
Role of ADCC, CDC, and CDCC in Vaccine-Mediated Protection against Her2 Mammary Carcinogenesis

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Cells
2.3. Immunization and Tumor Growth Monitoring
2.4. Antibody Response
2.5. Identification of Tumor-Infiltrating Leukocytes
2.6. Morphological Analysis
2.7. Histology Immunohistochemistry and Immunofluorescence
2.8. Statistical Analysis
3. Results
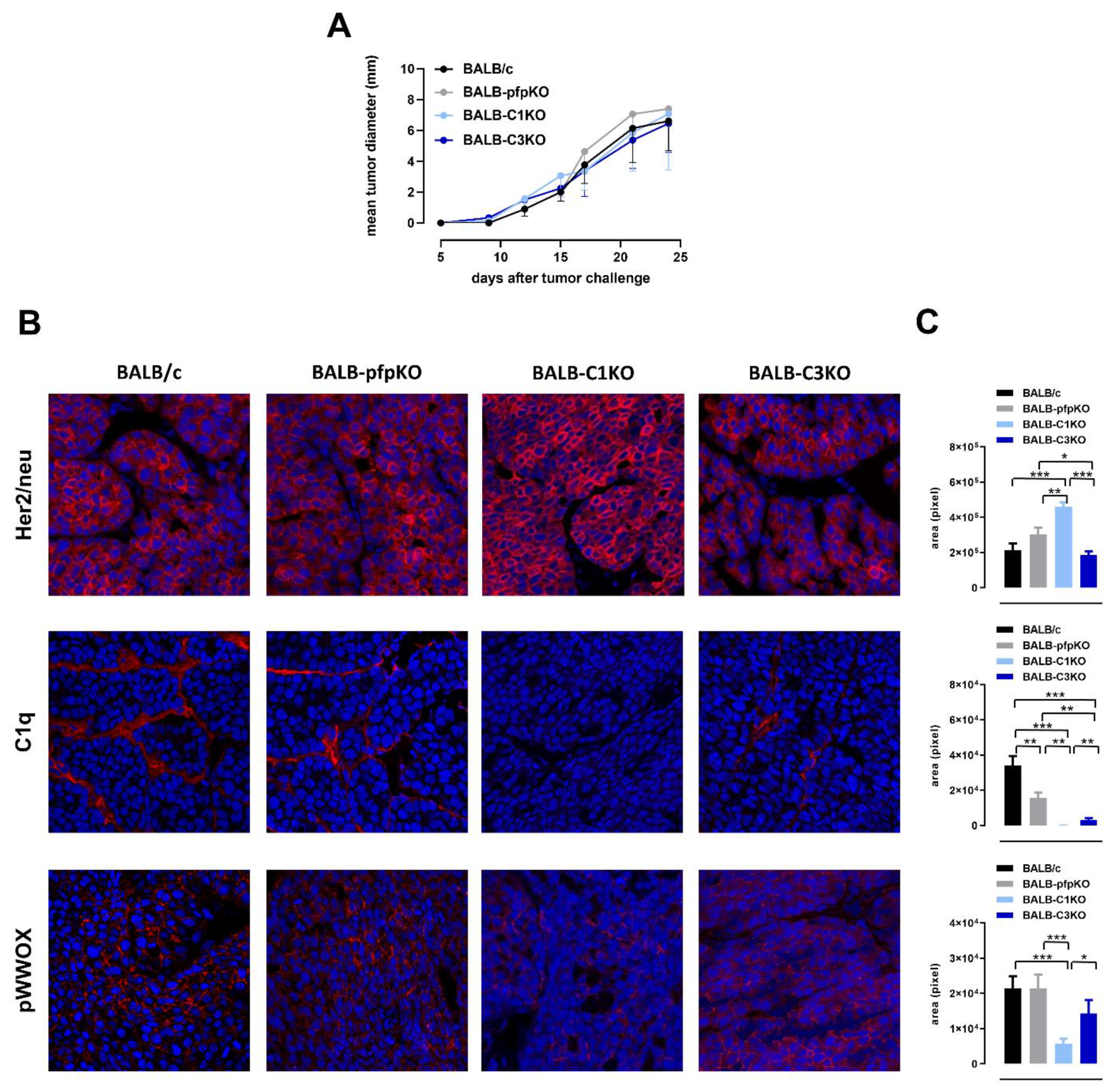
3.1. Perforin and Complement Deficiencies Do Not Affect the Growth of Transplantable Her2+ TUBO Tumors
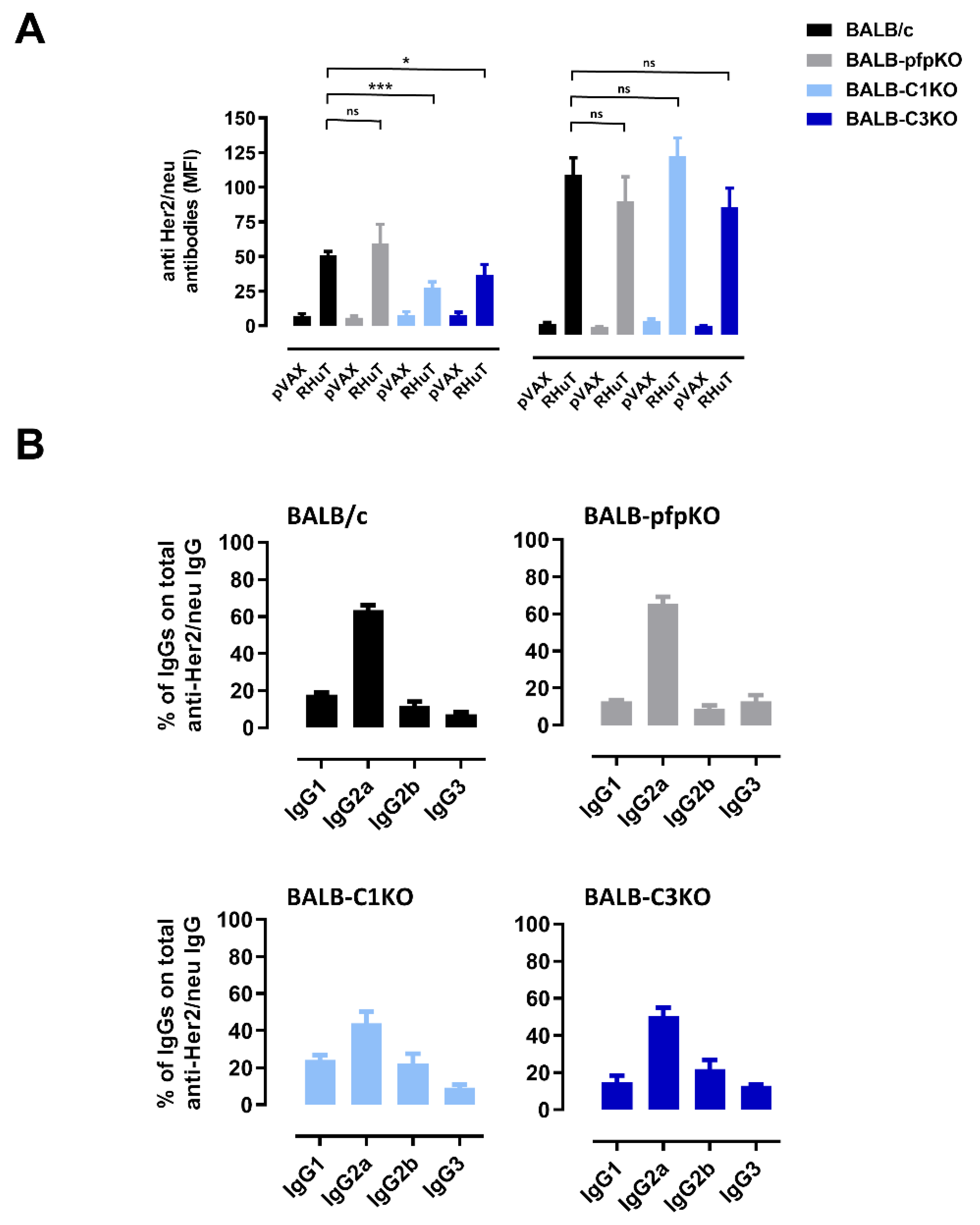
3.2. Perforin- and Complement-Deficient Mice Can Be Effectively Immunized against Her2
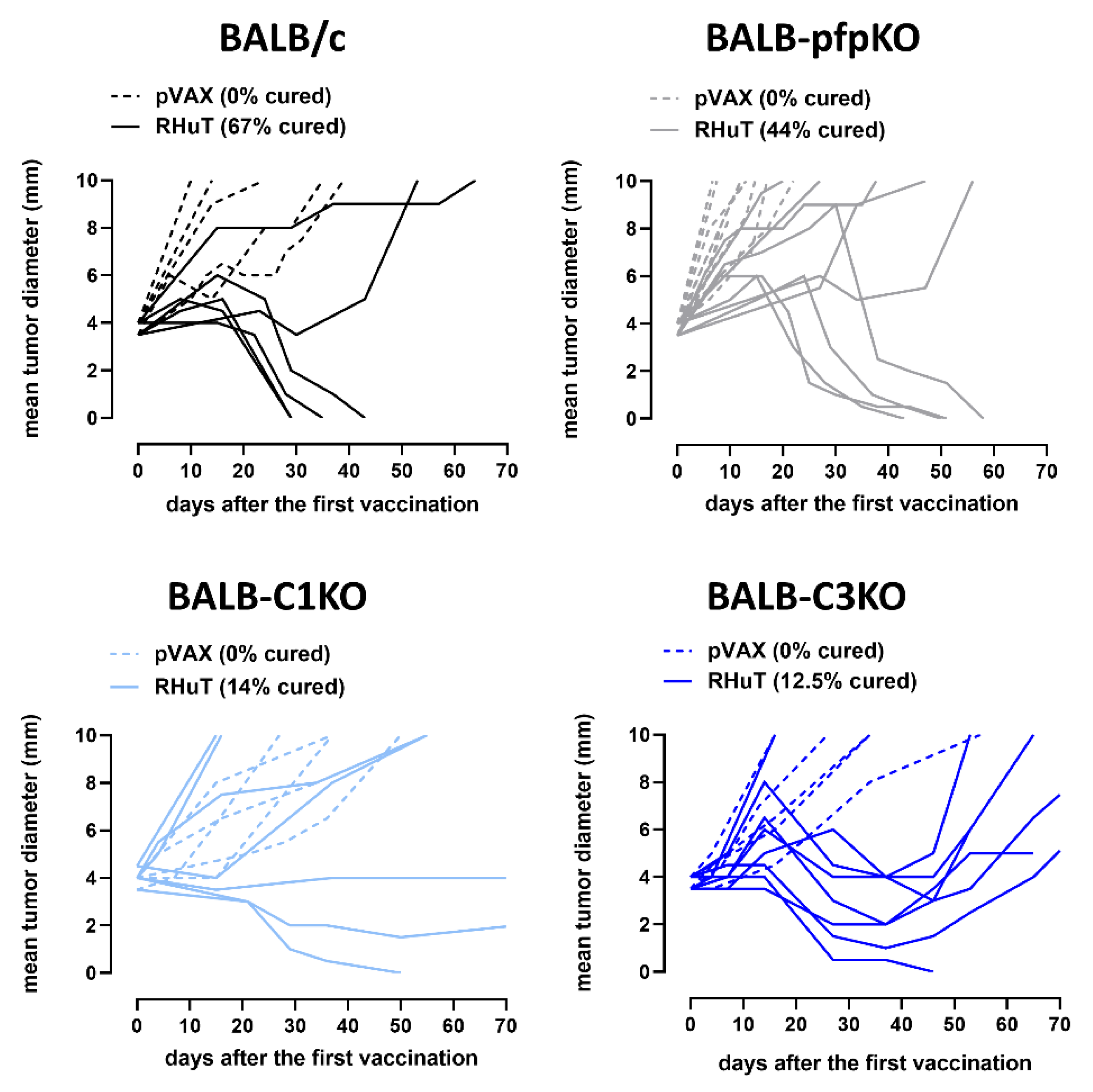
3.3. RHuT Vaccination in Perforin- and Complement-Deficient Mice Impacts the Growth of Her2+ TUBO Tumors
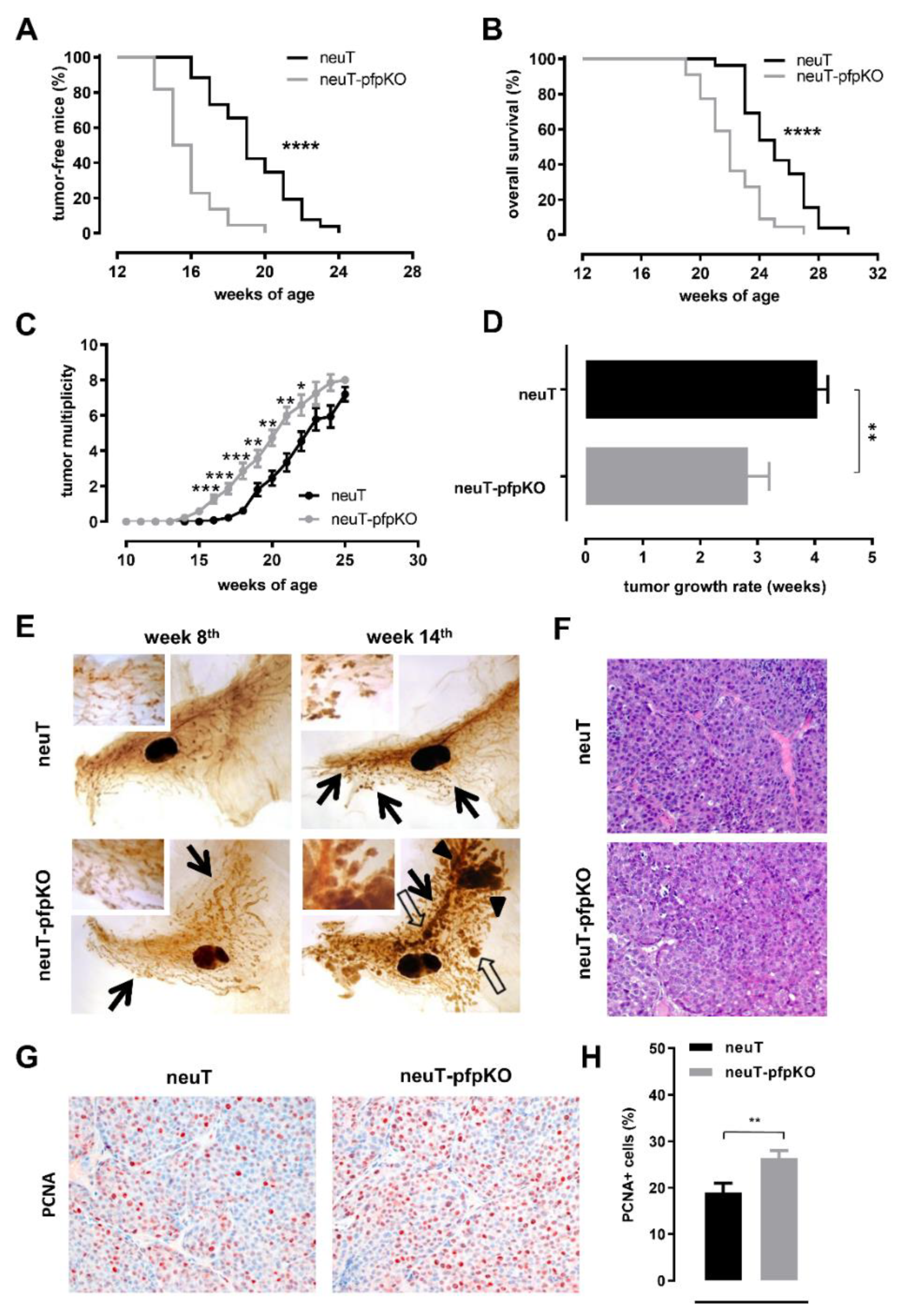
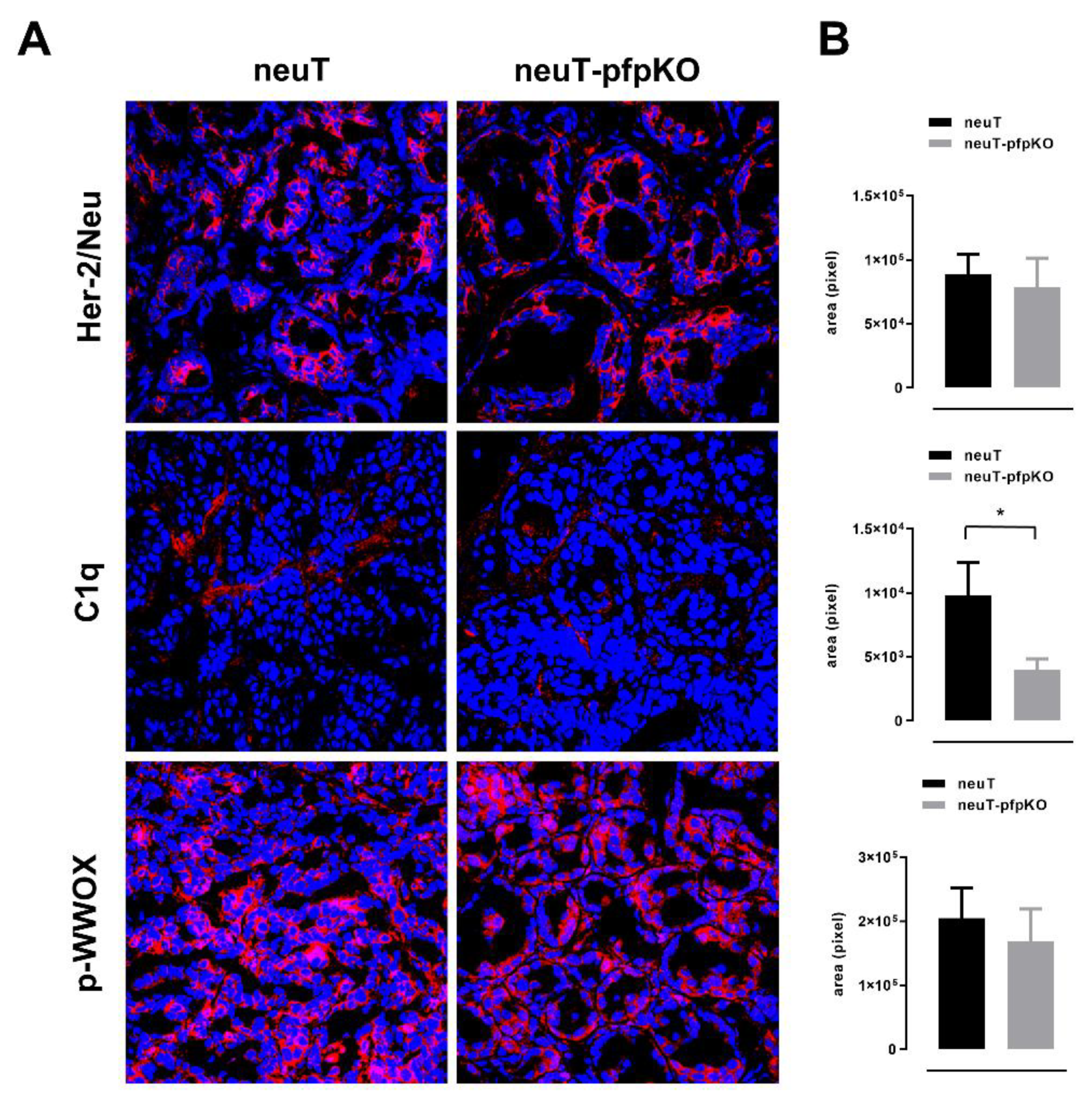
3.4. Perforin Deficiency Induces an Earlier Onset and an Accelerated Growth of Autochthonous Her2+ Carcinomas in neuT Female Mice
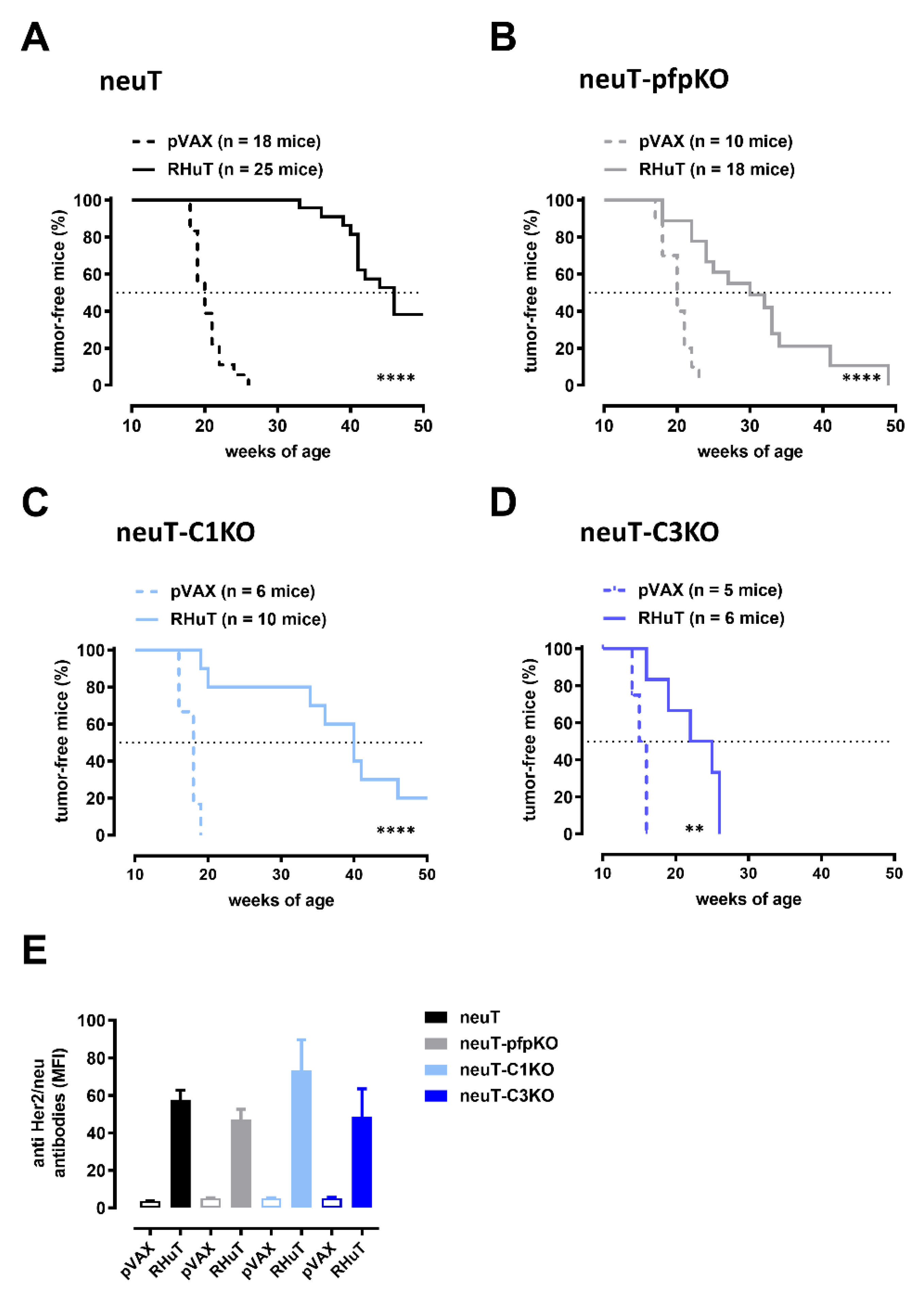
3.5. Perforin- and Complement-Mediated Mechanisms Contribute to the Antitumor Effects of Vaccine-Induced Anti-Rat Her2 Antibodies in neuT Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karunagaran, D.; Tzahar, E.; Beerli, R.R.; Chen, X.; Graus-Porta, D.; Ratzkin, B.J.; Seger, R.; Hynes, N.E.; Yarden, Y. ErbB-2 is a common auxiliary subunit of NDF and EGF receptors: Implications for breast cancer. EMBO J. 1996, 15, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Behravan, J.; Razazan, A.; Behravan, G. Towards Breast Cancer Vaccines, Progress and Challenges. Curr. Drug Discov. Technol. 2019, 16, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Whenham, N.; D’Hondt, V.; Piccart, M.J. HER2-positive breast cancer: From trastuzumab to innovatory anti-HER2 strategies. Clin. Breast Cancer. 