Identification of Novel Native Autoantigens in Rheumatoid Arthritis

 , ,
, , 
Abstract
:1. Introduction
2. Experimental Section
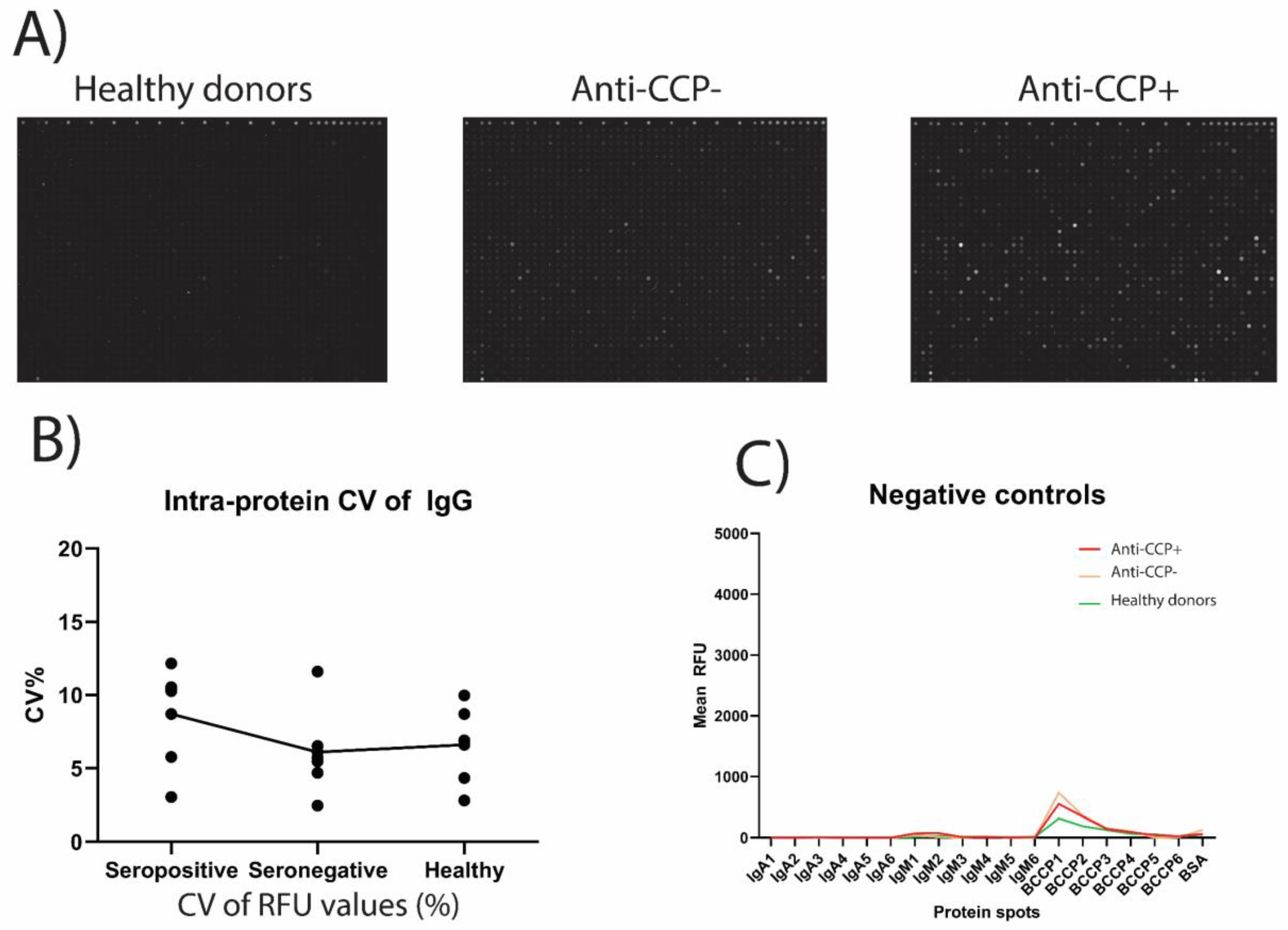
2.1. Collection of Plasma Samples
2.2. Protein Microarray Treatment
2.3. Extraction of Quantitative Expression Data
2.4. Data Analysis Pipeline
3. Results
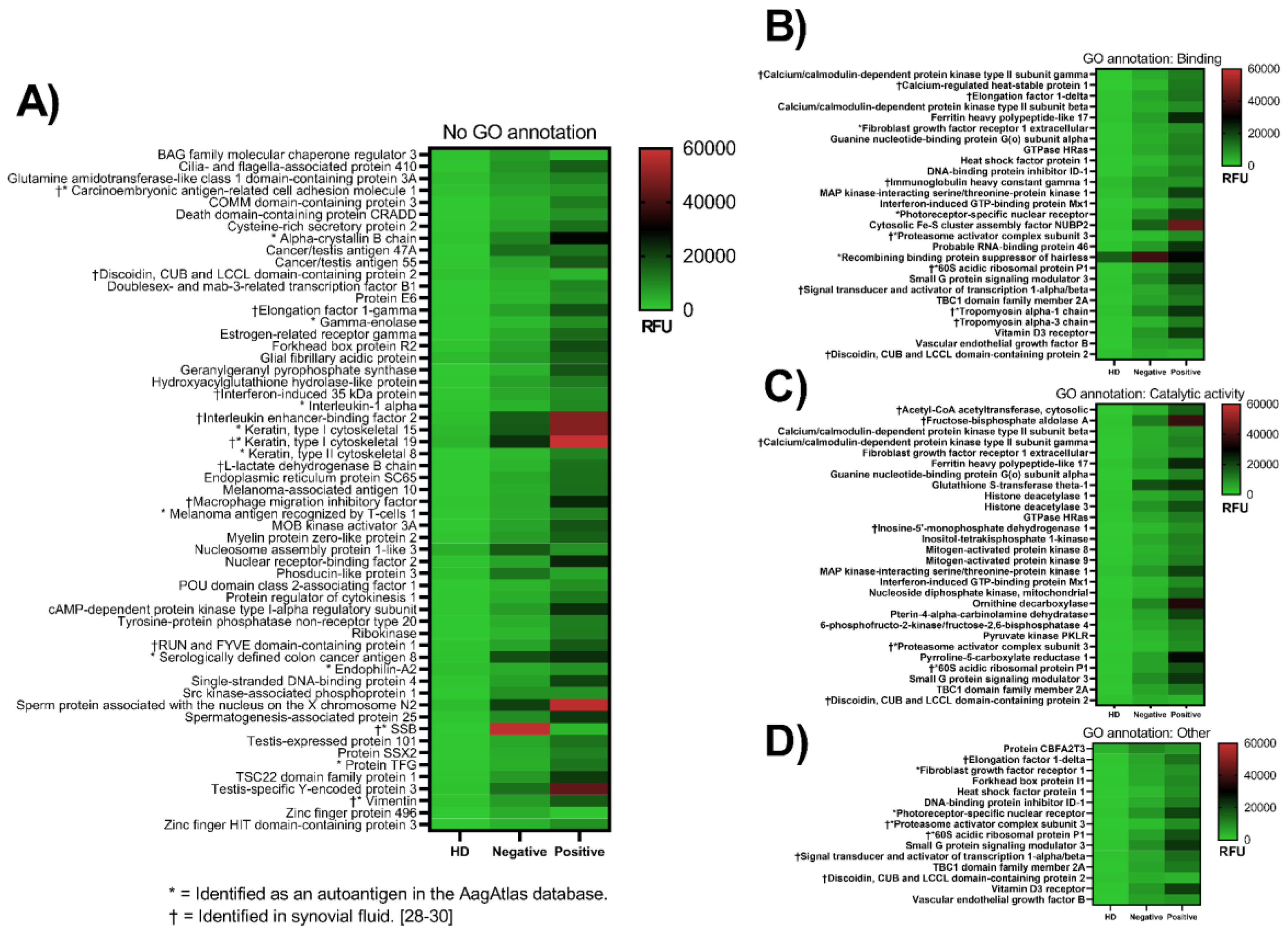
3.1. Identification of Native Autoantigens
3.2. Differences in Antibody Reactivity between Anti-CCP-Positive and Anti-CCP-Negative Plasma
3.3. Antigens Identified in Synovial Fluid
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smolen, J.S.; Aletaha, D.; Barton, A.; Burmester, G.R.; Emery, P.; Firestein, G.S.; Kavanaugh, A.; McInnes, I.B.; Solomon, D.H.; Strand, V.; et al. Rheumatoid arthritis. Nat. Rev. Dis. Primers 2018, 4, 18001. [Google Scholar] [CrossRef]
- Sirotti, S.; Generali, E.; Ceribelli, A.; Isailovic, N.; De Santis, M.; Selmi, C. Personalized medicine in rheumatology: The paradigm of serum autoantibodies. Autoimmun. Highlights 2017, 8. [Google Scholar] [CrossRef]
- Hu, Z.D.; Deng, A.M. Autoantibodies in pre-clinical autoimmune disease. Clin. Chim. Acta 2014, 437, 14–18. [Google Scholar] [CrossRef]
- Aggarwal, A. Role of autoantibody testing. Best Pract. Res. Clin. Rheumatol. 2014, 28, 907–920. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, K.; Barker, K.; Tang, Y.; Kahn, P.; Wiktor, P.; Brunner, A.; Knabben, V.; Takulapalli, B.; Buckner, J.; Nepom, G.; et al. A Contra Capture Protein Array Platform for Studying Post-translationally Modified (PTM) Auto-antigenomes. Mol. Cell. Proteom. 2016, 15, 2324–2337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Petritis, B.; Duan, H.; Xu, D.; LaBaer, J. Advances in cell-free protein array methods. Expert Rev. Proteom. 2018, 15, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Nielen, M.M.J.; van Schaardenburg, D.; Reesink, H.W.; van de Stadt, R.J.; van der Horst-Bruinsma, I.E.; de Koning, M.H.M.T.; Habibuw, M.R.; Vandenbroucke, J.P.; Dijkmans, B.A.C. Specific autoantibodies precede the symptoms of rheumatoid arthritis: A study of serial measurements in blood donors. Arthritis Rheum. 2004, 50, 380–386. [Google Scholar] [CrossRef]
- Vermeersch, P. Comparative Analysis of Different Approaches to Report Diagnostic Accuracy. Arch. Intern. Med. 2010, 170, 734. [Google Scholar] [CrossRef] [Green Version]
- Tenstad, H.B.; Nilsson, A.C.; Dellgren, C.D.; Lindegaard, H.M.; Rubin, K.H.; Lillevang, S.T. Use and utility of serologic tests for rheumatoid arthritis in primary care. Dan. Med. J. 2020, 67, A05190318. [Google Scholar]
- Van der Helm-van Mil, A.H.M.; Verpoort, K.N.; Breedveld, F.C.; Toes, R.E.M.; Huizinga, T.W.J. Antibodies to citrullinated proteins and differences in clinical progression of rheumatoid arthritis. Arthritis Res. Ther. 2005, 7, R949–R958. [Google Scholar] [CrossRef] [Green Version]
