
Characterization of Differentially Expressed Circulating miRNAs in Metabolically Healthy versus Unhealthy Obesity

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Blood Sampling
2.3. miRNA Isolation and Microarray Analysis
2.4. miRNA Target Prediction and Encyclopaedia of Genes and Genomes (KEGG) Pathway Analyses
2.5. miRNAs qPCR Validation
2.6. Data Analysis
3. Results
3.1. Characteristics of the Study Cohort
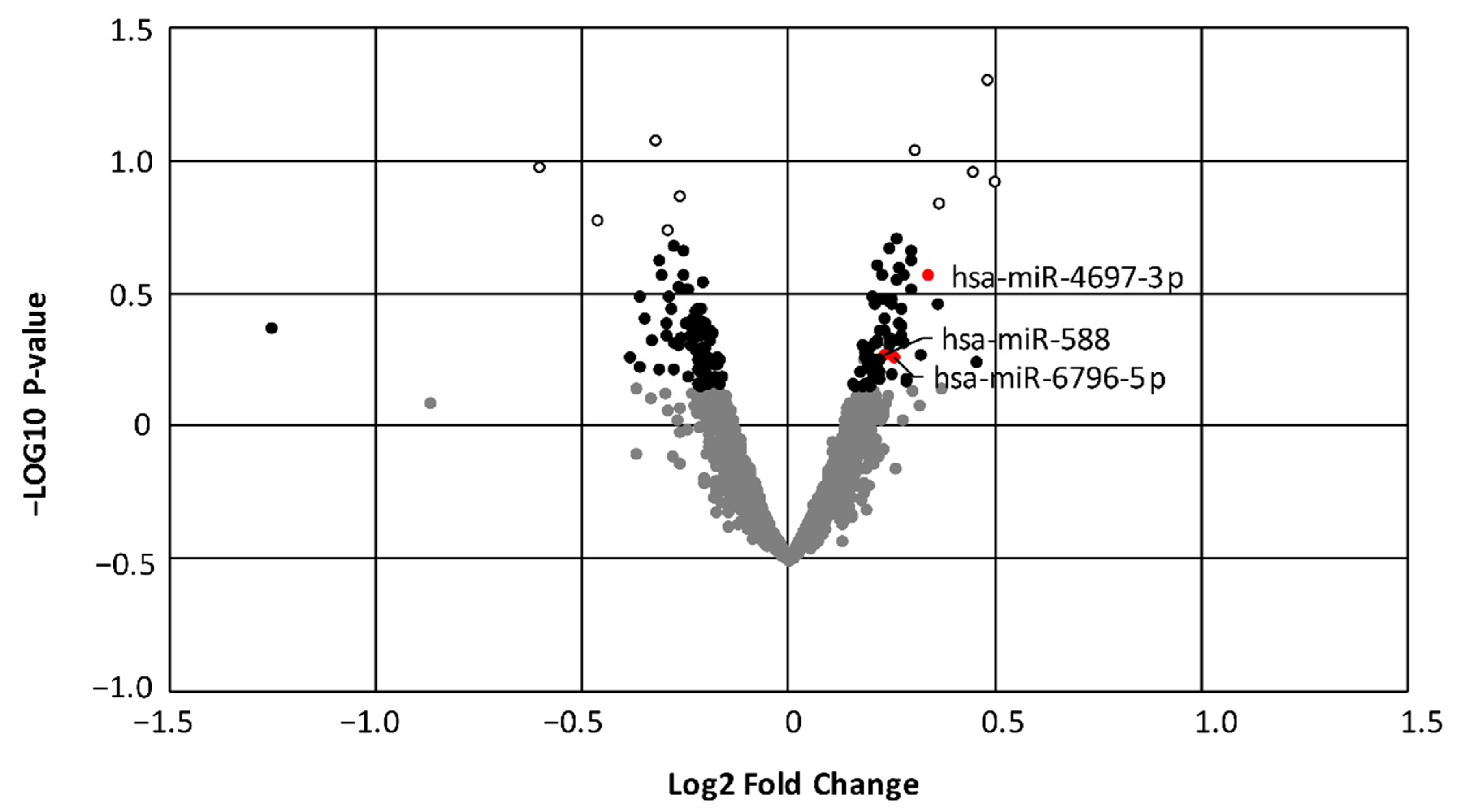
3.2. Evaluation of Differentially Expressed miRNA
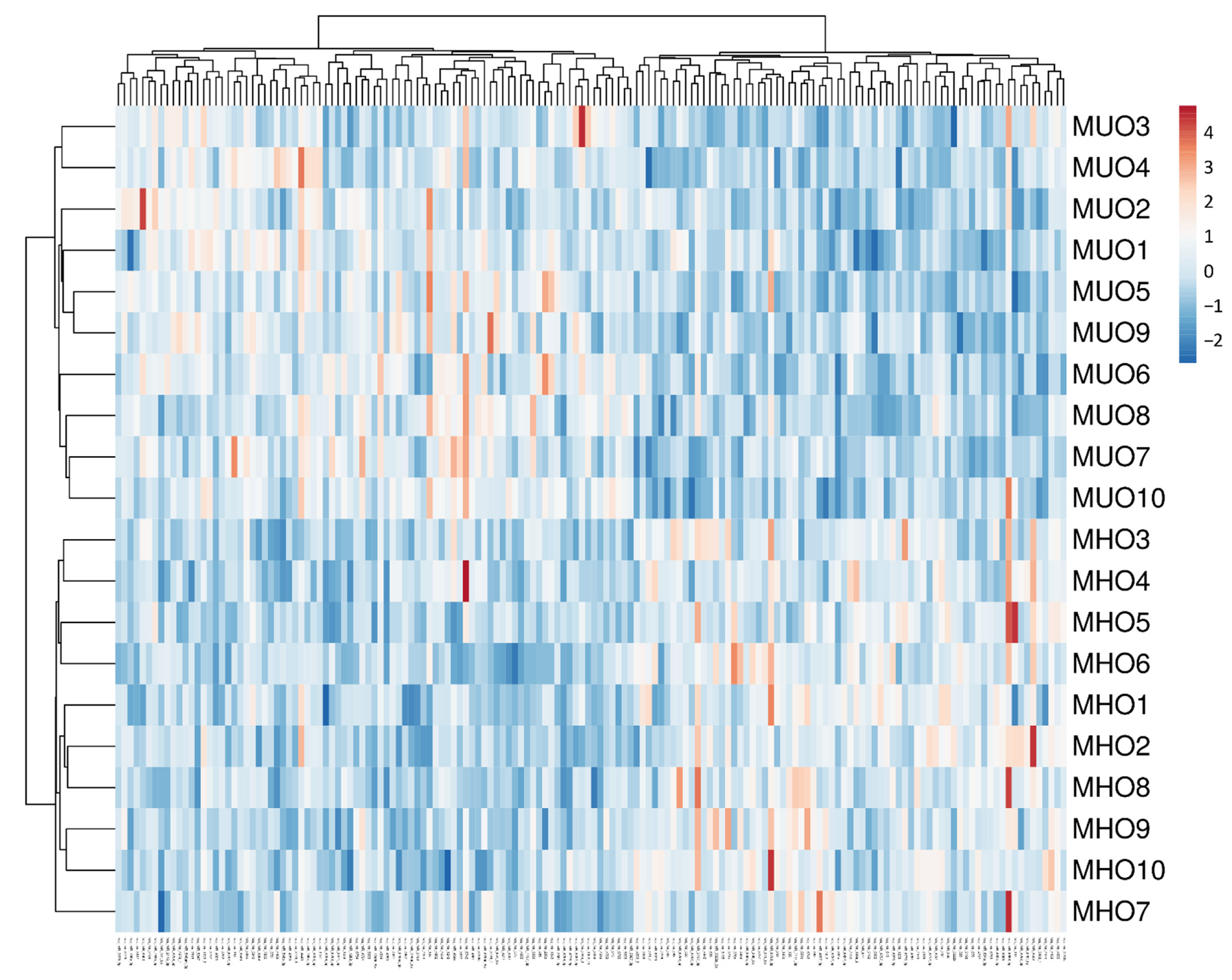
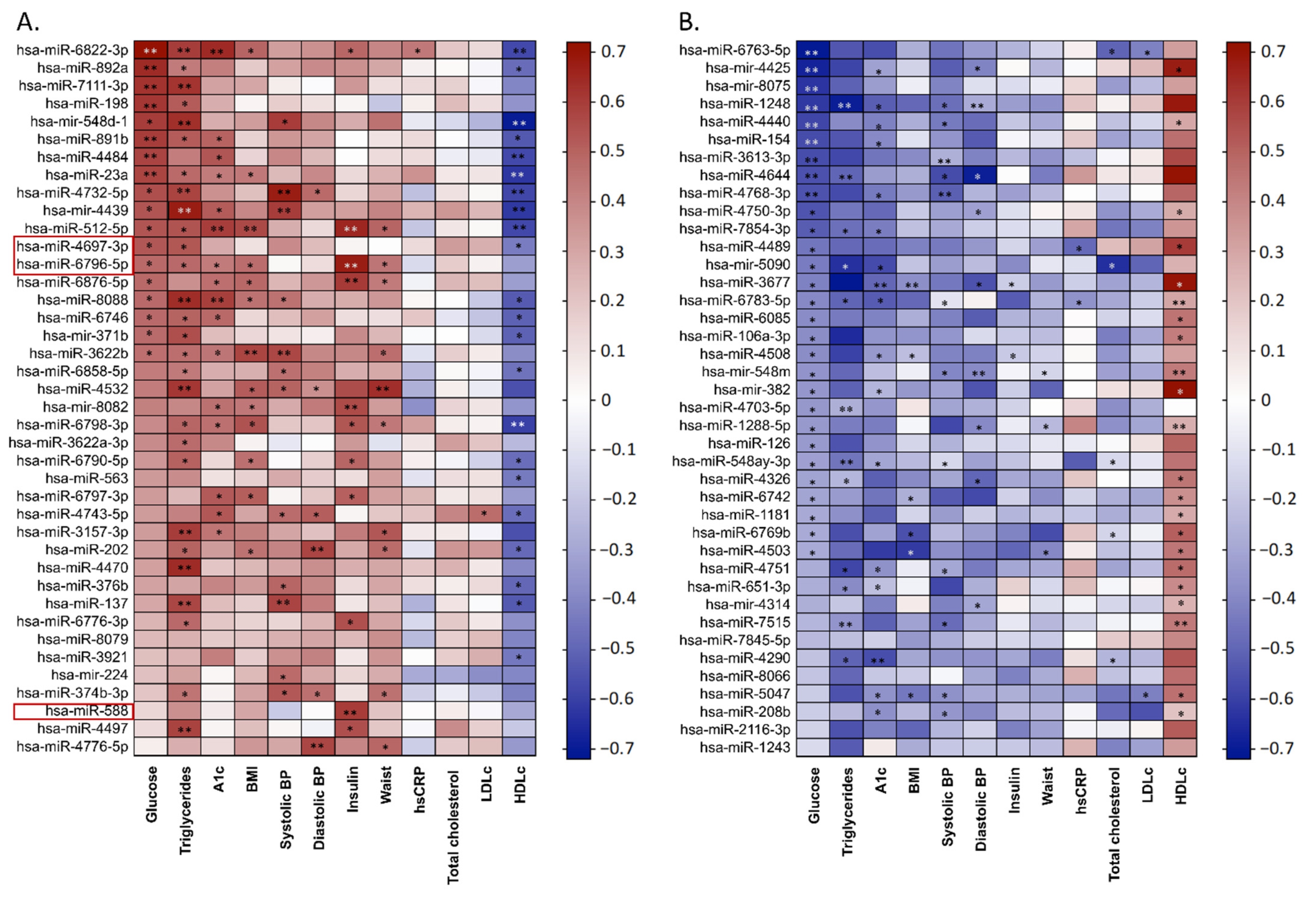
3.3. Heatmap
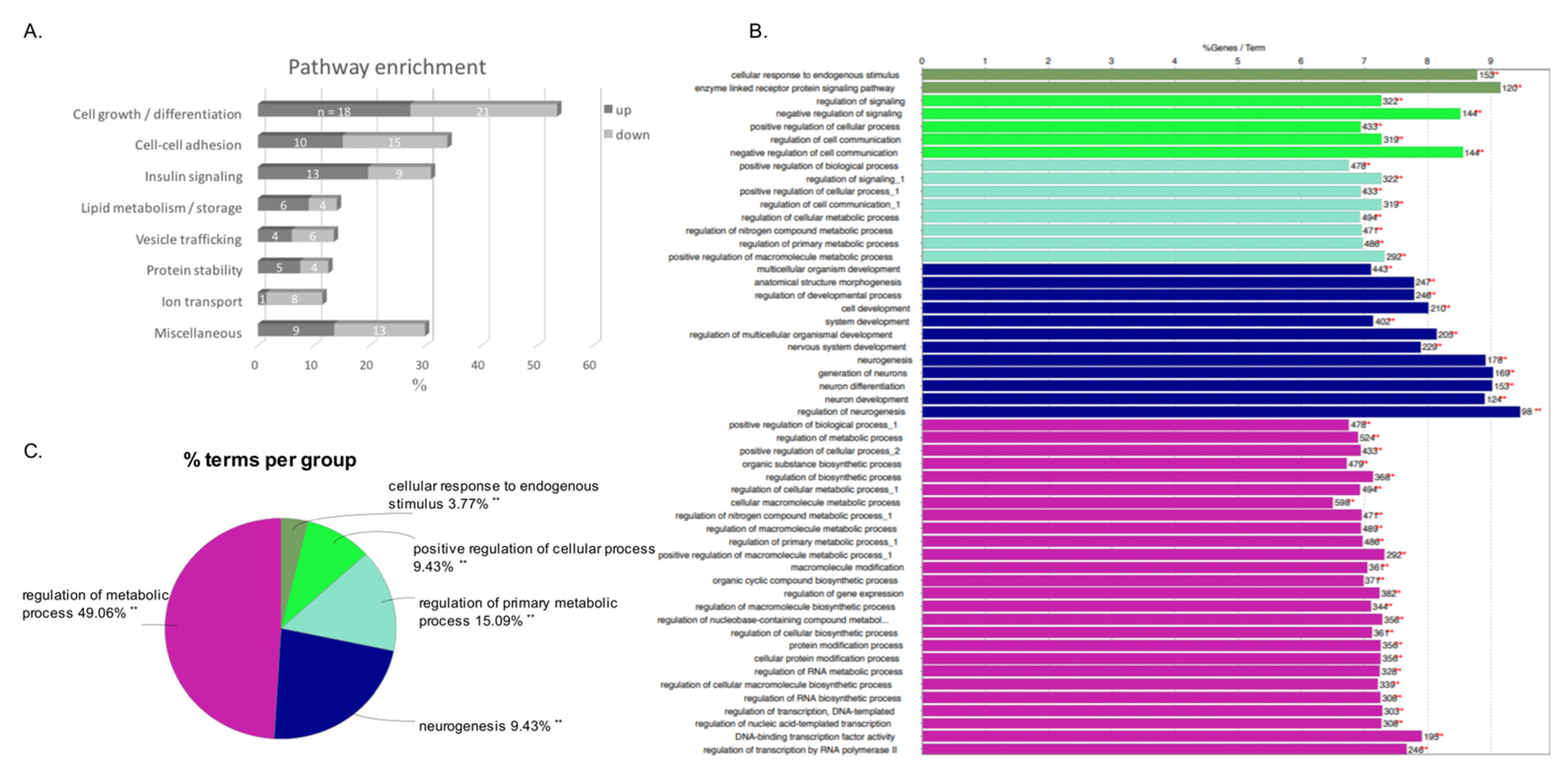
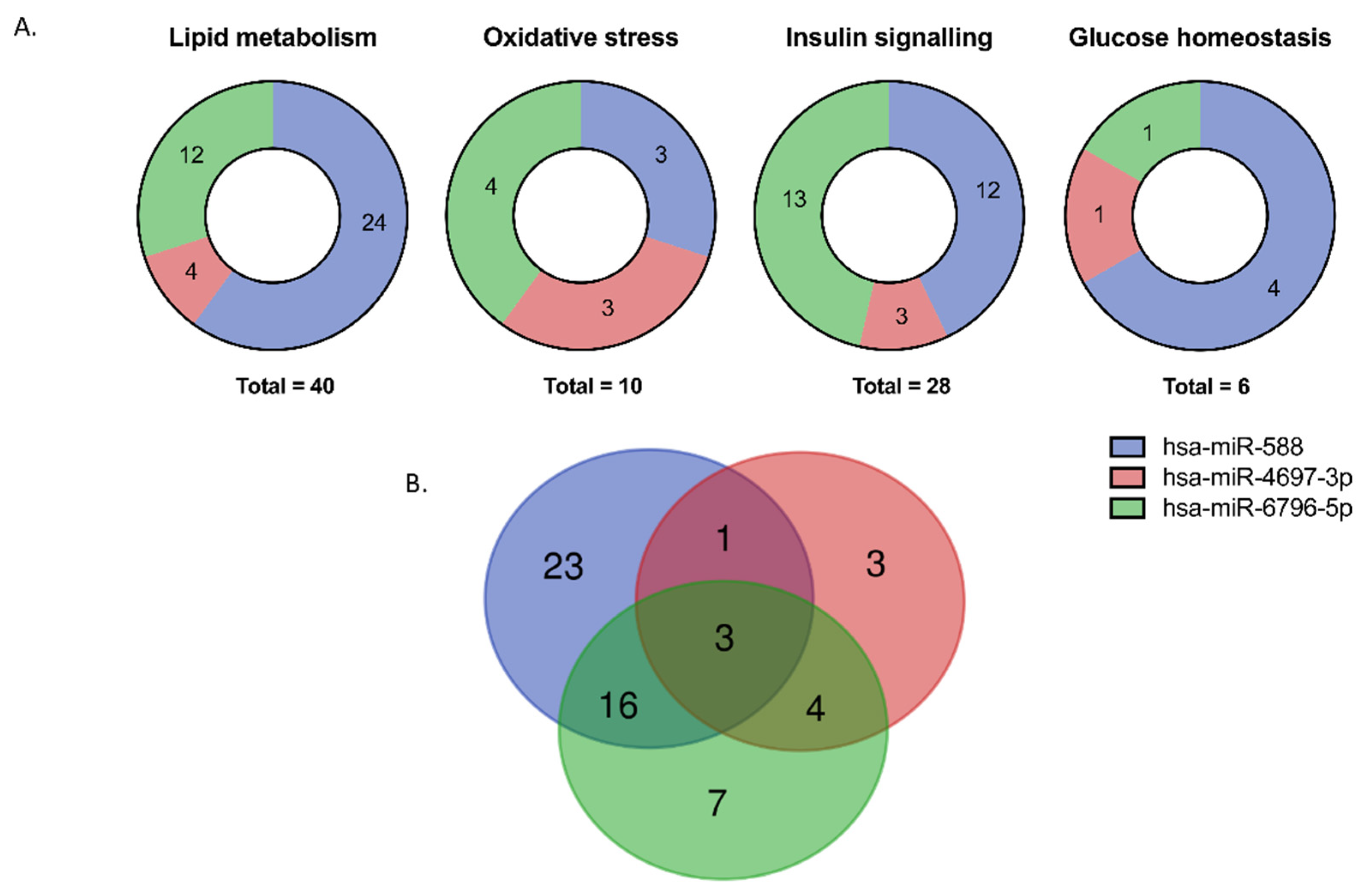
3.4. Pathway Enrichment
3.5. In Silico Analysis of Potential miRNA Target Genes
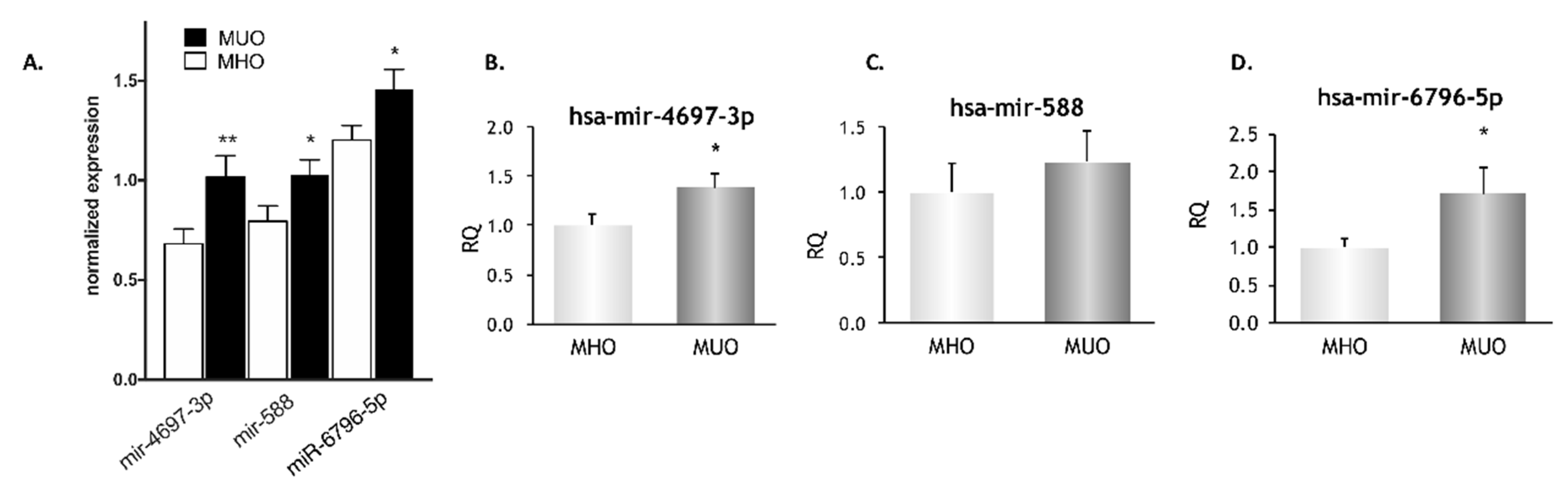
3.6. Validation of a Subgroup of miRNAs
3.7. Correlation Studies
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stefan, N.; Häring, H.U.; Hu, F.B.; Schulze, M.B. Metabolically healthy obesity: Epidemiology, mechanisms, and clinical implications. Lancet Diabetes Endocrinol. 2013, 1, 152–162. [Google Scholar] [CrossRef]
- Magkos, F. Metabolically healthy obesity: What’s in a name? Am. J. Clin. Nutr. 2019, 110, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Herrera, B.M.; Lindgren, C.M. The genetics of obesity. Curr. Diab. Rep. 2010, 10, 498–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albuquerque, D.; Stice, E.; Rodríguez-López, R.; Manco, L.; Nóbrega, C. Current review of genetics of human obesity: From molecular mechanisms to an evolutionary perspective. Mol. Genet. Genom. 