
Current Development of Nano-Drug Delivery to Target Macrophages


1
Centre for Biomedical Technologies, School of Mechanical, Medical & Process Engineering, Queensland University of Technology (QUT), Brisbane, QLD 4000, Australia
2
State Key Laboratory Breeding Base of Basic Science of Stomatology (Hubei MOST) and Key Laboratory of Oral Biomedicine Ministry of Education, School and Hospital of Stomatology, Wuhan University, Wuhan 430079, China
3
Australia-China Centre for Tissue Engineering and Regenerative Medicine, Queensland University of Technology, 60 Musk Ave., Kelvin Grove, Brisbane, QLD 4059, Australia
*
Authors to whom correspondence should be addressed.
Biomedicines 2022, 10(5), 1203; https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10051203
Submission received: 13 April 2022
/
Revised: 16 May 2022
/
Accepted: 18 May 2022
/
Published: 23 May 2022
(This article belongs to the Special Issue Advances in Nanomaterials for Drug Delivery)
{kind=link}
{kind=link}
Abstract
:Macrophages are the most important innate immune cells that participate in various inflammation-related diseases. Therefore, macrophage-related pathological processes are essential targets in the diagnosis and treatment of diseases. Since nanoparticles (NPs) can be preferentially taken up by macrophages, NPs have attracted most attention for specific macrophage-targeting. In this review, the interactions between NPs and the immune system are introduced to help understand the pharmacokinetics and biodistribution of NPs in immune cells. The current design and strategy of NPs modification for specific macrophage-targeting are investigated and summarized.
1. Introduction
Nanoparticles (NPs) have been extensively used for drug delivery in disease treatment, taking advantage of their stability, biocompatibility, blood circulation, immunogenicity, and capability to control drug release. Due to the nature of phagocytes, NPs can be preferably taken up by phagocytes in vivo, facilitating phagocyte-targeting drug delivery without influencing the function of other cells, which has become a new direction of drug delivery. Phagocytes, such as macrophages, are the most significant innate immune cells, which participate in the pathological processes of various inflammation-related diseases, making macrophages essential targets for developing novel diagnostic imaging and disease treatment. Therefore, increasing studies have used NPs for macrophage-targeting drug delivery. In this review, we introduced the current strategies for NPs modification for specific macrophage-targeting and their applications in inflammatory diseases to provide foundations for developing/optimizing macrophage-targeting NPs in the future.
2. The Physiology and Pathology Function of Macrophage
2.1. The Role of Macrophages in Immune Response
Macrophages, which are a class of innate immune cells that originated from the mononuclear phagocyte system (MPS), play an essential role in the inflammation and regeneration of injured tissues [1,2]. Macrophages can be derived from circulating monocytes, or exist in the body as resident cells, residing mainly in the liver (Kupffer cells), lungs (alveolar macrophages), spleen, lymph nodes, thymus, gut, marrow, brain, connective tissue, and serous cavities [3,4]. In acute inflammation, the released chemokines and cytokines induce the recruitment of monocytes which then differentiate into macrophages. As a class of phagocytic cells, macrophages mainly function in the removal of pathogens, cell debris, apoptotic cells, and small particles in immune responses [2]. Macrophages also act as antigen-presenting cells, which present antigens to T cells to stimulate the adaptive immune response [2].
Based on different functions, characteristics, surface markers, and inducers, macrophages can be broadly divided into M1 (including M1a and M1b sub-populations) and M2 phenotypes (including M2a, M2b, and M2c sub-populations) [5]. The M1 activation induced by lipopolysaccharide (LPS) or cytokines such as interferon-gamma (IFN-γ) and tumor necrosis factor-alpha (TNF-α), is characterized by high antigen presentation capacity, the killing of intracellular pathogens, secretion of pro-inflammatory cytokines and anti-tumor effects. The M2 polarization induced by interleukin-4 (IL-4) and IL-13, mainly results in the resolution of inflammation and repair of injured tissue. M1 phenotype induces the expression of inflammatory cytokines such as IL-1β, IL-6, nitric oxide (NO), and inducible nitric oxide synthase (iNOS), resulting in enhanced inflammation. In contrast, the M2 phenotype is characterized by an increase in the IL-10 cytokine and a decrease in the production of iNOS, leading to limitation of inflammation and promotion of tissue repair [6,7].
2.2. Macrophages in Diseases with Inflammation
2.2.1. Infection
Since macrophages are prominent in inflammation progress, they are closely related to the pathology of many inflammation-related diseases. In infectious diseases, macrophages contribute to the defense against pathogens and maintenance of tissue homeostasis. Pathogens can be detected by macrophages by combining with the specific receptors on the cell surface, such as Toll-like receptors (TLR) or other pathogen recognition receptors, leading to activation of downstream signaling pathways and enhanced release of cytokines to alarm other cells about bacterial invasion. Macrophages also engulf the alien microorganisms and even infected cells, degrade antigens in the early stage of an infectious process. Then, the activated macrophages will function as antigen-presenting cells and translocate degraded antigens to cell surfaces. CD4+ T cells can further recognize those antigens by T cell receptors (TCR) and be activated to initiate Th1 and Th2 immune responses. Activated macrophages can also release cytokines that can influence the adaptive immune response [1]. Macrophages are generally activated towards a M1 phenotype in the early stage of the infection, leading to the release of a large amount of pro-inflammatory factors (TNF-α, IL-1β, NO, etc.), thus killing the invading organisms and activating the adaptive immunity (through antigen-presenting). To avoid the excessive inflammatory response, macrophages undergo apoptosis or polarize to the anti-inflammatory M2 phenotype to protect the host from excessive injury and facilitate tissue healing.
In brief, the M1 phenotype dominates in infection and is strongly microbicidal, exhibiting high antigen presentation ability [8,9]. In contrast, M2 phenotype is less microbicidal and is associated with parasite infestation or chronic infection. It is worth noting that some bacteria can evade macrophage clearance through some special mechanisms. After being engulfed, the bacteria are supposed to either escape from the phagosome, or restrain the fusion of phagosome and lysosome, or survive inside phagolysosomes [10,11,12]. For example, Listeria monocytogenes can escape from phagosome [13]; bacteria from Shigella genus voids formation of autophagosome [14], Mycobaterium tuberculosis impedes phagosome maturation [15]; Legionella pneumophila modifies the phagosome [16]; and Coxiella burnetti replicates inside phagolysosomes [17].
2.2.2. Bone Healing
Macrophages have also received the most attention in bone healing than other immune cells due to their vital role in osteogenesis and osteoclastogenesis. It has been reported that depletion of resident macrophages leads to the failure of osteoblast-driven bone formation in vivo, indicating that resident macrophages are crucial for osteoblast differentiation and mineralization [18]. Macrophages are also the precursor of osteoclasts. Under the stimulation of macrophage-colony stimulating factor (M-CSF) and receptor activator of nuclear factor-κB (NF-κB) ligand (RANKL), macrophages can differentiate into osteoclasts and modulate the formation of new bone. It is still controversial about which macrophage phenotype is the most beneficial for osteogenesis. Traditionally, the pro-inflammatory M1 macrophages are regarded to induce osteoclastic activities by secreting various pro-inflammatory cytokines, resulting in enhanced bone resorption. On the contrary, activated M2 macrophages tend to be more anti-inflammatory and closely associated with late-stage of tissue repair, resulting in the formation of new bone. However, some recent studies have found that M1 macrophages are also capable of enhancing osteogenesis [19,20]. Moreover, premature of M2 macrophages and excessive M2 polarization lead to formation of scar tissue or delayed wound healing [21,22]. Therefore, it is assumed that both M1 and M2 macrophages play indispensable roles in osteogenesis and a timely switch of M1 to M2 phenotype is the key factor in bone healing. The pro-inflammatory M1 macrophages play a more prominent role in the early stage of the repair response, while the M2 macrophages dominate the mid to late stages of the repair response. The M1 polarization in the early stage of bone healing determines the cytokine release pattern of M2 macrophages. Prolonged M1 polarization can lead to an increase in fibrosis-inducing cytokine release patterns (such as TNF-α, tissue growth factor-β1 (TGF-β1), TGF-β3), which results in the formation of a fibrocapsule. By contrast, an effective and timely M1-to-M2 macrophage phenotype switch can result in an osteogenesis-favoring cytokine release pattern (such as bone morphogenetic protein (BMP) and vascular endothelial growth factor (VEGF)) to facilitate the formation of new bone tissue [23].
2.2.3. Cancer
Cancer refers to the rapid, abnormal, and uncontrolled proliferation of local tissue cells. It is commonly believed that tumor-associated macrophages (TAMs) assist in the angiogenesis, metastases, and immunosuppression of tumors, thus playing a prominent role in tumor survival [24,25]. TAMs express low levels of the polarization-associated macrophage antigens, carboxypeptidase M, TNF-α, and CD51, but show high expression of IL-1 and IL-6. Although TAMs can also be broadly divided into M1 and M2 phenotypes [26,27], it is believed that most resident TAMs exhibit a M2-like phenotype induced by chemokines, tumor-derived cytokines, and proteases, such as IL-4, IL-6, IL-10, TGF-β1, and polyethylene glycol (PEG) [28,29]. M2 macrophages promote tumor angiogenesis by secreting matrix metalloproteinases (MMPs) to release matrix-sequestered VEGF and by producing dozens of angiogenic factors that facilitate endothelial survival and proliferation. Enhanced angiogenesis further promotes tumor growth which is associated with faster cancer progression and shorter patient survival [4,30,31,32,33]. M2 macrophages also secrete anti-inflammatory factors, such as arginase, TGF-β, and IL-10 [34,35] to induce immunosuppression [36]. In contrast, M1 macrophages increase antigen presentation and enhance tumor cell clearance by secreting pro-inflammatory cytokines, including IL-6, IL-12, and TNF-α. Therefore, targeting and repolarizing TAMs from M2 to M1 phenotype can be a potential strategy to prevent tumor neovascularization and growth, thus being a promising therapeutic approach against cancer [37,38,39].
2.2.4. Atherosclerotic Cardiovascular Disease
Atherosclerotic cardiovascular disease is a chronic inflammatory disease, which causes morbidity and mortality worldwide [40]. During the pathological process of atherosclerotic cardiovascular disease, macrophages play a prominent role. Monocytes are recruited into the subendothelial space of the arterial intima by endothelial cells [41,42] and then differentiate into inflammatory macrophages, which phagocytose lipoproteins, leading to the formation of lipid-rich foam cells and early atherosclerotic lesions, resulting in the amplification of the local inflammatory response [43]. The atherosclerotic lesions progress to an advanced stage if the pro-inflammatory status persists, characterized by accumulated apoptotic macrophages and failure of their clearance [44,45,46]. Depending on local inducers in the micro-environment, macrophages can either polarize into a pro-inflammatory M1 phenotype or a reparative M2 phenotype [47]. Pro-inflammatory M1 macrophages are known to diminish lesion stability by inhibiting collagen production from smooth muscle cells and producing MMPs that degrade the protective fibrous capsule [48,49,50,51]. By contrast, pro-resolving M2 macrophages promote lesion stability by clearing apoptotic cells through efferocytosis, thereby diminishing plaque necrosis [52]. Therefore, macrophages are important targets in the treatment of atherosclerotic cardiovascular disease, considering their key role in the progression and regression of atherosclerotic lesions. Moreover, the so-called vulnerable atherosclerotic plaques lead to fatal sequelae, such as acute myocardial infarction and stroke, causing death worldwide. Given that atherogenesis processes slowly and sometimes over decades, early diagnosis and interventional therapy against high-risk atherosclerotic plaques before clinical symptoms are of great importance [53], and macrophages have been regarded as ideal targets of diagnostic imaging and treatment for atherosclerosis [54,55,56,57].
2.2.5. Ageing
Ageing populations are becoming a global trend, characterized by the accumulation of cells which have undergone the process of permanent cell cycle arrest, termed senescence [58,59]. In the immune system, cellular senescence has been observed in multiple cell types, including macrophages and T cells [60,61,62]. Macrophage senescence has been found to contribute to ageing, evidenced by the fact that in ageing individuals, multiple organs are under a long-term, chronic, and sustainable state of inflammation, also known as inflammageing [63]. Although macrophages are not the sole source of inflammageing, extensive studies have shown that macrophages are the central component in initiating ageing-related chronic inflammation. It has been found that the M1 macrophage level in ageing mice is higher than that in young mice, which is probably related to deficient tissue repair capacity and long-term inflammation state in ageing mice [64]. Meanwhile, the regenerative M2 macrophage population decreases with age, symbolized by the reduced expression of IL-10 (a M2 polarization marker) in ageing mice [65], and delaying macrophage senescence by knocking out tumor suppressor gene p16 (INK4a), which is associated with an increased risk of age-related inflammatory diseases, can upregulate the expression of Arg-1 (a M2 polarization marker) [66]. In addition to phenotype switch, the total macrophage population also changes with ageing. Studies have compared bone marrow in teenagers (<19 years of age) with adults, and found the former contains significantly more macrophages compared to the latter [67]. These macrophages have been further analyzed, and researchers have identified a higher population of CX3CR1+ macrophages and a reduction in Ly6C+ macrophages in old mice compared to young [68], suggesting that macrophages changed towards a more pro-angiogenic phenotype in old mice compared with young mice [69]. In the lungs of old mice, studies found the macrophage numbers declined, with the expression of over 3000 genes being altered, as compared to lungs of young mice [70]. Among these genes, the most affected ones include scavenging receptors, such as CD204 [70], and the macrophage receptor with collagenous domain (MARCO) [71], which impacts bacterial phagocytosis and efferocytosis of apoptotic neutrophils, adversely affecting inflammatory responses.
Macrophages also contribute to the pathology of age-related diseases, such as polymyalgia rheumatic (PMR) and giant cell arteritis (GCA), which only occur in people over 50 years old and are both characterized by increasing production of IL-1β and IL-6 by arterial macrophages [68]. Likewise, chronic obstructive pulmonary disease (COPD) is also strongly linked to inflammageing [72,73,74], due to the impaired bacterial phagocytosis and efferocytosis in alveolar macrophages, resulting in increased bacterial colonization and an elevated pro-inflammatory environment [75]. Therefore, it is believed that inflammation caused by aged macrophages has been potential target to reverse the detrimental effects of inflammageing and boost immunity in old individuals.
Considering the prominent role of macrophages in the pathological progression of various diseases, they are regarded as essential targets in modulating inflammation, thus developing new methods for disease treatment. However, specifically targeting macrophages involved in conditions without interfering with host immunity remains a challenge [76]. Given the phagocytic nature of macrophages in the clearance of cellular debris, apoptotic cells, pathogens, and especially foreign particles in immune response, NPshave attracted much attention for specific macrophage-targeting due to their feature to be preferentially engulfed by macrophages [2].
3. Nanotechnology-Based Drug Delivery System
NPs, with diameters ranging from 1 to 500 nm, have high loading capacities for drugs owing to their large surface-area-to-volume ratios. The small size of NPs allows them to be largely endocytosed by cells, thus increasing their accumulation in phagocytes [2]. Their small size also increases the tumor-targeting ability of NPs through the enhanced capillary permeability and retention effect as tumor tissues lack lymphatic drainage compared with normal tissues, which enables NPs to be taken up more efficiently by tumors [77]. NPs can be synthesized into various shapes and sizes based on different preparation methods with different raw materials (such as polymers, lipids, ceramics, and metals). Because of their tunable physiochemical properties and enhanced carrying capacity, NPs have been used for various biomedical applications [78,79,80], including diagnostic imaging and drug delivery. It is well known that drug-loaded nanosystems have many advantages compared with traditional drug therapy in terms of stability, biocompatibility, controlled drug release, and immunogenicity. Studies have shown that the encapsulation of therapeutics (such as small-molecule drugs, peptides, proteins, small interfering RNA (siRNA), and microRNA (miRNA)) into NPs prevents extensive metabolism and prolongs circulation time, improving the chemical stability and pharmacokinetics of the loaded therapeutics [81,82]. Moreover, NPs are known to interact with immune components in blood, such as opsonin, which causes NPs to be more easily recognized and preferentially taken up by macrophages [83,84], thus reducing the off-target and side effects on other cells and tissues compared with free drugs [85,86,87]. Until now, NP-based delivery strategies have shown great promise in suppressing specific pathological processes, including inflammation in mouse models [81].
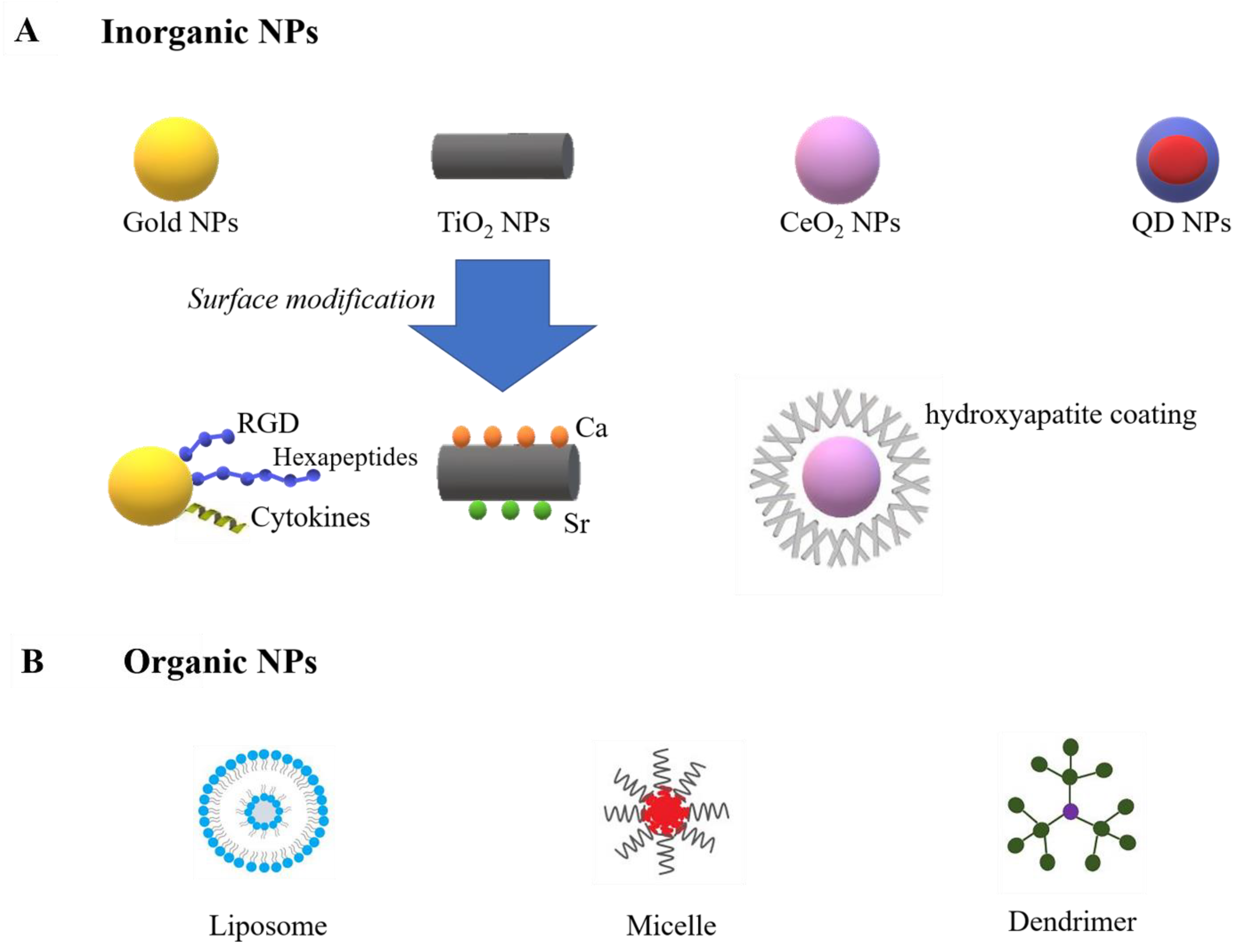
NPs can be assigned into two major groups: inorganic (e.g., gold, titanium oxide, cerium oxide, iron oxide, and quantum dot (QD) NPs) and organic (e.g., liposomes, micelles, protein-based nanomaterials, and dendrimers) (Figure 1).
Inorganic NPs are normally much smaller, contain metallic elements, and exhibit physical properties of specific nanoscale [88]. For example, gold NPs are extensively studied in cardiac tissue regeneration due to their capability to enhance scaffold conductivity [89]. Moreover, recent research reported their ability for immunomodulation by restraining inflammatory cytokines and enhancing anti-inflammatory cytokines [90]. The surface chemistry of gold NPs can be changed by bioactive molecules, allowing them to modulate macrophage polarization. Studies have decorated gold NPs with peptide arginine-glycine-aspartic acid (RGD) [91], hexapeptides Cys-Leu-Pro-Phe-Phe-Asp [92], and IL-4 [93], and those functioned gold NPs increased M2 macrophage polarization and decreased M1 phenotype. Titanium oxide NPs (TiO2 NPs), which are largely used in tissue engineering due to their ability to resist corrosion from the body, were also reported to accelerate M1 to M2 transition [94]. Research by Lee has reported an increased M2 macrophage population on calcium (Ca) and strontium (Sr)-modified titanium implants, leading to enhanced osteogenesis [95]. Moreover, it has been reported that TiO2 nanotubes can induce anti-inflammatory phenotype and promote endothelialization [96]. Cerium oxide NPs (CeO2 NPs) are also bioactive NPs with strong antioxidant properties. CeO2 NPs have been reported to enhance wound healing by enhancing fibroblast and vascular endothelial cells [97]. Moreover, by coating hydroxyapatite on the surface of CeO2 NPs, an increased M2 polarization can be achieved [98]. The Ce4+/Ce3+ ratio can influence the osteogenic ability of CeO2. The higher ratio enhanced M2 macrophages polarization and achieved better osseointegration in vivo [99]. Despite immunomodulation, inorganic NPs are also applied as imaging agents because of their superparamagnetic properties [100,101]. For example, both gold, iron oxide and QD NPs can be used in magnetic resonance imaging (MRI). Since the optical and electronic characteristics of QD NPs directly depend on the particle size, QD NPs can be applied in various imaging techniques other than MRI, such as X-ray computed tomography (CT), positron emission tomography (PET), and single-photon emission computed tomography (SPECT), by simply changing the QD NPs size [102,103,104,105].
