
Thrombin Activity in Rodent and Human Skin: Modified by Inflammation and Correlates with Innervation

 , , and
, , and 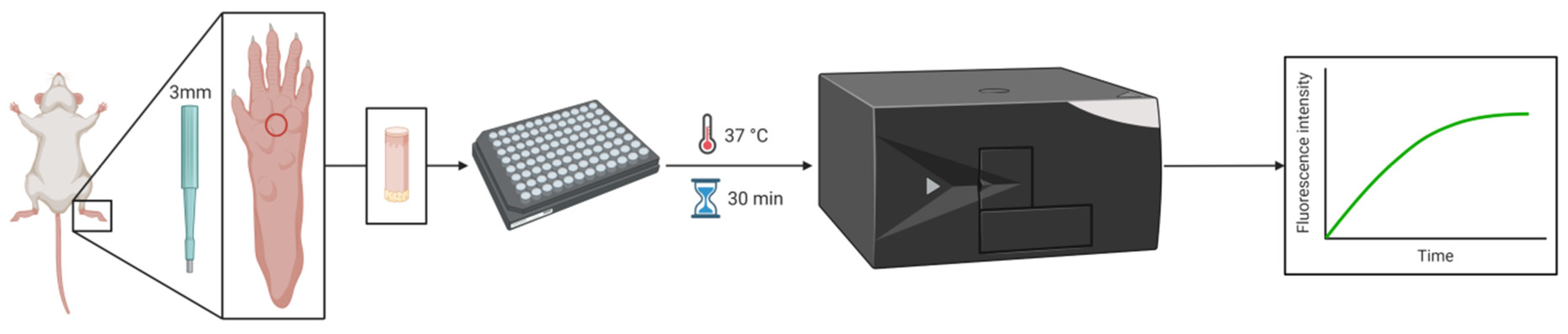{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Adjuvant-Induced Focal Inflammation
2.3. LPS-Induced Systemic Inflammation
2.4. Hot-Plate Test
2.5. Thrombin Activity Measurement
2.6. Histology
2.7. Human Patients
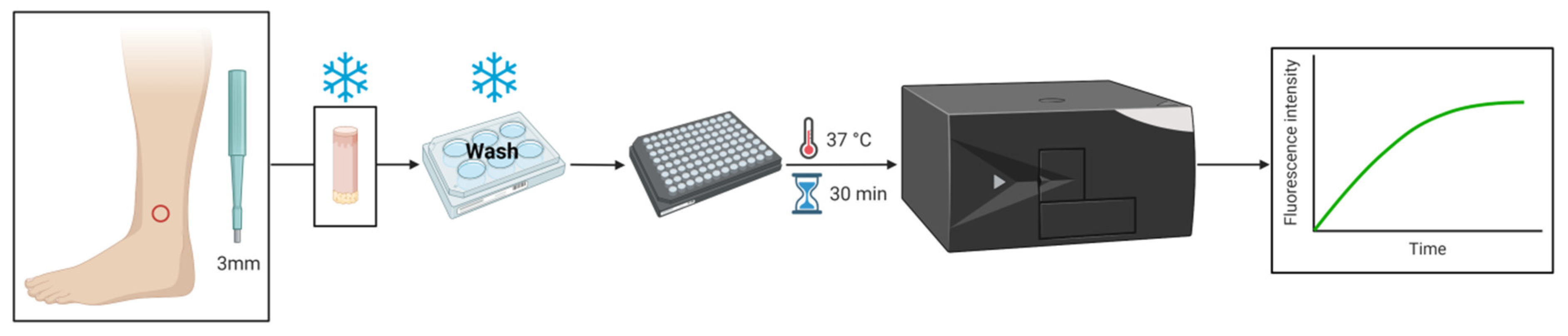
2.8. Skin Biopsy
2.9. Statistical Analysis
3. Results
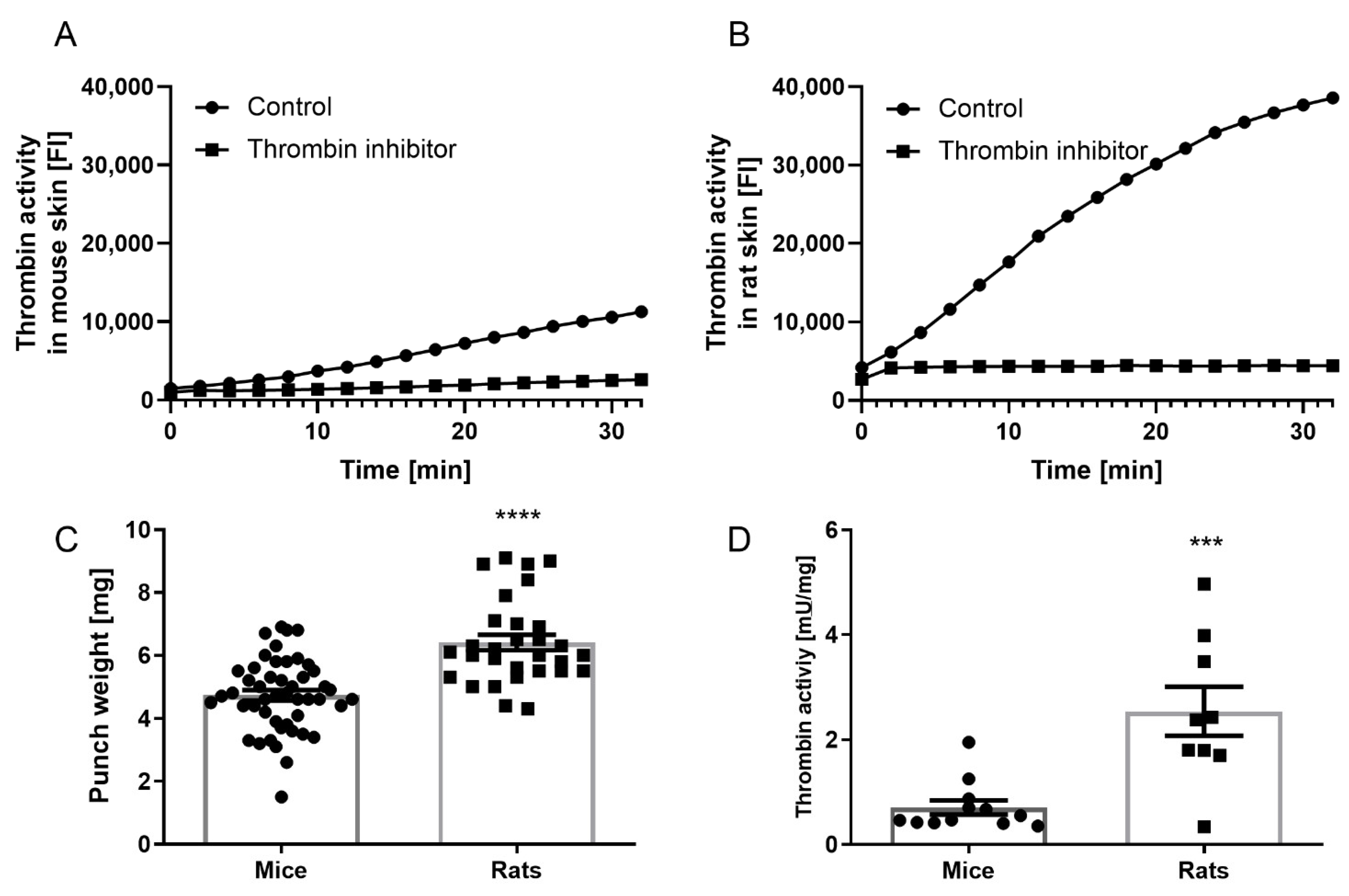
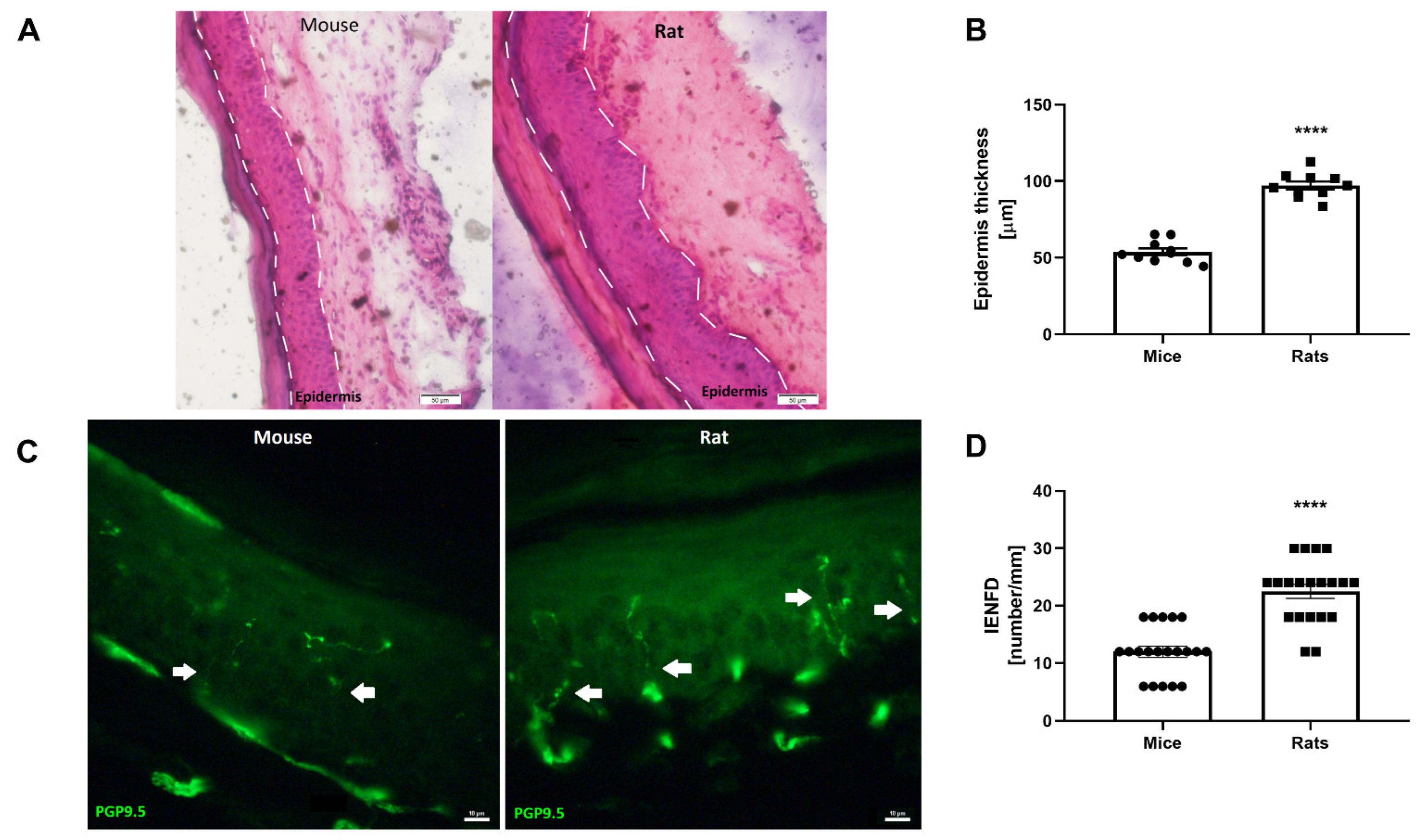
3.1. Mouse versus Rat Skin
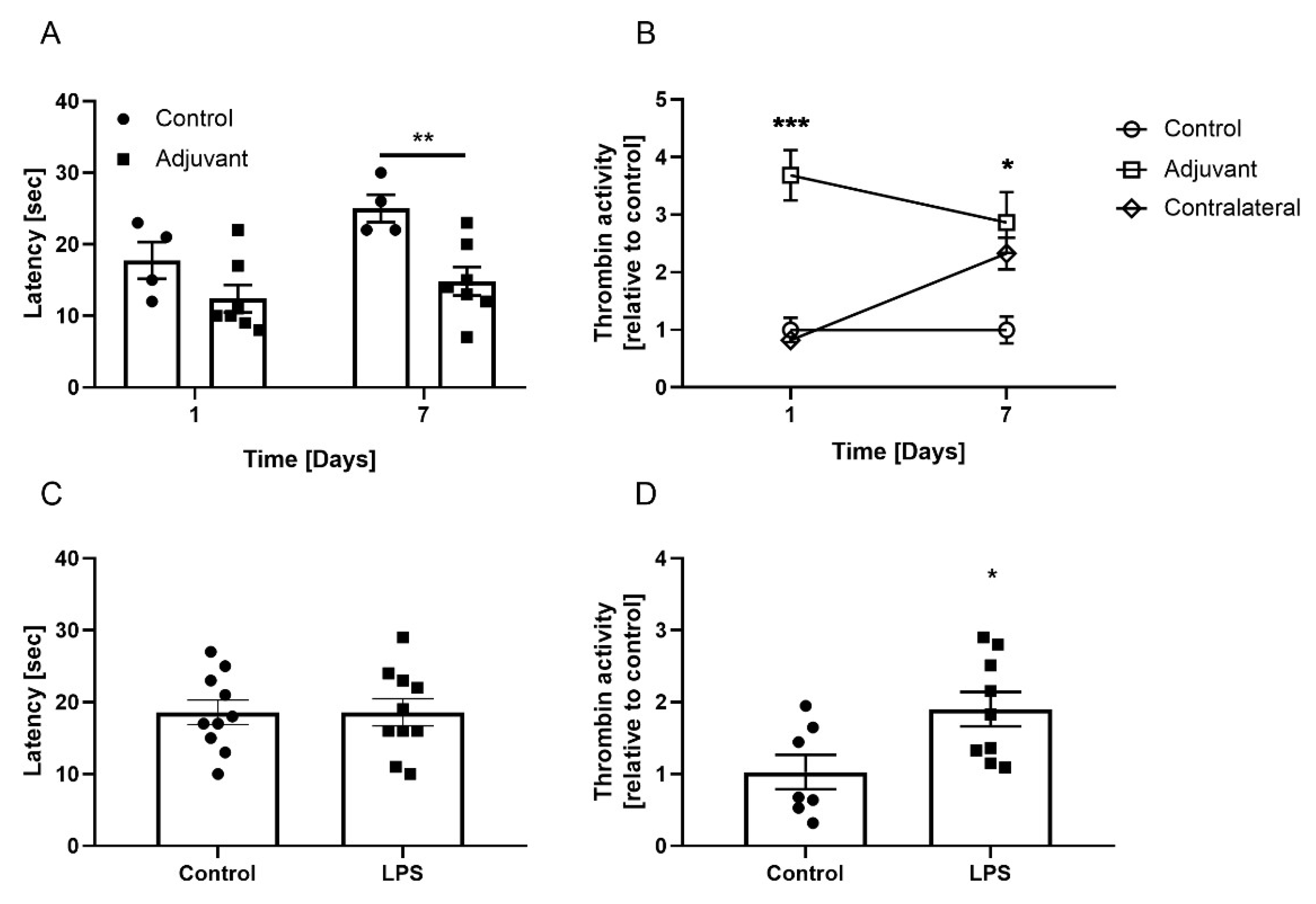
3.2. Skin Thrombin Activity in Adjuvant-Induced Focal Inflammation
3.3. Skin Thrombin Activity in LPS-Induced Systemic Inflammation
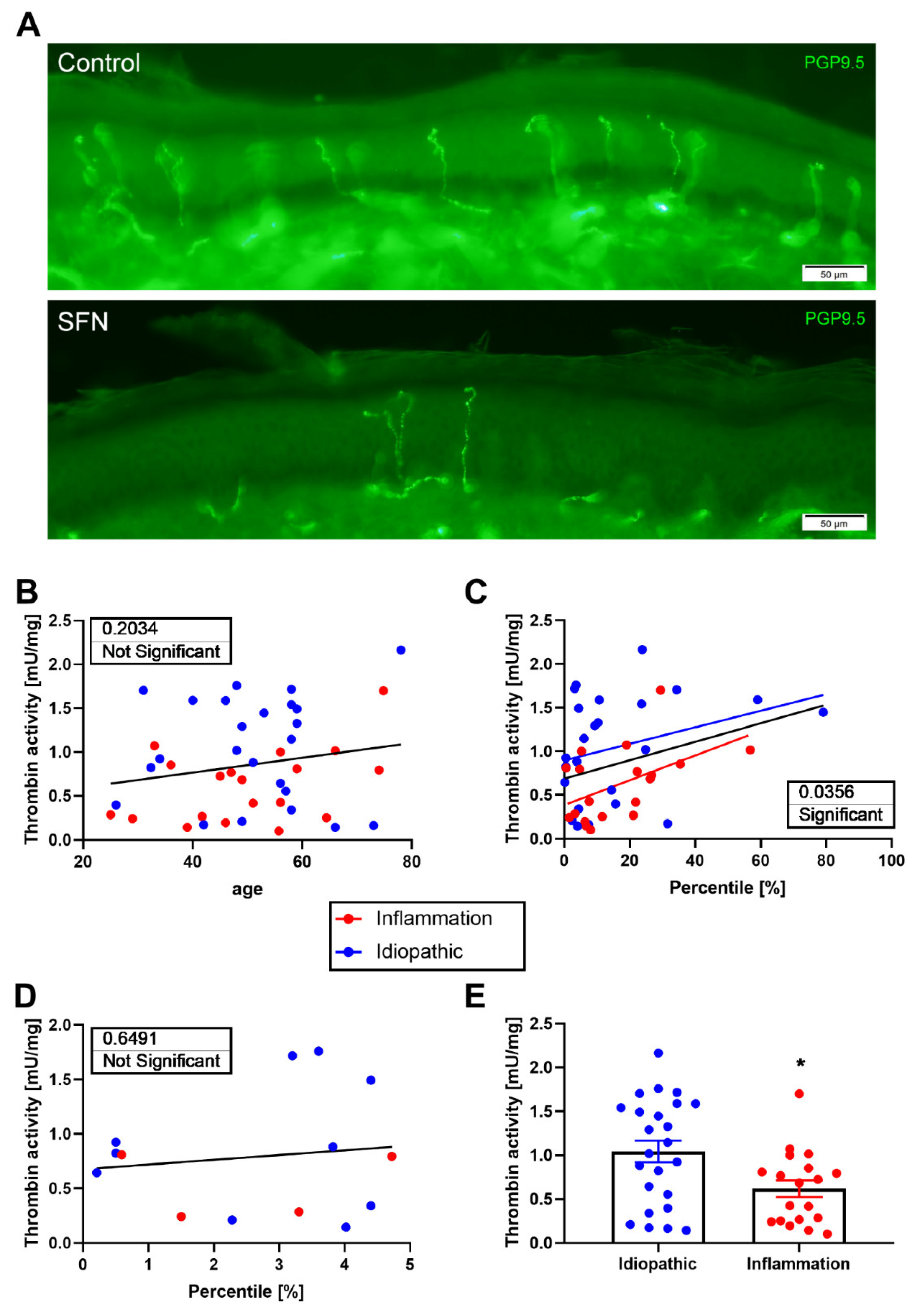
3.4. Thrombin Activity in Human Skin
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iba, T.; Levi, M.; Levy, J.H. Sepsis-Induced Coagulopathy and Disseminated Intravascular Coagulation. Semin. Thromb. Hemost. 2020, 46, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.N.; Ramachandran, R.; Yau, M.-K.; Suen, J.Y.; Fairlie, D.P.; Hollenberg, M.D.; Hooper, J.D. Structure, Function and Pathophysiology of Protease Activated Receptors. Pharmacol. Ther. 2011, 130, 248–282. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.-J.; Yamanaka, H.; Obata, K.; Dai, Y.; Kobayashi, K.; Kozai, T.; Tokunaga, A.; Noguchi, K. Expression of MRNA for Four Subtypes of the Proteinase-Activated Receptor in Rat Dorsal Root Ganglia. Brain Res. 2005, 1041, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Maggio, N.; Shavit, E.; Chapman, J.; Segal, M. Thrombin Induces Long-Term Potentiation of Reactivity to Afferent Stimulation and Facilitates Epileptic Seizures in Rat Hippocampal Slices: Toward Understanding the Functional Consequences of Cerebrovascular Insults. J. Neurosci. 2008, 28, 732–736. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.J.; Woo, D.H.; Lee, P.R.; Pajevic, S.; Bukalo, O.; Huffman, W.C.; Wake, H.; Basser, P.J.; SheikhBahaei, S.; Lazarevic, V.; et al. Regulation of myelin structure and conduction velocity by perinodal astrocytes. Proc. Natl. Acad. Sci. USA 2018, 115, 11832–11837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Almeida, V.H.; Monteiro, R.Q. Protease-Activated Receptor 1 (PAR1): A Promising Target for the Treatment of Glioblastoma? Transl. Cancer Res. 2016, 5, S1274–S1280. [Google Scholar] [CrossRef]
- Krenzlin, H.; Lorenz, V.; Alessandri, B. The Involvement of Thrombin in the Pathogenesis of Glioblastoma. J. Neurosci. Res. 2017, 95, 2080–2085. [Google Scholar] [CrossRef]
