, 30, 1208–1211. [Google Scholar] [CrossRef]
- Tressl, R. Formation of Flavor Components in Roasted Coffee. In Thermal Generation of Aromas; Parliment, T.H., McGorrin, R.J., Ho, C.-T., Eds.; American Chemical Society (ACS): Washington, DC, USA, 1989; pp. 285–301. [Google Scholar]
- Buttery, R.G.; Teranishi, R.; Flath, R.A.; Ling, L.C. Identification of additional tomato paste volatiles. J. Agric. Food Chem. 1990, 38, 792–795. [Google Scholar] [CrossRef]
- Teranishi, R.; Buttery, R.G.; Gaudagni, D.G. Odor Quality and Chemical Structure in Fruit and Vegetable Flavors. Ann. N. Y. Acad. Sci. 1974, 237, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Blank, I.; Lin, J.; Fumeaux, R.; Welti, D.H.; Fay, L.B. Formation of 3-hydroxy-4,5-dimethyl-2(5H)-furanone (sotolone) from 4-hydroxy-L-isoleucine and 3-amino-4,5-dimethyl-3,4-dihydro-2(5H)-furanone. J. Agric. Food Chem. 1996, 44, 1851–1856. [Google Scholar] [CrossRef]
- Ahmed, E.M.; Dennison, R.A.; Dougherty, R.H.; Shaw, P.E. Flavor and odor thresholds in water of selected orange juice components. J. Agric. Food Chem. 1978, 26, 187–191. [Google Scholar] [CrossRef]
- Plotto, A.; Margaria, C.A.; Goodner, K.L.; Goodrich, R.; Baldwin, E.A. Odour and flavor thresholds for key aroma components in an orange juice matrix: Ester and miscellaneous compounds. Flavour Fragr. J. 2008, 23, 398–406. [Google Scholar] [CrossRef]
- Demole, E.; Enggist, P.; Ohloff, G. p-Menthene-8-thiol: A powerful flavor impact constituent of grapefruit juice. Helv. Chim. Acta 1982, 65, 1785–1794. [Google Scholar] [CrossRef]
- Buttery, R.G.; Seifert, R.M.; Guadagni, D.G.; Ling, L.C. Characterization of some volatile constituents of bell peppers. J. Agric. Food Chem. 1969, 17, 1322–1327. [Google Scholar] [CrossRef] [PubMed]
- Buttery, R.G.; Guadagni, D.G.; Ling, L.C. Geosmin, a musty off-flavor of dry beans. J. Agric. Food Chem. 1976, 24, 419–420. [Google Scholar] [CrossRef]
- Ohloff, G. Recent Developments in the Field of Naturally Occurring Aroma Components. Prog. Chem. Org. Nat. Prod. 1978, 35, 431–527. [Google Scholar]
- Kaissling, K.E.; Priesner, E. Die Riechschwelle des Seidenspinners. Naturwissenschaften 1970, 57, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Kaissling, K.E. Chemo-electrical transduction in insect olfactory receptors. Annu. Rev. Neurosci. 1986, 9, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Kaissling, K.E. Pheromone Reception in Insects: The Example of Silk Moths. In Neurobiology of Chemical Communication; Mucignat-Caretta, C., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2014; Chapter 4. [Google Scholar]
- Firestein, S. How the olfactory system makes sense of scents. Nature 2001, 413, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Ronnett, G.V.; Moon, C. G proteins and olfactory signal transduction. Annu. Rev. Physiol. 2002, 64, 189–222. [Google Scholar] [CrossRef] [PubMed]
- Maresh, A.; Gil, D.R.; Whitman, M.C.; Greer, C.A. Principles of glomerular organization in the human olfactory bulb-implications for odor processing. PLoS ONE 2008, 3. [Google Scholar] [CrossRef] [PubMed]
- Gladysheva, O.; Kukushkina, D.; Martynova, G. Glycoprotein composition of olfactory mucus in vertebrates. Acta Histochem. 1986, 78, 141–146. [Google Scholar] [CrossRef]