Organic nanocarriers are carbon-based and are generally characterized by their biocompatibility and improved drug-loading capacity. Several major groups of organic nanocarriers include liposomes, micelles, dendrimers, and protein/peptide-based nanocarriers [88]. Liposomes are large NPs (25–1000 nm) that have been shown to accumulate in macrophages (by inducing phagocytosis), achieving macrophage-targeting in a simple and effective way [106]. However, due to their enhanced macrophage uptake in the mononuclear phagocyte system (MPS), liposomes can be rapidly cleared from circulation. Some studies coated PEG on the surface of liposomes and reduced their clearance by the MPS [107]. Micelles are natural nanocarriers with a large water-free environment in their core, which is surrounded by a hydrophilic surface. Micelles can be functioned by different bioactive molecules, such as CpG oligonucleotides and N-methylpyrrolidone. CpG oligonucleotides enable micelles to increase the CD8+ and CD4+ T cell responses, and N-methylpyrrolidone reduces the accumulation of micelles in the reticuloendothelial system (RES) [108,109]. Dendrimers are tree-like nanostructures, which have attracted significant attention as nanocarriers for small-molecular drugs, proteins, and nucleic acids. The subunits of dendrimers grow around the core in concentric layers to produce stepwise increases in size that are similar to globular proteins. This property is beneficial to NP–drug interaction [110].
4. Interaction between NPs and Immune System
The clearance and biodistribution of NPs are mediated by circulating and tissue-resident phagocytes of the MPS, including macrophages, which engulf and accumulate NPs in tissues [111,112]. This interaction between NPs and the immune cells can either result in immunosuppression or in immunostimulation, which may enhance or reduce the therapeutic effects of NPs [84,113]. Therefore, understanding the mechanisms involved in NP-uptake and the interaction between NP and MPS can facilitate the development of advanced NPs with features, such as prolonged systemic circulation time, enhanced selective delivery to the targeted tissues, as well as improved clinical implication.
Once entered the blood circulation system, plasma proteins will quickly adhere to the surface of NPs to form a “corona”, which contains about 3700 proteins [114]. Among these proteins, the adsorption of opsonin on the NP surfaces enables NPs to be more engulfed by phagocytes in the RES system, especially by macrophages [115]. The NP-uptake in macrophages is either non-selective or selective. The nonselective mechanisms include macropinocytosis and phagocytosis. Macropinocytosis refers to the uptake of tiny, soluble substances or extracellular fluids, while phagocytosis involves the uptake of larger, solid particles [116]. Previously mentioned opsonin can enhance phagocytosis by binding to macrophage surface receptors once adsorbed on the NP surface to trigger phagocytosis. Local actin will then be rearranged, contributing to the formation of a new vesicle called “phagosome” once tightly engulfed large NPs or NP aggregates. The fusion of phagosome with lysosome further forms a “phagolysosome”, which will eventually be degraded [117]. Unlike nonselective uptake, selective uptake requires the interaction and combination of cell membrane receptors with ligands decorated on NP surface [118,119,120]. The most common form of selective uptake is clathrin-mediated endocytosis, which strongly depends on NP size and only occurs in NPs with a diameter less than 50 nm [121,122]. As a coating protein presents in all cells, clathrin can be activated by the binding of ligands on the NPs to the AP-2 adaptor complex on the cell membrane, and then move to the binding site where a transport vesicle is formed. The vesicle will be coated with clathrin to travel through the cell. After vesicles are fused with lysosomes, the clathrin will fall off and return to the cell surface to repeat the process [123].
Many characteristics can influence the interaction between NPs and macrophages. For example, greater size may increase the extent of opsonization and enhance NP-uptake, and smaller size NPs (<100 nm) are more preferable due to slower removal from blood [124,125]. In addition to NP size, surface properties, such as hydrophilicity, and surface charge can also impact the adsorption of opsonin, thus influencing the uptake of NPs by phagocytes and clearance from the bloodstream [126,127]. Increased hydrophilicity results in a lower degree of protein adsorption and reduced uptake by macrophages [128]. Tabata et al. have tested the uptake of hydrophilic and hydrophobic NPs by macrophages and showed that the latter were more largely engulfed [129]. Many researchers coated NPs with PEG to reduce their uptake by macrophages and prolong the circulation lifetime. PEG is a hydrophilic polymer that increases the hydrophilicity of NPs and provides a steric hindrance via water shell formation, thus preventing the opsonin from binding to the NPs [128,130]. Purified cellular membranes from leukocytes and platelets are also used to avoid opsonization and delay the uptake of NPs by MPS [131,132]. Surface charge is another factor that influences phagocytosis. Positively or negatively charged surfaces show increased phagocytosis, as compared to neutral particles [120,133], which is resulted from the smaller charge magnitude of neutral particles [133]. Smaller aspect ratios also allow NPs to be phagocytosed more easily by macrophages compared with NPs with larger aspect ratios, due to the fact that smaller aspect ratios require less actin remodeling and less energy in the phagocytic process [134].
The uptake of NPs by phagocytes also has toxicological effects on immune cells [82], which might be contributed by the particle chemistry of NPs, according to some studies [135,136,137]. For example, the surface charge of silicon (Si) NPs and the size of zinc oxide (ZnO) NPs affected the interaction with cells and intracellular reactive oxygen species (ROS) production, thus influencing cytotoxicity [136,137]. Compared with T and B lymphocytes, macrophages play a more critical role in the biodistribution of NPs, which may be more sensitive to NP toxicity [138]. This indicates that NP-uptake by macrophages and intracellular release of the loaded drugs may contribute to the NP-mediated toxic effect on the MPS cells.
5. Macrophage-Targeting Nanotechnologies
5.1. Passive Macrophage-Targeting Nanotechnologies
There are two specific-targeting approaches in the design of NP-based drug delivery systems, namely passive and active targeting (Figure 2).
Passive targeting takes advantage of the NP pharmacokinetics, unique vascular pathophysiology, and immune responses of the targeted tissue, leading to the accumulation of NPs [139,140]. For example, large NPs (up to 500 nm) predominantly accumulate in the liver and lungs; medium NPs (10–300 nm) aggregate mainly in the liver and spleen after being opsonized and removed from the circulation; and small NPs (1–20 nm) are usually degraded by macrophages in the kidneys [139,141,142]. Therefore, the preference for unmodified NPs to accumulate in certain tissues has been utilized as a passive-targeting approach to deliver the payload to the macrophages. Inflammatory tissues and solid tumors are characterized by vascular leakage contributed by inflammatory mediators, cytokines, and growth factors that cause disruption of the endothelium [143,144,145] and, consequently, result in NP in situ accumulation. For example, Corvo et al. [146] intravenously injected liposomes coated with PEG into a mice model of rheumatoid arthritis and observed passive accumulation of these NPs at the arthritic sites. Thus, NPs with proper size can preferentially extravasate from the blood into the interstitial spaces and accumulate in inflammation sites or tumor tissues via the enhanced permeability and retention (EPR) effect [147]. Meanwhile, Keliher et al. [148] reported that TAMs are able to selectively capture NPs and translocate them from the periphery to the central hypoxic zone of tumor tissue. The ability of TAMs to take up NPs has been utilized in tumor imaging and quantification of TAMs in which zirconium (Zr)-labeled dextran-based NPs were applied as MRI imaging agents [142]. Since elevated infiltration of macrophages and lymphocytes are common in the extravascular spaces of inflammation sites and tumors (inducing excessive inflammatory responses and tissue damage) [149,150,151], macrophages are considered the primary therapeutic targets in inflammatory diseases [152,153], and particle uptake into macrophages could, therefore, allow the selective accumulation of NPs in the areas of inflammation [154,155].
5.2. Active Macrophage-Targeting Nanotechnologies
5.2.1. Phagocytosis-Related Cell Membrane Receptors Targeting
Active targeting can be used to enhance the selective delivery of drugs into the target cells or tissues by exploiting the specific interactions between drug carriers and targeted sites [139,156,157,158]. Currently, the most common approach to achieve active-targeting is to decorate the surface of NPs with an agent (e.g., ligand, antibody, and peptide) that can selectively interact and be recognized by the particular cell type in certain tissue [159]. Different receptors and lipid components on the cell membrane, thus playing a prominent role in active-targeting, for they can recognize specific agents on NPs. Moreover, compared with normal cells, pathological cells may uniquely express these receptors or express them at a different amount. Depending on the target cell and/or tissue types, various ligands, such as monoclonal antibodies, could be harnessed to the particle surface [160]. Thus, the active-targeting approach of NPs is based on the concept that the surface of the delivery vehicle is modified with a ligand or antigen to allow selective interaction with the target receptors [161]. The macrophage cell membrane surfaces contain many receptors that determine the activity of macrophages, including growth, differentiation, activation, recognition, endocytosis, migration, and secretion [162]. Among them, three main groups are frequently assumed: TLR, non-TLR, and opsonic receptors. Although TLRs do not participate in phagocytosis or endocytosis, they are involved in antimicrobial peptide production and innate immunity, playing a central role in recognition of pathogens and activation of innate immune responses [163,164,165]. Non-TLR, including the family of scavenger receptors [166] and the C-type lectins [167], are involved in phagocytosis and endocytosis. Opsonic receptors include complement receptors (integrins, such as CD11b) and Fc receptors (immunoglobulin superfamily, such as CD44), which dominate the phagocytosis and endocytosis of complement- or antibody-opsonised particles, respectively [168]. Those phagocytosis-related receptors are, thus, ideal structures for a macrophage-targeting therapy, which can facilitate nano-carriers to deliver therapeutic agents into macrophages selectively. As for non-TLR, for example, C-type lectin receptors (CLRs) recognize conserved carbohydrate structures, including mannose and galactose. Mannosylated liposomes have repeatedly been shown to preferentially target macrophages, enhancing cellular uptake both in vitro and in vivo [169,170,171,172]. Likewise, Lipinski et al. have decorated antibodies that specially interact with scavenger receptor (CD36, which is highly expressed on the surface of macrophages) on the surface of micelles for macrophage-targeting imaging in atherosclerosis [173]. Hyaluronic acid (HA), which is an unbranched non-sulphated glycosaminoglycan consisting of repeating disaccharides (β-1,3-N-acetyl-D-glucosamine and β-1,4-D-glucuronic acid), can be recognized by a large number of HA receptors on macrophages, such as Fc receptor (CD44), the receptor for HA-mediated motility (RHAMM), and several other receptors possessing HA-binding motifs [174]. Kamat et al. found that iron oxide magnetic NPs coated HA could be efficiently entrapped by activated THP-1 macrophages in vivo [175]. Tsai et al. reported a HA conjugated gold nanorod (HA-Au NR) as a drug carrier for anticancer doxorubicin (Dox) delivery, and HA significantly improved the recognition and uptake of HA/Dox-Au NRs by RAW 264.7 cells through the specific interaction with the HA receptor, CD44 on the cell surface [176]. Additionally, the modification effectively improved biocompatibility by switching the surface charge from positive (due to chitosan polymer) to negative (due to hyaluronate) [1]. To reduce the immunogenicity associated with the Fc portion, Gagne et al. coupled Fab’ fragment of the anti-HLA-DR (anti-class II major histocompatibility complex molecules) antibody rather than the entire antibody with PEG-modified immunoliposomes. Macrophages express a high level of HLA-DR, and the result showed a much more increased accumulation of modified NPs in lymphoid tissues compared with free drugs [177].
5.2.2. Pathogen Components-Mediated Macrophage-Targeting
In addition to components targeting phagocytosis-related cell membrane receptors, another strategy is to use pathogen-associated components to induce active-targeting, which takes advantage of the macrophage’s nature to engulf pathogens. Chavez-Santoscoy et al. decorated galactose and di-mannose on the surface of polyanhydride NPs to provide pathogen-like properties to target CLRs on alveolar macrophages [178]. Mannosylated fluorescent phenylboronic acid-containing NPs were also selectively taken up by murine RAW 264.7 macrophages, which were used as cell imaging agents [179]. More importantly, according to some in vivo studies, mannose significantly increased the uptake of gelatin NPs by macrophages in the liver, lymph nodes, and lung, compared with pure gelatin NPs [1]. Dextran sulphate, derived from certain lactic acid bacteria, is constituted by a linear glucose chain with one sulphate group per glucose unit [180]. Dextran is reported to be recognized by scavenger receptor class A on macrophages [181,182,183,184], which is, therefore, frequently used as decoration on NPs to enhance the specificity for targeting macrophages [148,185]. Keliher et al. has introduced a crosslinked, short-chain dextran nanoparticle that accumulated primarily in tissue-resident macrophages. Labeled with 89Zr, this NP can be used as a macrophage-specific PET imaging agent to quantify macrophage inflammation levels in the diagnosis of various diseases, such as cancer, atherosclerosis, and myocardial infarction [148].
Despite of ligands, Ren et al. encapsuled polymer−lipid hybrid NPs into porous and hollow yeast cell wall for macrophage-targeting drug delivery. The yeast cell wall composed of natural β-1,3-D glucan, can be recognized by the apical membrane receptor, dectin-1, which has a high expression on macrophages and intestinal M cells [186]. Soto et al. also incorporated NPs as cores inside glucan particles (GPs), which are hollow, porous 2–4 μm microspheres derived from the cell walls of Baker’s yeast (Saccharomyces cerevisiae), taking advantage of the macrophage-targeting property of GPs. The 1,3-β-glucan outer shell provides for receptor-mediated uptake by phagocytic cells expressing β-glucan receptors [187]. Except for the cell wall, Gram-negative bacteria outer membrane vehicles (OMVs), which exhibit various pathogen-associated molecular patterns and immunogenic antigens [188,189] that can be recognized by macrophages, have also been used for macrophage-targeting immunomodulation [190,191]. Gao et al. compared nanoparticles coated with the membrane of OMVs from Staphyloccocus. aureus (S. aureus) with counterparts coated with PEGylated lipid bilayer, and OMV membrane coating was found to facilitate NP internalization by S. aureus-infected macrophages [192].
Despite membrane coating, bio-nanocapsules (BNCs) derived from pathogens could be directly used as drug carriers. For example, hepatitis B virus (HBV) envelope particles, which are 50 nm BNCs consisting of approximately 110 molecules of HBV surface antigen L protein and lipid bilayer, have been explored to specifically deliver payloads to liver cells [193]. Li et al. then developed mutated BNC to selectively target the non-hepatic cells and tissues in vitro and in vivo, relying on the outwardly displayed tandem form of the S. aureus protein A-derived Z domain which could bind to animal IgG Fc domains [194,195,196]. This protein A-derived Z domain was recently replaced by Finegoldia magna protein L-derived B1 domain and showed enhanced uptake by the murine macrophage cell line RAW 264.7 [197].
5.2.3. Other Chemical Compounds-Mediated Macrophage-Targeting
The folate receptor (FR) is over-expressed on the activated macrophage surface in rheumatoid arthritis [198,199]. Thomas et al. used folate, which can be recognized by FR on macrophages, to decorate NPs loaded with methotrexate for macrophages-targeting. These NPs were demonstrated to offer a practical clinical approach to improving the drug delivery and efficacy at the inflammatory site of arthritis [200]. Similarly, another study fabricated FR-targeting fluorescence nanoprobes to detect and quantify the extent of biomaterial-mediated inflammatory responses in vivo. They found a good relationship between the extent of the inflammation and the intensity of nanoprobe-associated fluorescence signal in tissue [201]. Except for arthritis, ovarian TAMs also express a high level of folate receptor-2 (FOLR2), which can be selectively targeted using G5-methotrexate (act as both a ligand and a toxin) dendrimer NPs for cancer treatment [202].
Phosphatidylserine (PhoS) has significant potential to selectively target macrophages and has been frequently applied in developing drug delivery systems by anchoring PhoS on NPs [200,201,202,203]. Normal cells have PhoS inside the phospholipid bilayered plasma membrane, whereas apoptotic cells bring out PhoS to the outer surface of the plasma membrane and make themselves recognized by macrophages for phagocytosis. Thus, the expression of PhoS on apoptotic cells allows for PhoS-dependent identification and engulfment by macrophages [203].
Brain-derived neurotrophic factor (BDNF), which is a protein belonging to neurotrophins that support neuronal cells’ growth, differentiation, and survival, can bind to two neurotrophin receptors on the cell surface: the low-affinity neurotrophin receptor p75 and the high-affinity receptor TrkB [204,205]. In the research by Talvitie, chitosan NPs were functioned by TrkB binding targeting peptides and showed more efficient binding to RAW 264.7 macrophages than pure NPs [206].
Heparin, which is traditionally regarded as an anticoagulant, is also reported to have macrophage binding affinity and can further act as a macrophage-targeting agent [207]. Its uptake by macrophages is reported to be mediated by scavenger-like receptors. Interestingly, heparin-loading reduced the toxicity of cationic NPs in the rat macrophage NR8383 cell line [208,209].
5.2.4. Strategy to Specifically Target M1 or M2 Macrophage
As previously mentioned, macrophages in inflammation sites polarize into two phenotypes depending on local stimulations. A prolonged activation or dysregulation of M1 activity is closely related to the development of chronic diseases, such as rheumatoid arthritis, delayed/non-healing wounds, psoriasis, and septic shock, which can lead to multiple organ dysfunction syndromes (MODS). Therefore, selective M1 macrophage-targeting immunomodulation has become the focus of treatment, which can avoid the side effects on other cells [210]. However, it is challenging to find a suitable surface molecule specifically expressed or upregulated by M1 macrophages. This is because macrophages are highly plastic cells, exhibiting different surface markers depending on the inducers from the different microenvironments [27]. Recently, M1-specific and M2-specific surface marker screening have been performed, using mice and human macrophages exposed to their respective inflammatory stimulus (M1: IFN-γ and LPS, M2: IL-4) [211]. According to this study, the expression of two receptors (CD64, CD14) was increased in both mice and human M1 macrophages but reduced in the M2 macrophages. On the contrary, mannose receptor (CD206) and macrophage galactose-type C-type lectin (CD301) were down-regulated on M1 macrophages but upregulated on the M2 macrophages in both mice and human M1 macrophages [211]. Among them, attention has been paid to Fc γ RI (commonly referred to as CD64), which is considered a suitable target molecule on M1 macrophages owing to their ability to bind and rapidly internalize monomeric IgG [212]. The development of antibodies against CD64, such as monoclonal antibody (mAb) 197, can specifically recognize and bind to monocytes, allowing for the development of macrophage-targeting technologies [213]. mAb 197 was used for the clinical treatment of chronic immune thrombocytopenia purpura (cITP), as mAb 197 binding prevented CD64 mediated destruction of IgG-coated platelets [214]. The murine-derived mAb 197 was further humanized (H22), which contained both binding specificity and high affinity for CD64, to reduce immunogenic response in human body [215,216].
TAMs, which are normally M2-like macrophages, facilitate tumor angiogenesis and growth, thus playing an indispensable role in tumor development and progression. Therefore, specific M2 TAMs have been considered as therapeutic targets in tumor treatment [217]. Recently, a M2 macrophage-binding peptide (M2pep) identified by phage display was reported to have high selectivity and efficient targeting ability to M2 macrophages [218,219]. Research has functioned gold NPs with M2pep to deliver siRNA in a lung cancer mouse model to achieve specific and long-lasting gene therapy in inflammatory TAMs [218,219]. Other cancer treatments, such as NPs-based magnetic hyperthermia therapy (MHT), were also performed. By coating iron oxide NP with M2pep in an orthotopic breast cancer mouse model, the M2 TAM-targeting MHT significantly reduced the tumor volume by reducing the population of pro-tumoral M2 TAMs in tumor [217]. Except for M2pep, another α-peptide (a scavenger receptor B type 1 targeting peptide), also possesses great specificity to M2-like TAMs [219,220]. Han et al. developed poly (lactic-co-glycolic acid) (PLGA) NPs conjugated both M2pep and α-pep to target M2 TAMs. This successfully transformed M2 to M1 phenotype, and remodeled the tumor microenvironment to allow the killing of tumor cells [221].
6. Application of Macrophage-Targeting Nanotechnology in Disease Treatment
Given the important roles of macrophages in the progression of many diseases, macrophage-targeting therapeutics were also extensively studied in recent years. In this section, the latest developments in disease diagnosis and treatment based on macrophage-targeting NPs will be reviewed and discussed.
6.1. Macrophage-Targeting NPs for Diagnosis
The role of NPs as an imaging/contrast agent enhancer in noninvasive diagnostic technique has been explored due to their unique physical and chemical properties. Moreover, the NP surfaces could be modified to improve the signal intensity and specificity, allowing them to be suitable for the diagnosis of different diseases. As macrophages are involved in the pathology of many diseases (e.g., infection, tumor, atherosclerosis, and rheumatoid arthritis), this cell type has been considered as a suitable imaging target. Monitoring the role of macrophages in inflammation can help us learn more about the pathological process of inflammatory diseases [1].
