
Differential Expression of Genes Involved in Metabolism and Immune Response in Diffuse and Intestinal Gastric Cancers, a Pilot Ptudy

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Patients and Tissue Samples
2.2. Total RNA Preparation and Real-Time RT-PCR
2.3. Statistical Analysis
2.4. Immunocytochemistry
3. Results
3.1. Patient’s Characteristics
3.2. Expression of PD-L1, PD-L2 and PD1 in Gastric Cancers
3.3. Expression of IDO1, IDO2 and TDO2 in Gastric Cancers
3.4. Correlation of Expression between PD-L1 and IDO1 in Gastric Cancers
3.5. High AhR Expression in Gastric Cancers
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and mfajor patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Qu, Y.P.; Hou, P. Pathogenetic mechanisms in gastric cancer. World J. Gastroenterol. 2014, 20, 13804–13819. [Google Scholar] [CrossRef]
- Lauren, P. The Two Histological Main Types of Gastric Carcinoma: Diffuse and So-Called Intestinal-Type Carcinoma. An Attempt at a Histo-Clinical Classification. Acta Pathol. Microbiol. Scand. 1965, 64, 31–49. [Google Scholar] [CrossRef] [PubMed]
- Uemura, N.; Okamoto, S.; Yamamoto, S.; Matsumura, N.; Yamaguchi, S.; Yamakido, M.; Taniyama, K.; Sasaki, N.; Schlemper, R.J. Helicobacter pylori infection and the development of gastric cancer. N. Engl. J. Med. 2001, 345, 784–789. [Google Scholar] [CrossRef] [PubMed]
- Henson, D.E.; Dittus, C.; Younes, M.; Nguyen, H.; Albores-Saavedra, J. Differential trends in the intestinal and diffuse types of gastric carcinoma in the United States, 1973–2000: Increase in the signet ring cell type. Arch. Pathol. Lab. Med. 2004, 128, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Jezequel, J.; Bessaguet, C.; Verveur, C.; Faycal, J.; Richert, Z.; Metges, J.P.; Volant, A.; Nousbaum, J.B.; Robaszkiewicz, M. Trends in incidence, management, and survival of gastric and cardia carcinomas in the area of Finistere (France) between 1984 and 2003. Eur. J. Gastroenterol. Hepatol. 2010, 22, 1412–1419. [Google Scholar] [PubMed]
- Li, C.; Kim, S.; Lai, J.F.; Hyung, W.J.; Choi, W.H.; Choi, S.H.; Noh, S.H. Advanced gastric carcinoma with signet ring cell histology. Oncology 2007, 72, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Taghavi, S.; Jayarajan, S.N.; Davey, A.; Willis, A.I. Prognostic significance of signet ring gastric cancer. J. Clin. Oncol. 2012, 30, 3493–3498. [Google Scholar] [CrossRef] [Green Version]
- Pernot, S.; Voron, T.; Perkins, G.; Lagorce-Pages, C.; Berger, A.; Taieb, J. Signet-ring cell carcinoma of the stomach: Impact on prognosis and specific therapeutic challenge. World J. Gastroenterol. 2015, 21, 11428–11438. [Google Scholar] [CrossRef]
- Voron, T.; Messager, M.; Duhamel, A.; Lefevre, J.; Mabrut, J.Y.; Goere, D.; Meunier, B.; Brigand, C.; Hamy, A.; Glehen, O.; et al. Is signet-ring cell carcinoma a specific entity among gastric cancers? Gastric Cancer 2016, 19, 1027–1040. [Google Scholar] [CrossRef] [Green Version]
- Guilford, P.; Hopkins, J.; Harraway, J.; McLeod, M.; McLeod, N.; Harawira, P.; Taite, H.; Scoular, R.; Miller, A.; Reeve, A.E. E-cadherin germline mutations in familial gastric cancer. Nature 1998, 392, 402–405. [Google Scholar] [CrossRef] [PubMed]
- Humar, B.; Blair, V.; Charlton, A.; More, H.; Martin, I.; Guilford, P. E-cadherin deficiency initiates gastric signet-ring cell carcinoma in mice and man. Cancer Res. 2009, 69, 2050–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakiuchi, M.; Nishizawa, T.; Ueda, H.; Gotoh, K.; Tanaka, A.; Hayashi, A.; Yamamoto, S.; Tatsuno, K.; Katoh, H.; Watanabe, Y.; et al. Recurrent gain-of-function mutations of RHOA in diffuse-type gastric carcinoma. Nat. Genet. 2014, 46, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Perrot-Applanat, M.; Vacher, S.; Pimpie, C.; Chemlali, W.; Derieux, S.; Pocard, M.; Bieche, I. Differential gene expression in growth factors, epithelial mesenchymal transition and chemotaxis in the diffuse type compared with the intestinal type of gastric cancer. Oncol. Lett. 2019, 18, 674–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piessen, G.; Messager, M.; Leteurtre, E.; Jean-Pierre, T.; Mariette, C. Signet ring cell histology is an independent predictor of poor prognosis in gastric adenocarcinoma regardless of tumoral clinical presentation. Ann. Surg. 2009, 250, 878–887. [Google Scholar] [CrossRef] [PubMed]
