Probiotics-Containing Yogurt Ingestion and H. pylori Eradication Can Restore Fecal Faecalibacterium prausnitzii Dysbiosis in H. pylori-Infected Children


Abstract
:1. Introduction
2. Results
2.1. Prevalence of H. pylori Infection
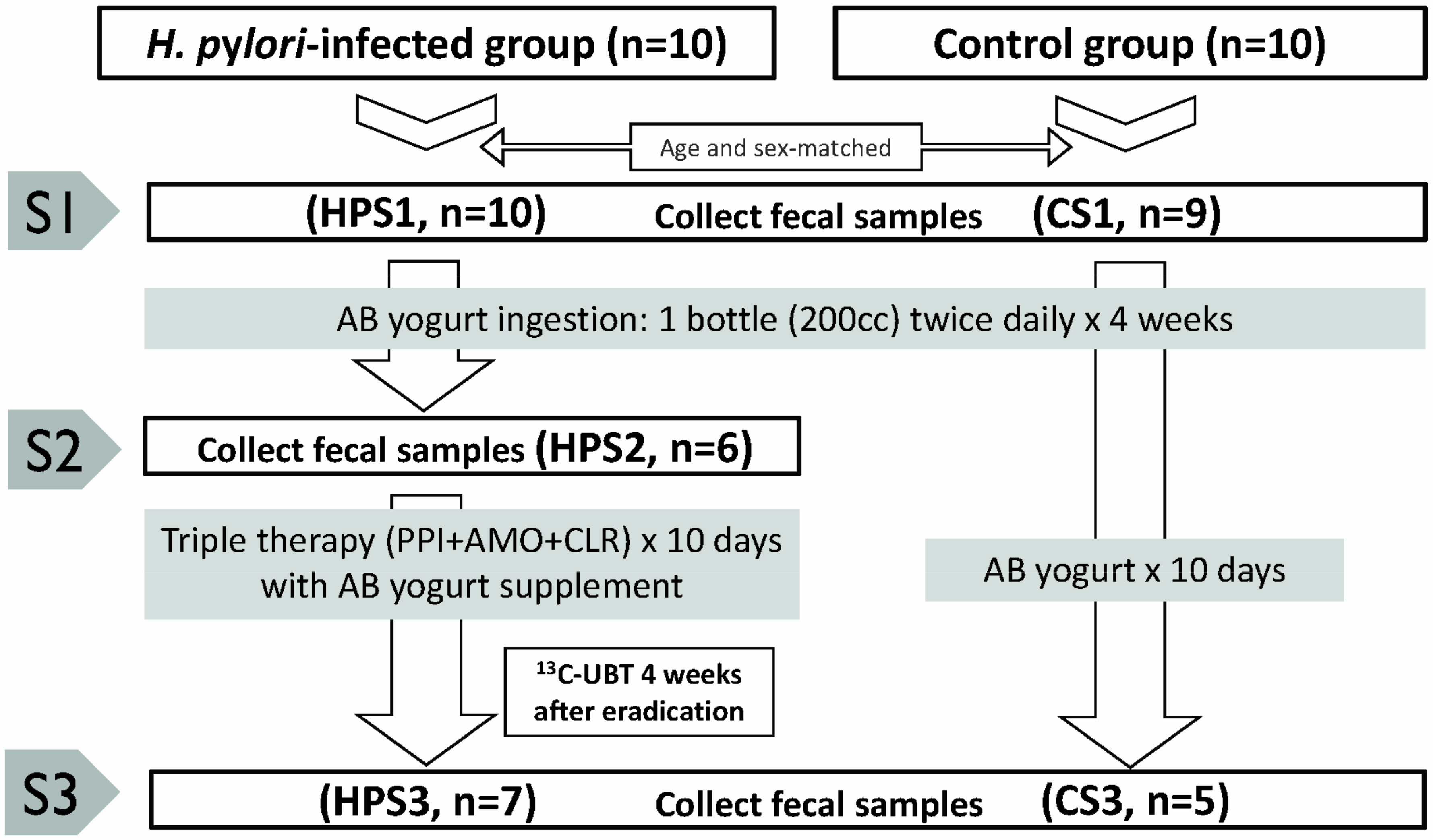
2.2. Numbers of Study Cases and Fecal Samples
2.3. Fecal Inflammatory Parameters between H. pylori-Infected Children and Controls
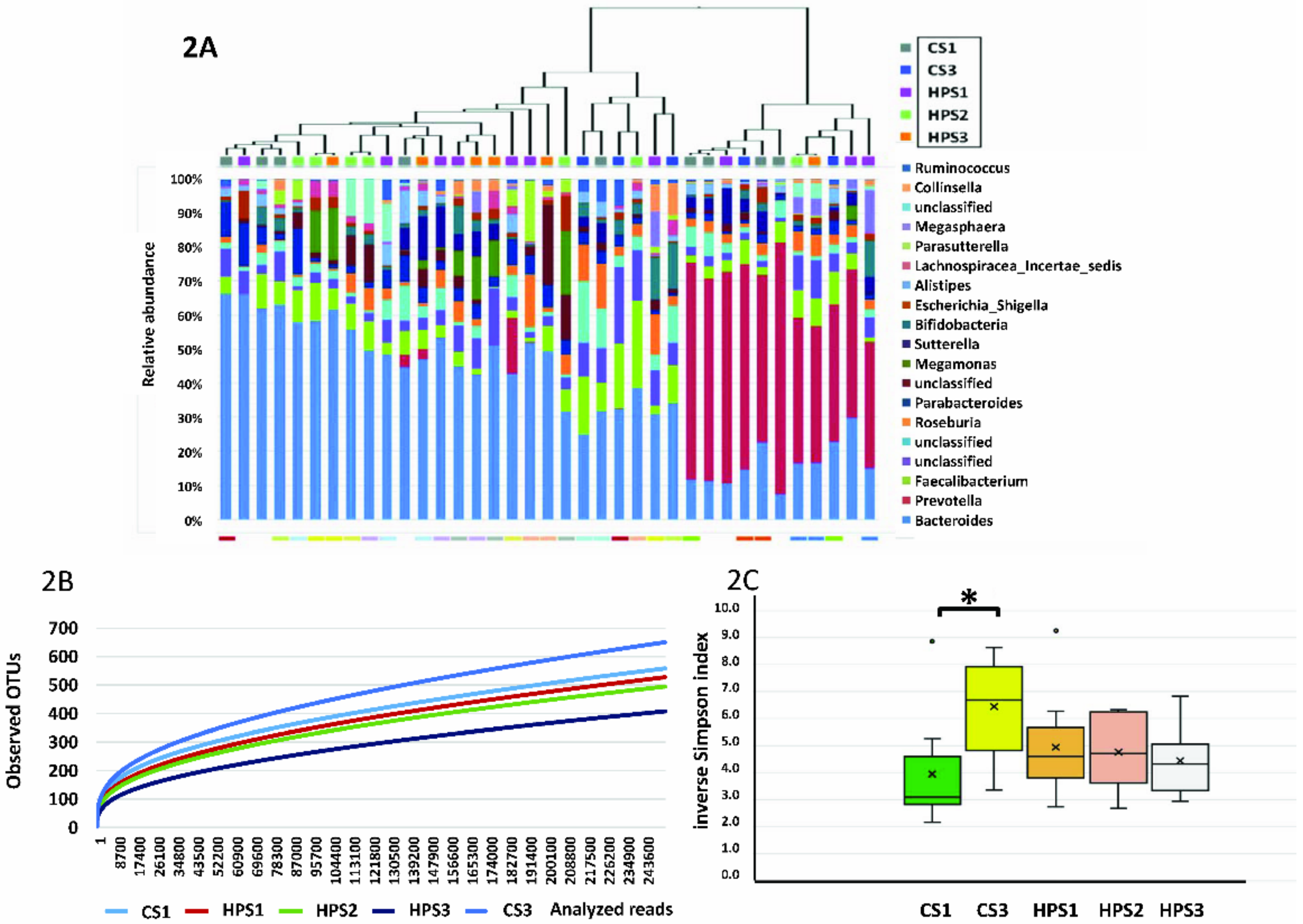
2.4. Sequencing Results
2.5. Library Coverage and Sequence Diversity
2.6. Significant Genus Difference between H. pylori-Infected Children and Controls
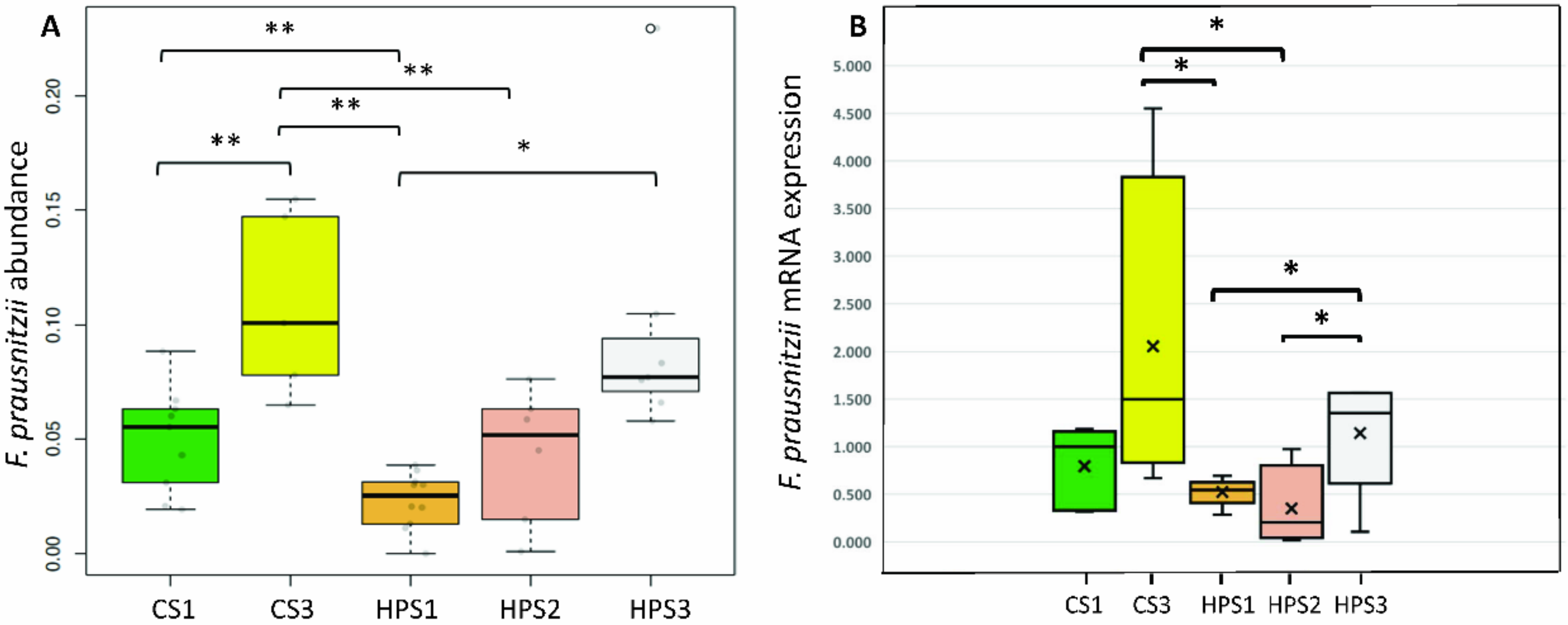
2.7. Serial Changes in the Abundance of F. prausnitzii after Yogurt Ingestion and H. pylori Eradication
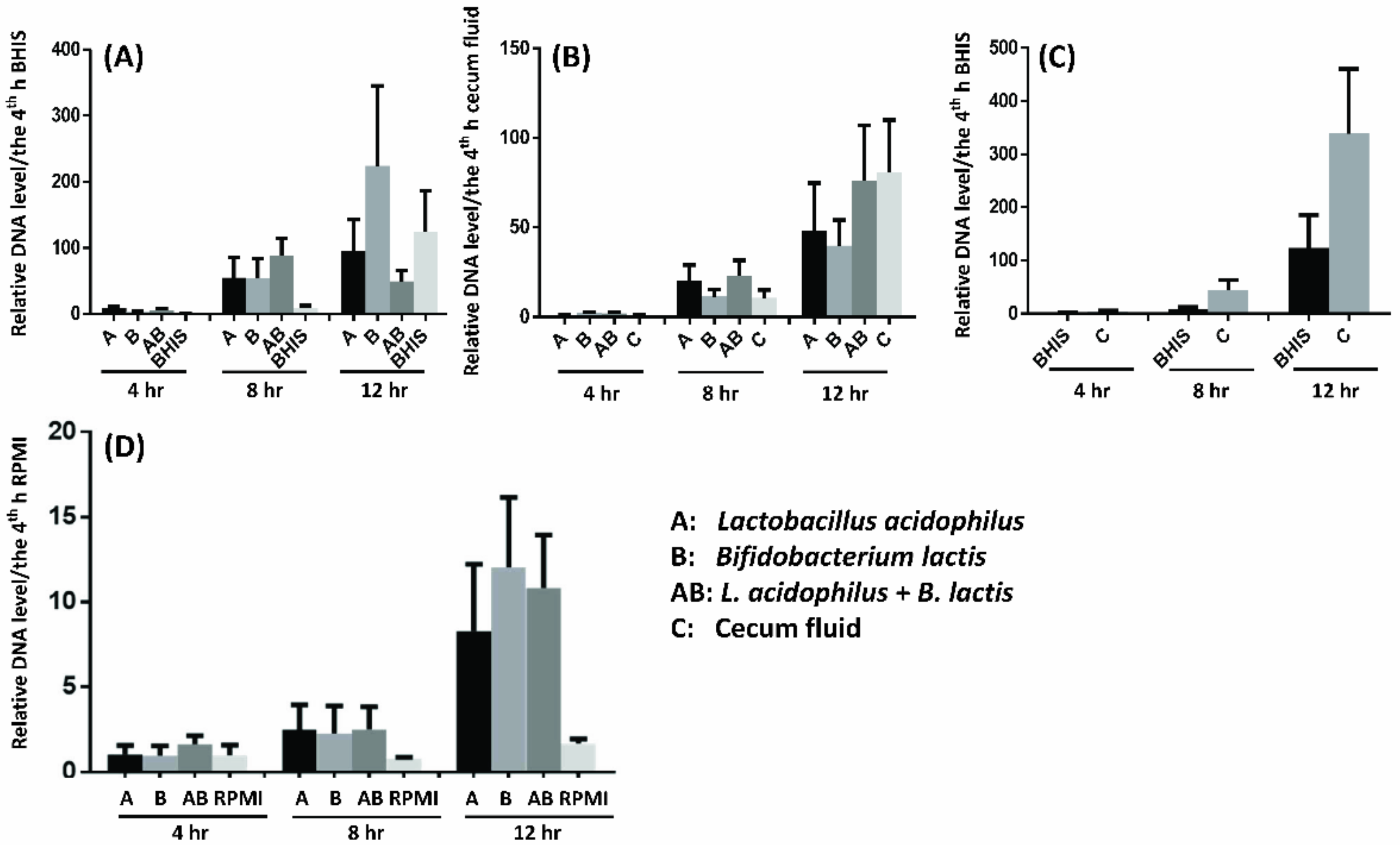
2.8. Probiotics Facilitate F. prausnitzii Growth in Vitro and In Ex Vivo
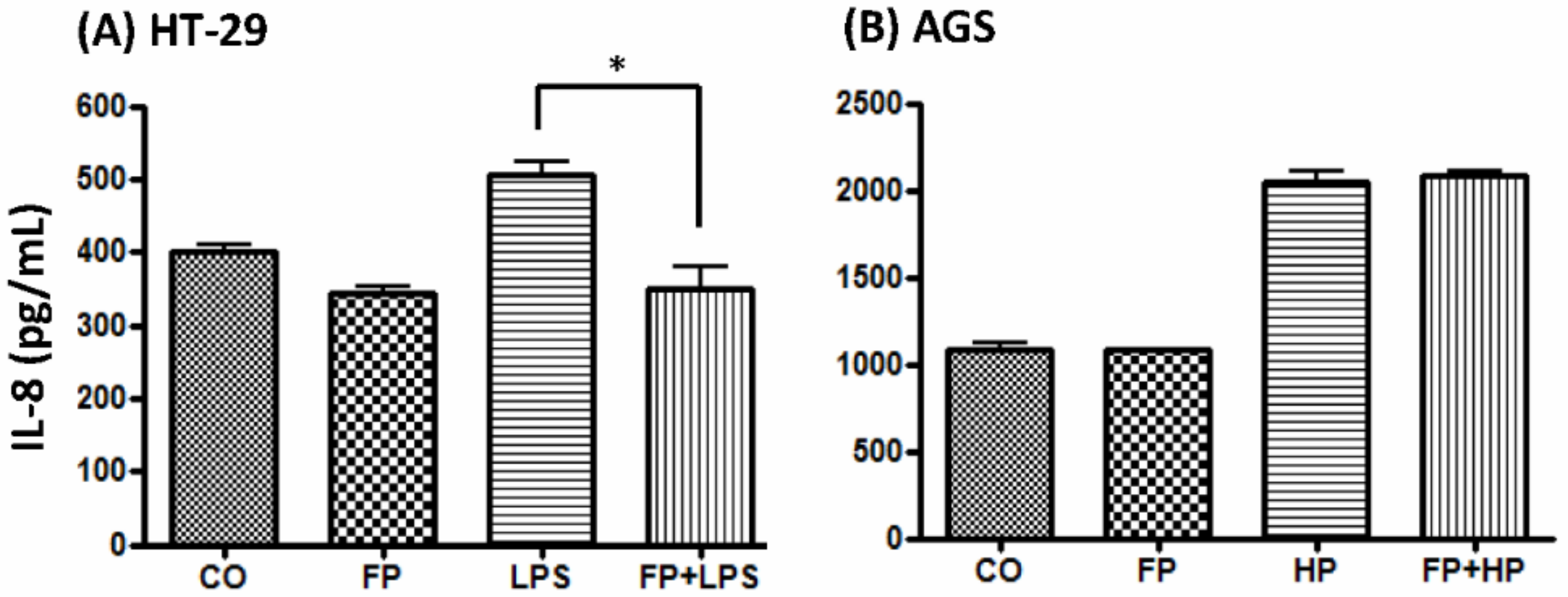
2.9. F. prausnitzii Ameliorated LPS-Induced IL-8 Expression in HT-29 Cells
3. Discussion
4. Materials and Methods
4.1. Subject Inclusion and Exclusion Criteria
4.2. Serum Collection and Diagnosis of H. pylori Infection
4.3. Probiotics-Containing Yogurt Ingestion, H. pylori Eradication and Follow-Up
4.4. Stool Collection and Preparation
4.5. Analysis of Fecal sIgA, IL-6, TGF-β, Lactoferrin, and Calprotectin
4.6. Amplicon Sequencing (16S-rRNA) for Fecal Microbiota Diversity
4.7. Probiotics Facilitate F. prausnitzii Growth In Vitro and Ex Vivo
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| TGF-β1 | Transforming growth factor-β1 |
| sIgA | Secretory immunoglobulin A |
| UBT | Urea breath test |
| PCoA | Principal coordinate analysis |
| OUT | Operational taxonomic unit. |
| ELISA | Enzyme-linked immunosorbent assay |
References