- Schleicher, S.; Boekhoff, I.; Arriza, J.; Lefkowitz, R.J.; Breer, H. A beta-adrenergic receptor kinase-like enzyme is involved in olfactory signal termination. Proc. Natl. Acad. Sci. USA 1993, 90, 1420–1424. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Zhao, A.Z.; Chan, G.C.; Baker, L.P.; Impey, S.; Beavo, J.A.; Storm, D.R. Phosphorylation and inhibition of olfactory adenylyl cyclase by CaM kinase II in Neurons: A mechanism for attenuation of olfactory signals. Neuron 1998, 21, 495–504. [Google Scholar] [CrossRef]
- Joshi, N.; Hayasaka, T.; Liu, Y.; Liu, H.; Oliveira, O.N., Jr.; Lin, L. A review on chemiresistive room temperature gas sensors based on metal oxide nanostructures, graphene and 2D transition metal dichalcogenides. Mikrochim. Acta 2018, 185. [Google Scholar] [CrossRef] [PubMed]
- Persaud, K.C.; Pelosi, P. An approach to an artifiacial nose. Trans. Am. Soc. Artif. Intern. Organs 1985, 31, 297–300. [Google Scholar] [PubMed]
- Amrani, M.E.H.; Persaud, K.C.; Payne, P.A. High-frequency measurements of conducting polymers: Development of a new technique for sensing volatile chemicals. Meas. Sci. Technol. 1995, 6, 1500–1507. [Google Scholar] [CrossRef]
- Hatfield, J.V.; Neaves, P.; Hicks, P.J.; Persaud, K.; Travers, P. Towards an integrated electronic nose using conducting polymer sensors. Sens. Actuators B Chem. 1994, 18, 221–228. [Google Scholar] [CrossRef]
- Lange, U.; Roznyatovskaya, N.V.; Mirsky, V.M. Conducting polymers in chemical sensors and arrays. Anal. Chim. Acta 2008, 614, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Persaud, K.C. Gas Sensors: Towards an artificial nose. In Sensors and Sensory Systems for Advanced Robots; Dario, P., Ed.; Springer: Berlin, Germany, 1988; pp. 361–382. [Google Scholar]
- Pelosi, P. The role of perireceptor events in vertebrate olfaction. Cell. Mol. Life Sci. 2001, 58, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Missbach, C.; Dweck, H.K.; Vogel, H.; Vilcinskas, A.; Stensmyr, M.C.; Hansson, B.S.; Grosse-Wilde, E. Evolution of insect olfactory receptors. eLife 2014, 26. [Google Scholar] [CrossRef] [PubMed]
- Buck, L.B. Olfactory receptors and odor coding in mammals. Nutr. Rev. 2004, 62, 184–188. [Google Scholar] [CrossRef]
- Touhara, K.; Vosshall, L.B. Sensing odorants and pheromones with chemosensory receptors. Annu. Rev. Physiol. 2009, 71, 307–332. [Google Scholar] [CrossRef] [PubMed]
- Wasilewskia, T.; Gebickib, J.; Kamysza, W. Advances in olfaction-inspired biomaterials applied to bioelectronic noses. Sens. Actuator B Chem. 2018, 257, 511–537. [Google Scholar] [CrossRef]
- Breer, H. Olfactory receptors: Molecular basis for recognition and discrimination of odors. Anal. Bioanal. Chem. 2003, 377, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Latek, D.; Modzelewska, A.; Trzaskowski, B.; Palczewski, K.; Filipek, S. G protein-coupled receptors-recent advances. Acta Biochim. Pol. 2012, 59, 515–529. [Google Scholar] [PubMed]
- Bianchet, M.A.; Bains, G.; Pelosi, P.; Pevsner, J.; Snyder, S.H.; Monaco, H.L.; Amzel, L.M. The three dimensional structure of bovine odorant-binding protein and its mechanism of odor recognition. Nat. Struct. Biol. 1996, 3, 934–939. [Google Scholar] [CrossRef] [PubMed]
- Tegoni, M.; Ramoni, R.; Bignetti, E.; Spinelli, S.; Cambillau, C. Domain swapping creates a third putative combining site in bovine odorant binding protein dimer. Nat. Struct. Biol. 1996, 3, 863–867. [Google Scholar] [CrossRef] [PubMed]
- Tegoni, M.; Pelosi, P.; Vincent, F.; Spinelli, S.; Campanacci, V.; Grolli, S.; Ramoni, R.; Cambillau, C. Mammalian odorant binding proteins. Biochim. Biophys. Acta 2000, 1482, 229–240. [Google Scholar] [CrossRef]