2008, 8, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Collins, D.M.; Madden, S.F.; Gaynor, N.; Alsultan, D.; Le Gal, M.; Eustace, A.J.; Gately, K.A.; Hughes, C.; Davies, A.M.; Mahgoub, T.; et al. Effects of HER Family–targeting Tyrosine Kinase Inhibitors on Antibody-dependent Cell-mediated Cytotoxicity in HER2-expressing Breast Cancer. Clin. Cancer Res. 2020, 27, 807–818. [Google Scholar] [CrossRef]
- Rinnerthaler, G.; Gampenrieder, S.P.; Greil, R. HER2 Directed Antibody-Drug-Conjugates beyond T-DM1 in Breast Cancer. Int. J. Mol. Sci. 2019, 20, 1115. [Google Scholar] [CrossRef] [Green Version]
- Vu, T.; Claret, F.X. Trastuzumab: Updated Mechanisms of Action and Resistance in Breast Cancer. Front. Oncol. 2012, 2, 62. [Google Scholar] [CrossRef] [Green Version]
- Scott, A.M.; Wolchok, J.D.; Old, L.J. Antibody therapy of cancer. Nat. Cancer 2012, 12, 278–287. [Google Scholar] [CrossRef]
- Mamidi, S.; Höne, S.; Kirschfink, M. The complement system in cancer: Ambivalence between tumour destruction and promotion. Immunobiology 2017, 222, 45–54. [Google Scholar] [CrossRef]
- Wang, B.; Yang, C.; Jin, X.; Du, Q.; Wu, H.; Dall’Acqua, W.; Mazor, Y. Regulation of antibody-mediated complement-dependent cytotoxicity by modulating the intrinsic affinity and binding valency of IgG for target antigen. mAbs 2019, 12, 1690959. [Google Scholar] [CrossRef] [Green Version]
- Jagosky, M.; Tan, A.R. Combination of Pertuzumab and Trastuzumab in the Treatment of HER2-Positive Early Breast Cancer: A Review of the Emerging Clinical Data. Breast Cancer Targets Ther. 2021, 13, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Spiridon, C.I.; Ghetie, M.A.; Uhr, J.; Marches, R.; Li, J.L.; Shen, G.L.; Vitetta, E.S. Targeting multiple Her-2 epitopes with monoclonal antibodies results in improved antigrowth activity of a human breast cancer cell line in vitro and in vivo. Clin. Cancer Res. 2002, 8, 1720–1730. [Google Scholar] [PubMed]
- Exman, P.; Tolaney, S.M. HER2-positive metastatic breast cancer: A comprehensive review. Clin. Adv. Hematol. Oncol. 2021, 19, 40–50. [Google Scholar] [PubMed]