2015, 290, 1191–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pigeyre, M.; Yazdi, F.T.; Kaur, Y.; Meyre, D. Recent progress in genetics, epigenetics and metagenomics unveils the pathophysiology of human obesity. Clin. Sci. 2016, 130, 943–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landrier, J.F.; Derghal, A.; Mounien, L. MicroRNAs in Obesity and Related Metabolic Disorders. Cells 2019, 8, 859. [Google Scholar] [CrossRef] [Green Version]
- Frost, R.J.; Olson, E.N. Control of glucose homeostasis and insulin sensitivity by the Let-7 family of microRNAs. Proc. Natl. Acad. Sci. USA 2011, 108, 21075–21080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Włodarski, A.; Strycharz, J.; Wróblewski, A.; Kasznicki, J.; Drzewoski, J.; Śliwińska, A. The Role of microRNAs in Metabolic Syndrome-Related Oxidative Stress. Int. J. Mol. Sci. 2020, 21, 6902. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liang, H.; Zhang, J.; Zen, K.; Zhang, C.Y. Secreted microRNAs: A new form of intercellular communication. Trends Cell Biol. 2012, 22, 125–132. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Clark, A.G. Impact of microRNA regulation on variation in human gene expression. Genome Res. 2012, 22, 1243–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arner, P.; Kulyté, A. MicroRNA regulatory networks in human adipose tissue and obesity. Nat. Rev. Endocrinol. 2015, 11, 276–288. [Google Scholar] [CrossRef]