The superparamagnetic iron oxide (SPIO) NPs coated with macrophage-targeting agents, such as dextran [222,223,224], human ferritin protein cage [225], osteopontin (OPN) [226] or annexin V [227], can induce signal loss in T2-weighted images and, therefore, have been applied as MRI agent to offer pivotal insights into plaque biology, thus assessing inflammatory burdens in atherosclerosis. Lipinski et al. described a scavenger receptor-targeted micelle-based nanoparticle-containing MRI contrast agent for imaging macrophages in atherosclerosis. They reported that the NP could especially target and accumulate in macrophages, which significantly increased the contrast-to-noise ratio (CNR) by 52.5%, while the untargeted NP only increased CNR by 18.7% [173]. Likewise, Li developed class AI scavenger receptor-targeting, glutathione-biomineralized gadolinium-based NPs for noninvasive precise MRI imaging to detect foam macrophages in atherosclerosis plaque. The imaging contrast showed an amplified T1 signal, which precisely targeted macrophages, and exhibited systematic clearance capabilities [228].
In addition to MRI, CT imaging, which has high spatial resolution and short acquisition time, is another frequently used technology for diagnosis. However, the low sensitivity of CT requires the administration of a high dose of NPs, which needs to be investigated in future research. For example, iodine-containing NPs have shown advantages in identifying pro-inflammatory macrophages in vulnerable plaques, but the dose of iodine-NP is 100 mg/kg for mice [229], and the potential toxicity needs to be further studied. PET, which has high tissue penetration and superior sensitivity, is another non-invasive imaging technique. Keliher et al. introduced dextran-coated iron oxide NPs labeled with 89Zr as macrophage-specific PET imaging agents due to their high affinity with macrophages [148]. Their later study described a PET radioactive tracer (18F)-labeled polyglucose NPs, which can be taken up by macrophages with high efficiency to visualize atherosclerotic plaques. This polyglucose NPs showed facilitated imaging in mouse and rabbit atherosclerosis models [230].
6.2. Macrophage-Targeting NPs for Tumor Treatment
As mentioned in this review, TAMs contribute to tumor development via degradation of tumor extracellular matrix, destruction of the basement membrane, promotion of angiogenesis, and recruitment of immunosuppressive cells [221]. Therefore, TAMs have been proposed as therapeutic targets. Strategies have been made to clear TAMs in the tumor environment. Miselis et al. used clodronate-containing liposomes labeled with fluorescent dye to selectively target macrophages in the tumor spheroids and eliminate these cells from the tumor environment. In the animal tumor model, the treatment resulted in a 4-fold decrease in tumor number and a 15-fold decrease in tumor size compared to the control, suggesting that the liposomes successfully decreased the tumor cell density and enhanced apoptosis of tumor cells [231]. Soto et al. used mesoporous silica NPs (MSNs) to load Dox as chemotherapeutics and resulted in enhanced tumor growth arrest. The macrophage-targeting was achieved by encapsulating MSN-Dox into GPs derived from the cell walls of Baker’s yeast. The outer shells of GPs express β-glucan receptors (1,3-D-glucan polysaccharide) that allow for receptor-mediated selective uptake by macrophages [187]. Likewise, Ren encapsuled polymer−lipid hybrid NPs into GPs for macrophage-targeting delivery of cabazitaxel, which showed a slower in vitro drug release and higher drug stability compared with non-coated NPs [186].
The M2-like TAMs can be reprogrammed to M1-like macrophages to induce tumor cell necrosis by secreting inflammatory cytokines. Therefore, another strategy in tumor treatment is to induce the M2-to-M1 TAM phenotype switch to remodel the tumor microenvironment [221]. According to previous studies, TLR agonists (e.g., CpG oligodeoxynucleotides (CpG ODNs), which can induce anti-tumor M1 polarization) and baicalin (which can increase the production of IFN-γ) were used as immune modulators for cancer treatment [232]. However, due to the lack of effective delivery approaches, their applications in vivo are limited, suggesting the potential of M2-targeting NPs in delivery of TLR agonists. In Han’s research, PLGA NPs conjugated M2pep and α-pep peptides were used to transform the M2-like TAMs into the M1-like phenotype by specifically delivering a combination of CpG ODN and baicalin. The result showed that the NPs were effectively ingested by M2-like TAMs both in vitro and in vivo, and the released biomolecules effectively reversed the macrophage phenotype [221]. Shan et al. also developed human ferritin heavy chain (rHF) nanocages modified with M2pep on their surfaces for the targeted delivery of CpG ODNs to M2-like TAMs. These NPs were found to inhibit tumor growth in tumor-bearing mice after intravenous injection by transforming M2 TAMs to the anti-tumor M1 type. Moreover, they discovered that the empty M2pep-rHF NPs without CpG ODNs also exhibited anti-tumor ability [233]. Opanasopit et al. [234] introduced a mannosylated liposome containing an immunomodulator called muramyl dipeptide (MDP), which is a component derived from bacterial cell wall, to target macrophages and activate the M1 polarization in an experimental liver metastases animal model. The result showed that after stimulated by MDP, the production of prostaglandins, collagenase and super-oxide anions by macrophages increased significantly, which induced cytolytic activity against tumor cells [235].
Despite immunomodulation, TAMs are also therapeutic targets for anti-angiogenic treatment. TAMs secret MMPs to release matrix-sequestered VEGF and produce dozens of angiogenic factors to facilitate endothelial survival and proliferation, thus promoting angiogenesis, as well as tumor growth [30,31,32,236]. Penn invented dendrimer NPs conjugated with methotrexate (G5-MTX NPs), a chemotherapeutic that can be recognized by highly expressed FOLR2 on the surface of ovarian TAMs, for anti-angiogenic therapy. G5-MTX NPs overcame the resistance to anti-angiogenic therapy and prevented the side-effects of anti-angiogenic therapy (which induced the generation of cancer stem-like cells) and depleted TAMs in both solid tumor and ascites models of ovarian cancer [202].
Hyperthermia therapy has attracted increasing attention for tumor treatment, as tumor tissues are particularly vulnerable to hyperthermia compared with normal tissues owing to their faster cell proliferation, enhanced hypoxia, low pH, and insufficient temperature regulation ability [237,238]. Chen et al. developed HA conjugated gold nanorods with Dox and acid-labile hydrazone linker attached to the surface as photothermal NPs. After near infrared (NIR) laser irradiation, they enhanced drug release and increased drug toxicity on tumor cells owing to the photothermal effect [176]. Similarly, Wang developed a M2 TAM-targeting iron oxide nanoparticle for MRI-guided MHT of breast tumors. MHT can penetrate deeply into tissues to treat deep tumors and magnetic NPs are commonly used hyperthermia agents. Wang’s NPs also served as contrast agents for MRI, providing diagnostic information and visualizing their distribution in vivo to guide the optimal therapeutic time window [217].
Radiation-induced fibrosis (RIF) is a dose-limiting complication of cancer radiotherapy and causes serious problems, such as restricted tissue flexibility, pain, ulceration, or necrosis [239]. The recruitment of macrophages in inflamed sites can promote inflammatory events and result in fibrosis. Therefore, macrophages are potential cellular targets for anti-inflammatory treatment by inhibiting their production of cytokines. A study successfully treated RIF in a mouse model by intraperitoneal administration of chitosan NPs carrying siRNA to silence TNF-α in local macrophage populations, which takes advantage of the natural homing potential of macrophages to inflammatory sites. They observed the uptake of fluorescently labeled siRNA NPs by peritoneal macrophages and their subsequent migration to lesion region in radiation-induced inflammatory skin, suggesting the chitosan-siRNA NPs may serve as a general therapeutic approach for inflammatory diseases [239].
6.3. Macrophage-Targeting NPs for Infection Control
Infectious diseases are an important health concern, as several pathogens have adapted to survive inside the phagocytic cells, especially macrophages. In some cases, macrophages even serve as nutrient reservoirs to facilitate pathogen growth and spread. Therefore, new therapeutic strategies should be developed to allow for macrophage-targeting drug delivery [162].
Mycobacterium tuberculosis (Mtb) is one of the most threatening pathogens for its latent infection in macrophages. The intracellular Mtb isolated itself from drugs and could spread via macrophages [172]. Mtb displays lipoarabinomannan (LAM) on their cell wall. LAM contains mannose oligosaccharides at the terminus of the molecule, which dominate the attachment of bacteria to macrophages [15,240,241]. Therefore, mannosylated carriers can specifically target and competitively bind to macrophages to deliver antitubercular drugs. A mannose-modified macrophage-targeting solid lipid NP has been developed to load the pH-sensitive prodrug of isoniazid (INH) to treat the latent tuberculosis infection, and the modified NPs showed a higher cell uptake in macrophages (97.2%) than unmodified ones (42.4%), thereby increasing intracellular antibiotic efficiency [172]. Other studies also reported mannosylated gelatin microspheres, gelatin NPs [242], and solid lipid NPs [243,244,245] to carry anti-tuberculosis drugs (such as INH and rifampicin) to target alveolar macrophages. In all cases, increased macrophage uptake and higher reduction in bacterial levels were observed compared with non-mannosylated particles, maintaining therapeutic concentrations for a prolonged period even upon the administration of a reduced clinical dose [170,242,246]. Despite of the mannose, Sharma et al. explored wheat germ agglutinin coated poly (lactide-co-glycolide) NPs as nanocarriers of rifampicin, INH, and pyrazinamide to treat tuberculosis. This nanosystem could reduce the frequency of antitubercular drug administration, therefore improving patient compliance with tuberculosis chemotherapy [247].
Leishmaniasis is a common tropical infectious disease characterized by fever, anemia, weight loss, and the enlargement of the spleen and liver [203]. The etiological agents of leishmaniasis are protozoan parasites called Leishmania donovani, which are intracellular parasites targeting mononuclear phagocytes (monocytes and macrophages) and replicating within membrane-bound subcellular organelles. The parasites develop several mechanisms to survive in macrophages and inhibit parasite-specific cell-mediated host immune response [248]. The classical treatment of leishmaniasis is not effective due to drug resistance, toxicity, bioavailability, and cost [249]. Current treatment for leishmaniasis mainly depends on amphotericin B (AmB), which also has limitations, such as dose-related hematologic toxicity [250], naphrotoxicity [251], stability, and high cost. Therefore, targeted intracellular delivery of AmB is required to enhance drug efficiency and facilitate pathogen clearance. As mentioned previously, mannose-based carriers can specifically target macrophages via interaction with mannose receptors on the cell surface, so studies have developed mannose-anchored thiolated chitosan AmB nanocarrier complexes (MTC AmB) for clearance of Leishmania in macrophages. The result showed a 71-fold increase in MTC AmB uptake compared with native drugs and a 13-fold enhancement in drug efficacy [252]. Despite the mannose, Singh used PhoS, which can be recognized by scavenger receptors (such as CD68 and CD14) on macrophage surfaces for macrophage-targeting. They fabricated PhoS anchored PLGA NPs to deliver AmB, and found that those NPs preferentially accumulated in macrophage-rich organs, which significantly increased anti-leishmanial activity and continually released the drug within 72 h [203].
Human immunodeficiency virus (HIV) infects approximately 35 million people globally and results in 1.8 million deaths every year [253,254]. HIV mainly targets and survives in macrophages, assembling and accumulating in intracellular compartments in macrophages, thus escaping from immune clearance [255]. Given that inhibition of HIV replication could enhance the host response and control infections, studies have been performed to develop macrophage-targeting nanosystems to deliver anti-HIV drugs [256,257]. Zhou et al. decorated NPs with folic acid (FA), which can target folate receptor-β expressed on the surface of activated macrophages [258]. Long-acting cabotegravir, which is an antiretroviral drug, was encapsulated in the nanoparticle and specifically delivered into macrophages. The result showed the slow release of drugs from macrophages, allowing for sustained intracellular drug levels, thus facilitating long-term viral suppression [258].
Among individuals infected by HIV, approximately one-third of them are co-infected with Mtb. HIV-1 infection causes severe damage to the immune system, which enables Mtb to infect and survive in the body more easily. Meanwhile, Mtb infection increases HIV-1 replication, thus enhancing the severity of HIV infection. Given that both HIV-1 virus and Mtb mainly reside in mononuclear macrophages, Narayanasamy and colleagues introduced macrophage-targeting long-acting gallium (Ga) nanoformulation for drug delivery. As a crucial element for the metabolism and growth of most microorganisms, including Mtb and HIV, Ga could stay inside the macrophages for a long time, exhibiting long-term antivirus effects [259].
S. aureus is a Gram-positive bacterium that predominantly infects the skin and the respiratory system, and how to treat S. aureus infection in deep tissue remains a major challenge. Local infection can process into the most serious systemic S. aureus infection-sepsis [260]. The current approach to treat S. aureus infection is using small molecule antibiotics, which cause side effects, such as toxicity and drug resistance [261]. Due to their crucial function in immune response, macrophages are potential targets for anti-infection. Kim synthesized porous Si NPs carrying siRNA against Irf5 to promote phagocytosis of macrophages and inhibit their inflammatory function [262]. The porous Si NPs contain an outer sheath of homing peptides and fusogenic liposomes, allowing them to selectively target macrophages. Irf5 gene highly exists in M1 macrophages, upregulating inflammatory factors while downregulating anti-inflammatory cytokines [263,264]. Knockdown of Irf5 in the early stages of staphylococcal pneumonia can prevent the excretion of inflammatory cytokines and reverse prolonged inflammation, allowing the immune system to clear bacteria and repair tissue [78,265].
Nanotechnology has also been applied to develop novel vaccines against pathogens capable of inducing robust and protective autoimmunity. Chavez-Santoscoy et al. decorated the surface of polyanhydride NPs with specific carbohydrates to provide pathogen-like properties. The carbohydrates, galactose and di-mannose, which were found on the surface of respiratory pathogens, can facilitate both macrophage-targeting and immune activation [266,267,268,269]. This nanovaccine can promote robust pulmonary immunity against many respiratory pathogens, including Yersinia pestis, Mtb, Streptococcus pneumoniae and influenza viruses [178]. To develop an effective vaccine against HIV infection, a macrophage-targeting HIV immunotherapeutic vaccine based on NPs was introduced. The Ebola virus envelope glycoprotein was incorporated with non-replicating virus-like particles (VLPs) to enhance the NP’s macrophage and dendritic cells targeting capability, resulting in a stronger HIV-specific humoral immune response [270]. Hattori et al. introduced mannosylated liposomes, as DNA vaccine carriers and revealed enhanced Th1 immune response, suggesting that nanotechnology was a potent method for DNA vaccine therapy [1].
6.4. Macrophage-Targeting NPs for Bone Regeneration
Bone defects caused by diseases, such as trauma, tumor, and infection, often lead to the non-union of bones, delayed healing or non-healing, and local dysfunction. Although autologous bone transplantation has been widely used to cure bone defects, it has several disadvantages, such as the limited availability of graft volume, the morbidity of the donor site, and the prolonged operation time [271,272,273,274]. To solve these problems, synthesized biomaterials incorporated with osteoinductive factors have become a promising way to treat bone defects [271,272,273,274]. Recent studies have found that the immune system is closely associated with the skeletal system by regulating the biological behavior of bone cells [275,276,277,278], especially for biomaterial applications. Once a foreign material enters the body, it is immediately recognized by the immune system. It triggers activation/inflammation of the immune system, which then influences the subsequent bone regeneration and eventually determines the success of bone biomaterial application in vivo [277,278]. The relationship between foreign biomaterials, host immune cells, and bone cells, is termed as “osteoimmunomodulation”, and biomaterials with appropriate osteoimmunomodulatory capacity can, therefore, modulate local immune microenvironment to favor osteogenesis [23,279].
Among immune cells, the macrophage is one of the most important cell types. The upregulated release of proinflammatory cytokines, including IL-1β, IL-6, and TNF-α from M1 macrophages results in suppression of osteogenesis [280,281] and increased osteoclastogenesis [92]. On the contrary, the anti-inflammatory M2 phenotype can release osteogenic cytokines, such as BMP2 and VEGF, to eliminate inflammation and promote bone healing [20,282,283]. Therefore, nanomaterials are emerging as effective agents able to target macrophages, inducing their M2 polarization and thus modulating bone regeneration. One way to stimulate M2 polarization is to change the surface chemistry of NPs by using bioactive molecules, such as conjugating gold NPs with RGD [91], hexapeptides Cys-Leu-Pro-Phe-Phe-Asp [92], and IL-4 [93], or coating hydroxyapatite on the surface of CeO2 NPs [98]. It is noteworthy that some NPs themselves can enhance M2 polarization, such as gold, TiO2, and CeO2 NPs [94,95,97]. In addition, the nanopore structure and pore size were found to affect the spreading and cell shape of macrophages by modulating their adhesion, which subsequently influences their autophagy, inflammatory response, and release of osteogenic factors [284,285]. For example, Chen found that macrophages on surfaces with larger sized pores (100 and 200 nm) become more anti-inflammatory, producing more pro-inflammatory cytokines and increasing the production of M2 surface-markers [284]. Surface roughness of biomaterials also influences macrophage polarization and cytokine secretion. Studies indicated that titanium with the smooth surface could stimulate M1 macrophage activation, expressing inflammatory cytokines, such as IL-1β, IL-6, and TNF-α, while rough and hydrophilic titanium surface enhanced anti-inflammatory macrophage polarization with the increased secretion of IL-4 and IL-10 [286].
Using nanosystems as carriers to deliver bioactive molecules (including growth factors, cytokines, gene-modulators, and signaling pathway regulators) is another way to induce M1-to-M2 polarization of macrophages. For example, exogenous addition of sphingosine-1-phosphate (S1P), which is a sphingolipid growth factor, stimulates macrophages toward M2-like phenotype [287]. In the study by Das et al., fused nanofibers loaded with S1P synthetic analog were used to direct macrophage polarization towards M2-like phenotype in a mandibular bone defect model and successfully facilitated the osseous repair [288]. Yin et al. developed gold nanocages (AuNC) coated with LPS-stimulated macrophages cell membranes to deliver an anti-inflammatory drug named esolving D1. After LPS stimulation, cytokine receptors on the cell membrane were overexpressed and were able to neutralize pro-inflammatory cytokines [289]. This nanosystem was found to effectively reverse inflammation, facilitate M2 activation and promote osteogenesis in the femoral defect. IL-4 is a well-known anti-inflammatory cytokine, so various nanocarriers have been developed to load IL-4 to induce M2 polarization [290,291,292]. A nanofibrous heparin-modified gelatin microsphere incorporated with IL-4 was developed to resolve the chronic inflammation caused by diabetes and enhance osteogenesis [291]. In another study, the gene of CD163 (a M2 macrophage marker belonging to the scavenger receptor cysteine-rich family) was encapsulated into polyethyleneimine NPs decorated with a mannose ligand to selectively target macrophages to transfer them into anti-inflammatory phenotype [293].
6.5. Macrophage-Targeting NPs for Atherosclerosis
Atherosclerotic cardiovascular disease is a chronic inflammatory disease. During the progress of atherosclerosis, macrophages give rise to early lesions by engulfing low-density lipoprotein (LDL)-derived cholesterol to form lipid-laden foam cells, and the accumulation of cholesterol in macrophages promotes inflammatory responses, thus amplifying inflammation and leading the lesions to progress to an advanced stage [43]. The increasing apoptosis of macrophages leads to plaque necrosis, a key feature of ‘vulnerable’ plaques [44,46,294] characterized by a thin fibrous cap and a large necrotic core. Traditional ways to reduce the risk of cardiovascular events, such as lipid-lowering therapies and anti-inflammatory therapies, have limitations, including low compliance, low bioavailability, poor target specificity, and safety concerns. Therefore, nanotechnology has addressed these challenges and improved disease therapy [295,296].
The pathological processes of atherosclerosis include macrophage recruitment and proliferation; defective efferocytosis, which results in a defect of dead cells movement; plaque inflammation, and cholesterol and oxidized LDL accumulation. Nanotechnology can thus target these processes for atherosclerosis treatment [76]. For instance, monocytes adhere to the arterial wall through cell adhesion molecules, such as intercellular adhesion molecule 1 (ICAM1) and vascular cell adhesion molecule 1 (VCAM1). Sager et al. used polymeric NPs to deliver siRNA to silence five endothelial cell adhesion molecules and inhibited monocyte recruitment to atherosclerotic lesions [297]. Rapamycin reduces the levels of pro-inflammatory cytokines via inhibition of the NF-κB pathway, so targeted delivery of rapamycin by biomimetic NPs [298,299,300] was developed to suppress the proliferation of macrophages and reduce inflammation. The upregulation of CD47 (a “do not eat me” signal) in plaque macrophages can lead to efferocytosis deficiency, so anti-CD47 antibody therapy can efficiently reactivate efferocytosis [301]. Flores et al. encapsulated inhibitor of signal-regulatory protein-α (SIRPα), an antiphagocytic target of CD47, in single-walled carbon nanotubes (SWNTs) for macrophage-specific delivery, and restored efferocytosis in mice model [302]. To reduce plaque inflammation, anti-inflammatory therapeutics were extensively used. IL-10, a well-known anti-inflammatory cytokine, was loaded in PLGA-PEG diblock copolymer NPs and RGD peptide-conjugated pluronic-based nanocarriers to effectively target plaque and decrease inflammation [303,304]. Inhibitors of NF-kB signaling pathways, such as celastrol, and agonists of anti-inflammatory peroxisome proliferator-activated receptor gamma (PPARγ) signaling pathways, such as pioglitazone, were also carried by macrophage-targeting NPs for anti-inflammatory treatment of atherosclerosis [305,306]. A recent study by Wu used M2-like macrophage-derived exosomes as a nanocarrier to deliver anti-inflammatory bio-products. The nanocarrier itself can release anti-inflammatory cytokines, and together with the drug inside of it, the nanosystem promotes the regression of atherosclerotic plaques in Apoe−/− mice [307]. Cholesterol accumulation in plaque macrophages is associated closely with the severity of atherosclerosis. The macrophage ATP-binding cassette (ABC) transporters ABCA1 and ABCG1 are able to transport excess cholesterol to extracellular apolipoprotein A-I (apoA-I) and high-density lipoprotein (HDL), thereby increasing ABCA1 or ABCG1 expression can promote cholesterol efflux from macrophages [308,309,310,311]. miR-33 inhibits cholesterol efflux while miR-206 and miR-223 increase cholesterol efflux, so miR-206, miR-223, and an antisense oligonucleotide against miR-33 were encapsuled in macrophage-targeting NPs to induce cholesterol efflux and reduce plaque burden. The macrophage-targeting ability of these NPs was afforded by cyclic pentapeptides cyclo(-Arg-Gly-Asp-D-Phe-Lys-)(cRGDfK) for integrin receptors on macrophages [312,313]. Despite of miRNA, activation of sterol-regulated transcription factor liver X receptor-α (LXRα) also upregulates express of ABCA1 and ABCG1. Yu and colleagues introduced LXR agonist GW3965 conjugated collagen type IV-targeted polymeric NPs and effectively promoted the cholesterol efflux [313].