- Sandler, B.H.; Nikonova, L.; Leal, W.S.; Clardy, J. Sexual attraction in the silkworm moth: Structure of the pheromone-binding-protein-bombykol complex. Chem. Biol. 2000, 7, 143–151. [Google Scholar] [CrossRef]
- Tegoni, M.; Campanacci, V.; Cambillau, C. Structural aspects of sexual attraction and chemical communication in insects. Trends Biochem. Sci. 2004, 29, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Mulla, M.Y.; Tuccori, E.; Magliulo, M.; Lattanzi, G.; Palazzo, G.; Persaud, K.; Torsi, L. Capacitance-modulated transistor detects odorant binding protein chiral interactions. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Arena, S.; Spinelli, S.; Liu, D.; Zhang, G.; Wei, R.; Cambillau, C.; Scaloni, A.; Wang, G.; Pelosi, P. Reverse chemical ecology: Olfactory proteins from the giant panda and their interactions with putative pheromones and bamboo volatiles. Proc. Natl. Acad. Sci. USA 2017, 114, 9802–9810. [Google Scholar] [CrossRef] [PubMed]
- Paolini, S.; Tanfani, F.; Fini, C.; Bertoli, E.; Pelosi, P. Porcine odorant-binding protein: Structural stability and ligand affinities measured by fourier-transform infrared spectroscopy and fluorescence spectroscopy. Biochim. Biophys. Acta 1999, 1431, 179–188. [Google Scholar] [CrossRef]
- Schwaighofer, A.; Pechlaner, M.; Oostenbrink, C.; Kotlowski, C.; Araman, C.; Mastrogiacomo, R.; Pelosi, P.; Knoll, W.; Nowak, C.; Larisika, M. Insights into structural features determining odorant affinities to honey bee odorant binding protein 14. Biochem. Biophys. Res. Commun. 2014, 446, 1042–1046. [Google Scholar] [CrossRef] [PubMed]
- Schwaighofer, A.; Kotlowski, C.; Araman, C.; Chu, N.; Mastrogiacomo, R.; Becker, C.; Pelosi, P.; Knoll, W.; Larisika, M.; Nowak, C. Honey bee odorant-binding protein 14: Effects on thermal stability upon odorant binding revealed by FT-IR spectroscopy and CD measurements. Eur. Biophys. J. 2014, 43, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Mastrogiacomo, R.; Iovinella, I.; Tuccori, E.; Persaud, K.C. Structure and biotechnological applications of odorant-binding proteins. Appl. Microbiol. Biotechnol. 2014, 98, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Paolini, S.; Scaloni, A.; Amoresano, A.; Marchese, S.; Napolitano, E.; Pelosi, P. Amino acid sequence, post-translational modifications, binding and labelling of porcine odorant-binding protein. Chem. Senses 1998, 23, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Brandazza, A.; Pelosi, P. Binding of Polyclycic Aromatic Hydrocarbons to Mutants of Odorant-Binding Protein: A first step towards biosensors for environmental monitoring. Biochim. Biophys. Acta 2008, 1784, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Ban, L.; Song, L.M.; Liu, Y.; Pelosi, P.; Wang, G. General odorant-binding proteins and sex pheromone guide larvae of Plutella xylostella to better food. Insect Biochem. Mol. Biol. 2016, 72, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Hotel, O.; Poli, J.P.; Mer-Calfati, C.; Scorsone, E.; Saada, S. SAW Sensor’s Frequency Shift Characterization for Odor Recognition and Concentration Estimation. IEEE Sens. J. 2017, 17, 7011–7018. [Google Scholar] [CrossRef]
- Hoß, S.G.; Bendas, G. Mass-Sensitive Biosensor Systems to Determine the Membrane Interaction of Analytes. Methods Mol. Biol. 2017, 1520, 145–157. [Google Scholar] [PubMed]