- Han, M.H.; Hwang, S.I.; Roy, D.B.; Lundgren, D.H.; Price, J.V.; Ousman, S.S.; Fernald, G.H.; Gerlitz, B.; Robinson, W.H.; Baranzini, S.E.; et al. Proteomic Analysis of Active Multiple Sclerosis Lesions Reveals Therapeutic Targets. Nature 2008, 451, 1076–1081. [Google Scholar] [CrossRef]
- Shavit-Stein, E.; Aronovich, R.; Sylantiev, C.; Gofrit, S.G.; Chapman, J.; Dori, A. The Role of Thrombin in the Pathogenesis of Diabetic Neuropathy. PLoS ONE 2019, 14, 0219453. [Google Scholar] [CrossRef]
- Shavit-Stein, E.; Aronovich, R.; Sylantiev, C.; Gera, O.; Gofrit, S.G.; Chapman, J.; Dori, A. Blocking Thrombin Significantly Ameliorates Experimental Autoimmune Neuritis. Front. Neurol. 2018, 9, 1139. [Google Scholar] [CrossRef] [Green Version]
- Shavit-Stein, E.; Beilin, O.; Korczyn, A.D.; Sylantiev, C.; Aronovich, R.; Drory, V.E.; Gurwitz, D.; Horresh, I.; Bar-Shavit, R.; Peles, E.; et al. Thrombin Receptor PAR-1 on Myelin at the Node of Ranvier: A New Anatomy and Physiology of Conduction Block. Brain 2008, 131, 1113–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahimi, S.; Jaberi, N.; Avan, A.; Ryzhikov, M.; Keramati, M.R.; Parizadeh, M.R.; Hassanian, S.M. Role of Thrombin in the Pathogenesis of Central Nervous System Inflammatory Diseases. J. Cell Physiol. 2017, 232, 482–485. [Google Scholar] [CrossRef] [PubMed]