6.6. Macrophage-Targeting NPs for Other Inflammation-Related Diseases
A surgical suture is a medical approach to closing the wound of skin and organs, whereas excessive inflammation surrounding the suture can disrupt the wound healing process. To solve this problem, poly(lactic-co-glycolic) NPs decorated with macrophage-targeting ligand mannose were developed to deliver diclofenac, which is an anti-inflammatory drug, as a local anti-inflammatory device. The nanosystem showed an enhanced anti-inflammatory effect in the excisional wound healing animal model compared to free drug-coated suture [314].
Acute lung injury (ALI) is an acute and life-threatening pulmonary inflammatory disease characterized by acute hypoxic respiratory failure caused by pulmonary and nonpulmonary insults [315,316,317]. The pathology of ALI includes infiltration of inflammatory cells, an increase in cytokine release, and uncontrolled inflammation, which further contributes to the accumulation of proteinaceous edema in pulmonary tissue, causing overall acute, diffuse, and inflammatory lung injuries [318]. So far, low tidal volume mechanical ventilation is the only effective therapy of ALI [319,320], but the mortality remains high. Therefore, new therapeutic approaches are needed to limit the acute lung inflammation and induce tissue repair. Macrophages play a crucial role in the inflammation of ALI and, therefore, are considered as a therapeutic target of ALI intervention [321,322]. As mentioned before, gold NPs coated with hexapeptides on the surface could stimulate the M2 restorative polarization of macrophages by inhibiting TLR signaling in macrophages [323], so Wang applied this nanoparticle in LPS induced ALI mouse model to reduce inflammatory cell infiltration, increased M2 polarization and alleviated lung inflammation [92].
Rheumatoid arthritis is an autoimmune disease which tightly related with the activation of macrophages in arthritic joints [324]. The release of inflammatory cytokines induces a rapid nuclear translocation of NF-kB, which then interacts with response-associated genes (such as TNF-α), and activates their transcription [325]. As such, blocking the NF-kB signaling pathway could reverse the inflammation, thus curing autoimmune diseases, such as rheumatoid arthritis. Hattori investigated the delivery of NF-kB-decoy by using folate-linked lipid-based NPs, which selectively target activated macrophages. The decoy was directed at a cognate sequence of NF-kB using a double-stranded oligonucleotide. It blocked the intracellular signaling pathways, resulting in inhibited translocation of NF-kB into the nucleus and decreased levels of inflammatory cytokines [326].
Obesity alters adipose tissue metabolic and endocrine function, which leads to an increased release of fatty acids, hormones, and proinflammatory molecules that contribute to obesity-associated complications [327]. Recent investigations suggest that obesity gives rise to a state of chronic, low-grade inflammation, contributed to by adipose tissue macrophages (ATM) [328,329]. In animal experimentation, lean mice were found to express more M2 phenotype-associated genes, while obese mice expressed more M1 macrophages genes, suggesting that obesity stimulates a M2-to-M1 phenotype change in ATM [330]. In many studies, nanotechnology has been used to delivery anti-obesity therapeutics to increase the stability, solubility, and bioavailability of therapeutics, protecting them from fast degradation in the body, thus prolonging their circulation time [331,332,333]. Moreover, given the crucial role of macrophages in obesity, using NPs to target ATMs in adipose tissue and modulate their polarization may be a direction for anti-obesity treatment. Zhao et al. investigated the function of celastrol-loaded nanomicelles in diet-induced obese mice. The nanosystem significantly reduced the expression of macrophage M1 biomarkers and increased the expression of macrophage M2 biomarkers in a dose-dependent manner, representing a translatable therapeutic opportunity for treating diet-induced obesity in humans. To selectively target ATM and reduce the off-target of small-molecule drugs, Ma invented nanoscale polysaccharides conjugated with dextran, a biocompatible glucosepolymer which can efficiently target macrophages. They loaded anti-inflammatory drugs in the nanoparticle for therapeutic modulation of macrophage phenotype and significantly reduced pro-inflammatory markers in adipose tissue of obese mice, providing promising nanomaterials-based delivery strategy to inhibit obesity [334].
The word “inflammageing” describes a chronic low-grade inflammation observed in ageing population, characterized by an elevated level of circulating inflammatory mediators, such as C reactive protein (CRP), IL-6, IL-1β, and TNF-α [335,336]. Recent data have suggested that although macrophages are not the only source of inflammageing, they are a central component in initiating the phenomenon [335]. Therefore, anti-inflammatory treatment targeting at macrophages may reverse age-related chronic inflammation, thus ameliorating the decline of physiological functions in ageing [337]. Arnardottir et al. applied novel nano-proresolving medicines carrying anti-inflammatory molecules (resolvins D1 and D3) in aged mice and observed promoted M2 macrophage polarization, as well as reduced exacerbated inflammatory response [338]. In our research, we developed an Escherichia. coli OMV coated AuNCs carrying anti-inflammatory drugs to reverse age-related inflammation. Since M1 macrophages are microbicidal, OMVs are supposed to be specially recognized by receptors on M1 macrophages membrane, thus achieving selectively M1 macrophage targeting. Although macrophage-targeting drug delivery nanotechnology has not been largely applied in inflammageing, it provides a new horizon for improving age-related diseases characterized by excessive innate inflammatory responses.
7. Conclusions and Future Direction
Cell-targeting nanotechnology is extensively investigated in a wide range of inflammation-related diseases. This novel approach successfully improves drug efficiency and reduces off-target therapeutics, thus lessening the toxicity of conventional small molecule drugs. Given the vital role of macrophages in immune response and pathology of various diseases, macrophage-targeting NPs have been reported to modulate macrophage behaviors, thus achieving more accurate diagnosis and treatment effects of diseases. NPs can either passively target macrophages by preferentially accumulating in inflammation sites and be taken up by phagocytes, or positively target macrophages by specific interaction between agents decorated on NP surfaces and receptors on macrophages membranes that recognize these agents. This review has summarized current approaches to developing macrophage-targeting NPs (including both passive and positive ways), and their applications in diagnosis and therapy. Despite the numerous advantages of NP-based therapy, there are still issues to be addressed for the optimization of NP treatment. Firstly, targeting M1/M2 macrophages is a challenge, as macrophage phenotype occupies a continuum between M1 and M2 designations, which means macrophages usually possess surface markers of both phenotypes, making it difficult to distinguish M1 from M2 macrophages. Thus, how to detect macrophage populations relying on markers remains to be problematic. Second, the most proper time to intervene the macrophage polarization still needs to be further explored. For example, in bone regeneration, the M1 phenotype dominates the early stage of inflammation, while M2 macrophages play a prominent role in the later stage. A timely M1–M2 switch determines the appropriate immune environment that decides later bone remodeling. However, the proper time to complete this transformation remains unknown and needs to be discovered in the future. Thirdly, although macrophage-targeting NPs have achieved the spatial control of drug release in the targeted cells, the temporal control of drug release still needs to be improved. Most NPs can enable a slow release of loaded therapeutics, avoiding the robust increase in local drug concentration, thereby reducing side effects and enhancing the efficiency of drugs. Some NPs are able to more precisely achieve temporal control of drug release, such as gold nanocages, which are supposed to release drugs only after near-infrared radiation [339]. However, drug leakage still exists before the radiation due to the slow melting of phase-change materials [340]. Therefore, novel NPs with reduced drug leakage and more accurate temporally release control need to be developed. Moreover, though modulation of the immune profile of macrophages by NPs has shown great promise in animal experiments, their efficacy in humans cannot be guaranteed due to the differences in pathobiology between animal models and humans. For example, the procession of atherosclerosis is usually faster in animal models compared with that in humans, suggesting a more complicated and heterogeneous pathology in the human body [341]. In addition, macrophage subtypes also vary in different individuals, making personalized nanomedicine particularly crucial to achieving the desired therapeutic outcomes [76]. In addition, the physicochemical interactions between NPs and targeted cell surfaces, the pharmacokinetics and biodistribution of NPs, and the underlying mechanism of their immunomodulation need further investigation to provide the foundation for improving nanotherapeutics in the future.
Author Contributions
D.C. wrote this paper and drew figures; W.G. assisted in drawing and reviewed this paper; Y.Z. and Z.L. reviewed this paper; L.X. and Y.X. designed the project and reviewed this paper. All authors have approved the submitted version and agreed to be personally accountable for their own contributions and for ensuring that questions related to the accuracy or integrity of any part of the work, even ones in which the author was not personally involved, are appropriately investigated, resolved, and documented in the literature. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the NHMRC Idea Grant (APP2000647) of Australia, JRC (2019) from Queensland Department of Environment and Science, ITI Research Grant 1260_2017, Young Researcher Grant (19-066) from the Osteology Foundation, Switzerland, QUT Centre for Biomedical Technologies ECR/MCR Grant Scheme 2021.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Abbreviation | Full Names |
| NPs | Nanoparticles |
| MPS | Mononuclear phagocyte system |
| LPS | Lipopolysaccharide |
| IFN-γ | Interferon-gamma |
| TNF-α | Tumor necrosis factor-alpha |
| IL-1β,4,6,10,12,13 | Interleukin-1β,4,6,10,12,13 |
| NO | Nitric oxide |
| iNOS | Inducible nitric oxide synthase |
| TLR | Toll-like receptors |
| TCR | T cell receptors |
| M-CSF | Macrophage-colony stimulating factor |
| NF-kB | Nuclear factor-κB |
| TGF-β1, β3 | Tissue growth factor-β1, β3 |
| BMP | Bone morphogenetic protein |
| VEGF | Vascular endothelial growth factor |
| TAMs | Tumor-associated macrophages |
| PEG | Polyethylene glycol |
| MMPs | Matrix metalloproteinases |
| Arg-1 | Arginase-1 |
| MARCO | Macrophage receptor with collagenous domain |
| PMR | Polymyalgia rheumatic |
| GCA | Giant cell arteritis |
| COPD | Chronic obstructive pulmonary disease |
| siRNA | Small interfering RNA |
| miRNA | MicroRNA |
| RGD | Arginine-glycine-aspartic acid |
| TiO2 | Titanium oxide |
| Ca | Calcium |
| Sr | Strontium |
| CeO2 | Cerium oxide |
| QD | Quantum dot |
| MRI | Magnetic resonance imaging |
| CT | X-ray computed tomography |
| PET | Positron emission tomography |
| SPECT | Single-photon emission computed tomography |
| RES | Reticuloendothelial system |
| ROS | Reactive oxygen species |
| Si | Silicon |
| ZnO | Zinc oxide |
| EPR | Enhanced permeability and retention |
| CLRs | C-type lectin receptors |
| HA | Hyaluronic acid |
| RHAMM | Receptor for hyaluronic acid-mediated motility |
| Au NR | Gold nanorod |
| Dox | Doxorubicin |
| Zr | Zirconium |
| GPs | Glucan particles |
| OMVs | Outer membrane vehicles |
| BNCs | Bio-nanocapsules |
| HBV | Hepatitis B virus |
| FR | Folate receptor |
| FOLR2 | Folate receptor-2 |
| PhoS | Phosphatidylserine |
| BDNF | Brain-derived neurotrophic factor |
| MODS | Multiple organ dysfunction syndromes |
| mAb | Monoclonal antibody |
| cITP | Chronic immune thrombocytopenia purpura |
| M2pep | M2 macrophage-binding peptide |
| MHT | Magnetic hyperthermia therapy |
| PLGA | Poly (lactic-co-glycolic acid) |
| SPIO | Superparamagnetic iron oxide |
| OPN | Osteopontin |
| CNR | Contrast-to-noise ratio |
| MSNs | Mesoporous silica NPs |
| ODNs | Oligodeoxynucleotides |
| rHF | Human ferritin heavy chain |
| MDP | Muramyl dipeptide |
| NIR | Near infrared |
| RIF | Radiation-induced fibrosis |
| Mtb | Mycobacterium tuberculosis |
| LAM | Lipoarabinomannan |
| INH | Isoniazid |
| AmB | Amphotericin B |
| HIV | Human immunodeficiency virus |
| FA | Folic acid |
| Ga | Gallium |
| S. aureus | Staphyloccocus. aureus |
| VLPs | Virus-like particles |
| S1P | Sphingosine-1-phosphate |
| AuNC | Gold nanocage |
| LDL | Low-density lipoprotein |
| ICAM1 | Intercellular adhesion molecule 1 |
| VCAM1 | Vascular cell adhesion molecule 1 |
| SIRPα | Signal-regulatory protein-α |
| SWNTs | Single-walled carbon nanotubes |
| PPARγ | Peroxisome proliferator-activated receptor gamma |
| ABC | ATP-binding cassette |
| apoA-I | Apolipoprotein A-I |
| HDL | High-density lipoprotein |
| LXRα | Liver X receptor-α |
| ALI | Acute lung injury |
| ATM | Adipose tissue macrophages |
| CRP | C reactive protein |
References
- Jain, S.; Amiji, M. Macrophage-targeted nanoparticle delivery systems. In Multifunctional Nanoparticles for Drug Delivery Applications; Springer: Boston, MA, USA, 2012; pp. 47–83. [Google Scholar]
- Pentecost, A.E.; Lurier, E.B.; Spiller, K.L. Nanoparticulate systems for controlling monocyte/macrophage behavior. In Microscale Technologies for Cell Engineering; Springer: Cham, Switzerland, 2016; pp. 291–304. [Google Scholar]
- Fujiwara, N.; Kazuo, K. Macrophages in inflammation. Curr. Drug Targets-Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.P.; Wynn, T.A. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Mills, C.D.; Kincaid, K.; Alt, J.M.; Heilman, M.J.; Hill, A.M. M-1/M-2 macrophages and the Th1/Th2 paradigm. J. Immunol. 2000, 164, 6166–6173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, H.M.; Robert, N.B.; Lars-Peter, E. Macrophages: Promising targets for the treatment of atherosclerosis. Curr. Vasc. Pharmacol. 2009, 7, 234–243. [Google Scholar] [CrossRef]
- Mosser, D.M. The many faces of macrophage activation. J. Leukoc. Biol. 2003, 73, 209–212. [Google Scholar] [CrossRef]
- Wang, N.; Liang, H.; Ke, Z. Molecular mechanisms that influence the macrophage M1–M2 polarization balance. Front. Immunol. 2014, 5, 614. [Google Scholar] [CrossRef] [Green Version]
- Mills, C. M1 and M2 macrophages: Oracles of health and disease. Crit. Rev. Immunol. 2012, 32, 463–488. [Google Scholar] [CrossRef] [Green Version]
- Dermine, J.; Michel, D. Survival of intracellular pathogens within macrophages. Protoplasma 1999, 210, 11–24. [Google Scholar] [CrossRef]
- Rosenberger, M.C.; Finlay, B.B. Phagocyte sabotage: Disruption of macrophage signalling by bacterial pathogens. Nat. Rev. Mol. Cell Biol. 2003, 4, 385–396. [Google Scholar] [CrossRef]
- Thi, P.E.; Lambertz, U.; Reiner, N.E. Sleeping with the enemy: How intracellular pathogens cope with a macrophage lifestyle. PLoS Pathog. 2012, 8, e1002551. [Google Scholar] [CrossRef] [Green Version]
- Radtke, A.L.; Anderson, K.L.; Davis, M.J.; DiMagno, M.J.; Swanson, J.A.; O’Riordan, M.X. Listeria monocytogenes exploits cystic fibrosis transmembrane conductance regulator (CFTR) to escape the phagosome. Proc. Natl. Acad. Sci. USA 2011, 108, 1633–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashida, H.; Ogawa, M.; Mimuro, H.; Kobayashi, T.; Sanada, T.; Sasakawa, C. Shigella are versatile mucosal pathogens that circumvent the host innate immune system. Curr. Opin. Immunol. 2011, 23, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Fratti, R.A.; Chua, J.; Vergne, I.; Deretic, V. Mycobacterium tuberculosis glycosylated phosphatidylinositol causes phagosome maturation arrest. Proc. Natl. Acad. Sci. USA 2003, 100, 5437–5442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilbi, H. (Ed.) Molecular Mechanisms in Legionella Pathogenesis; Springer Science & Business Media: Berlin, Germany, 2014; Volume 376. [Google Scholar]
- Newton, H.J.; Kohler, L.J.; McDonough, J.A.; Temoche-Diaz, M.; Crabill, E.; Hartland, E.L.; Roy, C.R. A screen of Coxiella burnetii mutants reveals important roles for Dot/Icm effectors and host autophagy in vacuole biogenesis. PLoS Pathog. 2014, 10, e1004286. [Google Scholar] [CrossRef] [Green Version]
- Chang, M.K.; Raggatt, L.J.; Alexander, K.A.; Kuliwaba, J.S.; Fazzalari, N.L.; Schroder, K.; Pettit, A.R. Osteal tissue macrophages are intercalated throughout human and mouse bone lining tissues and regulate osteoblast function in vitro and in vivo. J. Immunol. 2008, 181, 1232–1244. [Google Scholar] [CrossRef] [Green Version]
