Orthosiphon stamineus Standardized Extract Reverses Streptozotocin-Induced Alzheimer’s Disease-Like Condition in a Rat Model


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Extract Standardization of Orthosiphon stamineus 50% Ethanolic Extract
2.2. LC-MS Analysis
2.3. Animals
2.4. Intracerebroventricular (ICV) Infusion of Streptozotocin
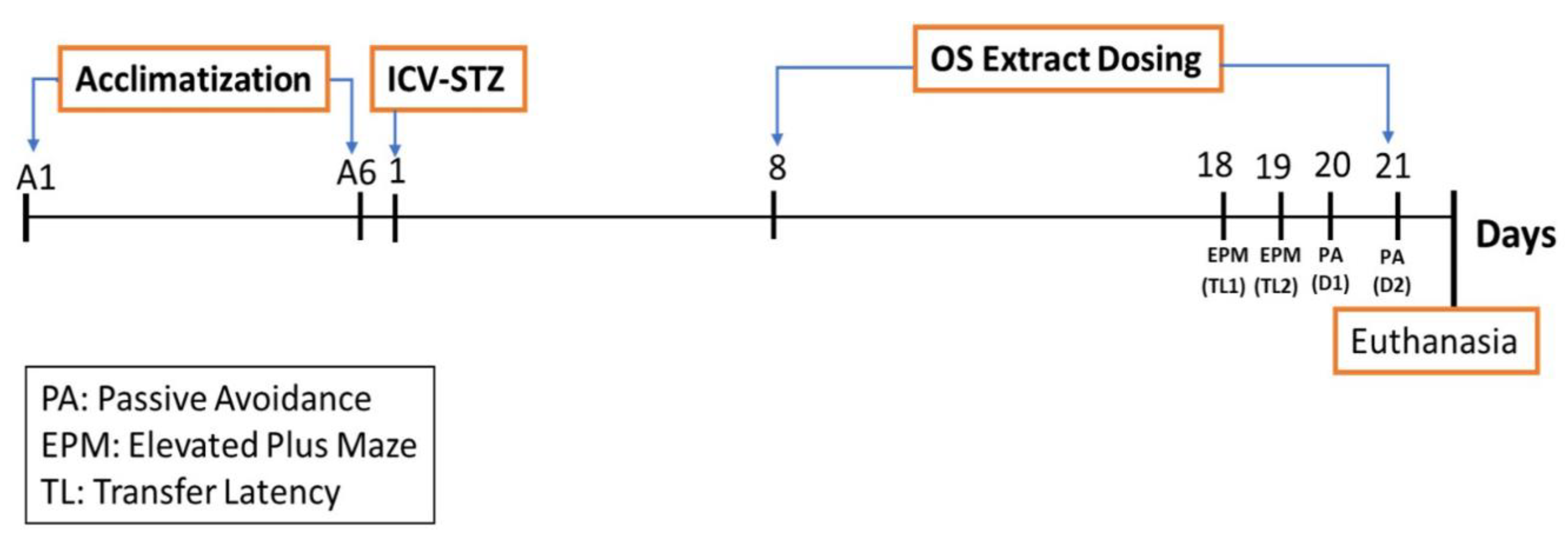
2.5. Experimental Design
2.6. Elevated Plus Maze (EPM)
2.7. Passive Avoidance (PA)
2.8. Gene Expression
2.9. Statistical Analysis
3. Results
3.1. Characterization of O. stamineus Ethanolic Extract
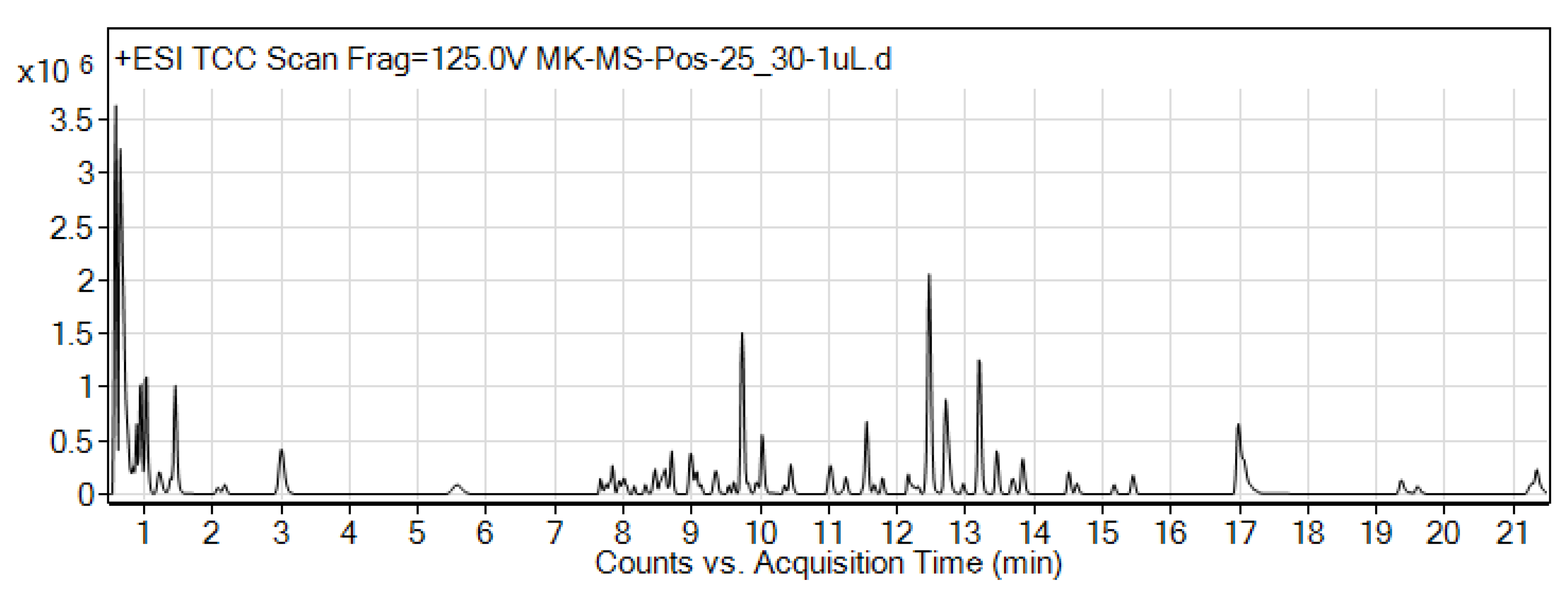
3.2. LC-MS Analysis
3.3. Effect of OS Extract on Memory Performance in EPM and PA Task in ICV-STZ Infused Rats
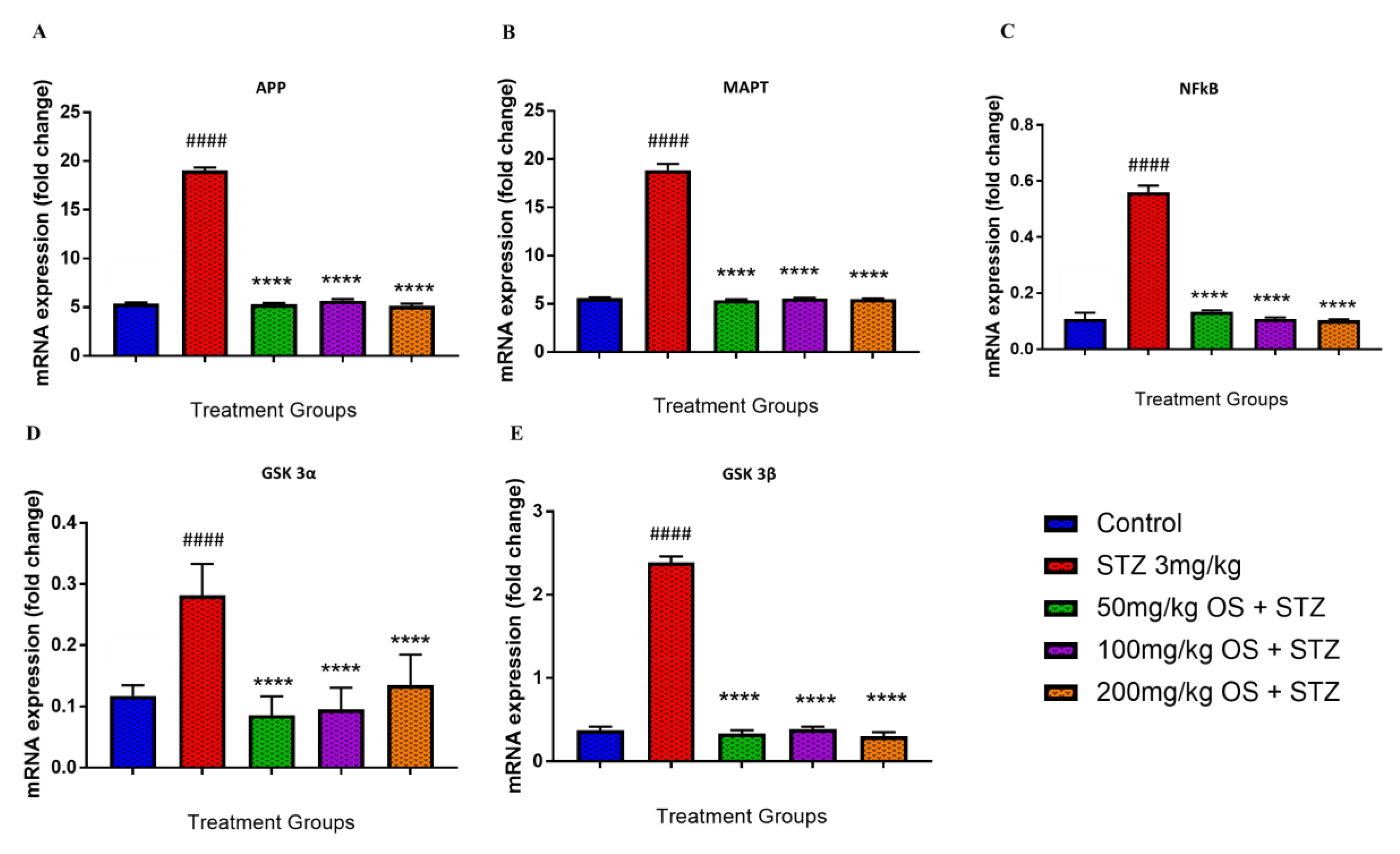
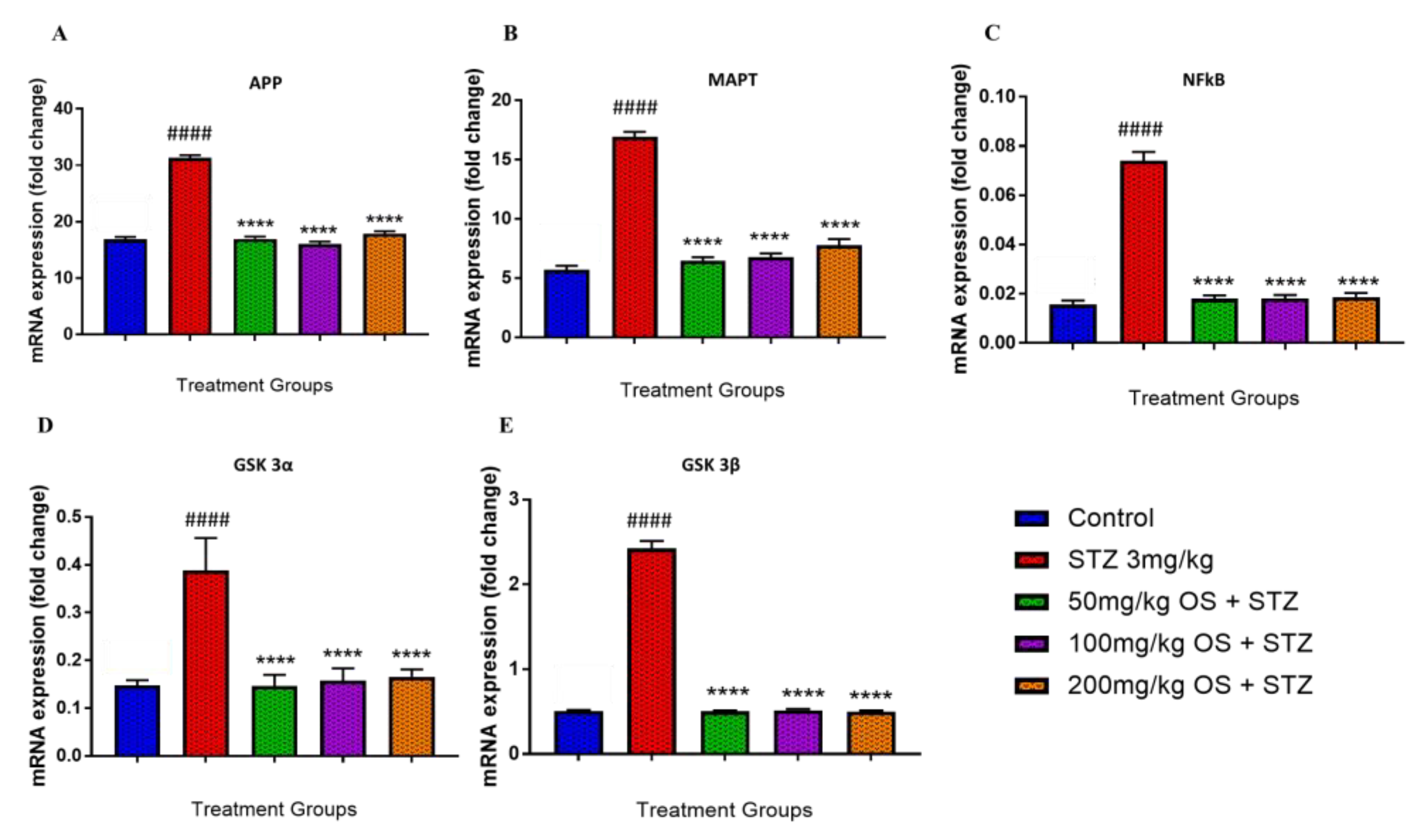
3.4. Effect of OS Extract on the Gene Expression in the Rat Hippocampal and Prefrontal Cortical Region
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zlokovic, B.V. Vascular disorder in Alzheimer’s disease: Role in pathogenesis of dementia and therapeutic targets. Adv. Drug Deliv. Rev. 2002, 54, 1553–1559. [Google Scholar] [CrossRef]
- Zhao, Y.; Gu, J.-H.; Dai, C.-L.; Liu, Q.; Iqbal, K.; Liu, F.; Gong, C.-X. Chronic cerebral hypoperfusion causes decrease of O-GlcNAcylation, hyperphosphorylation of tau and behavioral deficits in mice. Front. Aging Neurosci. 2014, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Brookmeyer, R.; Johnson, E.; Ziegler-Graham, K.; Arrighi, H.M. Forecasting the global burden of Alzheimer’s disease. Alzheimers Dement 2014, 3, 186–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluta, R.; Ulamek, M.; Jablonski, M. Alzheimer’s mechanisms in ischemic brain degeneration. Anat. Rec. (Hoboken) 2009, 292, 1863–1881. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Deshmukh, R. Embelin Attenuates Intracerebroventricular Streptozotocin-Induced Behavioral, Biochemical, and Neurochemical Abnormalities in Rats. Mol. Neurobiol. 2017, 54, 6670–6680. [Google Scholar] [CrossRef]
- Ghumatkar, P.J.; Patil, S.P.; Jain, P.D.; Tambe, R.M.; Sathaye, S. Nootropic, neuroprotective and neurotrophic effects of phloretin in scopolamine induced amnesia in mice. Pharmacol. Biochem. Behav. 2015, 135, 182–191. [Google Scholar] [CrossRef]
- Chen, X.-Q.; Mobley, W.C. Alzheimer Disease Pathogenesis: Insights From Molecular and Cellular Biology Studies of Oligomeric Aβ and Tau Species. Front. Neurosci. 2019, 13, 659. [Google Scholar] [CrossRef]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [Green Version]