- Harris, M.L.; Darrah, E.; Lam, G.K.; Bartlett, S.J.; Giles, J.T.; Grant, A.V.; Gao, P.; Scott, W.W.; El-Gabalawy, H.; Casciola-Rosen, L.; et al. Association of autoimmunity to peptidyl arginine deiminase type 4 with genotype and disease severity in rheumatoid arthritis. Arthritis Rheum. 2008, 58, 1958–1967. [Google Scholar] [CrossRef] [PubMed]
- Schellekens, G.A.; Visser, H.; de Jong, B.A.; van den Hoogen, F.H.; Hazes, J.M.; Breedveld, F.C.; van Venrooij, W.J. The diagnostic properties of rheumatoid arthritis antibodies recognizing a cyclic citrullinated peptide. Arthritis Rheum. 2000, 43, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Nell-Duxneuner, V.; Machold, K.; Stamm, T.; Eberl, G.; Heinzl, H.; Hoefler, E.; Smolen, J.S.; Steiner, G. Autoantibody profiling in patients with very early rheumatoid arthritis: A follow-up study. Ann. Rheum. Dis. 2010, 69, 169–174. [Google Scholar] [CrossRef]
- Darrah, E.; Giles, J.T.; Ols, M.L.; Bull, H.G.; Andrade, F.; Rosen, A. Erosive Rheumatoid Arthritis Is Associated with Antibodies That Activate PAD4 by Increasing Calcium Sensitivity. Sci. Transl. Med. 2013, 5, 186ra65. [Google Scholar] [CrossRef] [Green Version]
- Bonifacio, A.F.; Alunno, A.; La Paglia, G.M.C.; Valentini, E.; Leone, M.C.; Bartoloni, E.; Gerli, R. Novel autoantibodies in rheumatoid arthritis. Reumatismo 2019, 71, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Damgaard, D.; Bawadekar, M.; Senolt, L.; Stensballe, A.; Shelef, M.A.; Nielsen, C.H. Relative efficiencies of peptidylarginine deiminase 2 and 4 in generating target sites for anti-citrullinated protein antibodies in fibrinogen, alpha-enolase and histone H3. PLoS ONE 2018, 13, e0203214. [Google Scholar] [CrossRef]
- Konig, M.F.; Giles, J.T.; Nigrovic, P.A.; Andrade, F. Antibodies to native and citrullinated RA33 (hnRNP A2/B1) challenge citrullination as the inciting principle underlying loss of tolerance in rheumatoid arthritis. Ann. Rheum. Dis. 2016, 75, 2022–2028. [Google Scholar] [CrossRef]
- Schellekens, G.A.; de Jong, B.A.; van den Hoogen, F.H.; van de Putte, L.B.; van Venrooij, W.J. Citrulline is an essential constituent of antigenic determinants recognized by rheumatoid arthritis-specific autoantibodies. J. Clin. Investig. 1998, 101, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Wright, H.L.; Moots, R.J.; Edwards, S.W. The multifactorial role of neutrophils in rheumatoid arthritis. Nat. Rev. Rheumatol. 2014, 10, 593–601. [Google Scholar] [CrossRef]
- Zheng, Z.; Mergaert, A.M.; Fahmy, L.M.; Bawadekar, M.; Holmes, C.L.; Ong, I.M.; Bridges, A.J.; Newton, M.A.; Shelef, M.A. Disordered Antigens and Epitope Overlap Between Anti-Citrullinated Protein Antibodies and Rheumatoid Factor in Rheumatoid Arthritis. Arthritis Rheumatol. 2020, 72, 262–272. [Google Scholar] [CrossRef]