- Malaty, H.M.; Kumagai, T.; Tanaka, E.; Ota, H.; Kiyosawa, K.; Graham, D.Y.; Katsuyama, T. Evidence from a nine-year birth cohort study in Japan of transmission pathways of Helicobacter pylori infection. J. Clin. Microbiol. 2000, 38, 1971–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, B.J.; Warren, J.R. Unidentified curved bacilli in the stomach of patients with gastritis and peptic ulceration. Lancet 1984, 1, 1311–1315. [Google Scholar] [CrossRef]
- Suerbaum, S.; Michetti, P. Helicobacter pylori infection. N. Engl. J. Med. 2002, 347, 1175–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konno, M.; Muraoka, S.; Takahashi, M.; Imai, T. Iron deficiency anemia associated with Helicobacter pylori gastritis. J. Pediatr. Gastroenterol. Nutr. 2000, 31, 52–56. [Google Scholar] [CrossRef]
- Bravo, L.E.; Mera, R.; Reina, J.C.; Pradilla, A.; Alzate, A.; Fontham, E.; Correa, P. Impact of Helicobacter pylori infection on growth of children: A prospective cohort study. J. Pediatr. Gastroenterol. Nutr. 2003, 37, 614–619. [Google Scholar] [CrossRef]
- Blaser, M.J.; Chyou, P.H.; Nomura, A. Age at establishment of Helicobacter pylori infection and gastric carcinoma, gastric ulcer, and duodenal ulcer risk. Cancer Res. 1995, 55, 562–565. [Google Scholar]
- Wessler, S.; Krisch, L.M.; Elmer, D.P.; Aberger, F. From inflammation to gastric cancer—the importance of Hedgehog/GLI signaling in Helicobacter pylori-induced chronic inflammatory and neoplastic diseases. Cell Commun. Signal. 2017, 15, 15. [Google Scholar] [CrossRef] [Green Version]
- Kivrak Salim, D.; Sahin, M.; Köksoy, S.; Adanir, H.; Süleymanlar, I. Local immune response in Helicobacter pylori infection. Medicine 2016, 95, e3713. [Google Scholar] [CrossRef]