- Muir, J.L. Acetylcholine, aging, and Alzheimer’s disease. Pharmacol. Biochem. Behav. 1997, 56, 687–696. [Google Scholar] [CrossRef]
- Contestabile, A. The history of the cholinergic hypothesis. Behav. Brain Res. 2011, 221, 334–340. [Google Scholar] [CrossRef]
- Craig, L.A.; Hong, N.S.; Mcdonald, R.J. Revisiting the cholinergic hypothesis in the development of Alzheimer’s disease. Neurosci. Biobehav. Rev. 2011, 35, 1397–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salkovic-Petrisic, M.; Osmanovic-Barilar, J.; Brückner, M.K.; Hoyer, S.; Arendt, T.; Riederer, P. Cerebral amyloid angiopathy in streptozotocin rat model of sporadic Alzheimer’s disease: A long-term follow up study. J. Neural Transm. 2011, 118, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Salkovic-Petrisic, M.; Knezovic, A.; Hoyer, S.; Riederer, P. What have we learned from the streptozotocin-induced animal model of sporadic Alzheimer’s disease, about the therapeutic strategies in Alzheimer’s research. J. Neural. Transm. 2013, 120, 233–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lannert, H.; Hoyer, S. Intracerebroventricular administration of streptozotocin causes long-term diminutions in learning and memory abilities and in cerebral energy metabolism in adult rats. Behav. Neurosci. 1998, 112, 1199–1208. [Google Scholar] [CrossRef]
- Sharma, M.; Gupta, Y.K. Intracerebroventricular injection of streptozotocin in rats produces both oxidative stress in the brain and cognitive impairment. Life Sci. 2001, 68, 1021–1029. [Google Scholar] [CrossRef]
- Singh, H.; Kakalij, R.; Kshirsagar, R.; Kumar, B.; Santhosh, S.; Diwan, P.D. Cognitive effects of vanillic acid against streptozotocin-induced neurodegeneration in mice. Pharm. Biol. 2014, 53, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Essa, M.M.; Vijayan, R.K.; Castellano-Gonzalez, G.; Memon, M.A.; Braidy, N.; Guillemin, G.J. Neuroprotective effect of natural products against Alzheimer’s disease. Neurochem. Res. 2012, 37, 1829–1842. [Google Scholar] [CrossRef]
- Sumaryono, W.; Proksch, P.; Wray, V.; Witte, L.; Hartmann, T. Qualitative and Quantitative Analysis of the Phenolic Constituents from Orthosiphon aristatus. Planta Med. 1991, 57, 176–180. [Google Scholar] [CrossRef]
- Awale, S.; Tezuka, Y.; Banskota, A.H.; Adnyana, I.K.; Kadota, S. Nitric Oxide Inhibitory Isopimarane-type Diterpenes from Orthosiphon stamineus of Indonesia. J. Nat. Prod. 2003, 66, 255–258. [Google Scholar] [CrossRef]