- De Garavilla, L.; Vergnolle, N.; Young, S.H.; Ennes, H.; Steinhoff, M.; Ossovskaya, V.S.; D’Andrea, M.R.; Mayer, E.A.; Wallace, J.L.; Hollenberg, M.D.; et al. Agonists of Proteinase-Activated Receptor 1 Induce Plasma Extravasation by a Neurogenic Mechanism. Br. J. Pharmacol. 2001, 133, 975–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vellani, V.; Kinsey, A.M.; Prandini, M.; Hechtfischer, S.C.; Reeh, P.; Magherini, P.C.; Giacomoni, C.; McNaughton, P.A. Protease Activated Receptors 1 and 4 Sensitize TRPV1 in Nociceptive Neurones. Mol. Pain 2010, 6, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hovaguimian, A.; Gibbons, C.H. Diagnosis and Treatment of Pain in Small-Fiber Neuropathy. Curr. Pain Headache Rep. 2011, 15, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Lauria, G.; Lombardi, R. Skin Biopsy: A New Tool for Diagnosing Peripheral Neuropathy. Br. Med. J. 2007, 334, 1159–1162. [Google Scholar] [CrossRef] [Green Version]
- Malmberg, A.B.; Bannon, A.W. Models of Nociception: Hot-Plate, Tail-Flick, and Formalin Tests in Rodents. In Current Protocols in Neuroscience; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2001; Volume 6, pp. 8.9.1–8.9.15. [Google Scholar]