- Messager, M.; Lefevre, J.H.; Pichot-Delahaye, V.; Souadka, A.; Piessen, G.; Mariette, C.; FREGAT Working Group. The impact of perioperative chemotherapy on survival in patients with gastric signet ring cell adenocarcinoma: A multicenter comparative study. Ann. Surg. 2011, 254, 684–693, discussion 693. [Google Scholar] [CrossRef] [PubMed]
- Ohta, H.; Aoyagi, K.; Fukaya, M.; Danjoh, I.; Ohta, A.; Isohata, N.; Saeki, N.; Taniguchi, H.; Sakamoto, H.; Shimoda, T.; et al. Cross talk between hedgehog and epithelial-mesenchymal transition pathways in gastric pit cells and in diffuse-type gastric cancers. Br. J. Cancer 2009, 100, 389–398. [Google Scholar] [CrossRef]
- Ishimoto, T.; Miyake, K.; Nandi, T.; Yashiro, M.; Onishi, N.; Huang, K.K.; Joyce, L.S.; Kalpana, R.; Tay, S.T.; Suzuki, Y.; et al. Activation of Transforming Growth Factor Beta 1 Signaling in Gastric Cancer-associated Fibroblasts Increases Their Motility, via Expression of Rhomboid 5 Homolog 2, and Ability to Induce Invasiveness of Gastric Cancer Cells. Gastroenterology 2017, 153, 191–204.e16. [Google Scholar] [CrossRef]
- Ayers, M.; Lunceford, J.; Nebozhyn, M.; Murphy, E.; Loboda, A.; Kaufman, D.R.; Albright, A.; Cheng, J.D.; Kang, S.P.; Shankaran, V.; et al. IFN-gamma-related mRNA profile predicts clinical response to PD-1 blockade. J. Clin. Investig. 2017, 127, 2930–2940. [Google Scholar] [CrossRef]
- Seliger, B. Basis of PD1/PD-L1 Therapies. J. Clin. Med. 2019, 8, 2168. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.W.; Nam, K.H.; Ahn, S.H.; Park, D.J.; Kim, H.H.; Kim, S.H.; Chang, H.; Lee, J.O.; Kim, Y.J.; Lee, H.S.; et al. Prognostic implications of immunosuppressive protein expression in tumors as well as immune cell infiltration within the tumor microenvironment in gastric cancer. Gastric Cancer 2016, 19, 42–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, J.; Ock, C.Y.; Kim, J.W.; Nam, S.K.; Kwak, Y.; Yun, S.; Ahn, S.H.; Park, D.J.; Kim, H.H.; Kim, W.H.; et al. Clinicopathologic implications of immune classification by PD-L1 expression and CD8-positive tumor-infiltrating lymphocytes in stage II and III gastric cancer patients. Oncotarget 2017, 8, 26356–26367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, X.; Guo, J.; Ding, G.; Li, B.; Dong, B.; Feng, Q.; Li, S.; Zhang, J.; Ying, X.; Cheng, X.; et al. Analysis of PD1, PDL1, PDL2 expression and T cells infiltration in 1014 gastric cancer patients. Oncoimmunology 2018, 7, e1356144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.T.; Cristescu, R.; Bass, A.J.; Kim, K.M.; Odegaard, J.I.; Kim, K.; Liu, X.Q.; Sher, X.; Jung, H.; Lee, M.; et al. Comprehensive molecular characterization of clinical responses to PD-1 inhibition in metastatic gastric cancer. Nat. Med. 2018, 24, 1449–1458. [Google Scholar] [CrossRef]
- Ricci, A.D.; Rizzo, A.; Rojas Llimpe, F.L.; Di Fabio, F.; De Biase, D.; Rihawi, K. Novel HER2-Directed Treatments in Advanced Gastric Carcinoma: AnotHER Paradigm Shift? Cancers 2021, 13, 1664. [Google Scholar] [CrossRef]
- Platten, M.; von Knebel Doeberitz, N.; Oezen, I.; Wick, W.; Ochs, K. Cancer Immunotherapy by Targeting IDO1/TDO and Their Downstream Effectors. Front. Immunol. 2014, 5, 673. [Google Scholar] [CrossRef]
- Opitz, C.A.; Litzenburger, U.M.; Sahm, F.; Ott, M.; Tritschler, I.; Trump, S.; Schumacher, T.; Jestaedt, L.; Schrenk, D.; Weller, M.; et al. An endogenous tumour-promoting ligand of the human aryl hydrocarbon receptor. Nature 2011, 478, 197–203. [Google Scholar] [CrossRef]
- Mezrich, J.D.; Fechner, J.H.; Zhang, X.; Johnson, B.P.; Burlingham, W.J.; Bradfield, C.A. An interaction between kynurenine and the aryl hydrocarbon receptor can generate regulatory T cells. J. Immunol. 2010, 185, 3190–3198. [Google Scholar] [CrossRef] [Green Version]