- Di Pietrantonio, F.; Cannatà, D.; Benetti, M.; Verona, E.; Varriale, A.; Staiano, M.; D’Auria, S. Detection of odorant molecules via surface acoustic wave biosensor array based on odorant-binding proteins. Biosens. Bioelectron. 2013, 41, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Benilova, I.; Chegel, V.I.; Ushenin, Y.V.; Vidic, J.; Soldatkin, A.P.; Martelet, C.; Pajot, E.; Jaffrezic-Renault, N. Stimulation of human olfactory receptor 17–40 with odorants probed by surface plasmon resonance. Eur. Biophys. J. 2008, 37, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Sanmartí-Espinal, M.; Iavicoli, P.; Calò, A.; Taulés, M.; Galve, R.; Marco, M.P.; Samitier, J. Quantification of interacting cognate odorants with olfactory receptors in nanovesicles. Sci. Rep. 2017, 7, 17483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cennamo, N.; Di Giovanni, S.; Varriale, A.; Staiano, M.; Di Pietrantonio, F.; Notargiacomo, A.; Zeni, L.; D’Auria, S. Easy to use plastic optical fiber-based biosensor for detection of butanal. PLoS ONE 2015, 10, e0116770. [Google Scholar] [CrossRef] [PubMed]
- Larisika, M.; Kotlowski, C.; Steininger, C.; Mastrogiacomo, R.; Pelosi, P.; Schütz, S.; Peteu, S.F.; Kleber, C.; Reiner-Rozman, C.; Nowak, C.; et al. Electronic Olfactory Sensor Based on A. mellifera Odorant-Binding Protein 14 on a Reduced Graphene Oxide Field—Effect Transistor. Angew. Chem. Int. Ed. Engl. 2015, 54, 13245–13248. [Google Scholar] [CrossRef] [PubMed]
- Kotlowski, C.; Larisika, M.; Guerin, P.M.; Kleber, C.; Kröber, T.; Mastrogiacomo, R.; Nowak, C.; Schutz, S.; Schwaighofer, A.; Knoll, W. Fine discrimination of volatile compounds by graphene-immobilized odorant-binding proteins. Sens. Actuators B Chem. 2018, 256, 564–572. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, H.; Li, H.; Zhang, J.; Zhuang, S.; Zhang, F.; Hsia, K.J.; Wang, P. Impedance sensing and molecular modeling of an olfactory biosensor based on chemosensory proteins of honeybee. Biosens. Bioelectron. 2013, 40, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Li, H.; Zhuang, S.; Zhang, D.; Zhang, Q.; Zhou, J.; Dong, S.; Liu, Q.; Wang, P. Olfactory biosensor using odorant-binding proteins from honeybee: Ligands of floral odors and pheromones detection by electrochemical impedance. Sens. Actuators B Chem. 2014, 193, 420–427. [Google Scholar] [CrossRef]
- Lu, Y.; Yao, Y.; Zhang, Q.; Zhang, D.; Zhuang, S.; Li, H.; Liu, Q. Olfactory biosensor for insect semiochemicals analysis by impedance sensing of odorant-binding proteins on interdigitated electrodes. Biosens. Bioelectron. 2015, 67, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhang, D.; Zhang, Q.; Huang, Y.; Luo, S.; Yao, Y.; Li, S.; Liu, Q. Impedance spectroscopy analysis of human odorant binding proteins immobilized on nanopore arrays for biochemical detection. Biosens. Bioelectron. 2016, 79, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, E.; Magna, G.; Polese, D.; Vergara, A.; Schild, D.; Di Natale, C. Stable Odor Recognition by a neuro-adaptive Electronic Nose. Sci. Rep. 2015, 5, 10960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Complexity of odour coding. Smell can be only analysed in a multidimensional space, where different molecular parameters contribute to the perceived odour. The examples reported in the figure illustrate the effect of functional groups (nature and position), as well as stereochemical parameters on odour quality. Cases have been reported of enantiomer pairs smelling different, although this is not common in human olfaction.