- Awale, S.; Tezuka, Y.; Banskota, A.H.; Kadota, S. Inhibition of NO production by highly-oxygenated diterpenes of Orthosiphon stamineus and their structure-activity relationship. Biol. Pharm. Bull. 2003, 26, 468–473. [Google Scholar] [CrossRef] [Green Version]
- Yam, M.F.; Vuanghao, L.; Salman, I.; Ameer, O.; Fung-Ang, L.; Noersal, R.; Albaldawi, M.; Abdullah, G.; Basir, R.; Sadikun, A.; et al. HPLC and Anti-Inflammatory Studies of the Flavonoid Rich Chloroform Extract Fraction of Orthosiphon Stamineus Leaves. Molecules 2010, 15, 4452–4466. [Google Scholar] [CrossRef] [PubMed]
- Bhullar, K.S.; Rupasinghe, H.P. Polyphenols: Multipotent therapeutic agents in neurodegenerative diseases. Oxid. Med. Cell Longev. 2013, 2013, 891748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, A.; Chinnappan, S.; Choudhary, Y.; Choudhary, V.K.; Bommu, P.; Wong, H.J. Effects of a Proprietary Standardized Orthosiphon stamineus Ethanolic Leaf Extract on Enhancing Memory in Sprague Dawley Rats Possibly via Blockade of Adenosine A 2A Receptors. Evidence-based complementary and alternative medicine. eCAM 2015, 2015, 375837. [Google Scholar] [PubMed] [Green Version]
- Sree, N.V.; Sri, P.U.; Ramarao, N. Neuro-protective properties of orthosiphon staminus (benth) leaf methanolic fraction through antioxidant mechanisms on sh-sy5y cells: An in-vitro evaluation. Int. J. Pharm. Sci. Res. 2015, 6, 1115. [Google Scholar]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 5nd ed.; Academic Press: London, UK, 1982; pp. 1–5. [Google Scholar]
- Elcioglu, H.K.; Aslan, E.; Ahmad, S.; Alan, S.; Salva, E.; Elcioglu, Ö.H.; Kabasakal, L. Tocilizumab’s effect on cognitive deficits induced by intracerebroventricular administration of streptozotocin in Alzheimer’s model. Mol. Cell Biochem. 2016, 420, 21–28. [Google Scholar] [CrossRef]
- Nakahara, N.; Iga, Y.; Mizobe, F.; Kawanishi, G. Effects of intracerebroventricular injection of AF64A on learning behaviors in rats. Jpn. J. Pharm. 2013, 48, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Retinasamy, T.; Shaikh, M.F.; Kumari, Y.; Othman, I. Ethanolic Extract of Orthosiphon stamineus Improves Memory in Scopolamine-Induced Amnesia Model. Front. Pharm. 2019, 10, 1216. [Google Scholar] [CrossRef]
- Saidan, N.H.; Hamil, M.S.; Memon, A.H.; Abdelbari, M.M.; Hamdan, M.R.; Mohd, K.S.; Majid, A.M.; Ismail, Z. Selected metabolites profiling of Orthosiphon stamineus Benth leaves extracts combined with chemometrics analysis and correlation with biological activities. BMC Complement Altern. Med. 2015, 15, 350. [Google Scholar] [CrossRef] [Green Version]