- Liew, J.; Amir, A.; Chen, Y.; Fong, M.Y.; Razali, R.; Lau, Y.L. Autoantibody profile of patients infected with knowlesi malaria. Clin. Chim. Acta 2015, 448, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Wallstrom, G.; Yu, X.; Hopper, M.; Van Duine, J.; Steel, J.; Park, J.; Wiktor, P.; Kahn, P.; Brunner, A.; et al. Identification of Antibody Targets for Tuberculosis Serology using High-Density Nucleic Acid Programmable Protein Arrays. Mol. Cell. Proteom. 2017, 16, S277–S289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumera, A.; Anuar, N.D.; Radhakrishnan, A.K.; Ibrahim, H.; Rutt, N.H.; Ismail, N.H.; Tan, T.-M.; Baba, A.A. A Novel Method to Identify Autoantibodies against Putative Target Proteins in Serum from beta-Thalassemia Major: A Pilot Study. Biomedicines 2020, 8, 97. [Google Scholar] [CrossRef] [PubMed]
- Duarte, J.; Serufuri, J.; Mulder, N.; Blackburn, J. Bioinformatics of Human Proteomics; Springer: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Suwarnalata, G.; Tan, A.H.; Isa, H.; Gudimella, R.; Anwar, A.; Loke, M.F.; Mahadeva, S.; Lim, S.-Y.; Vadivelu, J. Augmentation of Autoantibodies by Helicobacter pylori in Parkinson’s Disease Patients May Be Linked to Greater Severity. PLoS ONE 2016, 11, e0153725. [Google Scholar] [CrossRef] [Green Version]
- Soe, H.J.; Yong, Y.K.; Al-Obaidi, M.M.J.; Raju, C.S.; Gudimella, R.; Manikam, R.; Sekaran, S.D. Identifying protein biomarkers in predicting disease severity of dengue virus infection using immune-related protein microarray. Medicine 2018, 97, 6–12. [Google Scholar] [CrossRef]
- Wang, D.; Yang, L.; Zhang, P.; LaBaer, J.; Hermjakob, H.; Li, D.; Yu, X. AAgAtlas 1.0: A human autoantigen database. Nucleic Acids Res. 2017, 45, D769–D776. [Google Scholar] [CrossRef] [Green Version]
- Bennike, T.; Ayturk, U.; Haslauer, C.M.; Froehlich, J.W.; Proffen, B.L.; Barnaby, O.; Birkelund, S.; Murray, M.M.; Warman, M.L.; Stensballe, A.; et al. A Normative Study of the Synovial Fluid Proteome from Healthy Porcine Knee Joints. J. Proteome Res. 2014, 13, 4377–4387. [Google Scholar] [CrossRef]
- Bhattacharjee, M.; Balakrishnan, L.; Renuse, S.; Advani, J.; Goel, R.; Sathe, G.; Keshava, P.T.S.; Nair, B.; Jois, R.; Shankar, S.; et al. Synovial fluid proteome in rheumatoid arthritis. Clin. Proteom. 2016, 13, 12. [Google Scholar] [CrossRef] [Green Version]
- Mahendran, S.M.; Keystone, E.C.; Krawetz, R.J.; Liang, K.; Diamandis, E.P.; Chandran, V. Elucidating the endogenous synovial fluid proteome and peptidome of inflammatory arthritis using label-free mass spectrometry. Clin. Proteom. 2019, 16, 23. [Google Scholar] [CrossRef] [Green Version]
- Tutturen, A.E.V.; Fleckenstein, B.; De Souza, G. Assessing the Citrullinome in Rheumatoid Arthritis Synovial Fluid with and without Enrichment of Citrullinated Peptides. J. Proteome Res. 2014, 13, 2867–2873. [Google Scholar] [CrossRef]
- Avrameas, S. Natural autoantibodies: From ‘horror autotoxicus’ to ‘gnothi seauton’. Immunol. Today 1991, 12, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, A.; Srivastava, R.; Agrawal, S.; Srivastava, R.K. T cell responses to citrullinated self-peptides in patients with rheumatoid arthritis. Rheumatol. Int. 2013, 33, 2359–2363. [Google Scholar] [CrossRef] [PubMed]