- Huang, Y.; Yan, Y.; Xv, W.; Qian, G.; Li, C.; Zou, H.; Li, Y. A New Insight into the Roles of MiRNAs in Metabolic Syndrome. Biomed Res. Int. 2018, 2018, 7372636. [Google Scholar] [CrossRef] [Green Version]
- Ji, C.; Guo, X. The clinical potential of circulating microRNAs in obesity. Nat. Rev. Endocrinol. 2019, 15, 731–743. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Lin, Y.C.D.; Li, J.; Huang, K.Y.; Shrestha, S.; Hong, H.C.; Tang, Y.; Chen, Y.G.; Jin, C.N.; Yu, Y.; et al. miRTarBase 2020: Updates to the experimentally validated microRNA-target interaction database. Nucleic Acids Res. 2020, 48, D148–D154. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Wong, N.; Wang, X. miRDB: An online resource for microRNA target prediction and functional annotations. Nucleic Acids Res. 2015, 43, D146–D152. [Google Scholar] [CrossRef]
- Bindea, G.; Galon, J.; Mlecnik, B. CluePediaCytoscape plugin: Pathway insights using integrated experimental and in silico data. Bioinformatics 2013, 29, 661–663. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: ACytoscape plugin to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]
- Bañuls, C.; Rovira-Llopis, S.; Lopez-Domenech, S.; Diaz-Morales, N.; Blas-Garcia, A.; Veses, S.; Morillas, C.; Victor, V.M.; Rocha, M.; Hernandez-Mijares, A. Oxidative and endoplasmic reticulum stress is impaired in leukocytes from metabolically unhealthy vs. healthy obese individuals. Int. J. Obes. 2017, 41, 1556–1563. [Google Scholar] [CrossRef]
- Choi, H.; Koh, H.W.; Zhou, L.; Cheng, H.; Loh, T.P.; ParvareshRizi, E.; Toh, S.A.; Ronnett, G.V.; Huang, B.E.; Khoo, C.M. Plasma Protein and MicroRNA Biomarkers of Insulin Resistance: A Network-Based Integrative-Omics Analysis. Front. Physiol. 2019, 10, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Yu, S.; Li, H.; Xiang, H.; Peng, J.; Jiang, S. MicroRNAs: Emerging roles in adipogenesis and obesity. Cell Signal. 2014, 26, 1888–1896. [Google Scholar] [CrossRef]
- Jones, A.; Danielson, K.M.; Benton, M.C.; Ziegler, O.; Shah, R.; Stubbs, R.S.; Das, S.; Macartney-Coxson, D. miRNA Signatures of Insulin Resistance in Obesity. Obesity 2017, 25, 1734–1744. [Google Scholar] [CrossRef] [Green Version]
- Doumatey, A.P.; He, W.J.; Gaye, A.; Lei, L.; Zhou, J.; Gibbons, G.H.; Adeyemo, A.; Rotimi, C.N. Circulating MiR-374a-5p is a potential modulator of the inflammatory process in obesity. Sci. Rep. 2018, 8, 7680. [Google Scholar] [CrossRef] [PubMed]
- Arner, E.; Mejhert, N.; Kulyté, A.; Balwierz, P.J.; Pachkov, M.; Cormont, M.; Lorente-Cebrián, S.; Ehrlund, A.; Laurencikiene, J.; Hedén, P.; et al. Adipose tissue microRNAs as regulators of CCL2 production in human obesity. Diabetes 2012, 61, 1986–1993. [Google Scholar] [CrossRef] [Green Version]