- Akowuah, G.A.; Zhari, I.; Norhayati, I.; Sadikun, A.; Khamsah, S.M. Sinensetin, eupatorin, 3′-hydroxy-5, 6, 7, 4′-tetramethoxyflavone and rosmarinic acid contents and antioxidative effect of Orthosiphon stamineus from Malaysia. Food Chem. 2004, 87, 559–566. [Google Scholar] [CrossRef]
- Park, S.A. A Common Pathogenic Mechanism Linking Type-2 Diabetes and Alzheimer’s Disease: Evidence from Animal Models. J. Clin. Neurol. 2011, 7, 10–18. [Google Scholar] [CrossRef]
- Arafat, O.M.; Roghani, M.; Khalili, M.; Baluchnejadmojarad, T. Studies on diuretic and hypouricemic effects of Orthosiphon stamineus methanol extracts in rats. J. Ethnopharmacol. 2008, 118, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Balouchnejadmojarad, T. The effect of genistein on intracerebroventricular streptozotocin-induced cognitive deficits in male rat. Basic Clin. Neurosci. J. 2009, 1, 17–21. [Google Scholar]
- Grunblatt, E.; Salkovic-Petrisic, M.; Osmanovic, J.; Riederer, P.; Hoyer, S. Brain insulin system dysfunction in streptozotocin intracerebroventricularly treated rats generates hyperphosphorylated tau protein. J. Neurochem. 2007, 101, 757–770. [Google Scholar] [CrossRef] [PubMed]
- Veerendra Kumar, M.H.; Gupta, Y.K. Effect of Centella asiatica on cognition and oxidative stress in an intracerebroventricular streptozotocin model of Alzheimer’s disease in rats. Clin. Exp. Pharmacol. Physiol. 2003, 30, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Tota, S.; Kamat, P.K.; Awasthi, H.; Singh, N.; Raghubir, R.; Nath, C.; Hanif, K. Candesartan improves memory decline in mice: Involvement of AT1 receptors in memory deficit induced by intracerebral streptozotocin. Behav. Brain. Res. 2009, 199, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, R.; Mishra, B.; Tyagi, E.; Nath, C.; Shukla, R. Effect of curcumin on brain insulin receptors and memory functions in STZ (ICV) induced dementia model of rat. Pharm. Res. 2010, 61, 247–252. [Google Scholar] [CrossRef]
- Mehla, J.; Mehla, J.; Pahuja, M.; Gupta, P.; Dethe, S.; Agarwal, A.; Gupta, Y.K. Clitoria ternatea ameliorated the intracerebroventricularly injected streptozotocin induced cognitive impairment in rats: Behavioral and biochemical evidence. Psychopharmacology 2013, 230, 589–605. [Google Scholar] [CrossRef]
- Martinez, A.; Perez, D.I. GSK-3 inhibitors: A ray of hope for the treatment of Alzheimer’s disease? J. Alzheimers Dis. 2008, 15, 181–191. [Google Scholar] [CrossRef]