- Vacher, S.; Castagnet, P.; Chemlali, W.; Lallemand, F.; Meseure, D.; Pocard, M.; Bieche, I.; Perrot-Applanat, M. High AHR expression in breast tumors correlates with expression of genes from several signaling pathways namely inflammation and endogenous tryptophan metabolism. PLoS ONE 2018, 13, e0190619. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Z.; Li, J.; Song, S.; Wang, J.; Cai, W.; Hu, W.; Ji, J.; Zhu, Z.; Zang, L.; Yan, R.; et al. A positive feedback between IDO1 metabolite and COL12A1 via MAPK pathway to promote gastric cancer metastasis. J. Exp. Clin. Cancer Res. 2019, 38, 314. [Google Scholar] [CrossRef] [Green Version]
- Feng, S.; Cao, Z.; Wang, X. Role of aryl hydrocarbon receptor in cancer. Biochim. Biophys. Acta 2013, 1836, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Schlezinger, J.J.; Liu, D.; Farago, M.; Seldin, D.C.; Belguise, K.; Sonenshein, G.E.; Sherr, D.H. A role for the aryl hydrocarbon receptor in mammary gland tumorigenesis. Biol. Chem. 2006, 387, 1175–1187. [Google Scholar] [CrossRef] [PubMed]
- Murray, I.A.; Patterson, A.D.; Perdew, G.H. Aryl hydrocarbon receptor ligands in cancer: Friend and foe. Nat. Rev. Cancer 2014, 14, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Quintana, F.J.; Sherr, D.H. Aryl hydrocarbon receptor control of adaptive immunity. Pharmacol. Rev. 2013, 65, 1148–1161. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Salguero, P.; Pineau, T.; Hilbert, D.M.; McPhail, T.; Lee, S.S.; Kimura, S.; Nebert, D.W.; Rudikoff, S.; Ward, J.M.; Gonzalez, F.J. Immune system impairment and hepatic fibrosis in mice lacking the dioxin-binding Ah receptor. Science 1995, 268, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Stockinger, B.; Di Meglio, P.; Gialitakis, M.; Duarte, J.H. The aryl hydrocarbon receptor: Multitasking in the immune system. Annu. Rev. Immunol. 2014, 32, 403–432. [Google Scholar] [CrossRef] [PubMed]
- Cella, M.; Colonna, M. Aryl hydrocarbon receptor: Linking environment to immunity. Semin. Immunol. 2015, 27, 310–314. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L. AHR Function in Lymphocytes: Emerging Concepts. Trends Immunol. 2016, 37, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Mulero-Navarro, S.; Fernandez-Salguero, P.M. New Trends in Aryl Hydrocarbon Receptor Biology. Front. Cell Dev. Biol. 2016, 4, 45. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Xu, K.; Wu, C.; Wang, Y.; Hu, Y.; Zhu, Y.; Chen, Y.; Shi, Q.; Yu, G.; Zhang, X. PD-L1 expression analysis in gastric carcinoma tissue and blocking of tumor-associated PD-L1 signaling by two functional monoclonal antibodies. Tissue Antigens 2007, 69, 19–27. [Google Scholar] [CrossRef]
- Soliman, H.; Khalil, F.; Antonia, S. PD-L1 expression is increased in a subset of basal type breast cancer cells. PLoS ONE 2014, 9, e88557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masugi, Y.; Nishihara, R.; Hamada, T.; Song, M.; da Silva, A.; Kosumi, K.; Gu, M.; Shi, Y.; Li, W.; Liu, L.; et al. Tumor PDCD1LG2 (PD-L2) Expression and the Lymphocytic Reaction to Colorectal Cancer. Cancer Immunol. Res. 2017, 5, 1046–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.Z.; Zhang, L.; Zhao, X.C.; Gao, S.H.; Qu, L.W.; Yu, H.; Fang, W.F.; Zhou, Y.C.; Liang, F.; Zhang, C.; et al. The Aryl hydrocarbon receptor mediates tobacco-induced PD-L1 expression and is associated with response to immunotherapy. Nat. Commun. 2019, 10, 1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yearley, J.H.; Gibson, C.; Yu, N.; Moon, C.; Murphy, E.; Juco, J.; Lunceford, J.; Cheng, J.; Chow, L.Q.M.; Seiwert, T.Y.; et al. PD-L2 Expression in Human Tumors: Relevance to Anti-PD-1 Therapy in Cancer. Clin. Cancer Res. 2017, 23, 3158–3167. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Patel, K.; Singhi, A.D.; Ren, B.; Zhu, B.; Shaikh, F.; Sun, W. Programmed Death-Ligand 1 Expression Is Common in Gastric Cancer Associated with Epstein-Barr Virus or Microsatellite Instability. Am. J. Surg. Pathol. 2016, 40, 1496–1506. [Google Scholar] [CrossRef] [PubMed]
- Boger, C.; Behrens, H.M.; Mathiak, M.; Kruger, S.; Kalthoff, H.; Rocken, C. PD-L1 is an independent prognostic predictor in gastric cancer of Western patients. Oncotarget 2016, 7, 24269–24283. [Google Scholar] [CrossRef] [Green Version]