- Guihard, P.; Danger, Y.; Brounais, B.; David, E.; Brion, R.; Delecrin, J.; Blanchard, F. Induction of osteogenesis in mesenchymal stem cells by activated monocytes/macrophages depends on oncostatin M signaling. Stem Cells 2012, 30, 762–772. [Google Scholar] [CrossRef] [Green Version]
- Rifas, L. T-cell cytokine induction of BMP-2 regulates human mesenchymal stromal cell differentiation and mineralization. J. Cell. Biochem. 2006, 98, 706–714. [Google Scholar] [CrossRef]
- Brown, N.B.; Badylak, S.F. Expanded applications, shifting paradigms and an improved understanding of host–biomaterial interactions. Acta Biomater. 2013, 9, 4948–4955. [Google Scholar] [CrossRef]
- Mokarram, N.; Ravi, V.B. A perspective on immunomodulation and tissue repair. Ann. Biomed. Eng. 2014, 42, 338–351. [Google Scholar] [CrossRef]
- Chen, Z.; Klein, T.; Murray, R.Z.; Crawford, R.; Chang, J.; Wu, C.; Xiao, Y. Osteoimmunomodulation for the development of advanced bone biomaterials. Mater. Today 2016, 19, 304–321. [Google Scholar] [CrossRef] [Green Version]
- Lewis, C.; Murdoch, C. Macrophage responses to hypoxia: Implications for tumor progression and anti-cancer therapies. Am. J. Pathol. 2005, 167, 627–635. [Google Scholar] [CrossRef]
- Ono, M. Molecular links between tumor angiogenesis and inflammation: Inflammatory stimuli of macrophages and cancer cells as targets for therapeutic strategy. Cancer Sci. 2008, 99, 1501–1506. [Google Scholar] [CrossRef] [PubMed]
- Germano, G.; Frapolli, R.; Belgiovine, C.; Anselmo, A.; Pesce, S.; Liguori, M.; Allavena, P. Role of macrophage targeting in the antitumor activity of trabectedin. Cancer Cell 2013, 23, 249–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosser, M.D.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Leek, D.R.; Harris, A.L. Tumor-associated macrophages in breast cancer. J. Mammary Gland. Biol. Neoplasia 2002, 7, 177–189. [Google Scholar] [CrossRef]
- Elgert, K.D.; David, G.A.; David, W.M. Tumor-induced immune dysfunction: The macrophage connection. J. Leukoc. Biol. 1998, 64, 275–290. [Google Scholar] [CrossRef]
- Conejo-Garcia, J.R.; Buckanovich, R.J.; Benencia, F.; Courreges, M.C.; Rubin, S.C.; Carroll, R.G.; Coukos, G. Vascular leukocytes contribute to tumor vascularization. Blood 2005, 105, 679–681. [Google Scholar] [CrossRef] [Green Version]
- Rehman, J.; Li, J.; Orschell, C.M.; March, K.L. Peripheral blood “endothelial progenitor cells” are derived from monocyte/macrophages and secrete angiogenic growth factors. Circulation 2003, 107, 1164–1169. [Google Scholar] [CrossRef] [Green Version]
- Gottfried, E.; Kreutz, M.; Haffner, S.; Holler, E.; Iacobelli, M.; Andreesen, R.; Eissner, G. Differentiation of human tumour-associated dendritic cells into endothelial-like cells: An alternative pathway of tumour angiogenesis. Scand. J. Immunol. 2007, 65, 329–335. [Google Scholar] [CrossRef]
- Kelly, C.; Jefferies, C.; Cryan, S. Targeted liposomal drug delivery to monocytes and macrophages. J. Drug Deliv. 2011, 2011, 727241. [Google Scholar] [CrossRef]
- Biswas, K.S.; Mantovani, A. Macrophage plasticity and interaction with lymphocyte subsets: Cancer as a paradigm. Nat. Immunol. 2010, 11, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Grohmann, U.; Belladonna, M.L.; Vacca, C.; Bianchi, R.; Fallarino, F.; Orabona, C.; Puccetti, P. Positive regulatory role of IL-12 in macrophages and modulation by IFN-γ. J. Immunol. 2001, 167, 221–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Ginderachter, J.A.; Movahedi, K.; Ghassabeh, G.H.; Meerschaut, S.; Beschin, A.; Raes, G.; De Baetselier, P. Classical and alternative activation of mononuclear phagocytes: Picking the best of both worlds for tumor promotion. Immunobiology 2006, 211, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Parayath, N.N.; Parikh, A.; Amiji, M.M. Repolarization of tumor-associated macrophages in a genetically engineered nonsmall cell lung cancer model by intraperitoneal administration of hyaluronic acid-based nanoparticles encapsulating microRNA-125b. Nano Lett. 2018, 18, 3571–3579. [Google Scholar] [PubMed]
- Kim, S.Y.; Kim, S.; Kim, J.E.; Lee, S.N.; Shin, I.W.; Shin, H.S.; Lim, Y.T. Lyophilizable and multifaceted toll-like receptor 7/8 agonist-loaded nanoemulsion for the reprogramming of tumor microenvironments and enhanced cancer immunotherapy. ACS Nano 2019, 13, 12671–12686. [Google Scholar] [PubMed]
- Singh, Y.; Pawar, V.K.; Meher, J.G.; Raval, K.; Kumar, A.; Shrivastava, R.; Chourasia, M.K. Targeting tumor associated macrophages (TAMs) via nanocarriers. J. Control. Release 2017, 254, 92–106. [Google Scholar] [PubMed]
- Virani, S.S.; Alonso, A.; Aparicio, H.J.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W. Heart disease and stroke statistics—2021 update: A report from the American Heart Association. Circulation 2021, 143, e254–e743. [Google Scholar] [CrossRef]
- Mestas, J.; Ley, K. Monocyte-endothelial cell interactions in the development of atherosclerosis. Trends Cardiovasc. Med. 2008, 18, 228–232. [Google Scholar]
- Glass, C.K.; Joseph, L.W. Atherosclerosis: The road ahead. Cell 2001, 104, 503–516. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.K.; Tabas, I. Macrophages in the pathogenesis of atherosclerosis. Cell 2011, 145, 341–355. [Google Scholar] [CrossRef] [Green Version]
- Doran, A.C.; Yurdagul, A.; Tabas, I. Efferocytosis in health and disease. Nat. Rev. Immunol. 2019, 20, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Yurdagul Jr, A.; Doran, A.C.; Cai, B.; Fredman, G.; Tabas, I.A. Mechanisms and consequences of defective efferocytosis in atherosclerosis. Front. Cardiovasc. Med. 2018, 4, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasikara, C.; Doran, A.C.; Cai, B.; Tabas, I. The role of non-resolving inflammation in atherosclerosis. J. Clin. Investig. 2018, 128, 2713–2723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeksema, M.A.; Stöger, J.L.; de Winther, M.P.J. Molecular pathways regulating macrophage polarization: Implications for atherosclerosis. Curr. Atheroscler. Rep. 2012, 14, 254–263. [Google Scholar] [CrossRef] [Green Version]
- Galis, Z.S.; Sukhova, G.K.; Lark, M.W.; Libby, P. Increased expression of matrix metalloproteinases and matrix degrading activity in vulnerable regions of human atherosclerotic plaques. J. Clin. Investig. 1994, 94, 2493–2503. [Google Scholar] [CrossRef] [Green Version]
- Quillard, T.; Tesmenitsky, Y.; Croce, K.; Travers, R.; Shvartz, E.; Koskinas, K.C.; Libby, P. Selective inhibition of matrix metalloproteinase-13 increases collagen content of established mouse atherosclerosis. Arter. Thromb. Vasc. Biol. 2011, 31, 2464–2472. [Google Scholar] [CrossRef] [Green Version]
- Herman, M.P.; Sukhova, G.K.; Libby, P.; Gerdes, N.; Tang, N.; Horton, D.B.; Schönbeck, U. Expression of neutrophil collagenase (matrix metalloproteinase-8) in human atheroma: A novel collagenolytic pathway suggested by transcriptional profiling. Circulation 2001, 104, 1899–1904. [Google Scholar] [CrossRef] [Green Version]
- Boyle, J.J.; Weissberg, P.L.; Bennett, M.R. Tumor Necrosis Factor-α Promotes Macrophage-Induced Vascular Smooth Muscle Cell Apoptosis by Direct and Autocrine Mechanisms. Arter. Thromb. Vasc. Biol. 2003, 23, 1553–1558. [Google Scholar] [CrossRef] [Green Version]
- Doran, A.C.; Ozcan, L.; Cai, B.; Zheng, Z.; Fredman, G.; Rymond, C.C.; Tabas, I. CAMKIIγ suppresses an efferocytosis pathway in macrophages and promotes atherosclerotic plaque necrosis. J. Clin. Investig. 2017, 127, 4075–4089. [Google Scholar] [CrossRef]
- Naghavi, M.; Libby, P.; Falk, E.; Casscells, S.W.; Litovsky, S.; Rumberger, J.; Willerson, J.T. From vulnerable plaque to vulnerable patient: A call for new definitions and risk assessment strategies: Part I. Circulation 2003, 108, 1664–1672. [Google Scholar] [CrossRef]
- Jaffer, A.F.; Libby, P.; Weissleder, R. Molecular and cellular imaging of atherosclerosis: Emerging applications. J. Am. Coll. Cardiol. 2006, 47, 1328–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaffer, A.F.; Weissleder, R. Molecular imaging in the clinical arena. JAMA 2005, 293, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Li, C.A.; Glass, C.K. The macrophage foam cell as a target for therapeutic intervention. Nat. Med. 2002, 8, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Paul, M.R.; Attilio, M. Inflammation and atherosclerosis. Circulation 2002, 105, 1135–1143. [Google Scholar] [CrossRef]
- Leon, D.A. Trends in European life expectancy: A salutary view. Int. J. Epidemiol. 2011, 40, 271–277. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Sharpless, N.E. Senescence in health and disease. Cell 2017, 169, 1000–1011. [Google Scholar] [CrossRef]
- Pereira, B.I.; Devine, O.P.; Vukmanovic-Stejic, M.; Chambers, E.S.; Subramanian, P.; Patel, N.; Akbar, A.N. Senescent cells evade immune clearance via HLA-E-mediated NK and CD8+ T cell inhibition. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Chen, H.; Ma, F.; Hu, X.; Jin, T.; Xiong, C.; Teng, X. Elevated COX2 expression and PGE2 production by downregulation of RXRα in senescent macrophages. Biochem. Biophys. Res. Commun. 2013, 440, 157–162. [Google Scholar] [CrossRef]
- Di Mitri, D.; Azevedo, R.I.; Henson, S.M.; Libri, V.; Riddell, N.E.; Macaulay, R.; Akbar, A.N. Reversible senescence in human CD4 + CD45RA + CD27− memory T cells. J. Immunol. 2011, 187, 2093–2100. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.Y.; Pan, Z.Y.; Yu, X.K.; Chen, Y.F.; Gao, C.H.; Yang, Y.T.; Pan, J.P. Bifidobacterium lactis BB-12 attenuates macrophage aging induced by D-Galactose and promotes M2 macrophage polarization. J. Immunol. Res. 2019, 2019, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Bailey, W.M.; Braun, K.J.; Gensel, J.C. Age decreases macrophage IL-10 expression: Implications for functional recovery and tissue repair in spinal cord injury. Exp. Neurol. 2015, 273, 83–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karuppagounder, V.; Giridharan, V.V.; Arumugam, S.; Sreedhar, R.; Palaniyandi, S.S.; Krishnamurthy, P.; Thandavarayan, R.A. Modulation of macrophage polarization and HMGB1-TLR2/TLR4 cascade plays a crucial role for cardiac remodeling in senescence-accelerated prone mice. PLoS ONE 2016, 11, e0152922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cudejko, C.; Wouters, K.; Fuentes, L.; Hannou, S.A.; Paquet, C.; Bantubungi, K.; Paumelle, R. p16INK4a deficiency promotes IL-4–induced polarization and inhibits proinflammatory signaling in macrophages. Blood 2011, 118, 2556–2566. [Google Scholar] [CrossRef]
- Ogawa, T.; Masanobu, K.; Katsuiku, H. Age-related changes of human bone marrow: A histometric estimation of proliferative cells, apoptotic cells, T cells, B cells and macrophages. Mech. Ageing Dev. 2000, 117, 57–68. [Google Scholar] [CrossRef]
- Jackaman, C.; Radley-Crabb, H.G.; Soffe, Z.; Shavlakadze, T.; Grounds, M.D.; Nelson, D.J. Targeting macrophages rescues age-related immune deficiencies in C57 BL/6J geriatric mice. Aging Cell 2013, 12, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.; Khan, A.A.; Yin, J.; Ferguson, T.A.; Apte, R.S. Senescence regulates macrophage activation and angiogenic fate at sites of tissue injury in mice. J. Clin. Investig. 2007, 117, 3421–3426. [Google Scholar] [CrossRef]
- Wong, C.K.; Smith, C.A.; Sakamoto, K.; Kaminski, N.; Koff, J.L.; Goldstein, D.R. Aging impairs alveolar macrophage phagocytosis and increases influenza-induced mortality in mice. J. Immunol. 2017, 199, 1060–1068. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Jiao, Y.; Fan, E.K.; Scott, M.J.; Li, Y.; Li, S.; Fan, J. Aging-impaired filamentous actin polymerization signaling reduces alveolar macrophage phagocytosis of bacteria. J. Immunol. 2017, 199, 3176–3186. [Google Scholar] [CrossRef]
- Wang, C.; Xu, J.; Yang, L.; Xu, Y.; Zhang, X.; Bai, C. Prevalence and risk factors of chronic obstructive pulmonary disease in China (the China Pulmonary Health [CPH] study): A national cross-sectional study. Lancet 2018, 391, 1706–1717. [Google Scholar] [CrossRef]
- Thannickal, V.J.; Murthy, M.; Balch, W.E.; Chandel, N.S.; Meiners, S.; Eickelberg, O.; Budinger, G.S. Blue journal conference. Aging and susceptibility to lung disease. Am. J. Respir. Crit. Care Med. 2015, 191, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Zhou, C.; Kalionis, B.; Shuang, X.; Ge, H.; Gao, W. Combined antioxidant, anti-inflammaging and mesenchymal stem cell treatment: A possible therapeutic direction in elderly patients with chronic obstructive pulmonary disease. Aging Dis. 2020, 11, 129. [Google Scholar] [PubMed] [Green Version]
- De Maeyer, R.P.H.; Emma, S.C. The impact of ageing on monocytes and macrophages. Immunol. Lett. 2021, 230, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Schilperoort, M.; Cao, Y.; Shi, J.; Tabas, I.; Tao, W. Macrophage-targeted nanomedicine for the diagnosis and treatment of atherosclerosis. Nat. Rev. Cardiol. 2021, 19, 228–249. [Google Scholar] [CrossRef]
- Maeda, H.; Matsumura, Y. Tumoritropic and lymphotropic principles of macromolecular drugs. Crit. Rev. Ther. Drug Carr. Syst. 1989, 6, 193–210. [Google Scholar]
- Leuschner, F.; Dutta, P.; Gorbatov, R.; Novobrantseva, T.I.; Donahoe, J.S.; Courties, G.; Nahrendorf, M. Therapeutic siRNA silencing in inflammatory monocytes in mice. Nat. Biotechnol. 2011, 29, 1005–1010. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.-M.; Thaxton, C.S.; Mirkin, C.A. Nanoparticle-Based Bio-Bar Codes for the Ultrasensitive Detection of Proteins. Science 2003, 301, 1884–1886. [Google Scholar] [CrossRef] [Green Version]
- Tsapis, N.; Bennett, D.; Jackson, B.; Weitz, D.A.; Edwards, D.A. Trojan particles: Large porous carriers of nanoparticles for drug delivery. Proc. Natl. Acad. Sci. USA 2002, 99, 12001–12005. [Google Scholar] [CrossRef] [Green Version]
- Lobatto, M.E.; Fuster, V.; Fayad, Z.A.; Mulder, W.J. Perspectives and opportunities for nanomedicine in the management of atherosclerosis. Nat. Rev. Drug Discov. 2011, 10, 835–852. [Google Scholar] [CrossRef] [Green Version]
- Song, G.; Petschauer, J.S.; Madden, A.J.; Zamboni, W.C. Nanoparticles and the mononuclear phagocyte system: Pharmacokinetics and applications for inflammatory diseases. Curr. Rheumatol. Rev. 2014, 10, 22–34. [Google Scholar] [CrossRef]
- Zamboni, W.C. Liposomal, nanoparticle, and conjugated formulations of anticancer agents. Clin. Cancer Res. 2005, 11, 8230–8234. [Google Scholar] [CrossRef] [Green Version]
- Zolnik, B.S.; González-Fernández, Á.; Sadrieh, N.; Dobrovolskaia, M.A. Minireview: Nanoparticles and the immune system. Endocrinology 2010, 151, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Peer, D.; Karp, J.M.; Hong, S.; Farokhzad, O.C.; Margalit, R.; Langer, R. Nanocarriers as an emerging platform for cancer therapy. Nat. Nanotechnol. 2007, 2, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Rosenblum, D.; Joshi, N.; Tao, W.; Karp, J.M.; Peer, D. Progress and challenges towards targeted delivery of cancer therapeutics. Nat. Commun. 2018, 9, 1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Glackin, C.A.; Horwitz, M.A.; Zink, J.I. Nanomachines and other caps on mesoporous silica nanoparticles for drug delivery. Accounts Chem. Res. 2019, 52, 1531–1542. [Google Scholar] [CrossRef]
- Chuang, S.Y.; Lin, C.F.; A Aljuffali, I.; Fang, J.Y. Specific targeting of engineered nanoparticles to activated macrophages. Curr. Nanosci. 2015, 12, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Hasan, A.; Morshed, M.; Memic, A.; Hassan, S.; Webster, T.J.; Marei, H.E.S. Nanoparticles in tissue engineering: Applications, challenges and prospects. Int. J. Nanomed. 2018, 13, 5637. [Google Scholar] [CrossRef] [Green Version]
- Ni, C.; Zhou, J.; Kong, N.; Bian, T.; Zhang, Y.; Huang, X.; Yan, F. Gold nanoparticles modulate the crosstalk between macrophages and periodontal ligament cells for periodontitis treatment. Biomaterials 2019, 206, 115–132. [Google Scholar] [CrossRef]
- Bartneck, M.; Ritz, T.; Keul, H.A.; Wambach, M.; Bornemann, J.; Gbureck, U.; Tacke, F. Peptide-functionalized gold nanorods increase liver injury in hepatitis. ACS Nano 2012, 6, 8767–8777. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, H.; Sun, L.; Gao, W.; Xiong, Y.; Ma, A.; Yang, H. Manipulation of macrophage polarization by peptide-coated gold nanoparticles and its protective effects on acute lung injury. J. Nanobiotechnol. 2020, 18, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Raimondo, T.M.; Mooney, D.J. Functional muscle recovery with nanoparticle-directed M2 macrophage polarization in mice. Proc. Natl. Acad. Sci. USA 2018, 115, 10648–10653. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Yu, Y.; Ma, P.; Luo, Z.; Hu, Y.; Li, M.; Cai, K. Titania nanotubes promote osteogenesis via mediating crosstalk between macrophages and MSCs under oxidative stress. Colloids Surf. B Biointerfaces 2019, 180, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Kim, Y.J.; Jang, J.H.; Park, J.W. Modulating macrophage polarization with divalent cations in nanostructured titanium implant surfaces. Nanotechnology 2016, 27, 085101. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.C.; Dong, X.; Ding, J.L.; Liu, J.C.; Xu, J.J.; Tang, Y.H.; Zhou, J.L. Nanotubular TiO2 regulates macrophage M2 polarization and increases macrophage secretion of VEGF to accelerate endothelialization via the ERK1/2 and PI3K/AKT pathways. Int. J. Nanomed. 2019, 14, 441. [Google Scholar] [CrossRef] [Green Version]