- Kremer, A.; Louis, J.V.; Jaworski, T.; VAN Leuven, F. GSK3 and Alzheimer’s Disease: Facts and Fiction. Front. Mol. Neurosci. 2011, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- DaRocha-Souto, B.; Coma, M.; Pérez-Nievas, B.G.; Scotton, T.C.; Siao, M.; Sánchez-Ferrer, P.; Hashimoto, T.; Fan, Z.; Hudry, E.; Barroeta, I.; et al. Activation of glycogen synthase kinase-3 beta mediates β-amyloid induced neuritic damage in Alzheimer’s disease. Neurobiol. Dis. 2012, 45, 425–437. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.H.; Johnson, G.V. Glycogen synthase kinase 3beta phosphorylates tau at both primed and unprimed sites. Differential impact on microtubule binding. J. Biol. Chem. 2003, 278, 187–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asuni, A.A.; Hooper, C.; Reynolds, C.H.; Lovestone, S.; Anderton, B.H.; Killick, R. GSK3alpha exhibits beta-catenin and tau directed kinase activities that are modulated by Wnt. Eur. J. Neurosci. 2006, 24, 3387–3392. [Google Scholar] [CrossRef] [PubMed]
- Engel, T.; Goni-Oliver, P.; Lucas, J.J.; Avila, J.; Hernandez, F. Chronic lithium administration to FTDP-17 tau and GSK-3beta overexpressing mice prevents tau hyperphosphorylation and neurofibrillary tangle formation, but pre-formed neurofibrillary tangles do not revert. J. Neurochem. 2006, 99, 1445–1455. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.J.; Hernández, F.; Gómez-Ramos, P.; Morán, M.A.; Hen, R.; Avila, J. Decreased nuclear beta-catenin, tau hyperphosphorylation and neurodegeneration in GSK-3beta conditional transgenic mice. Embo J. 2001, 20, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Phiel, C.J.; Wilson, C.A.; Lee, V.M.; Klein, P.S. GSK-3alpha regulates production of Alzheimer’s disease amyloid-beta peptides. Nature 2003, 423, 435–439. [Google Scholar] [CrossRef]
- Sun, X.; Sato, S.; Murayama, O.; Murayama, M.; Park, J.M.; Yamaguchi, H.; Takashima, A. Lithium inhibits amyloid secretion in COS7 cells transfected with amyloid precursor protein C100. Neurosci. Lett. 2002, 321, 61–64. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT (min) | Mass (m/z) | Compound Identified | DB Formula | Compound Class |
|---|---|---|---|---|
| 0.58 | 175.9546 | Methylselenopyruvate | C4 H6 O3 Se | Oxo carboxylic acid |
| 0.58 | 150.0317 | Piperonal | C8 H6 O3 | Benzodioxoles |
| 0.635 | 103.0993 | 2-Amino-3-methyl-1-butanol | C5 H13 N O | Valinol |
| 0.648 | 196.0369 | Haematommic Acid | C9 H8 O5 | Amides |