- Hestvik, E.; Tumwine, J.K.; Tylleskar, T.; Grahnquist, L.; Ndeezi, G.; Kaddu-Mulindwa, D.H.; Aksnes, L.; Olafsdottir, E. Faecal calprotectin concentrations in apparently healthy children aged 0–12 years in urban Kampala, Uganda: A community-based survey. BMC Pediatr. 2011, 11, 9. [Google Scholar] [CrossRef] [Green Version]
- Frank, D.N.; St Amand, A.L.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef] [Green Version]
- Willing, B.P.; Dicksved, J.; Halfvarson, J.; Andersson, A.F.; Lucio, M.; Zheng, Z.; Järnerot, G.; Tysk, C.; Jansson, J.K.; Engstrand, L. A pyrosequencing study in twins shows that gastrointestinal microbial profiles vary with inflammatory bowel disease phenotypes. Gastroenterology 2010, 139, 1844–1854.e1. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Shen, J.; Ran, Z.H. Association between Faecalibacterium prausnitzii reduction and inflammatory bowel disease: A meta-analysis and systematic review of the literature. Gastroenterol. Res. Pract. 2014, 2014, 872725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chassaing, B.; Gewirtz, A.T. Gut microbiota, low-grade inflammation, and metabolic syndrome. Toxicol. Pathol. 2014, 42, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Bik, E.M.; Eckburg, P.B.; Gill, S.R.; Nelson, K.E.; Purdom, E.A.; Francois, F.; Perez-Perez, G.; Blaser, M.J.; Relman, D.A. Molecular analysis of the bacterial microbiota in the human stomach. Proc. Natl. Acad. Sci. USA 2006, 103, 732–737. [Google Scholar] [CrossRef] [Green Version]
- Martin, M.E.; Bhatnagar, S.; George, M.D.; Paster, B.J.; Canfield, D.R.; Eisen, J.A.; Solnick, J.V. The impact of Helicobacter pylori infection on the gastric microbiota of the rhesus macaque. PLoS ONE 2013, 8, e76375. [Google Scholar] [CrossRef]