- Bushi, D.; Chapman, J.; Katzav, A.; Shavit-Stein, E.; Molshatzki, N.; Maggio, N.; Tanne, D. Quantitative Detection of Thrombin Activity in an Ischemic Stroke Model. J. Mol. Neurosci. 2013, 51, 844–850. [Google Scholar] [CrossRef]
- Shavit-Stein, E.; Sheinberg, E.; Golderman, V.; Sharabi, S.; Wohl, A.; Gofrit, S.G.; Zivli, Z.; Shelestovich, N.; Last, D.; Guez, D.; et al. A Novel Compound Targeting Protease Receptor 1 Activators for the Treatment of Glioblastoma. Front. Neurol. 2018, 9, 1087. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nature Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Lauria, G.; Hsieh, S.T.; Johansson, O.; Kennedy, W.R.; Leger, J.M.; Mellgren, S.I.; Nolano, M.; Merkies, I.S.J.; Polydefkis, M.; Smith, A.G.; et al. European Federation of Neurological Societies/Peripheral Nerve Society Guideline on the Use of Skin Biopsy in the Diagnosis of Small Fiber Neuropathy. Report of a Joint Task Force of the European Federation of Neurological Societies and the Peripheral Ner. J. Peripher. Nerv. Syst. 2010, 15, 79–92. [Google Scholar] [CrossRef]