- Arab, A.; Yazdian-Robati, R.; Behravan, J. HER2-Positive Breast Cancer Immunotherapy: A Focus on Vaccine Development. Arch. Immunol. Ther. Exp. 2020, 68, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Pallerla, S.; Abdul, A.; Comeau, J.; Jois, S. Cancer Vaccines, Treatment of the Future: With Emphasis on HER2-Positive Breast Cancer. Int. J. Mol. Sci. 2021, 22, 779. [Google Scholar] [CrossRef] [PubMed]
- Triulzi, C.; Vertuani, S.; Curcio, C.; Antognoli, A.; Seibt, J.; Akusjarvi, G.; Wei, W.-Z.; Cavallo, F.; Kiessling, R. Antibody-dependent natural killer cell-mediated cytotoxicity engendered by a kinase-inactive human HER2 adenovirus-based vaccination mediates resistance to breast tumors. Cancer Res. 2010, 70, 7431–7441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curcio, C.; Di Carlo, E.; Clynes, R.; Smyth, M.J.; Boggio, K.; Quaglino, E.; Spadaro, M.; Colombo, M.P.; Amici, A.; Lollini, P.-L.; et al. Nonredundant roles of antibody, cytokines, and perforin in the eradication of established Her-2/neu carcinomas. J. Clin. Invest. 2003, 111, 1161–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voskoboinik, I.; Whisstock, J.C.; Trapani, J.A. Perforin and granzymes: Function, dysfunction and human pathology. Nat. Rev. Immunol. 2015, 15, 388–400. [Google Scholar] [CrossRef]
- Meyer, S.; Leusen, J.H.W.; Boross, P. Regulation of complement and modulation of its activity in monoclonal antibody therapy of cancer. mAbs 2014, 6, 1133–1144. [Google Scholar] [CrossRef] [Green Version]
- Quaglino, E.; Mastini, C.; Amici, A.; Marchini, C.; Iezzi, M.; Lanzardo, S.; De Giovanni, C.; Montani, M.; Lollini, P.L.; Masucci, G.; et al. A Better Immune Reaction to Erbb-2 Tumors Is Elicited in Mice by DNA Vaccines Encoding Rat/Human Chimeric Proteins. Cancer Res. 2010, 70, 2604–2612. [Google Scholar] [CrossRef] [Green Version]