- Aebischer, T.; Fischer, A.; Walduck, A.; Schlötelburg, C.; Lindig, M.; Schreiber, S.; Meyer, T.F.; Bereswill, S.; Göbel, U.B. Vaccination prevents Helicobacter pylori-induced alterations of the gastric flora in mice. FEMS Immunol. Med. Microbiol. 2006, 46, 221–229. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.J.; Sheu, B.S. Probiotics-containing yogurts suppress H. pylori load, and modify immune response and intestinal microbiota in the H. pylori-infected children. Helicobacter 2012, 17, 297–304. [Google Scholar] [CrossRef]
- Kato, L.M.; Kawamoto, S.; Maruya, M.; Fagarasan, S. Gut TFH and IgA: Key players for regulation of bacterial communities and immune homeostasis. Immunol. Cell Biol. 2014, 92, 49–56. [Google Scholar] [CrossRef]
- Parvez, S.; Malik, K.A.; Ah Kang, S.; Kim, H.Y. Probiotics and their fermented food products are beneficial for health. J. Appl. Microbiol. 2006, 100, 1171–1185. [Google Scholar] [CrossRef]
- Yang, Y.J.; Chuang, C.C.; Yang, H.B.; Lu, C.C.; Sheu, B.S. Lactobacillus acidophilus ameliorates H. pylori-induced gastric inflammation by inactivating the Smad7 and NFκB pathways. BMC. Microbiol. 2012, 12, 38. [Google Scholar] [CrossRef] [Green Version]
- Gamallat, Y.; Meyiah, A.; Kuugbee, E.D.; Hago, A.M.; Chiwala, G.; Awadasseid, A.; Bamba, D.; Zhang, X.; Shang, X.; Luo, F.; et al. Lactobacillus rhamnosus induced epithelial cell apoptosis, ameliorates inflammation and prevents colon cancer development in an animal model. Biomed. Pharmacother. 2016, 83, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, Y.; Kita, M.; Kodama, T.; Sawai, N.; Kashima, K.; Imanishi, J. Induction of various cytokines and development of severe mucosal inflammation by cagA gene positive Helicobacter pylori strains. Gut 1997, 41, 442–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pabst, O. New concepts in the generation and functions of IgA. Nat. Rev. Immunol. 