- Jouvenne, P.; Garrone, P.; Djossou, O.; Banchereau, J.; Miossec, P. Increased incidence of neutralizing autoantibodies against interleukin-1? (IL-1?) in nondestructive chronic polyarthritis. J. Clin. Immunol. 1996, 16, 283–290. [Google Scholar] [CrossRef]
- Wang, X.; Chen, P.; Cui, J.; Yang, C.; Du, H. Keratin 8 is a novel autoantigen of rheumatoid arthritis. Biochem. Biophys. Res. Commun. 2015, 465, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Snir, O.; Widhe, M.; Hermansson, M.; Von Spee, C.; Lindberg, J.; Hensen, S.; Lundberg, K.; Engström, Å.; Venables, P.J.W.; Toes, R.; et al. Antibodies to several citrullinated antigens are enriched in the joints of rheumatoid arthritis patients. Arthritis Rheum. 2010, 62, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Kubuschok, B.; Preuss, K.-D.; Baier-Thoenes, K.; Regitz, E.; Thurner, L.; Assmann, G.; Pfreundschuh, M. Autoantibodies against lamin C, NA14 and CK15 in primary vasculitides or autoimmune diseases with secondary vasculitis. Clin. Exp. Rheumatol. 2016, 34. [Google Scholar]
- Van Noort, J.; Bsibsi, M.; Gerritsen, W.H.; Van Der Valk, P.; Bajramovic, J.; Steinman, L.; Amor, S. αB-Crystallin Is a Target for Adaptive Immune Responses and a Trigger of Innate Responses in Preactive Multiple Sclerosis Lesions. J. Neuropathol. Exp. Neurol. 2010, 69, 694–703. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.-H.; Nahm, D.-H.; Kim, S.-H.; Kim, Y.-S.; Suh, C.-H.; Park, H.-S.; Ahn, S.-W. Increased levels of IgG to cytokeratin 19 in sera of patients with toluene diisocyanate-induced asthma. Ann. Allergy Asthma Immunol. 2004, 93, 293–298. [Google Scholar] [CrossRef]
- Matsutani, T.; Hiwasa, T.; Takiguchi, M.; Oide, T.; Kunimatsu, M.; Saeki, N.; Iwadate, Y. Autologous antibody to src-homology 3-domain GRB2-like 1 specifically increases in the sera of patients with low-grade gliomas. J. Exp. Clin. Cancer Res. 2012, 31, 85. [Google Scholar] [CrossRef] [Green Version]
- Manoussakis, M.N.; Kistis, C.G.; Aidinis, V.; Guialis, A.; Piha, L.; Sekeris, C.E.; Moutsopoulos, H.M. Detection of human-specific anti-La(SSB) antibodies in patients with rheumatoid arthritis. J. Autoimmun. 1995, 8, 959–969. [Google Scholar] [CrossRef]
- Skopouli, F.N.; Andonopoulos, A.P.; Moutsopoulos, H.M. Clinical implications of the presence of anti-Ro(SSA) antibodies in patients with rheumatoid arthritis. J. Autoimmun. 1988, 1, 381–388. [Google Scholar] [CrossRef]
- Garg, A.; Barnes, P.F.; Porgador, A.; Roy, S.; Wu, S.; Nanda, J.S.; Griffith, D.E.; Girard, W.M.; Rawal, N.; Shetty, S.; et al. Vimentin expressed on Mycobacterium tuberculosis-infected human monocytes is involved in binding to the NKp46 receptor. J. Immunol. 2006, 177, 6192–6198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mor-Vaknin, N.; Punturieri, A.; Sitwala, K.; Markovitz, D.M. Vimentin is secreted by activated macrophages. Nat. Cell Biol. 2002, 5, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Goto, H.; Tanabe, K.; Manser, E.; Lim, L.; Yasui, Y.; Inagaki, M. Phosphorylation and reorganization of vimentin by p21-activated kinase (PAK). Genes Cells 2002, 7, 91–97. [Google Scholar] [CrossRef]