- Ma, E.; Fu, Y.; Garvey, W.T. Relationship of Circulating miRNAs with Insulin Sensitivity and Associated Metabolic Risk Factors in Humans. Metab. Syndr. Relat. Disord. 2018, 16, 82–89. [Google Scholar] [CrossRef]
- Krist, B.; Florczyk, U.; Pietraszek-Gremplewicz, K.; Józkowicz, A.; Dulak, J. The Role of miR-378a in Metabolism, Angiogenesis, and Muscle Biology. Int. J. Endocrinol. 2015, 2015, 281756. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Ding, Y.; Liang, B.; Lin, J.; Kim, T.K.; Yu, H.; Hang, H.; Wang, K. A Systematic Study of Dysregulated MicroRNA in Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2017, 18, 456. [Google Scholar] [CrossRef]
- Cӑtoi, A.F.; Pârvu, A.E.; Andreicuț, A.D.; Mironiuc, A.; Crӑciun, A.; Cӑtoi, C.; Pop, I.D. Metabolically Healthy versus Unhealthy Morbidly Obese: Chronic Inflammation, Nitro-Oxidative Stress, and Insulin Resistance. Nutrients 2018, 10, 1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Paik, J.K.; Kang, R.; Kim, S.Y.; Lee, S.H.; Lee, J.H. Increased oxidative stress in normal-weight postmenopausal women with metabolic syndrome compared with metabolically healthy overweight/obese individuals. Metabolism 2013, 62, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Yoo, H.J.; Ko, J.; Lee, J.H. Metabolically unhealthy overweight individuals have high lysophosphatide levels, phospholipase activity, and oxidative stress. Clin. Nutr. 