- Haas, M.; Dimmler, A.; Hohenberger, W.; Grabenbauer, G.G.; Niedobitek, G.; Distel, L.V. Stromal regulatory T-cells are associated with a favourable prognosis in gastric cancer of the cardia. BMC Gastroenterol. 2009, 9, 65. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.S.; da Silva, E.; Coit, D.G.; Tang, L.H. Intratumoral Immune Response to Gastric Cancer Varies by Molecular and Histologic Subtype. Am. J. Surg. Pathol. 2019, 43, 851–860. [Google Scholar] [CrossRef]
- Pernot, S.; Terme, M.; Radosevic-Robin, N.; Castan, F.; Badoual, C.; Marcheteau, E.; Penault-Llorca, F.; Bouche, O.; Bennouna, J.; Francois, E.; et al. Infiltrating and peripheral immune cell analysis in advanced gastric cancer according to the Lauren classification and its prognostic significance. Gastric Cancer 2020, 23, 73–81. [Google Scholar] [CrossRef]
- Andersson, P.; McGuire, J.; Rubio, C.; Gradin, K.; Whitelaw, M.L.; Pettersson, S.; Hanberg, A.; Poellinger, L. A constitutively active dioxin/aryl hydrocarbon receptor induces stomach tumors. Proc. Nat. Acad. Sci. USA 2002, 99, 9990–9995. [Google Scholar] [CrossRef] [Green Version]
- Safe, S.; Lee, S.O.; Jin, U.H. Role of the aryl hydrocarbon receptor in carcinogenesis and potential as a drug target. Toxicol. Sci. 2013, 135, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, X.F.; Chen, J.; Mao, W.; Wang, Y.H.; Chen, M.H. Downregulation of aryl hydrocarbon receptor expression decreases gastric cancer cell growth and invasion. Oncol. Rep. 2013, 30, 364–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, T.L.; Chen, J.; Mao, W.; Liu, X.; Tao, Y.; Chen, L.Z.; Chen, M.H. Potential therapeutic significance of increased expression of aryl hydrocarbon receptor in human gastric cancer. World J. Gastroenterol. 2009, 15, 1719–1729. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.; Hanieh, H.; Nakahama, T.; Kishimoto, T. The roles of aryl hydrocarbon receptor in immune responses. Int. Immunol. 2013, 25, 335–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julliard, W.; Fechner, J.H.; Mezrich, J.D. The aryl hydrocarbon receptor meets immunology: Friend or foe? A little of both. Front. Immunol. 2014, 5, 458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelante, T.; Iannitti, R.G.; Fallarino, F.; Gargaro, M.; De Luca, A.; Moretti, S.; Bartoli, A.; Romani, L. Tryptophan Feeding of the IDO1-AhR Axis in Host-Microbial Symbiosis. Front. Immunol. 2014, 5, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohinta, S.; Kannan, A.K.; Gowda, K.; Amin, S.G.; Perdew, G.H.; August, A. Differential regulation of Th17 and T regulatory cell differentiation by aryl hydrocarbon receptor dependent xenobiotic response element dependent and independent pathways. Toxicol. Sci. 2015, 145, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Munn, D.H.; Mellor, A.L. IDO in the Tumor Microenvironment: Inflammation, Counter-Regulation, and Tolerance. Trends Immunol. 2016, 37, 193–207. [Google Scholar] [CrossRef] [Green Version]
- Prendergast, G.C.; Smith, C.; Thomas, S.; Mandik-Nayak, L.; Laury-Kleintop, L.; Metz, R.; Muller, A.J. Indoleamine 2,3-dioxygenase pathways of pathogenic inflammation and immune escape in cancer. Cancer Immunol. Immunother. 2014, 63, 721–735. [Google Scholar] [CrossRef]
- Domblides, C.; Lartigue, L.; Faustin, B. Control of the Antitumor Immune Response by Cancer Metabolism. Cells 2019, 8, 104. [Google Scholar] [CrossRef] [Green Version]
- Takikawa, O.; Kuroiwa, T.; Yamazaki, F.; Kido, R. Mechanism of interferon-gamma action. Characterization of indoleamine 2,3-dioxygenase in cultured human cells induced by interferon-gamma and evaluation of the enzyme-mediated tryptophan degradation in its anticellular activity. J. Biol. Chem. 1988, 263, 2041–2048. [Google Scholar] [CrossRef]
- Litzenburger, U.M.; Opitz, C.A.; Sahm, F.; Rauschenbach, K.J.; Trump, S.; Winter, M.; Ott, M.; Ochs, K.; Lutz, C.; Liu, X.; et al. Constitutive IDO expression in human cancer is sustained by an autocrine signaling loop involving IL-6, STAT3 and the AHR. Oncotarget 2014, 5, 1038–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platten, M.; Wick, W.; Van den Eynde, B.J. Tryptophan catabolism in cancer: Beyond IDO and tryptophan depletion. Cancer Res. 2012, 72, 5435–5440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruyama, T.; Kono, K.; Mizukami, Y.; Kawaguchi, Y.; Mimura, K.; Watanabe, M.; Izawa, S.; Fujii, H. Distribution of Th17 cells and FoxP3(+) regulatory T cells in tumor-infiltrating lymphocytes, tumor-draining lymph nodes and peripheral blood lymphocytes in patients with gastric cancer. Cancer Sci. 2010, 101, 1947–1954. [Google Scholar] [CrossRef]