- Lauria, G.; Bakkers, M.; Schmitz, C.; Lombardi, R.; Penza, P.; Devigili, G.; Smith, A.G.; Hsieh, S.T.; Mellgren, S.I.; Umapathi, T.; et al. Intraepidermal Nerve Fiber Density at the Distal Leg: A Worldwide Normative Reference Study. J. Peripher. Nerv. Syst. 2010, 15, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Gerasimov, A.; Golderman, V.; Gofrit, S.; Aharoni, S.; Zohar, D.; Itsekson-Hayosh, Z.; Fay-Karmon, T.; Hassin-Baer, S.; Chapman, J.; Maggio, N.; et al. Markers for Neural Degeneration and Regeneration: Novel Highly Sensitive Methods for the Measurement of Thrombin and Activated Protein C in Human Cerebrospinal Fluid. Neural Regen. Res. 2021, 16, 2086–2092. [Google Scholar] [CrossRef] [PubMed]
- Gera, O.; Shavit-Stein, E.; Bushi, D.; Harnof, S.; Shimon, M.B.; Weiss, R.; Golderman, V.; Dori, A.; Maggio, N.; Finegold, K.; et al. Thrombin and Protein C Pathway in Peripheral Nerve Schwann Cells. Neuroscience 2016, 339, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Hanson, J. The Histogenesis of the Epidermis in the Rat and Mouse. J. Anat. 1947, 81, 174–17497. [Google Scholar] [PubMed]
- Shavit Stein, E.; ben Shimon, M.; Artan Furman, A.; Golderman, V.; Chapman, J.; Maggio, N. Thrombin Inhibition Reduces the Expression of Brain Inflammation Markers upon Systemic LPS Treatment. Neural Plast. 2018, 2018, 7692182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobo, R.; García-Piqueras, J.; Cobo, J.; Vega, J.A. The Human Cutaneous Sensory Corpuscles: An Update. J. Clin. Med. 2021, 10, 227. [Google Scholar] [CrossRef]