2012, 12, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Rol, N.; Favre, L.; Benyacoub, J.; Corthésy, B. The role of secretory immunoglobulin A in the natural sensing of commensal bacteria by mouse Peyer’s patch dendritic cells. J. Biol. Chem. 2012, 287, 40074–40082. [Google Scholar] [CrossRef] [Green Version]
- Borsutzky, S.; Cazac, B.B.; Roes, J.; Guzmán, C.A. TGF-beta receptor signaling is critical for mucosal IgA responses. J. Immunol. 2004, 173, 3305–3309. [Google Scholar] [CrossRef] [Green Version]
- Gloudemans, A.K.; Plantinga, M.; Guilliams, M.; Willart, M.A.; Ozir-Fazalalikhan, A.; van der Ham, A.; Boon, L.; Harris, N.L.; Hammad, H.; Hoogsteden, H.C.; et al. The mucosal adjuvant cholera toxin B instructs non-mucosal dendritic cells to promote IgA production via retinoic acid and TGF-β. PLoS ONE 2013, 8, e59822. [Google Scholar] [CrossRef] [Green Version]
- Montalto, M.; Gallo, A.; Ianiro, G.; Santoro, L.; D’Onofrio, F.; Ricci, R.; Cammarota, G.; Covino, M.; Vastola, M.; Gasbarrini, A.; et al. Can chronic gastritis cause an increase in fecal calprotectin concentrations? World J. Gastroenterol. 2010, 16, 3406–3410. [Google Scholar] [CrossRef]
- Eun, C.S.; Kim, B.K.; Han, D.S.; Kim, S.Y.; Kim, K.M.; Choi, B.Y.; Song, K.S.; Kim, Y.S.; Kim, J.F. Differences in gastric mucosal microbiota profiling in patients with chronic gastritis, intestinal metaplasia, and gastric cancer using pyrosequencing methods. Helicobacter 2014, 19, 407–416. [Google Scholar] [CrossRef]