- Chigurupati, S.; Mughal, M.R.; Okun, E.; Das, S.; Kumar, A.; McCaffery, M.; Mattson, M.P. Effects of cerium oxide nanoparticles on the growth of keratinocytes, fibroblasts and vascular endothelial cells in cutaneous wound healing. Biomaterials 2013, 34, 2194–2201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Shen, Q.; Xie, Y.; You, M.; Huang, L.; Zheng, X. Incorporation of cerium oxide into hydroxyapatite coating regulates osteogenic activity of mesenchymal stem cell and macrophage polarization. J. Biomater. Appl. 2016, 31, 1062–1076. [Google Scholar] [CrossRef]
- Li, J.; Wen, J.; Li, B.; Li, W.; Qiao, W.; Shen, J.; Chu, P.K. Valence state manipulation of cerium oxide nanoparticles on a titanium surface for modulating cell fate and bone formation. Adv. Sci. 2017, 5, 1700678. [Google Scholar] [CrossRef]
- Port, M.; Idée, J.M.; Medina, C.; Robic, C.; Sabatou, M.; Corot, C. Efficiency, thermodynamic and kinetic stability of marketed gadolinium chelates and their possible clinical consequences: A critical review. BioMetals 2008, 21, 469–490. [Google Scholar] [CrossRef]
- Beckmann, N.; Cannet, C.; Babin, A.L.; Blé, F.X.; Zurbruegg, S.; Kneuer, R.; Dousset, V. In vivo visualization of macrophage infiltration and activity in inflammation using magnetic resonance imaging. WIREs Nanomed. Nanobiotechnol. 2009, 1, 272–298. [Google Scholar] [CrossRef]
- Sosnovik, D.E.; Ralph, W. Emerging concepts in molecular MRI. Curr. Opin. Biotechnol. 2007, 18, 4–10. [Google Scholar] [CrossRef]
- Tejwani, A.; Lavaf, A.; Parikh, K.; Mokhtar, B.; Swamy, U.; Emmolo, J.; Ashamalla, H. The role of PET/CT in decreasing inter-observer variability in treatment planning and evaluation of response for cervical cancer. Am. J. Nucl. Med. Mol. Imaging 2012, 2, 307. [Google Scholar]
- Chen, D.; Dougherty, C.A.; Zhu, K.; Hong, H. Theranostic applications of carbon nanomaterials in cancer: Focus on imaging and cargo delivery. J. Control. Release 2015, 210, 230–245. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Hong, H.; Xu, Z.P.; Li, Z.; Cai, W. Quantum dot-based nanoprobes for in vivo targeted imaging. Curr. Mol. Med. 2013, 13, 1549–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartneck, M.; Scheyda, K.M.; Warzecha, K.T.; Rizzo, L.Y.; Hittatiya, K.; Luedde, T.; Tacke, F. Fluorescent cell-traceable dexamethasone-loaded liposomes for the treatment of inflammatory liver diseases. Biomaterials 2014, 37, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, R.; Hueber, A.J.; Hutton, A.; McInnes, I.B.; Graham, D. Nanoparticles and inflammation. Sci. World J. 2011, 11, 1300–1312. [Google Scholar] [CrossRef] [PubMed]
- Cabral, H.; Kataoka, K. Progress of drug-loaded polymeric micelles into clinical studies. J. Control. Release 2014, 190, 465–476. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.T.; Keller, S.; Manganiello, M.J.; Cheng, C.; Lee, C.C.; Opara, C.; Stayton, P.S. pH-Responsive nanoparticle vaccines for dual-delivery of antigens and immunostimulatory oligonucleotides. ACS Nano 2013, 7, 3912–3925. [Google Scholar] [CrossRef]
- Kannan, R.M.; Nance, E.; Kannan, S.; Tomalia, D.A. Emerging concepts in dendrimer-based nanomedicine: From design principles to clinical applications. J. Intern. Med. 2014, 276, 579–617. [Google Scholar] [CrossRef]
- Zamboni, W.C. Concept and clinical evaluation of carrier-mediated anticancer agents. Oncologist 2008, 13, 248–260. [Google Scholar] [CrossRef]
- Li, S.-D.; Huang, L. Pharmacokinetics and Biodistribution of Nanoparticles. Mol. Pharm. 2008, 5, 496–504. [Google Scholar] [CrossRef]
- Dobrovolskaia, A.M.; McNeil, S.E. Immunological properties of engineered nanomaterials. Nanosci. Technol. A Collect. Rev. Nat. J. 2010, 2, 278–287. [Google Scholar]
- Lynch, I.; Cedervall, T.; Lundqvist, M.; Cabaleiro-Lago, C.; Linse, S.; Dawson, K.A. The nanoparticle–protein complex as a biological entity; a complex fluids and surface science challenge for the 21st century. Adv. Colloid Interface Sci. 2007, 134-135, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Chonn, A.; Semple, S.C.; Cullis, P.R. Association of blood proteins with large unilamellar liposomes in vivo. Relation to circulation lifetimes. J. Biol. Chem. 1992, 267, 18759–18765. [Google Scholar] [CrossRef]
- Aderem, A.; Underhill, D.M. Mechanisms of phagocytosis in macrophages. Annu. Rev. Immunol. 1999, 17, 593–623. [Google Scholar] [CrossRef] [PubMed]
- May, C.R.; Machesky, L.M. Phagocytosis and the actin cytoskeleton. J. Cell Sci. 2001, 114, 1061–1077. [Google Scholar] [CrossRef] [PubMed]
- Gessner, A.; Waicz, R.; Lieske, A.; Paulke, B.R.; Mäder, K.; Müller, R.H. Nanoparticles with decreasing surface hydrophobicities: Influence on plasma protein adsorption. Int. J. Pharm. 2000, 196, 245–249. [Google Scholar] [CrossRef]
- Xie, J.; Xu, C.; Kohler, N.; Hou, Y.; Sun, S. Controlled PEGylation of monodisperse Fe3O4 nanoparticles for reduced non-specific uptake by macrophage cells. Adv. Mater. 2007, 19, 3163–3166. [Google Scholar] [CrossRef]
- Zahr, S.A.; Davis, C.A.; Pishko, M.V. Macrophage uptake of core− shell nanoparticles surface modified with poly (ethylene glycol). Langmuir 2006, 22, 8178–8185. [Google Scholar] [CrossRef]
- Gao, H.; Shi, W.; Freund, L.B. Mechanics of receptor-mediated endocytosis. Proc. Natl. Acad. Sci. USA 2005, 102, 9469–9474. [Google Scholar] [CrossRef] [Green Version]
- Osaki, F.; Kanamori, T.; Sando, S.; Sera, T.; Aoyama, Y. A quantum dot conjugated sugar ball and its cellular uptake. On the size effects of endocytosis in the subviral region. J. Am. Chem. Soc. 2004, 126, 6520–6521. [Google Scholar] [CrossRef]
- Mousavi, S.A.; Malerød, L.; Berg, T.; Kjeken, R. Clathrin-dependent endocytosis. Biochem. J. 2004, 377 Pt 1, 1. [Google Scholar] [CrossRef]
- França, A.; Aggarwal, P.; Barsov, E.V.; Kozlov, S.V.; Dobrovolskaia, M.A.; González-Fernández, Á. Macrophage scavenger receptor A mediates the uptake of gold colloids by macrophages in vitro. Nanomedicine 2011, 6, 1175–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senior, J.H. Fate and behavior of liposomes in vivo: A review of controlling factors. Crit. Rev. Ther. Drug Carr. Syst. 1987, 3, 123–193. [Google Scholar]
- Carrstensen, H.; Mueller, R.H.; Müller, B.W. Particle size, surface hydrophobicity and interaction with serum of parenteral fat emulsions and model drug carriers as parameters related to RES uptake. Clin. Nutr. 1992, 11, 289–297. [Google Scholar] [CrossRef]
- Owens, D.E., III; Peppas, N.A. Opsonization, biodistribution, and pharmacokinetics of polymeric nanoparticles. Int. J. Pharm. 2006, 307, 93–102. [Google Scholar] [CrossRef]
- Mosqueira, V.C.F.; Legrand, P.; Gulik, A.; Bourdon, O.; Gref, R.; Labarre, D.; Barratt, G. Relationship between complement activation, cellular uptake and surface physicochemical aspects of novel PEG-modified nanocapsules. Biomaterials 2001, 22, 2967–2979. [Google Scholar] [CrossRef]
- Tabata, Y.; Ikada, Y. Effect of the size and surface charge of polymer microspheres on their phagocytosis by macrophage. Biomaterials 1988, 9, 356–362. [Google Scholar] [CrossRef]
- Torchilin, V.P.; Shtilman, M.I.; Trubetskoy, V.S.; Whiteman, K.; Milstein, A.M. Amphiphilic vinyl polymers effectively prolong liposome circulation time in vivo. Biochim. Biophys. Acta (BBA) Biomembr. 1994, 1195, 181–184. [Google Scholar] [CrossRef]
- Parodi, A.; Quattrocchi, N.; Van De Ven, A.L.; Chiappini, C.; Evangelopoulos, M.; Martinez, J.O.; Tasciotti, E. Synthetic nanoparticles functionalized with biomimetic leukocyte membranes possess cell-like functions. Nat. Nanotechnol. 2012, 8, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, P.L.; Harada, T.; Christian, D.A.; Pantano, D.A.; Tsai, R.K.; Discher, D.E. Minimal “Self” peptides that inhibit phagocytic clearance and enhance delivery of nanoparticles. Science 2013, 339, 971–975. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Hu, Y.; Yin, L.; Tang, C.; Yin, C. Effects of particle size and surface charge on cellular uptake and biodistribution of polymeric nanoparticles. Biomaterials 2010, 31, 3657–3666. [Google Scholar] [CrossRef]
- Sharma, G.; Valenta, D.T.; Altman, Y.; Harvey, S.; Xie, H.; Mitragotri, S.; Smith, J.W. Polymer particle shape independently influences binding and internalization by macrophages. J. Control. Release 2010, 147, 408–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilhelmi, V.; Fischer, U.; van Berlo, D.; Schulze-Osthoff, K.; Schins, R.P.; Albrecht, C. Evaluation of apoptosis induced by nanoparticles and fine particles in RAW 264.7 macrophages: Facts and artefacts. Toxicol. Vitr. 2012, 26, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, S.; de Haan, L.H.; Evers, N.M.; Jiang, X.; Marcelis, A.; Zuilhof, H.; Alink, G.M. Role of surface charge and oxidative stress in cytotoxicity of organic monolayer-coated silicon nanoparticles towards macrophage NR8383 cells. Part. Fibre Toxicol. 2010, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, J.W.; Martinez, E.; Louka, P.; Wingett, D.G. Zinc oxide nanoparticles for selective destruction of tumor cells and potential for drug delivery applications. Expert Opin. Drug Deliv. 2010, 7, 1063–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanley, C.; Thurber, A.; Hanna, C.; Punnoose, A.; Zhang, J.; Wingett, D.G. The influences of cell type and ZnO nanoparticle size on immune cell cytotoxicity and cytokine induction. Res. Lett. 2009, 4, 1409–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghimi, M.S.; Hunter, A.C.; Murray, J.C. Long-circulating and target-specific nanoparticles: Theory to practice. Pharmacol. Rev. 2001, 53, 283–318. [Google Scholar]
- Vasir, J.; Reddy, M.; Labhasetwar, V. Nanosystems in Drug Targeting: Opportunities and Challenges. Curr. Nanosci. 2005, 1, 47–64. [Google Scholar] [CrossRef]
- Weissleder, R.; Nahrendorf, M.; Pittet, M.J. Imaging macrophages with nanoparticles. Nat. Mater. 2014, 13, 125–138. [Google Scholar] [CrossRef]
- López-Dávila, V.; Seifalian, A.M.; Loizidou, M. Organic nanocarriers for cancer drug delivery. Curr. Opin. Pharmacol. 2012, 12, 414–419. [Google Scholar] [CrossRef]
- Szekanecz, Z.; Besenyei, T.; Paragh, G.; Koch, A.E. Angiogenesis in rheumatoid arthritis. Autoimmunity 2009, 42, 563–573. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, V.P.; Stylianopoulos, T.; Boucher, Y.; Jain, R.K. Delivery of molecular and nanoscale medicine to tumors: Transport barriers and strategies. Annu. Rev. Chem. Biomol. Eng. 2011, 2, 281–298. [Google Scholar] [CrossRef] [PubMed]
- Jain, K.R.; Stylianopoulos, T. Delivering nanomedicine to solid tumors. Nat. Rev. Clin. Oncol. 2010, 7, 653–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corvo, M.L.; Boerman, O.C.; Oyen, W.J.; Van Bloois, L.; Cruz, M.E.M.; Crommelin, D.J.; Storm, G. Intravenous administration of superoxide dismutase entrapped in long circulating liposomes: II. In vivo fate in a rat model of adjuvant arthritis. Biochim. Biophys. Acta (BBA)-Biomembr. 1999, 1419, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Maeda, H.; Wu, J.; Sawa, T.; Matsumura, Y.; Hori, K. Tumor vascular permeability and the EPR effect in macromolecular therapeutics: A review. J. Control. Release 2000, 65, 271–284. [Google Scholar] [CrossRef]
- Keliher, E.J.; Yoo, J.; Nahrendorf, M.; Lewis, J.S.; Marinelli, B.; Newton, A.; Weissleder, R. 89Zr-labeled dextran nanoparticles allow in vivo macrophage imaging. Bioconjugate Chem. 2011, 22, 2383–2389. [Google Scholar] [CrossRef] [Green Version]
- Sommers, S.C. Cells, Tissues, and Disease: Principles of General Pathology. JAMA 1997, 277, 1170–1171. [Google Scholar] [CrossRef]
- Medzhitov, R. Inflammation 2010: New adventures of an old flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef] [Green Version]
- Pollard, J.W. Tumour-educated macrophages promote tumour progression and metastasis. Nat. Cancer 2004, 4, 71–78. [Google Scholar] [CrossRef]
- Barrera, P.; Blom, A.; Van Lent, P.L.; Van Bloois, L.; Beijnen, J.H.; Van Rooijen, N.; Van Den Berg, W.B. Synovial macrophage depletion with clodronate-containing liposomes in rheumatoid arthritis. Arthritis Rheum. Off. J. Am. Coll. Rheumatol. 2000, 43, 1951–1959. [Google Scholar] [CrossRef] [Green Version]
- Chellat, F.; Merhi, Y.; Moreau, A. Therapeutic potential of nanoparticulate systems for macrophage targeting. Biomaterials 2005, 26, 7260–7275. [Google Scholar] [CrossRef]
- Dams, E.T.; Reijnen, M.M.; Oyen, W.J.; Boerman, O.C.; Laverman, P.; Storm, G.; Van Goor, H. Imaging experimental intraabdominal abscesses with 99mTc-PEG liposomes and 99mTc-HYNIC IgG. Ann. Surg. 1999, 229, 551. [Google Scholar] [CrossRef] [PubMed]
- Metselaar, J.M.; Wauben, M.H.; Wagenaar-Hilbers, J.P.; Boerman, O.C.; Storm, G. Complete remission of experimental arthritis by joint targeting of glucocorticoids with long-circulating liposomes. Arthritis Rheum. Off. J. Am. Coll. Rheumatol. 2003, 48, 2059–2066. [Google Scholar] [CrossRef] [PubMed]
- Byrne, D.J.; Betancourt, T.; Brannon-Peppas, L. Active targeting schemes for nanoparticle systems in cancer therapeutics. Adv. Drug Deliv. Rev. 2008, 60, 1615–1626. [Google Scholar] [CrossRef] [PubMed]
- Koo, O.M.Y.; Rubinstein, I.; Önyüksel, H. Actively targeted low-dose camptothecin as a safe, long-acting, disease-modifying nanomedicine for rheumatoid arthritis. Pharm. Res. 2010, 28, 776–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, M.; Okano, T. Targetable drug carriers: Present status and a future perspective. Adv. Drug Deliv. Rev. 1996, 21, 77–80. [Google Scholar] [CrossRef]
- Nie, S.; Xing, Y.; Kim, G.J.; Simons, J.W. Nanotechnology applications in cancer. Annu. Rev. Biomed. Eng. 2007, 9, 257–288. [Google Scholar] [CrossRef] [Green Version]
- Nobs, L.; Buchegger, F.; Gurny, R.; Allémann, E. Current methods for attaching targeting ligands to liposomes and nanoparticles. J. Pharm. Sci. 2004, 93, 1980–1992. [Google Scholar] [CrossRef]
- Ulbrich, H.; Eriksson, E.E.; Lindbom, L. Leukocyte and endothelial cell adhesion molecules as targets for therapeutic interventions in inflammatory disease. Trends Pharmacol. Sci. 2003, 24, 640–647. [Google Scholar] [CrossRef]
- Rodrigues, S.; Grenha, A. Activation of Macrophages: Establishing a Role for Polysaccharides in Drug Delivery Strategies Envisaging Antibacterial Therapy. Curr. Pharm. Des. 2015, 21, 4869–4887. [Google Scholar] [CrossRef]
- Vasselon, T.; Detmers, P.A. Toll Receptors: A Central Element in Innate Immune Responses. Infect. Immun. 2002, 70, 1033–1041. [Google Scholar] [CrossRef] [Green Version]
- Doyle, S.L.; O’Neill, L. Toll-like receptors: From the discovery of NFκB to new insights into transcriptional regulations in innate immunity. Biochem. Pharmacol. 2006, 72, 1102–1113. [Google Scholar] [CrossRef] [PubMed]
- Beutler, B.; Jiang, Z.; Georgel, P.; Crozat, K.; Croker, B.; Rutschmann, S.; Du, X.; Hoebe, K. Genetic analysis of host resistance: Toll-like receptor Signaling and Immunity at Large. Annu. Rev. Immunol. 2006, 24, 353–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peiser, L.; Mukhopadhyay, S.; Gordon, S. Scavenger receptors in innate immunity. Curr. Opin. Immunol. 2002, 14, 123–128. [Google Scholar] [CrossRef]
- Geijtenbeek, T.B.H.; Gringhuis, S.I. Signalling through C-type lectin receptors: Shaping immune responses. Nat. Rev. Immunol. 2009, 9, 465–479. [Google Scholar] [CrossRef]
- Herwald, H.; Egesten, A. Macrophages: Past, present and future. J. Innate Immun. 2013, 5, 657. [Google Scholar] [CrossRef]
- Ishii, M.; Kojima, N. Mucosal adjuvant activity of oligomannose-coated liposomes for nasal immunization. Glycoconj. J. 2009, 27, 115–123. [Google Scholar] [CrossRef]
- Nimje, N.; Agarwal, A.; Saraogi, G.K.; Lariya, N.; Rai, G.; Agrawal, H.; Agrawal, G.P. Mannosylated nanoparticulate carriers of rifabutin for alveolar targeting. J. Drug Target. 2009, 17, 777–787. [Google Scholar] [CrossRef]
- Chono, S.; Tanino, T.; Seki, T.; Morimoto, K. Efficient drug targeting to rat alveolar macrophages by pulmonary administration of ciprofloxacin incorporated into mannosylated liposomes for treatment of respiratory intracellular parasitic infections. J. Control. Release 2008, 127, 50–58. [Google Scholar] [CrossRef]
- Ma, C.; Wu, M.; Ye, W.; Huang, Z.; Ma, X.; Wang, W.; Wu, C. Inhalable solid lipid nanoparticles for intracellular tuberculosis infection therapy: Macrophage-targeting and pH-sensitive properties. Drug Deliv. Transl. Res. 2021, 11, 218–1235. [Google Scholar] [CrossRef]
- Lipinski, M.J.; Frias, J.C.; Amirbekian, V.; Briley-Saebo, K.C.; Mani, V.; Samber, D.; Fayad, Z.A. Macrophage-specific lipid-based nanoparticles improve cardiac magnetic resonance detection and characterization of human atherosclerosis. JACC Cardiovasc. Imaging 2009, 2, 637–647. [Google Scholar] [CrossRef] [Green Version]
- Jaracz, S.; Chen, J.; Kuznetsova, L.V.; Ojima, I. Recent advances in tumor-targeting anticancer drug conjugates. Bioorganic. Med. Chem. 2005, 13, 5043–5054. [Google Scholar] [CrossRef] [PubMed]
- Kamat, M.; El-Boubbou, K.; Zhu, D.C.; Lansdell, T.; Lu, X.; Li, W.; Huang, X. Hyaluronic acid immobilized magnetic nanoparticles for active targeting and imaging of macrophages. Bioconjugate Chem. 2010, 21, 2128–2135. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.Y.; Chen, C.W.; Chen, K.H.; Chiu, H.T.; Huang, Y.F. Hyaluronic Acid/Doxorubicin/Gold Nanorods-Anchored Macrophage for Combined Dug Delivery and Photothermal Therapy; National Tsing Hua University: Hsinchu, Taiwan.