- Bornheim, R.; Reuter, U.; Herrmann, H.; Büssow, H.; Magin, T.M.; Müller, M. A dominant vimentin mutant upregulates Hsp70 and the activity of the ubiquitin-proteasome system, and causes posterior cataracts in transgenic mice. J. Cell Sci. 2008, 121, 3737–3746. [Google Scholar] [CrossRef] [Green Version]
- Ospelt, C.; Bang, H.; Feist, E.; Camici, G.G.; Keller, S.; Detert, J.; Krämer, A.; Gay, S.; Ghannam, K.; Burmester, G.R. Carbamylation of vimentin is inducible by smoking and represents an independent autoantigen in rheumatoid arthritis. Ann. Rheum. Dis. 2017, 76, 1176–1183. [Google Scholar] [CrossRef] [Green Version]
- Van Steendam, K.; Tilleman, K.; Deforce, D. The relevance of citrullinated vimentin in the production of antibodies against citrullinated proteins and the pathogenesis of rheumatoid arthritis. Rheumatology 2011, 50, 830–837. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Jia, R.; Liu, Y.; Tang, S.; Ma, X.; Shi, L.; Zhao, J.; Hu, F.; Li, Z. Antibodies against carbamylated vimentin exist in systemic lupus erythematosus and correlate with disease activity. Lupus 2020, 29, 239–247. [Google Scholar] [CrossRef]
- Bay-Jensen, A.C.; Karsdal, M.A.; Vassiliadis, E.; Wichuk, S.; Marcher-Mikkelsen, K.; Lories, R.; Christiansen, C.; Maksymowych, W.P. Circulating Citrullinated Vimentin Fragments Reflect Disease Burden in Ankylosing Spondylitis and Have Prognostic Capacity for Radiographic Progression. Arthritis Rheum. 2013, 65, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Mansour, H.; Metwaly, K.M.; Hassan, I.A.; ElShamy, H.-A.A.; Elbeblawy, M.M. Antibodies to Mutated Citrullinated Vimentin in Rheumatoid Arthritis: Diagnostic Value, Association with Radiological Damage and Axial Skeleton Affection. Clin. Med. Insights: Arthritis Musculoskelet. Disord. 2010, 3, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Mathsson, L.; Mullazehi, M.; Wick, M.C.; Sjöberg, O.; Van Vollenhoven, R.; Klareskog, L.; Rönnelid, J. Antibodies against citrullinated vimentin in rheumatoid arthritis: Higher sensitivity and extended prognostic value concerning future radiographic progression as compared with antibodies against cyclic citrullinated peptides. Arthritis Rheum. 2007, 58, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Bang, H.; Egerer, K.; Gauliard, A.; Lüthke, K.; Rudolph, P.E.; Fredenhagen, G.; Berg, W.; Feist, E.; Burmester, G.-R. Mutation and citrullination modifies vimentin to a novel autoantigen for rheumatoid arthritis. Arthritis Rheum. 2007, 56, 2503–2511. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-N.; Nie, L.-Y.; Lu, X.-Y.; Wu, H.-X. Meta-analysis: Compared with anti-CCP and rheumatoid factor, could anti-MCV be the next biomarker in the rheumatoid arthritis classification criteria? Clin. Chem. Lab. Med. 2019, 57, 1668–1679. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Gao, S.; Luo, G.; Cheng, G.; Huang, W.; Jiang, R.; Wang, Y.; Cui, T. Increased Circulating Autoantibodies Levels of IgG, IgA, IgM Against Cytokeratin 18 and Cytokeratin 19 in Chronic Obstructive Pulmonary Disease. Arch. Med. Res. 2017, 48, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Linn-Rasker, S.P.; Mil, A.V.D.H.