Figure 1.
Complexity of odour coding. Smell can be only analysed in a multidimensional space, where different molecular parameters contribute to the perceived odour. The examples reported in the figure illustrate the effect of functional groups (nature and position), as well as stereochemical parameters on odour quality. Cases have been reported of enantiomer pairs smelling different, although this is not common in human olfaction.

Figure 2.
Phylogenetic tree of the 50 human receptors so far deorphanised. Each receptor is associated with the structure of its best ligand. In several cases, two or more receptors are tuned to the same odour (indicated with same colour). Such redundancy likely insures that important odours can be detected in case some receptors become faulty of silenced. On the other hand, this observation allows us to predict that the total number of basic odours representing the olfactory code (and therefore the number of sensors required for an artificial nose) can be significantly lower than the 300–350 functional genes encoding for olfactory receptors in the human genome.
Figure 2.
Phylogenetic tree of the 50 human receptors so far deorphanised. Each receptor is associated with the structure of its best ligand. In several cases, two or more receptors are tuned to the same odour (indicated with same colour). Such redundancy likely insures that important odours can be detected in case some receptors become faulty of silenced. On the other hand, this observation allows us to predict that the total number of basic odours representing the olfactory code (and therefore the number of sensors required for an artificial nose) can be significantly lower than the 300–350 functional genes encoding for olfactory receptors in the human genome.

Figure 3.
Examples of volatile compounds with extremely low olfactory thresholds for humans. Together with names and structures, odour description is reported in italics, olfactory thresholds in ppb (parts per billion) and reference in parenthesis. The olfactory threshold values are given as an indication as differences by one order of magnitude are often found between published works. Moreover, sensitivity to each specific odorant can span two orders of magnitude across the human population.
Figure 3.
Examples of volatile compounds with extremely low olfactory thresholds for humans. Together with names and structures, odour description is reported in italics, olfactory thresholds in ppb (parts per billion) and reference in parenthesis. The olfactory threshold values are given as an indication as differences by one order of magnitude are often found between published works. Moreover, sensitivity to each specific odorant can span two orders of magnitude across the human population.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Features of biological and artificial noses.
| Feature | Biological Noses | Artificial Noses |
|---|---|---|
| Type of sensing elements | Membrane receptors of broad overlapping specificity | Macromolecules with a moderate degree of specificity (OBPs, other binding proteins, DNA) |
| Number of sensing elements | About 300 in humans, up to 1000 in other mammals, around 100 in insects | At least 100 types to mimic the discrimination and the wide range of odours detected by the human nose |
| Coding strategy | Discrimination mainly on the basis of stereochemical parameters, but in some cases also of functional groups | Stereochemical parameters and functional groups |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pelosi, P.; Zhu, J.; Knoll, W. From Gas Sensors to Biomimetic Artificial Noses. Chemosensors 2018, 6, 32. https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors6030032
AMA Style
Pelosi P, Zhu J, Knoll W. From Gas Sensors to Biomimetic Artificial Noses. Chemosensors. 2018; 6(3):32. https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors6030032
Chicago/Turabian StylePelosi, Paolo, Jiao Zhu, and Wolfgang Knoll. 2018. "From Gas Sensors to Biomimetic Artificial Noses" Chemosensors 6, no. 3: 32. https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors6030032
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.