- Li, Q.; Li, Q.; Chen, J.; Liu, Y.; Zhao, X.; Tan, B.; Ai, J.; Zhang, Z.; Song, J.; Shan, B. Prevalence of Th17 and Treg cells in gastric cancer patients and its correlation with clinical parameters. Oncol. Rep. 2013, 30, 1215–1222. [Google Scholar] [CrossRef] [Green Version]
- Puccetti, P. On the Non-Redundant Roles of TDO2 and IDO1. Front. Immunol. 2014, 5, 522. [Google Scholar] [CrossRef] [Green Version]
- D’Amato, N.C.; Rogers, T.J.; Gordon, M.A.; Greene, L.I.; Cochrane, D.R.; Spoelstra, N.S.; Nemkov, T.G.; D’Alessandro, A.; Hansen, K.C.; Richer, J.K. A TDO2-AhR signaling axis facilitates anoikis resistance and metastasis in triple-negative breast cancer. Cancer Res. 2015, 75, 4651–4664. [Google Scholar] [CrossRef] [Green Version]
- Honore, C.; Goere, D.; Messager, M.; Souadka, A.; Dumont, F.; Piessen, G.; Elias, D.; Mariette, C.; FRENCH, F.W.G. Risk factors of peritoneal recurrence in eso-gastric signet ring cell adenocarcinoma: Results of a multicentre retrospective study. Eur. J. Surg. Oncol. 2013, 39, 235–241. [Google Scholar] [CrossRef]


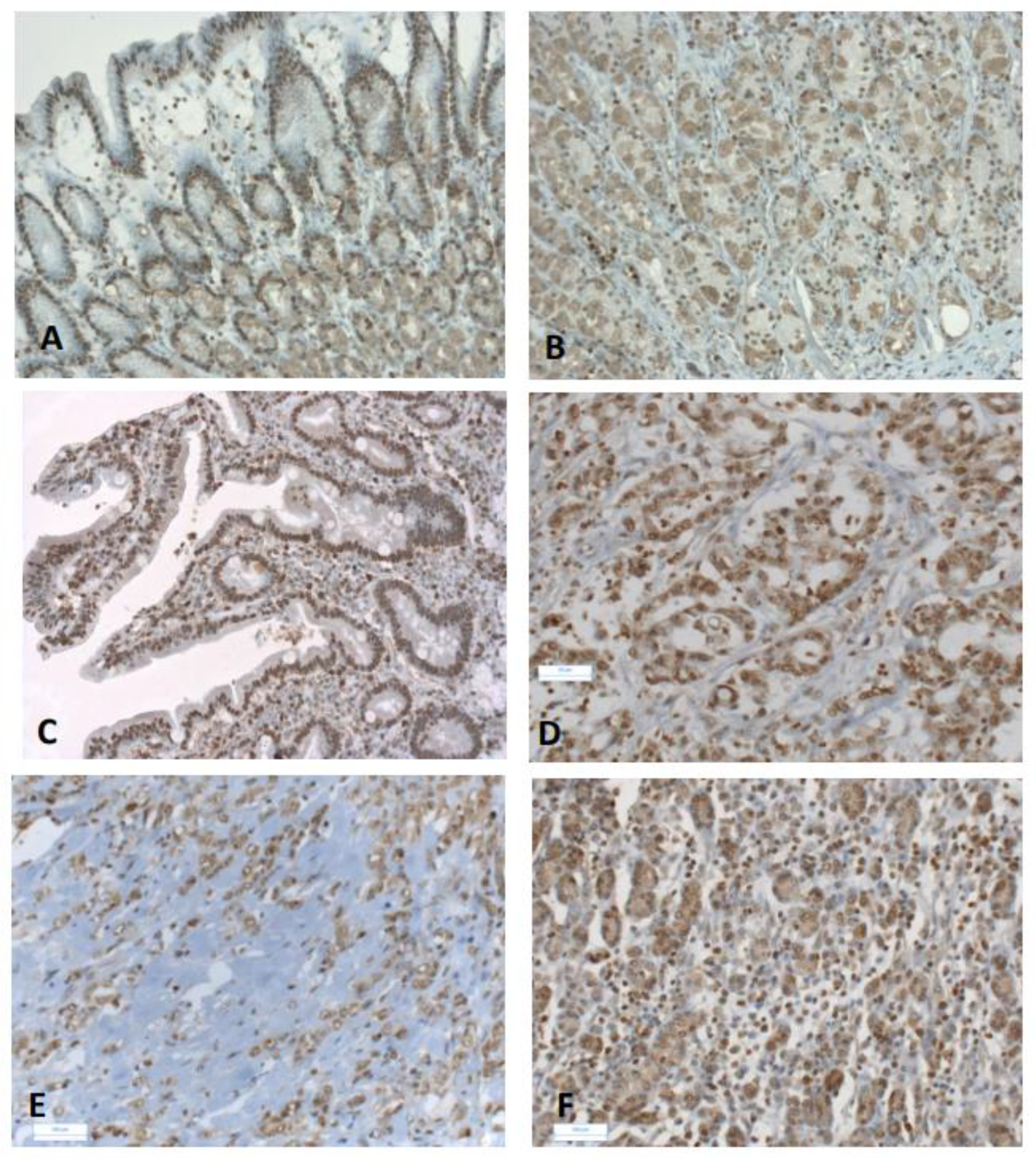{kind=link}
| Total GC (n = 29) | Poorly CohesiveGC (n = 13) (45%) | Intestinal-Subtype GC (n = 16) (n = 55%) | p-Value | |
|---|---|---|---|---|
| Gender, n (%) | ||||
| male | 13/29 | 6/13 (46%) | 7/16 (43%) | 0.90 (NS) a |
| female | 16/29 | 7/13 (54%) | 9/16 (56%) | |
| Age (years, median) | 63 +/−17 | 57(27–71) | 75(59–82) | 0.0004 b |
| Tumor size(mm), n | ||||
| <50 | 10/27 | 4/11 (36%) | 6/16 (37%) | 0.10 (NS) b |
| >=50 | 17/27 | 7/11 (64%) | 10/16 (63%) | 0.95 (NS) a |
| Depth of tumor invasion | ||||
| T1-T2 | 6/29 | 2/13 (15%) | 4/16 (33%) | 0.5 (NS) a |
| T3-T4 | 23/29 | 11/13 (85%) | 12/16 (67%) | |
| Lymphatic invasion, n (%) | ||||
| negative | 11/28 | 1/13 (7%) | 10/15 (67%) | 0.0014 a |
| positive | 17/28 | 12/13 (92%) | 5/15 (33%) | |
| Vascular invasion, n + (%) | ||||
| negative | 9/29 | 3/13 (23%) | 6/16 (38%) | 0.67 (NS) a |
| positive | 20/29 | 10/13 (77%) | 10/16 (62%) | |
| Neural invasion, n (%) | ||||
| negative | 23/29 | 2/13 (15%) | 4/16 (25%) | 0.66 (NS) a |
| positive | 6/29 | 11/13 (68%) | 12/16 (75%) | |
| Metastasis (M), n (%) | ||||
| negative | 24/29 | 9/13 (69%) | 15/16 (94%) | 0.14 (NS) a |
| positive | 5/29 | 4/13 (31%) | 1/16 (6%) | |
| TNM status | ||||
| I-II | 16/29 | 5/13 (38.5%) | 11/16 (69%) | 0.10 (NS) a |
| III-IV | 13/29 | 8/13 (61.5%) | 5/16 (31%) | |
| Smoking | ||||
| negative | 12/22 | 4/12 | 8/12 | 0.77 (NS) a |
| positive | 10/22 | 3/10 | 7/10 |
| Genes | PT (n = 11) | All Tumors (n = 29) | p-Value a | Intestinal-GC vs. PT (n = 16) | p-Value a | Diffuse-GC vs. PT (n = 13) | p-Value a | p-Value subtypes |
|---|---|---|---|---|---|---|---|---|
| Immunity | ||||||||
| PD/PDCD1 | 1 (0.35–2.80) | 1.63 (0.81–5.3) | 0.001 | 1.71 (0.81–5.30) | 0.003 | 1.53 (0.87–3.08) | 0.009 | 0.35 (NS) |
| PDL1/CD274 | 1 (0.57–2.70) | 1.27 (0.52–6.63) | 0.33 (NS) | 1.46 (0.52–6.63) | 0.08 (NS) | 1.03 (0.57–1.56) | 0.84 (NS) | 0.018 |
| PDL2/PDCDL2 | 1 (0.54–1.63) | 1.52 (0.7–2.84) | 0.009 | 1.67 (0.70–2.79) | 0.014 | 1.21 (0.89–2.84) | 0.036 | 0.51 (NS) |
| Trypt metabolism | ||||||||
| IDO1 | 1 (0.19–1.46) | 2.17 (0.34–205) | <0.0001 | 3 (0.34–205) | 0.0006 | 1.96 (0.57–4.78) | 0.002 | 0.14 (NS) |
| TDO2 | 1 (0.45–2.95) | 5.41 (1.36–25.2) | <0.0001 | 7.45 (1.4–25.2) | <0.0001 | 3.33 (1.36–11.9) | 0.0002 | 0.049 |
| Arylhydrocarbon receptor | ||||||||
| AhR | 1 (0.37–1.64) | 1.94 (0.55–3.53) | 0.002 | 1.60 (0.65–3.53) | 0.003 | 2.12 (0.55–3.35) | 0.001 | 0.13 (NS) |
| All Gastric Tumors (n = 29) | Intestinal Sub-Type (n = 16) | Diffuse Sub-Type (n = 13) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| PD1 | PDL1 | PDL2 | PD1 | PDL1 | PDL2 | PD1 | PDL1 | PDL2 | |||
| Gender. | p = 0.25 | p = 0.51 | p = 0.65 | Gender. | p = 0.11 | p = 0.58 | p = 0.15 | Gender. | p = 0.81 | p = 0.80 | p = 0.19 |