| 0.650 | 360.318 | (R)-O-(3,4-Dihydroxycinnamoyl)-3-(3,4- dihydroxyphenyl)lactic acid (Rosmarinic acid) | C18 H16 O8 | Flavonoids |
| 0.651 | 265.1152 | D-1-[(3-Carboxypropyl)amino]-1-deoxyfructose | C10 H19 N O7 | Carboxylic acid |
| 0.651 | 404.0875 | Asp-Tyr-OH | C18 H16 N2 O9 | Amino acid |
| 0.654 | 309.1058 | N-Acetyl-a-neuraminic acid | C11 H19 N O9 | Sialic acid (antioxidants) |
| 0.663 | 117.0789 | Valine | C5 H11 N O2 | Amino acid |
| 0.673 | 115.0632 | 3-Acetamidopropanal | C5 H9 N O2 | Monocarboxylic acid amide |
| 0.684 | 174.0999 | Gly Val | C7 H14 N2 O3 | Amino acid |
| 0.686 | 192.0633 | Quinic acid | C7 H12 O6 | Cyclitol carboxylic acid (plant metabolite) |
| 0.693 | 232.1057 | Asp Val | C9 H16 N2 O5 | Amino acid |
| 0.716 | 125.0477 | 3-Hydroxyaminophenol | C6 H7 N O2 | Phenols |
| 0.717 | 304.1276 | 2′-Deoxymugineic acid | C12 H20 N2 O7 | Tricarboxylic acid |
| 0.717 | 135.0546 | Adenine | C5 H5 N5 | Amino acid |
| 0.753 | 117.0791 | Isoamyl nitrite | C5 H11 N O2 | Nitrites |
| 0.758 | 279.1308 | N-(1-Deoxy-1-fructosyl)valine | C11 H21 N O7 | Amino acid |
| 0.828 | 123.0321 | Isonicotinic acid | C6 H5 N O2 | Carboxylic acid |
| 0.882 | 208.094 | Ethyl beta-D-glucopyranoside | C8 H16 O6 | Glucoside |
| 0.883 | 162.0526 | 3-Hydroxy-3-methyl-glutaric acid | C6 H10 O5 | Carboxylic acid (plant metabolites) |
| 0.886 | 256.0589 | Piscidic Acid | C11 H12 O7 | Phenols |
| 0.892 | 204.0271 | Oxaloglutarate | C7 H8 O7 | Tricarboxylic acid |
| 0.939 | 271.1054 | Deidaclin | C12 H17 N O6 | Glycoside |
| 0.947 | 174.0162 | trans-Aconitate | C6 H6 O6 | Carboxylic acid anion (metabolite) |
| 0.947 | 146.0212 | Methyloxaloacetate | C5 H6 O5 | Dicarboxylic acid |
| 0.948 | 192.0271 | Citric acid | C6 H8 O7 | Tricarboxylic acid |
| 0.956 | 187.048 | 1-(Malonylamino)cyclopropanecarboxylic acid | C7 H9 N O5 | Carboxylic acid |
| 1.024 | 189.064 | L-2-Amino-6-oxoheptanedioate | C7 H11 N O5 | Oxo dicarboxylic acid |
| 1.025 | 171.053 | Tetrahydrodipicolinate | C7 H9 N O4 | Dicarboxylic acid anion |
| 1.038 | 293.1473 | N-(1-Deoxy-1-fructosyl)isoleucine | C12 H23 N O7 | Amino acid |
| 1.074 | 131.0945 | N,N-Diethylglycine | C6 H13 N O2 | Amino acid |
| 1.222 | 100.0164 | Succinic anhydride | C4 H4 O3 | Tetrahydrofurandione |
| 1.223 | 118.0263 | Erythrono-1,4-lactone | C4 H6 O4 | Lactone (butan-4-olide) |
| 1.381 | 131.0944 | L-Leucine | C6 H13 N O2 | Amino acid |
| 1.403 | 283.0918 | 8-hydroxy-2′-deoxy Guanosine | C10 H13 N5 O5 | Nucleoside |
| 1.452 | 127.0631 | Guvacine | C6 H9 N O2 | Amino acid |