-V.; Van Gaalen, F.A.; Kloppenburg, M.; De Vries, R.R.P.; Le Cessie, S.; Breedveld, F.C.; Toes, R.; Huizinga, T.W.J. Smoking is a risk factor for anti-CCP antibodies only in rheumatoid arthritis patients who carry HLA-DRB1 shared epitope alleles. Ann. Rheum. Dis. 2005, 65, 366–371. [Google Scholar] [CrossRef] [Green Version]
- Vincent, T. Fibroblast growth factor 2: Good or bad guy in the joint? Arthritis Res. Ther. 2011, 13, 127. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.; Sun, D.; Li, H.; Li, X.; Pan, W.; Yan, C.; Tang, R.; Liu, X. The Effect of SHH-Gli Signaling Pathway on the Synovial Fibroblast Proliferation in Rheumatoid Arthritis. Inflammation 2015, 39, 503–512. [Google Scholar] [CrossRef]
- Sun, H.-J.; Cai, W.; Gong, L.-L.; Wang, X.; Zhu, X.; Wan, M.-Y.; Wang, P.-Y.; Qiu, L. FGF-2-mediated FGFR1 signaling in human microvascular endothelial cells is activated by vaccarin to promote angiogenesis. Biomed. Pharmacother. 2017, 95, 144–152. [Google Scholar] [CrossRef]
- Paleolog, E. Angiogenesis in rheumatoid arthritis. Arthritis Res. 2002, 4, S81–S90. [Google Scholar] [CrossRef]
- Yan, D.; Chen, D.; Cool, S.M.; Van Wijnen, A.J.; Mikecz, K.; Murphy, G.; Im, H.-J. Fibroblast growth factor receptor 1 is principally responsible for fibroblast growth factor 2-induced catabolic activities in human articular chondrocytes. Arthritis Res. Ther. 2011, 13, R130. [Google Scholar] [CrossRef] [Green Version]
- Provost, T.T. Anti-Ro(SSA)and Anti-La(SSB)Antibodies in Lupus Erythematosus and Sjoegren’s Syndrome. Keio J. Med. 1991, 40, 72–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Türkçapar, N.; Demir, Ö.; Atli, T.; Kopuk, M.; Turgay, M.; Kinikli, G.; Duman, M. Late onset rheumatoid arthritis: Clinical and laboratory comparisons with younger onset patients. Arch. Gerontol. Geriatr. 2006, 42, 225–231. [Google Scholar] [CrossRef]
- Panush, R.S.; Edwards, N.L.; Longley, S.; Webster, E. “Rhupus” syndrome. Arch. Intern. Med. 1988, 148, 1633–1636. [Google Scholar] [CrossRef] [PubMed]
- Nordström, E.; Möller, E.; Abedi-Valugerdi, M. Induction of IgG Rheumatoid Factor (RF) Production by Antibody–antibody (RF-like) Immune Complexes: The Role of T cells, Complement and Fcγ Receptors. J. Autoimmun. 1998, 11, 131–140. [Google Scholar] [CrossRef]
- Seegobin, S.D.; Ma, M.H.; Dahanayake, C.; Cope, A.; Scott, D.L.; Lewis, C.M.; Scott, I.C. ACPA-positive and ACPA-negative rheumatoid arthritis differ in their requirements for combination DMARDs and corticosteroids: Secondary analysis of a randomized controlled trial. Arthritis Res. Ther. 2014, 16, R13. [Google Scholar] [CrossRef] [Green Version]
- Schmid, K.; Macnair, M.B. CHARACTERIZATION OF THE PROTEINS OF CERTAIN POSTMORTEM HUMAN SYNOVIAL FLUIDS12. J. Clin. Investig. 1958, 37, 708–718. [Google Scholar] [CrossRef]