- Jacob, J.B.; Quaglino, E.; Radkevich-Brown, O.; Jones, R.F.; Piechocki, M.P.; Reyes, J.D.; Weise, A.; Amici, A.; Wei, W.-Z. Combining human and rat sequences in her-2 DNA vaccines blunts immune tolerance and drives antitumor immunity. Cancer Res. 2010, 70, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Barutello, G.; Curcio, C.; Spadaro, M.; Arigoni, M.; Trovato, R.; Bolli, E.; Zheng, Y.; Ria, F.; Quaglino, E.; Iezzi, M.; et al. Antitumor immunization of mothers delays tumor development in cancer-prone offspring. OncoImmunology 2015, 4, e1005500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quaglino, E.; Mastini, C.; Forni, G.; Cavallo, F. ErbB2 Transgenic Mice: A Tool for Investigation of the Immune Prevention and Treatment of Mammary Carcinomas. Curr. Protoc. Immunol. 2008, 80, 20.9.1–20.9.10. [Google Scholar] [CrossRef]
- Hüsemann, Y.; Geigl, J.; Schubert, F.; Musiani, P.; Meyer, M.; Burghart, E.; Forni, G.; Eils, R.; Fehm, T.; Riethmüller, G.; et al. Systemic Spread Is an Early Step in Breast Cancer. Cancer Cell 2008, 13, 58–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rovero, S.; Amici, A.; Di Carlo, E.; Bei, R.; Nanni, P.; Quaglino, E.; Porcedda, P.; Boggio, K.; Smorlesi, A.; Lollini, P.-L.; et al. DNA Vaccination Against Rat Her-2/Neu p185 More Effectively Inhibits Carcinogenesis Than Transplantable Carcinomas in Transgenic BALB/c Mice. J. Immunol. 2000, 165, 5133–5142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quaglino, E.; Iezzi, M.; Mastini, C.; Amici, A.; Pericle, F.; DI Carlo, E.; Pupa, S.; De Giovanni, C.; Spadaro, M.; Curcio, C.; et al. Electroporated DNA Vaccine Clears Away Multifocal Mammary Carcinomas in Her-2/neu Transgenic Mice. Cancer Res. 2004, 64, 2858–2864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacca, S.; Rolih, V.; Quaglino, E.; Franceschi, V.; Tebaldi, G.; Bolli, E.; Rosamilia, A.; Ottonello, S.; Cavallo, F.; Donofrio, G. Bovine herpesvirus 4-based vector delivering a hybrid rat/human HER-2 oncoantigen efficiently protects mice from autochthonous Her-2+ mammary cancer. OncoImmunology 2015, 5, e1082705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, L.; Bolli, E.; Di Lorenzo, A.; Franceschi, V.; Macchi, F.; Riccardo, F.; Ruiu, R.; Russo, L.; Quaglino, E.; Donofrio, G.; et al. Immunotargeting of the xCT Cystine/Glutamate Antiporter Potentiates the Efficacy of HER2-Targeted Immunotherapies in Breast Cancer. Cancer Immunol. Res. 2020, 8, 1039–1053. [Google Scholar] [CrossRef]
- Porzia, A.; Lanzardo, S.; Citti, A.; Cavallo, F.; Forni, G.; Santoni, A.; Galandrini, R.; Paolini, R. Attenuation of PI3K/Akt-Mediated Tumorigenic Signals through PTEN Activation by DNA Vaccine-Induced Anti-ErbB2 Antibodies. J. Immunol. 2010, 184, 4170–4177. [Google Scholar] [CrossRef] [Green Version]
- Street, S.E.A.; Zerafa, N.; Iezzi, M.; Westwood, J.A.; Stagg, J.; Musiani, P.; Smyth, M. Host Perforin Reduces Tumor Number but Does Not Increase Survival in Oncogene-Driven Mammary Adenocarcinoma. Cancer Res. 2007, 67, 5454–5460. [Google Scholar] [CrossRef] [Green Version]
- Macagno, M.; Bandini, S.; Stramucci, L.; Quaglino, E.; Conti, L.; Balmas, E.; Smyth, M.J.; Lollini, P.-L.; Musiani, P.; Forni, G.; et al. Multiple roles of perforin in hampering ERBB-2 (Her-2/neu) carcinogenesis in transgenic male mice. J. Immunol. 2014, 192, 5434–5441. [Google Scholar] [CrossRef] [Green Version]
- Trendelenburg, M.; Fossati-Jimack, L.; Cortes-Hernandez, J.; Turnberg, D.; Lewis, M.; Izui, S.; Cook, H.T.; Botto, M. The role of complement in cryoglobulin-induced immune complex glomerulonephritis. J. Immunol. 2005, 175, 6909–6914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandini, S.; Macagno, M.; Hysi, A.; Lanzardo, S.; Conti, L.; Bello, A.; Riccardo, F.; Ruiu, R.; Merighi, I.F.; Forni, G.; et al. The non-inflammatory role of C1q during Her2/neu-driven mammary carcinogenesis. OncoImmunology 2016, 5, e1253653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandini, S.; Curcio, C.; Macagno, M.; Quaglino, E.; Arigoni, M.; Lanzardo, S.; Hysi, A.; Barutello, G.; Consolino, L.; Longo, D.L.; et al. Early onset and enhanced growth of autochthonous mammary carcinomas in C3-deficient Her2/neu transgenic mice. OncoImmunology 2013, 2, e26137. [Google Scholar] [CrossRef] [Green Version]