| 1.453 | 145.074 | Isobutyrylglycine | C6 H11 N O3 | Carboxylic acid (N-acylglycine) |
| 2.075 | 327.1319 | N-(1-Deoxy-1-fructosyl)phenylalanine | C15 H21 N O7 | Monosaccharide derivative |
| 2.176 | 165.0784 | Gentiatibetine | C9 H11 N O2 | Alkaloids |
| 3.004 | 198.0526 | 2-Hydroxy-3,4-dimethoxybenzoic Acid | C9 H10 O5 | Phenolic acid |
| 3.005 | 152.0465 | p-Anisic acid | C8 H8 O3 | Phenolic acid |
| 5.57 | 162.0313 | 3-Hydroxycoumarin | C9 H6 O3 | Coumarin |
| 7.663 | 712.2231 | Isoliquiritigenin 4′-O-glucoside 4-O-apiofuranosyl-(1′′′->2′′′)-glucoside | C32 H40 O18 | Flavonoids |
| 7.742 | 684.1694 | Cosmosiin Hexaacetate | C33 H32 O16 | Phenols |
| 7.802 | 180.0416 | 4-Hydroxyphenylpyruvic acid | C9 H8 O4 | Carboxylic acid (oxo carboxylic acid) |
| 7.923 | 206.1302 | 2-Phenylethyl 3-methylbutanoate | C13 H18 O2 | Carboxylic ester |
| 7.944 | 656.1391 | Quercetagetin 4′-methyl ether 7-(6-(E)-caffeylglucoside) | C31 H28 O16 | Flavonoids |
| 7.993 | 517.1616 | Piperacillin | C23 H27 N5 O7S | Penicillin |
| 8.013 | 594.1598 | Luteolin 7-rhamnosyl(1->6)galactoside | C27 H30 O15 | Flavonoids |
| 8.442 | 226.1203 | 12-hydroxyjasmonic acid | C12 H18 O4 | Oxo carboxylic acid |
| 8.467 | 608.1184 | Prodelphinidin A1 | C30 H24 O14 | Flavonoids |
| 8.538 | 206.1301 | 2-Phenylethyl 3-methylbutanoate | C13 H18 O2 | Carboxylic ester |
| 8.582 | 596.1388 | Quercetin 3-alpha-arabinopyranosyl-(1->2)-glucoside | C26 H28 O16 | Flavonoid glycoside |
| 8.612 | 448.1012 | 6-Hydroxyluteolin 7-rhamnoside | C21 H20 O11 | Flavonoids |
| 8.702 | 464.0979 | Robinetin 7-glucoside | C21 H20 O12 | Flavonoids |
| 8.985 | 464.096 | 5,6,7,3′,4′-Pentahydroxy-8-methoxyflavone 7-apioside | C21 H20 O12 | Flavonoids |
| 9.062 | 294.0376 | Tricrozarin A | C13 H10 O8 | Quinone |
| 9.316 | 196.1096 | 4-(2-hydroxypropoxy)-3,5-dimethyl-Phenol | C11 H16 O3 | Phenols |
| 9.339 | 448.1013 | 6-Hydroxyluteolin 5-rhamnoside | C21 H20 O11 | Flavonoids |
| 9.365 | 720.1688 | Xanthochymuside | C36 H32 O16 | Flavonoids |
| 9.612 | 520.1583 | 5-Hydroxy-7,8,2′,3′-tetramethoxyflavone 5-glucoside | C25 H28 O12 | Flavonoids |
| 9.733 | 342.0744 | Iriskumaonin | C18 H14 O7 | Flavonoids |
| 9.734 | 162.0318 | 3-Hydroxycoumarin | C9 H6 O3 | Coumarins |
| 9.734 | 180.0424 | 4-Hydroxyphenylpyruvic acid | C9 H8 O4 | Phenols |
| 9.823 | 520.1575 | Quercetin 5,7,3′,4′-tetramethyl ether 3-galactoside | C25 H28 O12 | Flavonoids |
| 10.028 | 538.1122 | Lithospermic acid | C27 H22 O12 | Benzofuran |
| 10.028 | 718.1537 | Salvianolic acid L | C36 H30 O16 | Stilbenoids |