- Gagné, J.F.; Désormeaux, A.; Perron, S.; Tremblay, M.J.; Bergeron, M.G. Targeted delivery of indinavir to HIV-1 primary reservoirs with immunoliposomes. Biochim. Biophys. Acta (BBA) Biomembr. 2001, 1558, 198–210. [Google Scholar] [CrossRef] [Green Version]
- Chavez-Santoscoy, A.V.; Roychoudhury, R.; Pohl, N.L.; Wannemuehler, M.J.; Narasimhan, B.; Ramer-Tait, A.E. Tailoring the immune response by targeting C-type lectin receptors on alveolar macrophages using “pathogen-like” amphiphilic polyanhydride nanoparticles. Biomaterials 2012, 33, 4762–4772. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, U.; Inubushi, R.; Uyama, H.; Uematsu, T.; Kuwabata, S.; van der Vlies, A.J. Mannose-displaying fluorescent framboidal nanoparticles containing phenylboronic acid groups as a potential drug carrier for macrophage targeting. Colloids Surfaces B Biointerfaces 2015, 136, 1174–1181. [Google Scholar] [CrossRef]
- Ricketts, C.R. Dextran sulphate—A synthetic analogue of heparin. Biochem. J. 1952, 51, 129. [Google Scholar] [CrossRef] [Green Version]
- Chao, Y.; Makale, M.; Karmali, P.P.; Sharikov, Y.; Tsigelny, I.; Merkulov, S.; Simberg, D. Recognition of dextran–superparamagnetic iron oxide nanoparticle conjugates (Feridex) via macrophage scavenger receptor charged domains. Bioconjugate Chem. 2012, 23, 1003–1009. [Google Scholar] [CrossRef] [Green Version]
- De Villiers, W.J.; Westherry, K.; Kang, Y.; Lee, E.; Varilek, G. Macrophage scavenger receptor type A attenuates the development of dextran sodium sulfate-induced colitis. Gastroenterology 2001, 5, A183. [Google Scholar] [CrossRef]
- Limmon, G.V.; Arredouani, M.; McCann, K.L.; Minor, R.A.C.; Kobzik, L.; Imani, F. Scavenger receptor class-A is a novel cell surface receptor for double-stranded RNA. FASEB J. 2008, 22, 159–167. [Google Scholar] [CrossRef]
- Thelen, T.; Hao, Y.; Medeiros, A.I.; Curtis, J.L.; Serezani, C.H.; Kobzik, L.; Aronoff, D.M. The class A scavenger receptor, macrophage receptor with collagenous structure, is the major phagocytic receptor for Clostridium sordellii expressed by human decidual macrophages. J. Immunol. 2010, 187, 4328–4335. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, J.R.; Jaffer, F.; Weissleder, R. A Macrophage-Targeted Theranostic Nanoparticle for Biomedical Applications. Small 2006, 2, 983–987. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Gou, J.; Sun, W.; Tao, X.; Tan, X.; Wang, P.; Tang, X. Entrapping of nanoparticles in yeast cell wall microparticles for macrophage-targeted oral delivery of cabazitaxel. Mol. Pharm. 2018, 15, 2870–2882. [Google Scholar] [CrossRef] [PubMed]
- Soto, E.R.; Caras, A.C.; Kut, L.C.; Castle, M.K.; Ostroff, G.R. Glucan particles for macrophage targeted delivery of nanoparticles. J. Drug Deliv. 2011, 2012, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poetsch, A.; Wolters, D. Bacterial membrane proteomics. Proteomics 2008, 8, 4100–4122. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.Y.; Bang, J.Y.; Park, G.W.; Choi, D.S.; Kang, J.S.; Kim, H.J.; Gho, Y.S. Global proteomic profiling of native outer membrane vesicles derived from Escherichia coli. Proteomics 2007, 7, 3143–3153. [Google Scholar] [CrossRef]
- Kuehn, M.J.; Kesty, N.C. Bacterial outer membrane vesicles and the host–pathogen interaction. Genes Dev. 2005, 19, 2645–2655. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Lee, J.; Park, J.; Gho, Y.S. Gram-negative and Gram-positive bacterial extracellular vesicles. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2015; Volume 40. [Google Scholar]
- Gao, F.; Xu, L.; Yang, B.; Fan, F.; Yang, L. Kill the real with the fake: Eliminate intracellular Staphylococcus aureus using nanoparticle coated with its extracellular vesicle membrane as active-targeting drug carrier. ACS Infect. Dis. 2018, 5, 218–227. [Google Scholar] [CrossRef]
- Kasuya, T.; Nomura, S.; Matsuzaki, T.; Jung, J.; Yamada, T.; Tatematsu, K.; Kuroda, S.I. Expression of squamous cell carcinoma antigen-1 in liver enhances the uptake of hepatitis B virus envelope-derived bio-nanocapsules in transgenic rats. FEBS J. 2008, 275, 5714–5724. [Google Scholar] [CrossRef]
- Kurata, N.; Shishido, T.; Muraoka, M.; Tanaka, T.; Ogino, C.; Fukuda, H.; Kondo, A. Specific protein delivery to target cells by antibody-displaying bionanocapsules. J. Biochem. 2008, 144, 701–707. [Google Scholar] [CrossRef]
- Iijima, M.; Matsuzaki, T.; Kadoya, H.; Hatahira, S.; Hiramatsu, S.; Jung, G.; Kuroda, S.I. Bionanocapsule-based enzyme–antibody conjugates for enzyme-linked immunosorbent assay. Anal. Biochem. 2010, 396, 257–261. [Google Scholar] [CrossRef]
- Tsutsui, Y.; Tomizawa, K.; Nagita, M.; Michiue, H.; Nishiki, T.I.; Ohmori, I.; Matsui, H. Development of bionanocapsules targeting brain tumors. J. Control. Release 2007, 122, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Erntell, M.; Myhre, E.B.; Sjöbring, U.; Björck, L. Streptococcal protein G has affinity for both Fab-and Fc-fragments of human IgG. Mol. Immunol. 1988, 25, 121–126. [Google Scholar] [CrossRef]
- Van Der Heijden, J.W.; Oerlemans, R.; Dijkmans, B.A.; Qi, H.; Laken, C.J.V.D.; Lems, W.F.; Jansen, G. Folate receptor β as a potential delivery route for novel folate antagonists to macrophages in the synovial tissue of rheumatoid arthritis patients. Arthritis Rheum. 2009, 60, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Paulos, C.M.; Turk, M.J.; Breur, G.J.; Low, P.S. Folate receptor-mediated targeting of therapeutic and imaging agents to activated macrophages in rheumatoid arthritis. Adv. Drug Deliv. Rev. 2004, 56, 1205–1217. [Google Scholar] [CrossRef]
- Thomas, T.P.; Goonewardena, S.N.; Majoros, I.J.; Kotlyar, A.; Cao, Z.; Leroueil, P.R.; Baker, J.R., Jr. Folate-targeted nanoparticles show efficacy in the treatment of inflammatory arthritis. Arthritis Care Res. 2011, 63, 2671–2680. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Tsai, Y.T.; Weng, H.; Baker, D.W.; Tang, L. Real time monitoring of biomaterial-mediated inflammatory responses via macrophage-targeting NIR nanoprobes. Biomaterials 2011, 32, 9383–9390. [Google Scholar] [CrossRef] [Green Version]
- Penn, C.A.; Yang, K.; Zong, H.; Lim, J.Y.; Cole, A.; Yang, D.; Buckanovich, R.J. Therapeutic impact of nanoparticle therapy targeting tumor-associated macrophages. Mol. Cancer Ther. 2017, 17, 96–106. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.K.; Jaiswal, A.K.; Pawar, V.K.; Raval, K.; Kumar, A.; Bora, H.K.; Chourasia, M.K. Fabrication of 3-O-sn-phosphatidyl-L-serine anchored PLGA nanoparticle bearing amphotericin B for macrophage targeting. Pharm. Res. 2018, 35, 60. [Google Scholar] [CrossRef]
- García-Suárez, O.; Hannestad, J.; Esteban, I.; Sainz, R.; Naves, F.J.; Vega, J.A. Expression of the TrkB neurotrophin receptor by thymic macrophages. Immunology 1998, 94, 235–241. [Google Scholar] [CrossRef]
- Ma, Z.; Wu, X.; Cao, M.; Pan, W.; Zhu, F.; Chen, J.; Qi, Z. Selection of trkB-binding peptides from a phage-displayed random peptide library. Sci. China Ser. C Life Sci. 2003, 46, 77–86. [Google Scholar] [CrossRef]
- Talvitie, E.; Leppiniemi, J.; Mikhailov, A.; Hytönen, V.P.; Kellomäki, M. Peptide-functionalized chitosan–DNA nanoparticles for cellular targeting. Carbohydr. Polym. 2012, 89, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Sebollela, A.; Cagliari, T.C.; Limaverde, G.S.; Chapeaurouge, A.; Sorgine, M.H.; Coelho-Sampaio, T.; Ferreira, S.T. Heparin-binding sites in granulocyte-macrophage colony-stimulating factor: Localization and regulation by histidine ionization. J. Biol. Chem. 2005, 280, 31949–31956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falcone, D.J. Heparin stimulation of plasminogen activator secretion by macrophage-like cell line RAW264. 7: Role of the scavenger receptor. J. Cell. Physiol. 1989, 140, 219–226. [Google Scholar] [CrossRef]
- Lindstedt, K.; Kokkonen, J.; Kovanen, P. Soluble heparin proteoglycans released from stimulated mast cells induce uptake of low density lipoproteins by macrophages via scavenger receptor-mediated phagocytosis. J. Lipid Res. 1992, 33, 65–75. [Google Scholar] [CrossRef]
- Akinrinmade, O.A.; Chetty, S.; Daramola, A.K.; Islam, M.U.; Thepen, T.; Barth, S. CD64: An attractive immunotherapeutic target for M1-type macrophage mediated chronic inflammatory diseases. Biomedicines 2017, 5, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hristodorov, D.; Mladenov, R.; von Felbert, V.; Huhn, M.; Fischer, R.; Barth, S.; Thepen, T. Targeting CD64 mediates elimination of M1 but not M2 macrophages in vitro and in cutaneous inflammation in mice and patient biopsies. MAbs 2015, 7, 853–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thepen, T.; Van Vuuren, A.J.; Kiekens, R.; Damen, C.A.; Vooijs, W.C.; van de Winkel, J.G. Resolution of cutaneous inflammation after local elimination of macrophages. Nat. Biotechnol. 2000, 18, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Wallace, P.K.; Keler, T.; Coleman, K.; Fisher, J.; Vitale, L.; Graziano, R.F.; Fanger, M.W. Humanized mAb H22 binds the human high affinity Fc receptor for IgG (FcγRI), blocks phagocytosis, and modulates receptor expression. J. Leukoc. Biol. 1997, 62, 469–479. [Google Scholar] [CrossRef]
- Ericson, S.G.; Coleman, K.D.; Wardwell, K.; Baker, S.; Fanger, M.W.; Guyre, P.M.; Ely, P. Monoclonal antibody 197 (anti-FcγRI) infusion in a patient with immune thrombocytopenia purpura (ITP) results in down-modulation of FcγRI on circulating monocytes. Br. J. Haematol. 1996, 92, 718–724. [Google Scholar] [CrossRef]
- Guyre, P.M.; Graziano, R.F.; Vance, B.A.; Morganelli, P.M.; Fanger, M.W. Monoclonal antibodies that bind to distinct epitopes on Fc gamma RI are able to trigger receptor function. J. Immunol. 1989, 143, 1650–1655. [Google Scholar]
- Graziano, R.F.; Tempest, P.R.; White, P.; Keler, T.; Deo, Y.; Ghebremariam, H.; Guyre, P.M. Construction and characterization of a humanized anti-gamma-Ig receptor type I (Fc gamma RI) monoclonal antibody. J. Immunol. 1995, 155, 4996–5002. [Google Scholar] [PubMed]
- Wang, W.; Li, F.; Li, S.; Hu, Y.; Xu, M.; Zhang, Y.; Qiu, B. M2 macrophage-targeted iron oxide nanoparticles for magnetic resonance image-guided magnetic hyperthermia therapy. J. Mater. Sci. Technol. 2021, 81, 77–87. [Google Scholar] [CrossRef]
- Conde, J.; Bao, C.; Tan, Y.; Cui, D.; Edelman, E.R.; Azevedo, H.S.; Tian, F. Dual targeted immunotherapy via in vivo delivery of biohybrid RNAi-peptide nanoparticles to tumor-associated macrophages and cancer cells. Adv. Funct. Mater. 2015, 25, 4183–4194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cieslewicz, M.; Tang, J.; Jonathan, L.Y.; Cao, H.; Zavaljevski, M.; Motoyama, K.; Pun, S.H. Targeted delivery of proapoptotic peptides to tumor-associated macrophages improves survival. Proc. Natl. Acad. Sci. USA 2013, 110, 15919–15924. [Google Scholar] [CrossRef] [Green Version]
- Ngambenjawong, C.; Gustafson, H.H.; Pineda, J.M.; Kacherovsky, N.A.; Cieslewicz, M.; Pun, S.H. Serum stability and affinity optimization of an M2 macrophage-targeting peptide (M2pep). Theranostics 2016, 6, 1403–1414. [Google Scholar] [CrossRef]
- Han, S.; Wang, W.; Wang, S.; Yang, T.; Zhang, G.; Wang, D.; Wang, L. Tumor microenvironment remodeling and tumor therapy based on M2-like tumor associated macrophage-targeting nano-complexes. Theranostics 2021, 11, 2892–2916. [Google Scholar] [CrossRef]
- Schmitz, S.A.; Taupitz, M.; Wagner, S.; Wolf, K.J.; Beyersdorff, D.; Hamm, B. Magnetic resonance imaging of atherosclerotic plaques using superparamagnetic iron oxide particles. J. Magn. Reson. Imaging Off. J. Int. Soc. Magn. Reson. Med. 2001, 14, 355–361. [Google Scholar] [CrossRef]
- Kooi, M.E.; Cappendijk, V.C.; Cleutjens, K.B.J.M.; Kessels, A.G.H.; Kitslaar, P.J.E.H.M.; Borgers, M.; Van Engelshoven, J.M.A. Accumulation of ultrasmall superparamagnetic particles of iron oxide in human atherosclerotic plaques can be detected by in vivo magnetic resonance imaging. Circulation 2003, 107, 2453–2458. [Google Scholar] [CrossRef] [Green Version]
- Morishige, K.; Kacher, D.F.; Libby, P.; Josephson, L.; Ganz, P.; Weissleder, R.; Aikawa, M. High-resolution magnetic resonance imaging enhanced with superparamagnetic nanoparticles measures macrophage burden in atherosclerosis. Circulation 2010, 122, 1707–1715. [Google Scholar] [CrossRef] [Green Version]
- Terashima, M.; Uchida, M.; Kosuge, H.; Tsao, P.S.; Young, M.J.; Conolly, S.M.; McConnell, M.V. Human ferritin cages for imaging vascular macrophages. Biomaterials 2011, 32, 1430–1437. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.; Wang, Y.; Zhang, R.; Gao, Q.; Liang, X.; Gao, L.; Cao, F. MRI/optical dual-modality imaging of vulnerable atherosclerotic plaque with an osteopontin-targeted probe based on Fe3O4 nanoparticles. Biomaterials 2016, 112, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Li, X.; Zhang, C.; Tan, H.; Wang, C.; Pang, L.; Shi, H. Detection of vulnerable atherosclerosis plaques with a dual-modal single-photon-emission computed tomography/magnetic resonance imaging probe targeting apoptotic macrophages. ACS Appl. Mater. Interfaces 2015, 7, 2847–2855. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, J.; Wu, M.; He, Y.; Zhang, H.; Xu, G.; Zhang, X. Macrophage-targeted and clearable glutathione-based MRI nanoprobes for atherosclerosis molecular imaging. J. Nanoparticle Res. 2019, 21, 231. [Google Scholar] [CrossRef]
- Ding, J.; Wang, Y.; Ma, M.; Zhang, Y.; Lu, S.; Jiang, Y.; Gu, N. CT/fluorescence dual-modal nanoemulsion platform for investigating atherosclerotic plaques. Biomaterials 2013, 34, 209–216. [Google Scholar] [CrossRef]
- Keliher, E.J.; Ye, Y.X.; Wojtkiewicz, G.R.; Aguirre, A.D.; Tricot, B.; Senders, M.L.; Nahrendorf, M. Polyglucose nanoparticles with renal elimination and macrophage avidity facilitate PET imaging in ischaemic heart disease. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef]
- Miselis, N.R.; Wu, Z.J.; Van Rooijen, N.; Kane, A.B. Targeting tumor-associated macrophages in an orthotopic murine model of diffuse malignant mesothelioma. Mol. Cancer Ther. 2008, 7, 788–799. [Google Scholar] [CrossRef] [Green Version]
- Orzechowska, B.; Chaber, R.; Wiśniewska, A.; Pajtasz-Piasecka, E.; Jatczak, B.; Siemieniec, I.; Błach-Olszewska, Z. Baicalin from the extract of Scutellaria baicalensis affects the innate immunity and apoptosis in leukocytes of children with acute lymphocytic leukemia. Int. Immunopharmacol. 2014, 23, 558–567. [Google Scholar] [CrossRef]
- Shan, H.; Dou, W.; Zhang, Y.; Qi, M. Targeted ferritin nanoparticle encapsulating CpG oligodeoxynucleotides induces tumor-associated macrophage M2 phenotype polarization into M1 phenotype and inhibits tumor growth. Nanoscale 2020, 12, 22268–22280. [Google Scholar] [CrossRef]
- Opanasopit, P.; Sakai, M.; Nishikawa, M.; Kawakami, S.; Yamashita, F.; Hashida, M. Inhibition of liver metastasis by targeting of immunomodulators using mannosylated liposome carriers. J. Control. Release 2002, 80, 283–294. [Google Scholar] [CrossRef]
- Aniyama, T.; Holden, H.T. Direct augmentation of cytolytic activity of tumor-derived macrophages and macrophage cell lines by muramyl dipeptide. Cell. Immunol. 1979, 48, 369–374. [Google Scholar] [CrossRef]
- Conejo-Garcia, J.R.; Benencia, F.; Courreges, M.C.; Kang, E.; Mohamed-Hadley, A.; Buckanovich, R.J.; Coukos, G. Tumor-infiltrating dendritic cell precursors recruited by a β-defensin contribute to vasculogenesis under the influence of Vegf-A. Nat. Med. 2004, 10, 950–958. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, D.K.; Diagaradjane, P.; Krishnan, S. Nanoparticle-mediated hyperthermia in cancer therapy. Ther. Deliv. 2011, 2, 1001–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IIssels, R.D. Hyperthermia adds to chemotherapy. Eur. J. Cancer 2008, 44, 2546–2554. [Google Scholar] [CrossRef] [PubMed]
- Nawroth, I.; Alsner, J.; Deleuran, B.W.; Dagnaes-Hansen, F.; Yang, C.; Horsman, M.R.; Gao, S. Peritoneal macrophages mediated delivery of chitosan/siRNA nanoparticle to the lesion site in a murine radiation-induced fibrosis model. Acta Oncol. 2012, 52, 1730–1738. [Google Scholar] [CrossRef] [Green Version]
- Vergne, I.; Chua, J.; Singh, S.B.; Deretic, V. Cell biology of Mycobacterium tuberculosis phagosome. Annu. Rev. Cell Dev. Biol. 2004, 20, 367–394. [Google Scholar] [CrossRef]
- Hunter, S.W.; Brennan, P.J. Evidence for the presence of a phosphatidylinositol anchor on the lipoarabinomannan and lipomannan of Mycobacterium tuberculosis. J. Biol. Chem. 1990, 265, 9272–9279. [Google Scholar] [CrossRef]
- Saraogi, G.K.; Sharma, B.; Joshi, B.; Gupta, P.; Gupta, U.D.; Jain, N.K.; Agrawal, G.P. Mannosylated gelatin nanoparticles bearing isoniazid for effective management of tuberculosis. J. Drug Target. 2010, 19, 219–227. [Google Scholar] [CrossRef]
- Maretti, E.; Costantino, L.; Rustichelli, C.; Leo, E.; Croce, M.A.; Buttini, F.; Iannuccelli, V. Surface engineering of Solid Lipid Nanoparticle assemblies by methyl α-d-mannopyranoside for the active targeting to macrophages in anti-tuberculosis inhalation therapy. Int. J. Pharm. 2017, 528, 440–451. [Google Scholar] [CrossRef]
- Maretti, E.; Costantino, L.; Buttini, F.; Rustichelli, C.; Leo, E.; Truzzi, E.; Iannuccelli, V. Newly synthesized surfactants for surface mannosylation of respirable SLN assemblies to target macrophages in tuberculosis therapy. Drug Deliv. Transl. Res. 2018, 9, 298–310. [Google Scholar] [CrossRef]
- Costa, A.; Sarmento, B.; Seabra, V. Mannose-functionalized solid lipid nanoparticles are effective in targeting alveolar macrophages. Eur. J. Pharm. Sci. 2018, 114, 103–113. [Google Scholar] [CrossRef]
- Tiwari, S.; Chaturvedi, A.P.; Tripathi, Y.B.; Mishra, B. Macrophage-specific targeting of isoniazid through mannosylated gelatin microspheres. AAPS PharmSciTech 2011, 12, 900–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Sharma, S.; Khuller, G.K. Lectin-functionalized poly (lactide-co-glycolide) nanoparticles as oral/aerosolized antitubercular drug carriers for treatment of tuberculosis. J. Antimicrob. Chemother. 2004, 54, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Asthana, S.; Jaiswal, A.K.; Gupta, P.K.; Dube, A.; Chourasia, M.K. Th-1 biased immunomodulation and synergistic antileishmanial activity of stable cationic lipid–polymer hybrid nanoparticle: Biodistribution and toxicity assessment of encapsulated amphotericin B. Eur. J. Pharm. Biopharm. 2015, 89, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Jain, K.; Verma, A.K.; Mishra, P.R.; Jain, N.K. Surface-engineered dendrimeric nanoconjugates for macrophage-targeted delivery of amphotericin B: Formulation development and in vitro and in vivo evaluation. Antimicrob. Agents Chemother. 2015, 59, 2479–2487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundar, S.; More, D.K.; Singh, M.K.; Singh, V.P.; Sharma, S.; Makharia, A.; Murray, H.W. Failure of pentavalent antimony in visceral leishmaniasis in India: Report from the center of the Indian epidemic. Clin. Infect. Dis. 2000, 31, 1104–1107. [Google Scholar] [CrossRef] [Green Version]
- Boelaert, M.; Criel, B.; Leeuwenburg, J.; Van Damme, W.; Le Ray, D.; Van Der Stuyft, P. Visceral leishmaniasis control: A public health perspective. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 465–471. [Google Scholar] [CrossRef]
- Shahnaz, G.; Edagwa, B.J.; McMillan, J.; Akhtar, S.; Raza, A.; Qureshi, N.A.; Gendelman, H.E. Development of mannose-anchored thiolated amphotericin B nanocarriers for treatment of visceral leishmaniasis. Nanomedicine 2017, 12, 99–115. [Google Scholar] [CrossRef]
- Narayanasamy, P.; Eoh, H.; Brennan, P.J.; Crick, D.C. Synthesis of 4-diphosphocytidyl-2-C-methyl-D-erythritol 2-phosphate and kinetic studies of Mycobacterium tuberculosis IspF. Chem. Biol. 2010, 17, 117–122. [Google Scholar] [CrossRef] [Green Version]
- Drobniewski, F. Is death inevitable with multiresistant TB plus HIV infection? Lancet 1997, 349, 71–72. [Google Scholar] [CrossRef]
- Tan, J.; Sattentau, Q.J. The HIV-1-containing macrophage compartment: A perfect cellular niche? Trends Microbiol. 2013, 21, 405–412. [Google Scholar] [CrossRef]
- Batrakova, V.E.; Gendelman, H.E.; Kabanov, A.V. Cell-mediated drug delivery. Expert Opin. Drug Deliv. 2011, 8, 415–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brynskikh, A.M.; Zhao, Y.; Mosley, R.L.; Li, S.; Boska, M.D.; Klyachko, N.L.; Batrakova, E.V. Macrophage delivery of therapeutic nanozymes in a murine model of Parkinson’s disease. Nanomedicine 2010, 5, 379–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, W.; Hilgenbrink, A.R.; Matteson, E.L.; Lockwood, M.B.; Cheng, J.X.; Low, P.S. A functional folate receptor is induced during macrophage activation and can be used to target drugs to activated macrophages. Blood J. Am. Soc. Hematol. 2009, 113, 438–446. [Google Scholar] [CrossRef] [Green Version]