- Jacob, J.; Radkevich, O.; Forni, G.; Zielinski, J.; Shim, D.; Jones, R.F.; Wei, W.-Z. Activity of DNA vaccines encoding self or heterologous Her-2/neu in Her-2 or neu transgenic mice. Cell. Immunol. 2006, 240, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Quaglino, E.; Riccardo, F.; Macagno, M.; Bandini, S.; Cojoca, R.; Ercole, E.; Amici, A.; Cavallo, F. Chimeric DNA Vaccines against ErbB2+ Carcinomas: From Mice to Humans. Cancers 2011, 3, 3225–3241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudis, C.A. Trastuzumab--mechanism of action and use in clinical practice. N. Engl. J. Med. 2007, 357, 39–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quaglino, E.; Rolla, S.; Iezzi, M.; Spadaro, M.; Musiani, P.; De Giovanni, C.; Lollini, P.L.; Lanzardo, S.; Forni, G.; Sanges, R.; et al. Concordant morphologic and gene expression data show that a vaccine halts HER-2/neu preneoplastic lesions. J. Clin. Investig. 2004, 113, 709–717. [Google Scholar] [CrossRef]
- Bulla, R.; Tripodo, C.; Rami, D.; Ling, G.S.; Agostinis, C.; Guarnotta, C.; Zorzet, S.; Durigutto, P.; Botto, M.; Tedesco, F. C1q acts in the tumour microenvironment as a cancer-promoting factor independently of complement activation. Nat. Commun. 2016, 7, 10346. [Google Scholar] [CrossRef] [Green Version]
- Riihilä, P.; Viiklepp, K.; Nissinen, L.; Farshchian, M.; Kallajoki, M.; Kivisaari, A.; Meri, S.; Peltonen, J.; Kähäri, V.; Peltonen, S. Tumour-cell-derived complement components C1r and C1s promote growth of cutaneous squamous cell carcinoma. Br. J. Dermatol. 2019, 182, 658–670. [Google Scholar] [CrossRef]