- Llorca, L.; Pérez-Pérez, G.; Urruzuno, P.; Martinez, M.J.; Iizumi, T.; Gao, Z.; Sohn, J.; Chung, J.; Cox, L.; Simón-Soro, A.; et al. Characterization of the gastric microbiota in a pediatric population according to Helicobacter pylori status. Pediatr. Infect. Dis. J. 2017, 36, 173–178. [Google Scholar] [CrossRef]
- Foditsch, C.; Santos, T.M.; Teixeira, A.G.; Pereira, R.V.; Dias, J.M.; Gaeta, N.; Bicalho, R.C. Isolation and characterization of Faecalibacterium prausnitzii from calves and piglets. PLoS ONE 2014, 9, e116465. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Siles, M.; Duncan, S.H.; Garcia-Gil, L.J.; Martinez-Medina, M. Faecalibacterium prausnitzii: From microbiology to diagnostics and prognostics. ISME J. 2017, 11, 841–852. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.J.; Sheu, B.S. Metabolic interaction of Helicobacter pylori infection and gut microbiota. Microorganisms 2016, 4, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, X.; Zhang, M.; Yang, X.; Hong, N.; Yu, C. Faecalibacterium prausnitzii upregulates regulatory T cells and anti-inflammatory cytokines in treating TNBS-induced colitis. J. Crohns Colitis. 2013, 7, e558–e568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breyner, N.M.; Michon, C.; de Sousa, C.S.; Vilas Boas, P.B.; Chain, F.; Azevedo, V.A.; Langella, P.; Chatel, J.M. Microbial anti-inflammatory molecule (MAM) from Faecalibacterium prausnitzii shows a protective effect on DNBS and DSS-Induced colitis model in mice through inhibition of NF-κB pathway. Front. Microbiol. 2017, 8, 114. [Google Scholar] [CrossRef] [Green Version]
- Miquel, S.; Martín, R.; Rossi, O.; Bermúdez-Humarán, L.G.; Chatel, J.M.; Sokol, H.; Thomas, M.; Wells, J.M.; Langella, P. Faecalibacterium prausnitzii and human intestinal health. Curr. Opin. Microbiol. 2013, 16, 255–261. [Google Scholar] [CrossRef]