- Shavit-Stein, E.; Gofrit, S.G.; Gayster, A.; Teldan, Y.; Ron, A.; Bandora, E.A.; Golderman, V.; Gera, O.; Harnof, S.; Chapman, J.; et al. Treatment of Diabetic Neuropathy with A Novel PAR1-Targeting Molecule. Biomolecules 2020, 10, 1552. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.H.; Jeng, S.F.; Lu, T.H.; Yang, J.C.S.; Hsieh, M.W.; Chen, Y.C.; Rau, C.S. Correlation between Skin Biopsy with Quantification of Intraepidermal Nerve Fiber and the Severity of Sciatic Nerve Traction Injury in Rats. J. Trauma 2009, 66, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Marchand, F.; Perretti, M.; McMahon, S.B. Role of the Immune System in Chronic Pain. Nat. Rev. Neurosci. 2005, 6, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Gera, O.; Shavit-Stein, E.; Chapman, J. The Effect of Neuronal Activity on Glial Thrombin Generation. J. Mol. Neurosci. 2019, 67, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Reuveni, G.; Golderman, V.; Shavit-Stein, E.; Rosman, Y.; Shrot, S.; Chapman, J.; Harnof, S. Measuring Thrombin Activity in Frozen Brain Tissue. NeuroReport 2017, 28, 1176–1179. [Google Scholar] [CrossRef] [PubMed]