| Male (n = 13) | 1.5 (0.8–3.1) | 1.14 (0.5–6.6) | 1.21 (0.7–2.8) | Male (n = 7) | 1.5 (0.81–3.13) | 1.4 (0.52–6.63) | 1.17 (0.7–2.65) | Male (n = 6) | 1.51 (0.87–3.08) | 1.04 (0.62–1.27) | 1.71 (0.89–2.84) |
| Female (n = 16) | 1.7 (1.2–5.3) | 1.33 (0.6–5.5) | 1.53 (0.9–2.8) | Female (n = 9) | 2.18 (1.42–5.3) | 2.03 (0.78–5.55) | 1.83 (1.12–2.8) | Female (n = 7) | 1.58 (1.22–2.01) | 1.03 (0.57–1.56) | 1.16 (0.89–1.83) |
| Age | p = 0.08 | p = 0.18 | p = 0.20 | Age | ND | ND | ND | Age | p = 0.72 | p = 0.12 | p = 0.80 |
| <60 years (n = 9) | 1.49 (0.8–2.0) | 1.09 (0.5–1.6) | 1.21 (0.8–1.8) | <60 years (n = 1) | 0.81 | 0.52 | 0.76 | <60 years (n = 8) | 1.51 (0.87–2.01) | 1.11 (0.62–1.56) | 1.38 (0.89–1.83) |
| >60 years (n = 20) | 1.71 (0.9–5.3) | 1.33 (0.6–6.6) | 1.67 (0.7–2.8) | >60 years (n = 15) | 1.76 (0.94–5.3) | 1.52 (0.8–6.63) | 1.83 (0.7–2.8) | >60 years (n = 5) | 1.58 (1.22–3.08) | 0.78 (0.57–1.27) | 1.16 (0.90–2.84) |
| Tumor invasion | p = 0.74 | p = 0.72 | p = 0.32 | Tumor invasion | p = 0.86 | p > 0.9999 | p > 0.9999 | Tumor invasion | ND | ND | ND |
| T1-T2 (n = 6) | 1.42 (1–5.3) | 1.14 (0.6–85) | 0.93 (0.7–2.8) | T1-T2 (n = 4) | 2.85 (0.94–5.3) | 1.67 (0.96–4.25) | 1.66 (0.7–2.8) | T1-T2 (n = 2) | 1.42 (1.27–1.58) | 0.67 (0.57–0.78) | 0.93 (0.9–0.96) |
| T3-T4 (n = 23) | 1.65 (0.8–3.1) | 1.27 (0.5–6.6) | 1.55 (0.8–2.8) | T3-T4 (n = 12) | 1.71 (0.81–3.13) | 1.46 (0.52–6.63) | 1.67 (0.76–2.65) | T3-T4 (n = 11) | 1.53 (0.9–3.1) | 1.1 (0.62–1.56) | 1.55 (0.89–2.84) |
| Vascular invasion | p = 0.23 | p = 0.48 | p = 0.04 * | Vascular invasion | p = 0.14 | p = 0.56 | p = 0.03 * | Vascular invasion | p > 0.9999 | p = 0.83 | p = 0.79 |
| negative (n = 9) | 1.63 (1.3–5.3) | 1.56 (0.6–6.6) | 1.83 (1.2–2.8) | negative (n = 6) | 2.66 (1.42–5.3) | 3.07 (0.78–6.63) | 2.16 (1.21–2.8) | negative (n = 3) | 1.53 (1.32–1.63) | 0.88 (0.62–1.56) | 1.21 (1.18–1.72) |
| positive (n = 20) | 1.61 (3.1–5.3) | 1.22 (0.5–5.5) | 1.17 (0.7–2.8) | positive (n = 10) | 1.65 (0.81–2.5) | 1.37 (0.52–5.55) | 1.17 (0.7–2.65) | positive (n = 10) | 1.53 (0.87–3.08) | 1.06 (0.57–1.35) | 1.35 (0.89–2.84) |
| Lymphatic invasion | p = 0.05 | p = 0.009 * | p = 0.72 | Lymphatic invasion | p = 0.054 | p = 0.04 * | p = 0.39 | Lymphatic invasion | ND | ND | ND |
| negative (n = 11) | 2.18 (0.9–3.1) | 2.03 (0.6–6.6) | 1.21 (0.7–2.8) | negative (n = 10) | 2.24 (0.94–5.3) | 3.07 (0.96–6.63) | 1.36 (0.7–2.8) | negative (n = 1) | 1.53 | 0.62 | 1.18 |
| positive (n = 17) | 1.49 (0.8–3.1) | 1.03 (0.5–1.6) | 1.71 (0.8–2.9) | positive (n = 5) | 1.48 (0.81–1.66) | 0.87 (0.52–1.52) | 2.22 (0.76–2.65) | positive (n = 12) | 1.53 (0.87–3.08) | 1.06 (0.57–1.56) | 1.38 (0.89–2.84) |
| Metastasis | p = 0.25 | p = 0.15 | p = 0.20 | Metastasis | ND | ND | ND | Metastasis | p = 0.93 | p = 0.82 | p = 0.79 |
| negative (n = 24) | 1.65 (0.9–5.3) | 1.29 (0.6–6.6) | 1.71 (0.7–2.8) | negative (n = 15) | 1.76 (0.94–5.3) | 1.52 (0.8–6.63) | 1.83 (0.7–2.8) | negative (n = 9) | 1.49 (0.87–3.08) | 1.03 (0.57–1.32) | 1.71 (0.89–2.84) |
| positive (n = 5) | 1.53 (0.8–2) | 0.72 (0.5–1.6) | 1.18 (0.8–1.5) | positive (n = 1) | 0.81 | 0.52 | 0.76 | positive (n = 4) | 1.58 (1.22–2.01) | 1.03 (0.62–1.56) | 1.19 (1.16–1.55) |
| TNM | p = 0.37 | p = 0.008 * | p = 0.43 | TNM | p = 0.037 * | p = 0.03 * | p = 0.33 | TNM | p = 0.21 | p = 0.52 | p = 0.80 |
| I-II (n = 16) | 1.8 (0.9–5.3) | 1.33 (0.9–6.6) | 1.36 (0.7–2.8) | I-II (n = 11) | 2.18 (0.94–5.3) | 2.03 (0.96–6.63) | 1.21 (0.7–2.8) | I-II (n = 5) | 1.32 (0.87–1.9) | 1.09 (0.88–1.32) | 1.71 (0.89–1.83) |
| III-IV (n = 13) | 1.53 (0.8–3.1) | 0.87 (0.5–1.6) | 1.55 (0.76–2.8) | III-IV (n = 5) | 1.48 (0.81–1.66) | 0.87 (0.52–1.52) | 2.22 (0.76–2.65) | III-IV (n = 8) | 1.6 (1.22–3.08) | 0.88 (0.57–1.56) | 1.19 (0.9–2.84) |
| EPN | p = 0.13 | p > 0.999 | p = 0.54 | EPN | p = 0.02 * | p = 0.86 | p = 0.11 | EPN | ND | ND | ND |
| negative (n = 6) | 2.1 (1.3–5.3) | 1.26 (0.6–4.25) | 1.67 (0.9–2.8) | negative (n = 4) | 3.5 (1.76–5.3) | 1.69 (1.18–4.25) | 2.16 (1.52–2.8) | negative (n = 2) | 1.42 (1.27–1.58) | 0.67 (0.57–0.78) | 0.93 (0.9–0.96) |