- Yang, Y.J.; Sheu, B.S.; Lee, S.C.; Wu, J.J. Short-term recurrent abdominal pain related to Helicobacter pylori infection in children. J. Gastroenterol. Hepatol. 2005, 20, 395–400. [Google Scholar] [CrossRef]
- Yang, Y.J.; Sheu, B.S.; Lee, S.C.; Yang, H.B.; Wu, J.J. Children of H. pylori-infected dyspeptic mothers are predisposed to H. pylori acquisition with subsequent iron deficiency and growth retardation. Helicobacter 2005, 10, 249–255. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Siles, M.; Khan, T.M.; Duncan, S.H.; Harmsen, H.J.; Garcia-Gil, L.J.; Flint, H.J. Cultured representatives of two major phylogroups of human colonic Faecalibacterium prausnitzii can utilize pectin, uronic acids, and host-derived substrates for growth. Appl. Environ. Microbiol. 2012, 78, 420–428. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Siles, M.; Martinez-Medina, M.; Surís-Valls, R.; Aldeguer, X.; Sabat-Mir, M.; Duncan, S.H.; Flint, H.J.; Garcia-Gil, L.J. Changes in the abundance of Faecalibacterium prausnitzii phylogroups I and II in the intestinal mucosa of inflammatory bowel disease and patients with colorectal cancer. Inflamm. Bowel Dis. 2016, 22, 28–41. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, R.M.; Pereira-Marques, J.; Pinto-Ribeiro, I.; Costa, J.L.; Carneiro, F.; Machado, J.C.; Figueiredo, C. Gastric microbial community profiling reveals a dysbiotic cancer-associated microbiota. Gut 2018, 67, 226–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OTUs; Taxonomy | Mean Proportional Abundance (%) | Percent Change | p Value | |
|---|---|---|---|---|
| H. pylori-Infected (HPS1, n = 10) | Controls (CS1, n = 9) | |||
| 004; Firmicutes; Clostridia; Clostridiales; Ruminococcaceae; Faecalibacterium | 2.3347 | 5.0 | −2.14 | 0.005994 |