2020, 39, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Marques-Rocha, J.L.; Samblas, M.; Milagro, F.I.; Bressan, J.; Martínez, J.A.; Marti, A. Noncoding RNAs, cytokines, and inflammation-related diseases. FASEB J. 2015, 29, 3595–3611. [Google Scholar] [CrossRef] [Green Version]
- Gaia‑Oltean, A.I.; Braicu, C.; Gulei, D.; Ciortea, R.; Mihu, D.; Roman, H.; Irimie, A.; Berindan‑Neagoe, I. Ovarian endometriosis, a precursor of ovarian cancer: Histological aspects, gene expression and microRNA alterations. Exp. Ther. Med. 2021, 21, 243. [Google Scholar] [CrossRef]
- Igelmann, S.; Neubauer, H.A.; Ferbeyre, G. STAT3 and STAT5 Activation in Solid Cancers. Cancers 2019, 11, 1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghafouri-Fard, S.; Abak, A.; TondroAnamag, F.; Shoorei, H.; Majidpoor, J.; Taheri, M. The emerging role of non-coding RNAs in the regulation of PI3K/AKT pathway in the carcinogenesis process. Biomed Pharmacother. 2021, 137, 111279. [Google Scholar] [CrossRef]
- Chen, Q.; Chen, S.; Zhao, J.; Zhou, Y.; Xu, L. MicroRNA-126: A new and promising player in lung cancer. Oncol. Lett. 2021, 21, 35. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, R.; Zhou, X.; Guo, R.; Yin, J.; Li, Y.; Ma, G. miR-137: A Novel Therapeutic Target for Human Glioma. Mol. Ther. Nucleic Acids 2020, 21, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, J.; Wei, T.; Li, J. Down-Regulation of MicroRNA-137 Improves High Glucose-Induced Oxidative Stress Injury in Human Umbilical Vein Endothelial Cells by Up-Regulation of AMPKα1. Cell. Physiol. Biochem. 