- Schmid, K.; Macnair, M.B. CHARACTERIZATION OF THE PROTEINS OF HUMAN SYNOVIAL FLUID IN CERTAIN DISEASE STATES 1. J. Clin. Investig. 1956, 35, 814–824. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Romero, C.; Calamia, V.; Carreira, V.; Mateos, J.; Fernandez-Puente, P.; Blanco, F.J.; Blanco, F.J. Strategies to optimize two-dimensional gel electrophoresis analysis of the human joint proteome. Talanta 2010, 80, 1552–1560. [Google Scholar] [CrossRef]
- Kinloch, A.; Lundberg, K.; Wait, R.; Wegner, N.; Lim, N.H.; Zendman, A.J.W.; Saxne, T.; Malmstr, V.; Venables, P.J.; Kinloch, A.J. Synovial fluid is a site of citrullination of autoantigens in inflammatory arthritis. Arthritis Rheum. 2008, 58, 2287–2295. [Google Scholar] [CrossRef]
- Van De Putte, L.B.A.; Atkins, C.; Malaise, M.; Sany, J.; Russell, A.S.; Van Riel, P.L.C.M.; Settas, L.; Bijlsma, J.W.; Todesco, S.; Dougados, M.; et al. Efficacy and safety of adalimumab as monotherapy in patients with rheumatoid arthritis for whom previous disease modifying antirheumatic drug treatment has failed. Ann. Rheum. Dis. 2004, 63, 508–516. [Google Scholar] [CrossRef]
- Cohen, S.B.; Emery, P.; Greenwald, M.W.; Dougados, M.; Furie, R.A.; Genovese, M.C.; Keystone, E.C.; Loveless, J.E.; Burmester, G.-R.; Cravets, M.W.; et al. Rituximab for rheumatoid arthritis refractory to anti–tumor necrosis factor therapy: Results of a multicenter, randomized, double-blind, placebo-controlled, phase III trial evaluating primary efficacy and safety at twenty-four weeks. Arthritis Rheum. 2006, 54, 2793–2806. [Google Scholar] [CrossRef] [PubMed]
- Wijbrandts, C.; Tak, P. Prediction of Response to Targeted Treatment in Rheumatoid Arthritis. Mayo Clin. Proc. 2017, 92, 1129–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tak, P.-P. A personalized medicine approach to biologic treatment of rheumatoid arthritis: A preliminary treatment algorithm. Rheumatology 2011, 51, 600–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thurlings, R.M.; Wijbrandts, C.A.; Mebius, R.E.; Cantaert, T.; Dinant, H.J.; Van Der Pouw-Kraan, T.C.T.M.; Verweij, C.L.; Baeten, M.; Tak, P.-P. Synovial lymphoid neogenesis does not define a specific clinical rheumatoid arthritis phenotype. Arthritis Rheum. 2008, 58, 1582–1589. [Google Scholar] [CrossRef] [PubMed]
- Klimiuk, P.A.; Goronzy, J.J.; Nsson, J.B.; Beckenbaugh, R.D.; Weyand, C.M. Tissue cytokine patterns distinguish variants of rheumatoid synovitis. Am. J. Pathol. 1997, 151, 1311–1319. [Google Scholar] [PubMed]
- Ulfgren, A.-K.; Gröndal, L.; Lindblad, S.; Khademi, M.; Johnell, O.; Klareskog, L.; Andersson, U. Interindividual and intra-articular variation of proinflammatory cytokines in patients with rheumatoid arthritis: Potential implications for treatment. Ann. Rheum. Dis. 2000, 59, 439–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Baarsen, L.; Wijbrandts, C.A.; Timmer, T.C.G.; Kraan, T.C.T.M.V.D.P.; Tak, P.P.; Verweij, C.L. Synovial tissue heterogeneity in rheumatoid arthritis in relation to disease activity and biomarkers in peripheral blood. Arthritis Rheum. 2010, 62, 1602–1607. [Google Scholar] [CrossRef]