| positive (n = 23) | 1.53 (0.8–3.1) | 1.29 (0.5–6.6) | 1.21 (0.7–2.8) | positive (n= 12) | 1.57 (0.81–3.13) | 1.46 (0.52–6.63) | 1.19 (0.7–2.65) | positive (n = 11) | 1.53 (0.9–3.1) | 1.1 (0.62–1.56) | 1.55 (0.89–2.84) |
| Smoking | p = 0.75 | p = 0.91 | p = 0.0006 * | Smoking | p = 0.45 | p = 0.46 | p = 0.0003 * | Smoking | p = 0.63 | p = 0.57 | p = 0.74 |
| negative (n = 12) | 1.58 (0.9–5.3) | 1.26 (0.6–4.2) | 1.83 (0.9–2.6) | negative (n = 8) | 1.71 (1.42–5.3) | 1.37 (0.78–4.25) | 2.05 (1.52–2.65) | negative (n = 4) | 1.51 (0.87–1.63) | 1.11 (0.62–1.56) | 1.19 (0.89–1.71) |
| positive (n = 10) | 1.61 (0.8–3.1) | 1.33 (0.5–6.6) | 1.04 (0.7–1.5) | positive (n = 7) | 1.65 (0.81–3.13) | 1.54 (0.52–6.63) | 1.12 (0.7–1.21) | positive (n = 3) | 1.58 (1.27–2.01) | 0.78 (0.57–1.35) | 0.96 (0.9–1.55) |
| All Gastric Tumors | IDO1 | TDO2 | Intestinal Sub-Type (n = 16) | IDO1 | TDO2 | Diffuse Sub-Type (n = 13) | IDO1 | TDO2 |
|---|---|---|---|---|---|---|---|---|
| Gender. | p = 0.23 | p = 0.08 | Gender. | p = 0.09 | p = 0.53 | Gender. | p = 0.71 | p = 0.035 * |
| Male (n = 13) | 2 (0.34–44.5) | 8.5 (1.4–25) | Male (n = 7) | 1.77 (0.34–44.5) | 9.45 (1.38–25) | Male (n = 6) | 2.37(0.57–4.78) | 6.82 (1.92–1.9) |
| Female (n = 16) | 2.25 (1.3–205) | 4.23 (1.4–20) | Female (n = 9) | 6.36 (1.43–205) | 6.19 (2.33–20) | Female (n = 7) | 1.75(1.31–3.43) | 2.42 (1.36–5.26) |
| Age | p = 0.06 | p = 0.045 * | Age | ND | ND | Age | p = 0.72 | p = 0.78 |
| <60 years (n = 9) | 1.96 (0.3–3.4) | 3.33 (1.4–7.2) | <60 years (n = 1) | 0.34 | 1.38 | <60 years (n = 8) | 1.98 (0.57–0.43) | 4.16 (1.92–7.17) |
| >60 years (n = 20) | 2.85 (1–205) | 7.45 (1.4–25.2) | >60 years (n = 15) | 3.15 (0.98–205) | 8.5 (2.33–25.2) | >60 years (n = 5) | 1.75 (1.32–4.78) | 2.42(1.36–11.9) |
| Tumor invasion | p = 0.38 | p = 0.74 | Tumor invasion | p = 0.32 | p > 0.9999 | Tumor invasion | ND | ND |
| T1–T2 (n = 6) | 3.53 (1.3–53) | 5.1 (1.6–20) | T1–T2 (n = 4) | 5.14 (3.15–53) | 6.95 (4.73–20) | T1–T2 (n = 2) | 1.4 (1.32–1.5) | 2 (1.6–2.4) |
| T3–T4 (n = 23) | 2 (0.3–205) | 6.19 (1.4–25.2) | T3–T4 (n = 12) | 2.05 (0.34–205) | 7.7 (1.38–25) | T3–T4 (n = 11) | 2 (0.57–4.8) | 5 (1.36–11.9) |
| Vascular invasion | p = 0.33 | p = 0.39 | Vascular invasion | p = 0.18 | p = 0.22 | Vascular invasion | p = 0.32 | p = 0.50 |
| negative (n = 9) | 3.43 (0.6–205) | 6.19 (1.9–20) | negative (n = 6) | 25.4 (1.43–205) | 9.22 (5.4–20) | negative (n = 3) | 0.81 (0.57–3.43) | 2.91 (1.92–5.0) |
| positive (n = 20) | 1.98 (0.3–141) | 5.19 (1.4–25.2) | positive (n = 10) | 2.05 (0.34–141) | 5.78 (1.38–25) | positive (n = 10) | 1.98(1.31–4.78) | 4.29 (1.36–11.9) |
| Lymphatic invasion | p = 0.005 * | p = 0.85 | Lymphatic invasion | p = 0.004 * | p = 0.37 | Lymphatic invasion | ND | ND |
| negative (n = 11) | 6.3 (0.57–205) | 5.4 (1.9–20) | negative (n = 10) | 25.4 (1.74–205) | 5.91 (2.33–20) | negative (n = 1) | 0.57 | 1.92 |
| positive (n = 17) | 1.77 (0.3–4.8) | 5.2 (1.4–25.2) | positive (n = 5) | 1.43 (0.34–2.96) | 12.47 (1.38–25) | positive (n = 12) | 1.98 (0.8–4.8) | 4.16 (1.36–11.9) |
| Metastasis | p = 0.12 | p = 0.005 * | Metastasis | ND | ND | Metastasis | p = 0.75 | p = 0.034 * |
| negative (n = 24) | 2.53 (0.8–205) | 6.44 (1.6–25) | negative (n = 15) | 3.15 (0.98–205) | 8.5 (2.33–25) | negative (n = 9) | 2 (0.81–4.78) | 5.26 (1.58–11.9) |
| positive (n = 5) | 1.75 (0.3–3.4) | 1.92 (1.4–2.9) | positive (n = 1) | 0.34 | 1.38 | positive (n = 4) | 0.85 (0.57–3.43) | 2 (1.36–2.91) |
| TNM | p = 0.035 * | p = 0.44 | TNM | p = 0.02 * | p = 0.38 | TNM | p = 0.78 | p = 0.17 |
| I–II (n = 16) | 3.25 (0.8–205) | 5.91 (2.3–20) | I–II (n = 11) | 6.36 (0.98–205) | 6.41 (2.33–20) | I–II (n = 5) | 2 (0.81–3.35) | 5.26 (3.33–7.17) |
| III–IV (n = 13) | 1.75 (0.3–4.8) | 2.91 (1.4–25.2) | III–IV (n = 5) | 1.43 (0.34–2.96) | 12.47 (1.38–25) | III–IV (n = 8) | 1.85(0.57–4.78) | 2.24(1.36–11.9) |
| EPN | p = 0.94 | p = 0.22 | EPN | p = 0.86 | p = 0.34 | EPN | ND | ND |
| negative (n= 6) | 2.03 (1.3–53) | 2.49 (1.6–20) | negative (n= 4) | 4.34 (1.74–53) | 3.99 (2.33–20) | negative (n = 2) | 1.4 (1.32–1.5) | 2 (1.6–2.4) |
| positive (n= 23) | 2.17 (0.3–205) | 6.41 (1.4–25.2) | positive (n= 12) | 3.05 (0.34–205) | 8.75 (1.38–25) | positive (n = 11) | 2 (0.57–4.8) | 5 (1.36–11.9) |