| 10.352 | 506.143 | Morin 3,7,4′-trimethyl ether 2′-glucoside | C24 H26 O12 | Flavonoids |
| 10.441 | 520.1012 | Melitric acid B | C27 H20 O11 | Cinnamic acids |
| 10.444 | 538.1113 | Melitric acid A | C27 H22 O12 | Cinnamic acids |
| 10.996 | 208.0736 | 2,5-Dimethoxycinnamic acid | C11 H12 O4 | Cinnamic acids |
| 11.032 | 330.0741 | Hypolaetin 8,3′-dimethyl ether | C17 H14 O7 | Flavonoids |
| 11.246 | 254.1879 | Kikkanol A | C15 H26 O3 | Sesquiterpenoid |
| 11.55 | 358.1057 | Corymbosin | C19 H18 O7 | Flavonoids |
| 11.664 | 374.1366 | (2S)-5,6,7,3′,4′-Pentamethoxyflavanone | C20 H22 O7 | Flavonoids |
| 11.783 | 328.0949 | Luteolin 7,3′,4′-trimethyl ether | C18 H16 O6 | Flavonoids |
| 12.256 | 342.1106 | 5,7-Dihydroxy-3′,4′-dimethoxy-6,8-dimethylflavone | C19 H18 O6 | Flavonoids |
| 12.306 | 272.2345 | 16-hydroxy hexadecanoic acid | C16 H32 O3 | Juniperic acid |
| 12.42 | 180.1145 | 3-tert-Butyl-5-methylcatechol | C11 H16 O2 | Phenols |
| 12.465 | 372.1213 | 7,8,3′,4′,5′-Pentamethoxyflavone | C20 H20 O7 | Flavnonoids |
| 12.706 | 344.09 | Wightin | C18 H16 O7 | Flavonoids |
| 12.764 | 314.0793 | Luteolin 5,3′-dimethyl ether | C17 H14 O6 | Flavonoids |
| 13.198 | 342.1108 | 5,7,2′,5′-tetramethoxyflavone | C19 H18 O6 | Flavonoids |
| 13.673 | 310.1784 | methyl 8-[2-(2-formyl-vinyl)-3-hydroxy-5-oxo-cyclopentyl]-octanoate | C17 H26 O5 | Long chain fatty acid |
| 14.622 | 328.0942 | Luteolin 7,3′,4′-trimethyl ether | C18 H16 O6 | Flavonoids |
| 21.342 | 390.2771 | 3α,12α-Dihydroxy-5β-chol-8(14)-en-24-oic Acid | C24 H38 O4 | Cholanoids |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Retinasamy, T.; Shaikh, M.F.; Kumari, Y.; Zainal Abidin, S.A.; Othman, I. Orthosiphon stamineus Standardized Extract Reverses Streptozotocin-Induced Alzheimer’s Disease-Like Condition in a Rat Model. Biomedicines 2020, 8, 104. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8050104
Retinasamy T, Shaikh MF, Kumari Y, Zainal Abidin SA, Othman I. Orthosiphon stamineus Standardized Extract Reverses Streptozotocin-Induced Alzheimer’s Disease-Like Condition in a Rat Model. Biomedicines. 2020; 8(5):104. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8050104
Chicago/Turabian StyleRetinasamy, Thaarvena, Mohd. Farooq Shaikh, Yatinesh Kumari, Syafiq Asnawi Zainal Abidin, and Iekhsan Othman. 2020. "Orthosiphon stamineus Standardized Extract Reverses Streptozotocin-Induced Alzheimer’s Disease-Like Condition in a Rat Model" Biomedicines 8, no. 5: 104. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8050104