2016, 39, 847–859. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MHO | MUO | p-Value | |
|---|---|---|---|
| n (% men) | 10 (20) | 10 (30) | ns |
| Age (years) | 44.2 ± 10.5 | 45.8 ± 6.5 | ns |
| BMI (kg/m2) | 38.1 ± 2.86 | 42.9 ± 4.37 | <0.05 |
| Waist (cm) | 115.3 ± 11.6 | 129.4 ± 11.8 | <0.05 |
| Systolic BP (mm Hg) | 122 ± 10 | 142 ± 14 | <0.01 |
| Diastolic BP (mm Hg) | 76 ± 6 | 89 ± 11 | <0.05 |
| Total cholesterol (mg/dL) | 178 ± 29 | 194 ± 25 | ns |
| LDLc (mg/dL) | 108 ± 23 | 115 ± 21 | ns |
| HDLc (mg/dL) | 55 ± 7.7 | 35 ± 3.3 | <0.001 |
| Triglycerides (mg/dL) | 70.5 (47.0–102.0) | 225 (186–244) | <0.001 |
| Apo AI (mg/dL) | 164 ± 18 | 134 ± 15 | <0.01 |
| Apo B (mg/dL) | 89 ± 20 | 114 ± 19 | 0.05 |
| Glucose (mg/dL) | 87.1 ± 8.4 | 107.2 ± 8.1 | <0.001 |
| Insulin (μU/mL) | 13.5 ± 6.9 | 27.3 ± 16.4 | <0.05 |
| HOMA-IR | 2.85 ± 1.29 | 7.16 ± 4.12 | <0.01 |
| HbA1c (%) | 5.22 ± 0.37 | 5.91 ± 0.40 | <0.01 |
| hsCRP (mg/L) | 4.57 (2.83–11.17) | 5.33 (2.27–11.78) | ns |
| Direction | Name | Target Gene | Fold Change | p-Value |
|---|---|---|---|---|
| Up | hsa-miR-4532 | POU3F1 | 1.39 | 0.0002 |
| hsa-miR-548d-1 | PPARA * | 1.23 | 0.0008 | |
| hsa-miR-3157-3p | OR11A1 | 1.36 | 0.0012 | |
| hsa-miR-137 | CDK6 * | 1.41 | 0.0014 | |
| hsa-miR-6798-3p | PEBP1 | 1.29 | 0.0020 | |