- Csomor, E.; Bajtay, Z.; Sándor, N.; Kristóf, K.; Arlaud, G.J.; Thiel, S.; Erdei, A. Complement protein C1q induces maturation of human dendritic cells. Mol. Immunol. 2007, 44, 3389–3397. [Google Scholar] [CrossRef]
- Bajtay, Z.; Csomor, E.; Sándor, N.; Erdei, A. Expression and role of Fc- and complement-receptors on human dendritic cells. Immunol. Lett. 2006, 104, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Baruah, P.; Dumitriu, I.E.; Malik, T.H.; Cook, H.T.; Dyson, J.; Scott, D.; Simpson, E.; Botto, M. C1q enhances IFN-gamma production by antigen-specific T cells via the CD40 costimulatory pathway on dendritic cells. Blood. 2009, 113, 3485–3493. [Google Scholar] [CrossRef] [PubMed]
- Carroll, M.C. Complement and humoral immunity. Vaccine 2008, 26, I28–I33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Broek, M.; Kägi, D.; Ossendorp, F.; Toes, R.; Vamvakas, S.; Lutz, W.K.; Melief, C.J.; Zinkernagel, R.M.; Hengartner, H. Decreased tumor surveillance in perforin-deficient mice. J. Exp. Med. 1996, 184, 1781–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prang, N.; Preithner, S.; Brischwein, K.; Göster, P.; Wöppel, A.; Müller, J.; Steiger, C.; Peters, M.; Baeuerle, P.A.; Da Silva, A.J. Cellular and complement-dependent cytotoxicity of Ep-CAM-specific monoclonal antibody MT201 against breast cancer cell lines. Br. J. Cancer 2005, 92, 342–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petricevic, B.; Laengle, J.; Singer, J.; Sachet, M.; Fazekas, J.; Steger, G.; Bartsch, R.; Jensen-Jarolim, E.; Bergmann, M. Trastuzumab mediates antibody-dependent cell-mediated cytotoxicity and phagocytosis to the same extent in both adjuvant and metastatic HER2/neu breast cancer patients. J. Transl. Med. 2013, 11, 307. [Google Scholar] [CrossRef] [Green Version]
- Tallerico, R.; Conti, L.; Lanzardo, S.; Sottile, R.; Garofalo, C.; Wagner, A.; Johansson, M.H.; Cristiani, C.M.; Kärre, K.; Carbone, E.; et al. NK cells control breast cancer and related cancer stem cell hematological spread. Oncoimmunology 2017, 6, e1284718. [Google Scholar] [CrossRef] [Green Version]