- Galligan, C.L.; Baig, E.; Bykerk, V.; Keystone, E.C.; Fish, E.N. Distinctive gene expression signatures in rheumatoid arthritis synovial tissue fibroblast cells: Correlates with disease activity. Genes Immun. 2007, 8, 480–491. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Gene Name | Protein Name | Localization † |
|---|---|---|
| IGHG1 (IgG1) | Immunoglobulin heavy constant gamma 1 | Extracellular/cell associated |
| ACAT2 | Acetyl-CoA acetyltransferase | Intracellular and extracellular |
| ALDOA | Fructose-bisphosphate aldolase A | Intracellular and extracellular |
| CEACAM1 | Carcinoembryonic antigen-related cell adhesion molecule 1 | Intracellular and extracellular |
| EEF1G | Elongation factor 1-gamma | Intracellular and extracellular |
| IMPDH1 | Inosine-5’-monophosphate dehydrogenase 1 | Intracellular and extracellular |
| KRT19 | Keratin 19 | Intracellular and extracellular |
| LDHB | L-lactate dehydrogenase B chain | Intracellular and extracellular |
| MIF | Macrophage migration inhibitory factor | Intracellular and extracellular |
| TKT | Transketolase | Intracellular and extracellular |
| TPM3 | Tropomyosin alpha-3 chain | Intracellular and extracellular |
| VIM | Vimentin | Intracellular and extracellular |
| CAMK2G | Calcium/calmodulin-dependent protein kinase type II subunit gamma | Intracellular |
| CARHSP1 | Calcium-regulated heat-stable protein 1 | Intracellular |
| EEF1D | Elongation factor 1-delta | Intracellular |
| IFI35 | Interferon-induced 35 kDa protein | Intracellular |
| ILF2 | Interleukin enhancer-binding factor 2 | Intracellular |
| PRKAR1A | cAMP-dependent protein kinase type I-alpha regulatory subunit | Intracellular |
| PSME3 | Proteasome activator complex subunit 3 | Intracellular |
| RPLP1 | 60S acidic ribosomal protein P1 | Intracellular |
| RUFY1 | RUN and FYVE domain-containing protein 1 | Intracellular |
| SSB | Lupus La protein / SSB | Intracellular |
| STAT1 | Signal transducer and activator of transcription 1-alpha/beta | Intracellular |
| TPM1 | Tropomyosin alpha-1 chain | Intracellular |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poulsen, T.B.G.; Damgaard, D.; Jørgensen, M.M.; Senolt, L.; Blackburn, J.M.; Nielsen, C.H.; Stensballe, A. Identification of Novel Native Autoantigens in Rheumatoid Arthritis. Biomedicines 2020, 8, 141. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8060141
Poulsen TBG, Damgaard D, Jørgensen MM, Senolt L, Blackburn JM, Nielsen CH, Stensballe A. Identification of Novel Native Autoantigens in Rheumatoid Arthritis. Biomedicines. 2020; 8(6):141. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8060141
Chicago/Turabian StylePoulsen, Thomas B. G., Dres Damgaard, Malene Møller Jørgensen, Ladislav Senolt, Jonathan M. Blackburn, Claus H. Nielsen, and Allan Stensballe. 2020. "Identification of Novel Native Autoantigens in Rheumatoid Arthritis" Biomedicines 8, no. 6: 141. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8060141