| Down | hsa-miR-5047 | TFRC | 0.79 | 0.0007 |
| hsa-miR-4508 | BOK | 0.66 | 0.0011 | |
| hsa-miR-1181 | STAT3 * | 0.83 | 0.0018 | |
| hsa-miR-126 | CRK * | 0.72 | 0.0027 | |
| hsa-miR-5090 | WNK2 | 0.81 | 0.0033 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rovira-Llopis, S.; Díaz-Rúa, R.; Grau-del Valle, C.; Iannantuoni, F.; Abad-Jimenez, Z.; Bosch-Sierra, N.; Panadero-Romero, J.; Victor, V.M.; Rocha, M.; Morillas, C.; et al. Characterization of Differentially Expressed Circulating miRNAs in Metabolically Healthy versus Unhealthy Obesity. Biomedicines 2021, 9, 321. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9030321
Rovira-Llopis S, Díaz-Rúa R, Grau-del Valle C, Iannantuoni F, Abad-Jimenez Z, Bosch-Sierra N, Panadero-Romero J, Victor VM, Rocha M, Morillas C, et al. Characterization of Differentially Expressed Circulating miRNAs in Metabolically Healthy versus Unhealthy Obesity. Biomedicines. 2021; 9(3):321. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9030321
Chicago/Turabian StyleRovira-Llopis, Susana, Rubén Díaz-Rúa, Carmen Grau-del Valle, Francesca Iannantuoni, Zaida Abad-Jimenez, Neus Bosch-Sierra, Joaquín Panadero-Romero, Víctor M. Victor, Milagros Rocha, Carlos Morillas, and et al. 2021. "Characterization of Differentially Expressed Circulating miRNAs in Metabolically Healthy versus Unhealthy Obesity" Biomedicines 9, no. 3: 321. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9030321