- Salemme, V.; Centonze, G.; Cavallo, F.; Defilippi, P.; Conti, L. The Crosstalk Between Tumor Cells and the Immune Microenvironment in Breast Cancer: Implications for Immunotherapy. Front. Oncol. 2021, 11, 289. [Google Scholar] [CrossRef]
- Lee, S.-C.; Shimasaki, N.; Lim, J.S.; Wong, A.L.; Yadav, K.; Yong, W.P.; Tan, L.K.; Koh, L.P.; Poon, M.L.; Tan, S.H.; et al. Phase I Trial of Expanded, Activated Autologous NK-cell Infusions with Trastuzumab in Patients with HER2-positive Cancers. Clin. Cancer Res. 2020, 26, 4494–4502. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mouse Strain | Tumor Take/Mice Vaccinated with: | |
|---|---|---|
| pVAX | RHuT | |
| BALB/c | 8/8 (0%) a | 0/8 (100%) a |
| BALB-pfpKO | 6/6 (0%) | 0/5 (100%) |
| BALB-C1KO | 8/8 (0%) | 0/6 (100%) |
| BALB-C3KO | 6/6 (0%) | 0/8 (100%) |
| BALB-C1KO-pfpKO | 5/5 (0%) | 0/6 (100%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macagno, M.; Bandini, S.; Bolli, E.; Bello, A.; Riccardo, F.; Barutello, G.; Merighi, I.F.; Forni, G.; Lamolinara, A.; Del Pizzo, F.; et al. Role of ADCC, CDC, and CDCC in Vaccine-Mediated Protection against Her2 Mammary Carcinogenesis. Biomedicines 2022, 10, 230. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10020230
Macagno M, Bandini S, Bolli E, Bello A, Riccardo F, Barutello G, Merighi IF, Forni G, Lamolinara A, Del Pizzo F, et al. Role of ADCC, CDC, and CDCC in Vaccine-Mediated Protection against Her2 Mammary Carcinogenesis. Biomedicines. 2022; 10(2):230. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10020230
Chicago/Turabian StyleMacagno, Marco, Silvio Bandini, Elisabetta Bolli, Amanda Bello, Federica Riccardo, Giuseppina Barutello, Irene Fiore Merighi, Guido Forni, Alessia Lamolinara, Francesco Del Pizzo, and et al. 2022. "Role of ADCC, CDC, and CDCC in Vaccine-Mediated Protection against Her2 Mammary Carcinogenesis" Biomedicines 10, no. 2: 230. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10020230