- Narayanasamy, P.; Switzer, B.L.; Britigan, B.E. Prolonged-acting, Multi-targeting Gallium Nanoparticles Potently Inhibit Growth of Both HIV and Mycobacteria in Co-Infected Human Macrophages. Sci. Rep. 2015, 5, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergstrøm, B.; Aune, M.H.; Awuh, J.A.; Kojen, J.F.; Blix, K.J.; Ryan, L.; Stenvik, J. TLR8 senses Staphylococcus aureus RNA in human primary monocytes and macrophages and induces IFN-β production via a TAK1–IKKβ–IRF5 signaling pathway. J. Immunol. 2015, 195, 1100–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levinson, W. Review of Medical Microbiology and Immunology; McGraw-Hill Education: New York, NY, USA, 2014. [Google Scholar]
- Kim, B.; Pang, H.B.; Kang, J.; Park, J.H.; Ruoslahti, E.; Sailor, M.J. Immunogene therapy with fusogenic nanoparticles modulates macrophage response to Staphylococcus aureus. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Courties, G.; Heidt, T.; Sebas, M.; Iwamoto, Y.; Jeon, D.; Truelove, J.; Nahrendorf, M. In vivo silencing of the transcription factor IRF5 reprograms the macrophage phenotype and improves infarct healing. J. Am. Coll. Cardiol. 2013, 63, 1556–1566. [Google Scholar] [CrossRef]
- Hall, J.L.; Wei, L.-N. Could Silencing IRF5 Improve Healing of a Myocardial Infarct Through the Reprogramming of the Macrophage Population? J. Am. Coll. Cardiol. 2013, 63, 1567–1568. [Google Scholar] [CrossRef] [Green Version]
- Weiss, M.; Byrne, A.J.; Blazek, K.; Saliba, D.G.; Pease, J.E.; Perocheau, D.; Udalova, I.A. IRF5 controls both acute and chronic inflammation. Proc. Natl. Acad. Sci. USA 2015, 112, 11001–11006. [Google Scholar] [CrossRef] [Green Version]
- Dobos, K.M.; Swiderek, K.; Khoo, K.H.; Brennan, P.J.; Belisle, J.T. Evidence for glycosylation sites on the 45-kilodalton glycoprotein of Mycobacterium tuberculosis. Infect. Immun. 1995, 63, 2846–2853. [Google Scholar] [CrossRef] [Green Version]
- Hartley, J.L.; Adams, G.A.; Tornabene, T.G. Chemical and Physical Properties of Lipopolysaccharide of Yersinia pestis. J. Bacteriol. 1974, 118, 848–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.J.; Fraser, B.A.; Szu, S.; Lin, K.T. Chemical structure of and immune response to polysaccharides of Streptococcus pneumoniae. Clin. Infect. Dis. 1981, 3, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.; Yoshima, H.; Kobata, A. Carbohydrates of influenza virus hemagglutinin: Structures of the whole neutral sugar chains. Biochemistry 1983, 22, 188–196. [Google Scholar] [CrossRef]
- Ao, Z.; Wang, L.; Mendoza, E.J.; Cheng, K.; Zhu, W.; Cohen, E.A.; Yao, X. Incorporation of Ebola glycoprotein into HIV particles facilitates dendritic cell and macrophage targeting and enhances HIV-specific immune responses. PLoS ONE 2019, 14, e0216949. [Google Scholar] [CrossRef] [PubMed]
- Kneser, U.; Schaefer, D.J.; Polykandriotis, E.; Horch, R.E. Tissue engineering of bone: The reconstructive surgeon’s point of view. J. Cell. Mol. Med. 2006, 10, 7–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, L.; Liu, B.; Liu, G.; Zhang, W.; Cen, L.; Sun, J.; Cao, Y. Repair of cranial bone defects with adipose derived stem cells and coral scaffold in a canine model. Biomaterials 2007, 28, 5477–5486. [Google Scholar] [CrossRef]
- Bueno, E.M.; Glowacki, J. Cell-free and cell-based approaches for bone regeneration. Nat. Rev. Rheumatol. 2009, 5, 685–697. [Google Scholar] [CrossRef]
- Wang, B.; Guo, Y.; Chen, X.; Zeng, C.; Hu, Q.; Yin, W.; Yu, F. Nanoparticle-modified chitosan-agarose-gelatin scaffold for sustained release of SDF-1 and BMP-2. Int. J. Nanomed. 2018, 13, 7395–7408. [Google Scholar] [CrossRef] [Green Version]
- Takayanagi, H. Osteoimmunology: Shared mechanisms and crosstalk between the immune and bone systems. Nat. Rev. Immunol. 2007, 7, 292–304. [Google Scholar] [CrossRef]
- Franz, S.; Rammelt, S.; Scharnweber, D.; Simon, J.C. Immune responses to implants–a review of the implications for the design of immunomodulatory biomaterials. Biomaterials 2011, 32, 6692–6709. [Google Scholar] [CrossRef]
- Takayanagi, H. Inflammatory bone destruction and osteoimmunology. J. Periodontal Res. 2005, 40, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yuen, J.; Crawford, R.; Chang, J.; Wu, C.; Xiao, Y. The effect of osteoimmunomodulation on the osteogenic effects of cobalt incorporated β-tricalcium phosphate. Biomaterials 2015, 61, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, M.; Mukai, T.; Otsuka, F.; Yamashita, M.; Takasugi, K.; Makino, H. Inhibition of bone morphogenetic protein-induced osteoblast differentiation by tumor necrosis factor-a. Ann. Rheum. Dis. 2007, 66, 155. [Google Scholar]
- Yamashita, M.; Otsuka, F.; Mukai, T.; Otani, H.; Inagaki, K.; Miyoshi, T.; Makino, H. Simvastatin antagonizes tumor necrosis factor-α inhibition of bone morphogenetic proteins-2-induced osteoblast differentiation by regulating Smad signaling and Ras/Rho-mitogen-activated protein kinase pathway. J. Endocrinol. 2008, 196, 601–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caetano-Lopes, J.; Canhao, H.; Fonseca, J.E. Osteoimmunology—the hidden immune regulation of bone. Autoimmun. Rev. 2009, 8, 250–255. [Google Scholar]
- Freytes, D.O.; Wan, L.; Vunjak-Novakovic, G. Geometry and force control of cell function. J. Cell. Biochem. 2009, 108, 1047–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schett, G. Osteoimmunology in rheumatic diseases. Arthritis Res. Ther. 2009, 11, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Ni, S.; Han, S.; Crawford, R.; Lu, S.; Wei, F.; Xiao, Y. Nanoporous microstructures mediate osteogenesis by modulating the osteo-immune response of macrophages. Nanoscale 2016, 9, 706–718. [Google Scholar] [CrossRef]
- Yang, C.; Zhao, C.; Wang, X.; Shi, M.; Zhu, Y.; Jing, L.; Chang, J. Stimulation of osteogenesis and angiogenesis by micro/nano hierarchical hydroxyapatite via macrophage immunomodulation. Nanoscale 2019, 11, 17699–17708. [Google Scholar] [CrossRef]
- Hotchkiss, K.M.; Reddy, G.B.; Hyzy, S.L.; Schwartz, Z.; Boyan, B.D.; Olivares-Navarrete, R. Titanium surface characteristics, including topography and wettability, alter macrophage activation. Acta Biomater. 2016, 31, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Hughes, E.J.; Srinivasan, S.R.; Hedrick, C.C. Sphingosine-1-Phosphate Induces an Anti-Inflammatory Phenotype in Macrophages. Circulation research. 2008, 102, 950–958. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Segar, C.E.; Hughley, B.B.; Bowers, D.T.; Botchwey, E.A. The promotion of mandibular defect healing by the targeting of S1P receptors and the recruitment of alternatively activated macrophages. Biomaterials 2013, 34, 9853–9862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, C.; Zhao, Q.; Li, W.; Zhao, Z.; Wang, J.; Deng, T.; Zhang, Y. Biomimetic anti-inflammatory nano-capsule serves as a cytokine blocker and M2 polarization inducer for bone tissue repair. Acta Biomater. 2019, 102, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Feng, X.; He, Y.; Gao, Y.; Yang, S.; Shao, Z.; Ye, Z. IL-4 administration exerts preventive effects via suppression of underlying inflammation and TNF-α-induced apoptosis in steroid-induced osteonecrosis. Osteoporos. Int. 2016, 27, 1827–1837. [Google Scholar] [CrossRef] [PubMed]
- He, X.T.; Li, X.; Xia, Y.; Yin, Y.; Wu, R.X.; Sun, H.H.; Chen, F.M. Building capacity for macrophage modulation and stem cell recruitment in high-stiffness hydrogels for complex periodontal regeneration: Experimental studies in vitro and in rats. Acta Biomater. 2019, 88, 162–180. [Google Scholar] [CrossRef]
- Kwon, D.; Cha, B.G.; Cho, Y.; Min, J.; Park, E.B.; Kang, S.J.; Kim, J. Extra-large pore mesoporous silica nanoparticles for directing in vivo M2 macrophage polarization by delivering IL-4. Nano Lett. 2017, 17, 2747–2756. [Google Scholar] [CrossRef]
- Alvarado-Vazquez, P.A.; Bernal, L.; Paige, C.A.; Grosick, R.L.; Vilrriales, C.M.; Ferreira, D.W.; Romero-Sandoval, E.A. Macrophage-specific nanotechnology-driven CD163 overexpression in human macrophages results in an M2 phenotype under inflammatory conditions. Immunobiology 2017, 222, 900–912. [Google Scholar] [CrossRef]
- Thorp, E.; Tabas, I. Mechanisms and consequences of efferocytosis in advanced atherosclerosis. J. Leukoc. Biol. 2009, 86, 1089–1095. [Google Scholar] [CrossRef] [Green Version]
- Lieb, W.; Enserro, D.M.; Larson, M.G.; Vasan, R.S. Residual cardiovascular risk in individuals on lipid-lowering treatment: Quantifying absolute and relative risk in the community. Open Hear. 2018, 5, e000722. [Google Scholar] [CrossRef]
- Dhindsa, D.S.; Sandesara, P.B.; Shapiro, M.D.; Wong, N.D. The evolving understanding and approach to residual cardiovascular risk management. Front. Cardiovasc. Med. 2020, 7, 88. [Google Scholar] [CrossRef]
- Sager, H.B.; Dutta, P.; Dahlman, J.E.; Hulsmans, M.; Courties, G.; Sun, Y.; Nahrendorf, M. RNAi targeting multiple cell adhesion molecules reduces immune cell recruitment and vascular inflammation after myocardial infarction. Sci. Transl. Med. 2016, 8, 342ra80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, K.; Qin, X.; Li, T.; Qiu, J.; Yin, T.; Wang, G. Biomimetic nanotherapies: Red blood cell based core–shell structured nanocomplexes for atherosclerosis management. Adv. Sci. 2019, 6, 1900172. [Google Scholar] [CrossRef] [PubMed]
- Boada, C.; Zinger, A.; Tsao, C.; Zhao, P.; Martinez, J.O.; Hartman, K.; Tasciotti, E. Rapamycin-loaded biomimetic nanoparticles reverse vascular inflammation. Circ. Res. 2020, 126, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Dou, Y.; Chen, Y.; Zhang, X.; Xu, X.; Chen, Y.; Guo, J.; Zhang, J. Non-proinflammatory and responsive nanoplatforms for targeted treatment of atherosclerosis. Biomaterials 2017, 143, 93–108. [Google Scholar] [CrossRef]
- Kojima, Y.; Volkmer, J.P.; McKenna, K.; Civelek, M.; Lusis, A.J.; Miller, C.L.; Leeper, N.J. CD47-blocking antibodies restore phagocytosis and prevent atherosclerosis. Nature 2016, 536, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Flores, A.M.; Hosseini-Nassab, N.; Jarr, K.U.; Ye, J.; Zhu, X.; Wirka, R.; Leeper, N.J. Pro-efferocytic nanoparticles are specifically taken up by lesional macrophages and prevent atherosclerosis. Nat. Nanotechnol. 2020, 15, 154–161. [Google Scholar] [CrossRef]
- Kamaly, N.; Fredman, G.; Fojas, J.J.R.; Subramanian, M.; Choi, W.I.; Zepeda, K.; Farokhzad, O.C. Targeted interleukin-10 nanotherapeutics developed with a microfluidic chip enhance resolution of inflammation in advanced atherosclerosis. ACS Nano 2016, 10, 5280–5292. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Sahu, A.; Hwang, Y.; Kim, G.B.; Nam, G.H.; Kim, I.S.; Tae, G. Targeted delivery of anti-inflammatory cytokine by nanocarrier reduces atherosclerosis in Apo E−/−mice. Biomaterials 2019, 226, 119550. [Google Scholar] [CrossRef]
- Allen, S.D.; Liu, Y.G.; Kim, T.; Bobbala, S.; Yi, S.; Zhang, X.; Scott, E.A. Celastrol-loaded PEG-b-PPS nanocarriers as an anti-inflammatory treatment for atherosclerosis. Biomater. Sci. 2018, 7, 657–668. [Google Scholar] [CrossRef]
- Nakashiro, S.; Matoba, T.; Umezu, R.; Koga, J.I.; Tokutome, M.; Katsuki, S.; Egashira, K. Pioglitazone-incorporated nanoparticles prevent plaque destabilization and rupture by regulating monocyte/macrophage differentiation in ApoE−/− mice. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 491–500. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Zhang, J.; Zhao, Q.; Zhuang, W.; Ding, J.; Zhang, C.; Xie, H.Y. Molecularly engineered macrophage-derived exosomes with inflammation tropism and intrinsic heme biosynthesis for atherosclerosis treatment. Angewandte Chemie Angew. Chem. 2019, 132, 4097–4103. [Google Scholar] [CrossRef]
- Kennedy, M.A.; Barrera, G.C.; Nakamura, K.; Baldán, Á.; Tarr, P.; Fishbein, M.C.; Edwards, P.A. ABCG1 has a critical role in mediating cholesterol efflux to HDL and preventing cellular lipid accumulation. Cell Metab. 2005, 1, 121–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oram, J.F.; Lawn, R.M.; Garvin, M.R.; Wade, D.P. ABCA1 is the cAMP-inducible apolipoprotein receptor that mediates cholesterol secretion from macrophages. J. Biol. Chem. 2000, 274, 34508–34511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Lan, D.; Chen, W.; Matsuura, F.; Tall, A.R. ATP-binding cassette transporters G1 and G4 mediate cellular cholesterol efflux to high-density lipoproteins. Proc. Natl. Acad. Sci. USA 2004, 101, 9774–9779. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Silver, D.L.; Costet, P.; Tall, A.R. Specific binding of ApoA-I, enhanced cholesterol efflux, and altered plasma membrane morphology in cells expressing ABC1. J. Biol. Chem. 2000, 275, 33053–33058. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Dou, Y.; Chen, Y.; Qi, Y.; Li, L.; Han, S.; Zhang, J. Site-specific microRNA-33 antagonism by pH-responsive nanotherapies for treatment of atherosclerosis via regulating cholesterol efflux and adaptive immunity. Adv. Funct. Mater. 2020, 30, 2002131. [Google Scholar] [CrossRef]
- Yu, M.; Amengual, J.; Menon, A.; Kamaly, N.; Zhou, F.; Xu, X.; Farokhzad, O.C. Targeted nanotherapeutics encapsulating liver X receptor agonist GW3965 enhance antiatherogenic effects without adverse effects on hepatic lipid metabolism in Ldlr−/− Mice. Adv. Healthc. Mater. 2017, 6, 1700313. [Google Scholar] [CrossRef]
- Kim, H.; Kim, B.H.; Huh, B.K.; Yoo, Y.C.; Heo, C.Y.; Choy, Y.B.; Park, J.H. Surgical suture releasing macrophage-targeted drug-loaded nanoparticles for an enhanced anti-inflammatory effect. Biomater. Sci. 2017, 5, 1670–1677. [Google Scholar] [CrossRef]
- Bellani, G.; Laffey, J.G.; Pham, T.; Fan, E.; Brochard, L.; Esteban, A. Epidemiology, patterns of care, and mortality for patients with acute respiratory distress syndrome in intensive care units in 50 countries. JAMA 2016, 315, 788–800. [Google Scholar] [CrossRef]
- Riviello, E.D.; Kiviri, W.; Twagirumugabe, T.; Mueller, A.; Banner-Goodspeed, V.M.; Officer, L.; Fowler, R.A. Hospital incidence and outcomes of the acute respiratory distress syndrome using the Kigali modification of the Berlin definition. Am. J. Respir. Crit. Care Med. 2016, 193, 52–59. [Google Scholar] [CrossRef]
- Fan, E.; Brodie, D.; Slutsky, A.S. Acute respiratory distress syndrome: Advances in diagnosis and treatment. JAMA 2018, 319, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Thompson, T.B.; Chambers, R.C.; Liu, K.D. Acute respiratory distress syndrome. N. Engl. J. Med. 2017, 377, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Laffey, J.G.; Kavanagh, B.P. Acute Respiratory Distress Syndrome Network. Ventilation with lower tidal volumes as compared with traditional tidal volumes for acute lung injury and the acute respiratory distress syndrome. N. Engl. J. Med. 2000, 342, 1301–1308. [Google Scholar]
- Villar, J.; Kacmarek, R.M.; Pérez-Méndez, L.; Aguirre-Jaime, A. A high positive end-expiratory pressure, low tidal volume ventilatory strategy improves outcome in persistent acute respiratory distress syndrome: A randomized, controlled trial. Crit. Care Med. 2006, 34, 1311–1318. [Google Scholar] [CrossRef] [PubMed]
- Matthay, M.A.; Zemans, R.L.; Zimmerman, G.A.; Arabi, Y.M.; Beitler, J.R.; Mercat, A.; Calfee, C.S. Acute respiratory distress syndrome. Nat. Rev. Dis. Primers 2019, 5, 1–22. [Google Scholar] [CrossRef]
- Huang, X.; Xiu, H.; Zhang, S.; Zhang, G. The role of macrophages in the pathogenesis of ALI/ARDS. Mediat. Inflamm. 2018, 2018, 1264913. [Google Scholar] [CrossRef]
- Xiong, Y.; Gao, W.; Xia, F.; Sun, Y.; Sun, L.; Wang, L.; Li, Q. Peptide–Gold Nanoparticle Hybrids as Promising Anti-Inflammatory Nanotherapeutics for Acute Lung Injury: In Vivo Efficacy, Biodistribution, and Clearance. Adv. Healthc. Mater. 2018, 7, 1800510. [Google Scholar] [CrossRef]
- Kinne, R.W.; Bräuer, R.; Stuhlmüller, B.; Palombo-Kinne, E.; Burmester, G.R. Macrophages in rheumatoid arthritis. Arthritis Res. Ther. 2000, 2, 189. [Google Scholar]
- Blackwell, S.T.; Christman, J.W. The role of nuclear factor-κ B in cytokine gene regulation. Am. J. Respir. Cell Mol. Biol. 1997, 17, 3–9. [Google Scholar] [CrossRef]
- Hattori, Y.; Sakaguchi, M.; Maitani, Y. Folate-linked lipid-based nanoparticles deliver a NFκB decoy into activated murine macrophage-like RAW264. 7 cells. Biol. Pharm. Bull. 2006, 29, 1516–1520. [Google Scholar] [CrossRef] [Green Version]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, A.S.; Obin, M.S. Obesity and the role of adipose tissue in inflammation and metabolism. Am. J. Clin. Nutr. 2006, 83, 461S–465S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luca, D.; Carl, O.J.M. Inflammation and insulin resistance. FEBS Lett. 2008, 582, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Investig. 2007, 117, 175–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goktas, Z.; Zu, Y.; Abbasi, M.; Galyean, S.; Wu, D.; Fan, Z.; Wang, S. Recent advances in nanoencapsulation of phytochemicals to combat obesity and its comorbidities. J. Agric. Food Chem. 2020, 68, 8119–8131. [Google Scholar] [CrossRef]
- Ahmad, M.; Gani, A. Development of novel functional snacks containing nano-encapsulated resveratrol with anti-diabetic, anti-obesity and antioxidant properties. Food Chem. 2021, 352, 129323. [Google Scholar] [CrossRef]
- Sharifan, P.; Ziaee, A.; Darroudi, S.; Rezaie, M.; Safarian, M.; Eslami, S.; Ghayour Mobarhan, M. Effect of low-fat dairy products fortified with 1500IU nano encapsulated vitamin D3 on cardiometabolic indicators in adults with abdominal obesity: A total blinded randomized controlled trial. Curr. Med Res. Opin. 2021, 37, 579–588. [Google Scholar] [CrossRef]
- Ma, L.; Liu, T.W.; Wallig, M.A.; Dobrucki, I.T.; Dobrucki, L.W.; Nelson, E.R.; Smith, A.M. Efficient targeting of adipose tissue macrophages in obesity with polysaccharide nanocarriers. ACS Nano 2016, 10, 6952–6962. [Google Scholar] [CrossRef]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging: An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef]
- lvarez-Rodríguez, L.; López-Hoyos, M.; Muñoz-Cacho, P.; Martínez-Taboada, V.M. Aging is associated with circulating cytokine dysregulation. Cell. Immunol. 2012, 273, 124–132. [Google Scholar] [CrossRef]
- Shaw, A.C.; Goldstein, D.R.; Montgomery, R.R. Age-dependent dysregulation of innate immunity. Nat. Rev. Immunol. 2013, 13, 875–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnardottir, H.H.; Dalli, J.; Colas, R.A.; Shinohara, M.; Serhan, C.N. Aging delays resolution of acute inflammation in mice: Reprogramming the host response with novel nano-proresolving medicines. J. Immunol. 2014, 193, 4235–4244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, G.D.; Choi, S.W.; Cai, X.; Li, W.; Cho, E.C.; Jeong, U.; Xia, Y. A new theranostic system based on gold nanocages and phase-change materials with unique features for photoacoustic imaging and controlled release. J. Am. Chem. Soc. 2011, 133, 4762–4765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Huo, D.; Chen, Q.; Xue, J.; Shen, S.; Xia, Y. A Eutectic Mixture of Natural Fatty Acids Can Serve as the Gating Material for Near-Infrared-Triggered Drug Release. Adv. Mater. 2017, 29, 1703702. [Google Scholar] [CrossRef] [PubMed]
- Bentzon, J.F.; Falk, E. Atherosclerotic lesions in mouse and man: Is it the same disease? Curr. Opin. Lipidol. 2010, 21, 434–440. [Google Scholar] [CrossRef]
Figure 1.
Current nanomaterials for regulating macrophage polarization. (A) Surface modification of inorganic NPs to modulate macrophage polarization. (B) Organic NPs.
Figure 1.
Current nanomaterials for regulating macrophage polarization. (A) Surface modification of inorganic NPs to modulate macrophage polarization. (B) Organic NPs.

Figure 2.
Summary of macrophage-targeting nanotechnologies. (A) Passive macrophage-targeting. NPs accumulate in tissues through the vascular leakage. (B) Passive macrophage-targeting. NPs with different sizes preferentially accumulate in different organs. (C) Active macrophage-targeting. With various surface modification methods, NPs can specially target macrophages via recognition by receptors on macrophage membrane.
Figure 2.
Summary of macrophage-targeting nanotechnologies. (A) Passive macrophage-targeting. NPs accumulate in tissues through the vascular leakage. (B) Passive macrophage-targeting. NPs with different sizes preferentially accumulate in different organs. (C) Active macrophage-targeting. With various surface modification methods, NPs can specially target macrophages via recognition by receptors on macrophage membrane.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cai, D.; Gao, W.; Li, Z.; Zhang, Y.; Xiao, L.; Xiao, Y. Current Development of Nano-Drug Delivery to Target Macrophages. Biomedicines 2022, 10, 1203. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10051203
AMA Style
Cai D, Gao W, Li Z, Zhang Y, Xiao L, Xiao Y. Current Development of Nano-Drug Delivery to Target Macrophages. Biomedicines. 2022; 10(5):1203. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10051203
Chicago/Turabian StyleCai, Donglin, Wendong Gao, Zhelun Li, Yufeng Zhang, Lan Xiao, and Yin Xiao. 2022. "Current Development of Nano-Drug Delivery to Target Macrophages" Biomedicines 10, no. 5: 1203. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10051203
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.