| Smoking | p = 0.97 | p = 0.18 | Smoking | p = 0.68 | p = 0.28 | Smoking | p = 0.40 | p = 0.23 |
| negative (n = 12) | 2.16 (0.6–205) | 6.82 (1.9–25) | negative (n = 8) | 2.05 (1.3–205) | 10.74 (2.33–25) | negative (n = 4) | 2.67(0.57–3.43) | 4.69(1.92–7.17) |
| positive (n = 10) | 2.55 (0.3–141) | 4.93 (1.4–10) | positive (n = 7) | 3.92 (0.34–141) | 6.41 (1.38–10) | positive (n = 3) | 1.48(1.32–1.96) | 2.07(1.58–2.42) |
| All Tumors n = 29 | Intestinal Sub-Type n = 16 | Diffuse Sub- Type n = 13 | |||
|---|---|---|---|---|---|
| AhR | AhR | AhR | |||
| Gender. | p = 0.51 | Gender. | p = 0.19 | Gender. | p = 0.81 |
| Male (n = 13) | 1.54 (0.55–3.33) | Male (n = 7) | 1.71 (0.89–2.84) | Male (n = 6) | 1.51 (0.87–3.08) |
| Female (n = 16) | 1.35 (0.65–3.53) | Female (n = 9) | 1.16 (0.89–1.83) | Female (n = 7) | 1.58 (1.22–2.01) |
| Age | p = 0.82 | Age | ND | Age | p = 0.72 |
| <60 years (n = 9) | 1.94 (0.55–3.35) | <60 years (n = 1) | 1.38 (0.89–1.83) | <60 years (n = 8) | 1.51 (0.87–2.01) |
| >60 years (n = 20) | 1.80 (0.65–3.53) | >60 years (n = 15) | 1.16 (0.90–2.84) | >60 years (n = 5) | 1.58 (1.22–3.08) |
| Tumor invasion | p = 0.21 | Tumor invasion | ND | Tumor invasion | ND |
| T1–T2 (n = 6) | 1.45 (0.65–2.86) | T1–T2 (n = 4) | 0.93 (0.9–0.96) | T1–T2 (n = 2) | 1.42 (1.27–1.58) |
| T3–T4 (n = 23) | 1.94 (0.55–3.53) | T3–T4 (n = 12) | 1.55 (0.89–2.84) | T3–T4 (n = 11) | 1.53 (0.9–3.1) |
| Vascular invasion | p = 0.14 | Vascular invasion | p = 0.79 | Vascular invasion | p>0.9999 |
| negative (n = 9) | 1.25 (0.82–2.96) | negative (n = 6) | 1.21 (1.18–1.72) | negative (n = 3) | 1.53 (1.32–1.63) |
| positive (n = 20) | 2.05 (0.55–3.53) | positive (n = 10) | 1.35 (0.89–2.84) | positive (n = 10) | 1.53 (0.87–3.08) |
| Lymphatic invasion | p = 0.11 | Lymphatic invasion | ND | Lymphatic invasion | ND |
| negative (n = 11) | 1.25 (0.65–3.18) | negative (n = 10) | 1.18 | negative (n = 1) | 1.53 |
| positive (n = 17) | 2.11 (0.55–3.53) | positive (n = 5) | 1.38 (0.89–2.84) | positive (n = 12) | 1.53 (0.87–3.08) |
| Metastasis | p = 0.92 | Metastasis | ND | Metastasis | p = 0.93 |
| negative (n = 24) | 1.96 (0.55–3.53) | negative (n = 15) | 1.71 (0.89–2.84) | negative (n = 9) | 1.49 (0.87–3.08) |
| positive (n = 5) | 2.1 (0.82–2.55) | positive (n = 1) | 1.19 (1.16–1.55) | positive (n = 4) | 1.58 (1.22–2.01) |
| TNM | p = 0.30 | TNM | p = 0.80 | TNM | p = 0.21 |
| I–II (n = 16) | 1.77 (0.54–3.35) | I–II (n = 11) | 1.71 (0.89–1.83) | I–II (n = 5) | 1.32 (0.87–1.9) |
| III–IV (n = 13) | 2.11 (0.82–3.53) | III–IV (n = 5) | 1.19 (0.9–2.84) | III–IV (n = 8) | 1.6 (1.22–3.08) |
| EPN | p = 0.47 | EPN | ND | EPN | ND |
| negative (n = 6) | 1.58 (0.90–2.86) | negative (n = 4) | 0.93 (0.9–0.96) | negative (n = 2) | 1.42 (1.27–1.58) |
| positive (n = 23) | 1.94 (0.55–3.53) | positive (n = 12) | 1.55 (0.89–2.84) | positive (n = 11) | 1.53 (0.9–3.1) |
| Smoking | p = 0.85 | Smoking | p = 0.74 | Smoking | p = 0.63 |
| negative (n = 12) | 1.77 (0.55–3.35) | negative (n = 8) | 1.19 (0.89–1.71) | negative (n = 4) | 1.51 (0.87–1.63) |
| positive (n = 10) | 1.85 (0.82–3.53) | positive (n = 7) | 0.96 (0.9–1.55) | positive (n = 3) | 1.58 (1.27–2.01) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perrot-Applanat, M.; Pimpie, C.; Vacher, S.; Bieche, I.; Pocard, M.; Baud, V. Differential Expression of Genes Involved in Metabolism and Immune Response in Diffuse and Intestinal Gastric Cancers, a Pilot Ptudy. Biomedicines 2022, 10, 240. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10020240
Perrot-Applanat M, Pimpie C, Vacher S, Bieche I, Pocard M, Baud V. Differential Expression of Genes Involved in Metabolism and Immune Response in Diffuse and Intestinal Gastric Cancers, a Pilot Ptudy. Biomedicines. 2022; 10(2):240. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10020240
Chicago/Turabian StylePerrot-Applanat, Martine, Cynthia Pimpie, Sophie Vacher, Ivan Bieche, Marc Pocard, and Véronique Baud. 2022. "Differential Expression of Genes Involved in Metabolism and Immune Response in Diffuse and Intestinal Gastric Cancers, a Pilot Ptudy" Biomedicines 10, no. 2: 240. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10020240