- Mcglone, F.; Olausson, H.; Boyle, J.A.; Jones-Gotman, M.; Dancer, C.; Guest, S.; Essick, G. Touching and Feeling: Differences in Pleasant Touch Processing between Glabrous and Hairy Skin in Humans. Eur. J. Neurosci. 2012, 35, 1782–1788. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golderman, V.; Berkowitz, S.; Guly Gofrit, S.; Gera, O.; Aharoni, S.A.; Zohar, D.N.; Keren, D.; Dori, A.; Chapman, J.; Shavit-Stein, E. Thrombin Activity in Rodent and Human Skin: Modified by Inflammation and Correlates with Innervation. Biomedicines 2022, 10, 1461. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10061461
Golderman V, Berkowitz S, Guly Gofrit S, Gera O, Aharoni SA, Zohar DN, Keren D, Dori A, Chapman J, Shavit-Stein E. Thrombin Activity in Rodent and Human Skin: Modified by Inflammation and Correlates with Innervation. Biomedicines. 2022; 10(6):1461. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10061461
Chicago/Turabian StyleGolderman, Valery, Shani Berkowitz, Shani Guly Gofrit, Orna Gera, Shay Anat Aharoni, Daniela Noa Zohar, Daria Keren, Amir Dori, Joab Chapman, and Efrat Shavit-Stein. 2022. "Thrombin Activity in Rodent and Human Skin: Modified by Inflammation and Correlates with Innervation" Biomedicines 10, no. 6: 1461. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10061461