| 065; Bacteroidetes; Bacteroidia; Bacteroidales; Porphyromonadaceae | 0.0004 | 0.1859 | −464.75 | 0.008991 |
| 051; Proteobacteria; Deltaproteobacteria; Desulfovibrionales; Desulfovibrionaceae; Desulfovibrio | 0.002 | 0.2433 | −121.65 | 0.015984 |
| 122; Firmicutes; Clostridia; Clostridiales; Eubacteriaceae; Eubacterium | 0 | 0.0031 | − | 0.005351 |
| 082; Proteobacteria; Alphaproteobacteria | 0.1157 | 0 | − | 0.000999 |
| 070; Actinobacteria; Actinobacteria; Coriobacteriales; Coriobacteriaceae | 0.0101 | 0 | − | 0.003996 |
| 136; Firmicutes; Clostridia; Clostridiales; Lachnospiraceae; Howardella | 0.0036 | 0 | − | 0.004299 |
| 063; Firmicutes; Clostridia; Clostridiales; Ruminococcaceae; Pseudoflavonifractor | 0.0653 | 0.0013 | 0.02 | 0.007992 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.-J.; Chen, P.-C.; Lai, F.-P.; Tsai, P.-J.; Sheu, B.-S. Probiotics-Containing Yogurt Ingestion and H. pylori Eradication Can Restore Fecal Faecalibacterium prausnitzii Dysbiosis in H. pylori-Infected Children. Biomedicines 2020, 8, 146. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8060146
Yang Y-J, Chen P-C, Lai F-P, Tsai P-J, Sheu B-S. Probiotics-Containing Yogurt Ingestion and H. pylori Eradication Can Restore Fecal Faecalibacterium prausnitzii Dysbiosis in H. pylori-Infected Children. Biomedicines. 2020; 8(6):146. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8060146
Chicago/Turabian StyleYang, Yao-Jong, Peng-Chieh Chen, Fu-Ping Lai, Pei-Jane Tsai, and Bor-Shyang Sheu. 2020. "Probiotics-Containing Yogurt Ingestion and H. pylori Eradication Can Restore Fecal Faecalibacterium prausnitzii Dysbiosis in H. pylori-Infected Children" Biomedicines 8, no. 6: 146. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8060146