
Probiotic Bifidobacterium bifidum G9-1 Has a Preventive Effect on the Acceleration of Colonic Permeability and M1 Macrophage Population in Maternally Separated Rats

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Maternal Separation and Experimental Design
2.2. Real-Time RT-PCR
2.3. Tissue Specimen and Immunohistochemistry
2.4. Ussing Chamber Assay
2.5. Measurement of Transepithelial Electrical Resistance
2.6. Statistical Analysis
3. Results
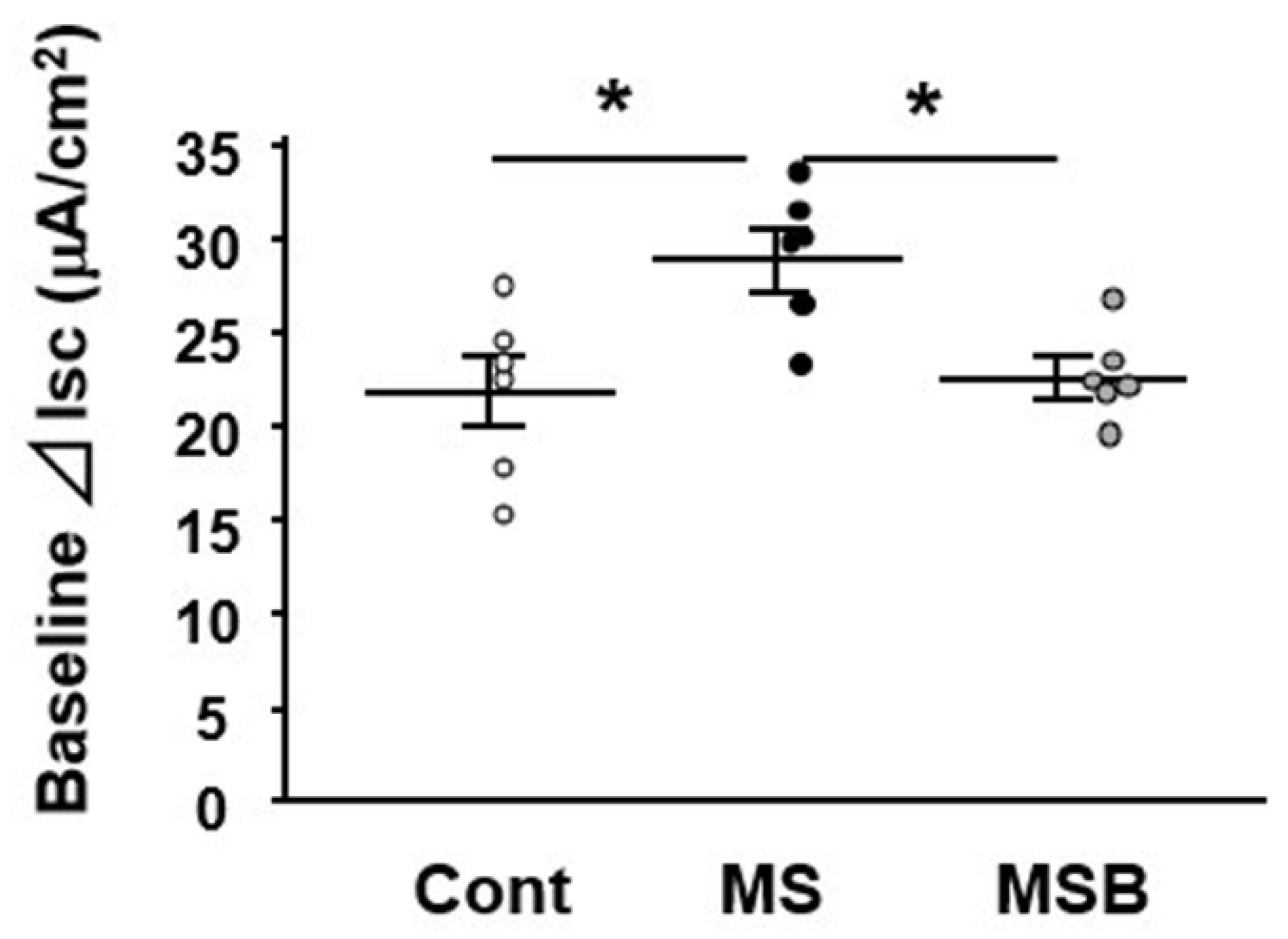
3.1. Effect of BBG9-1 on Intestinal Permeability in MS Rats
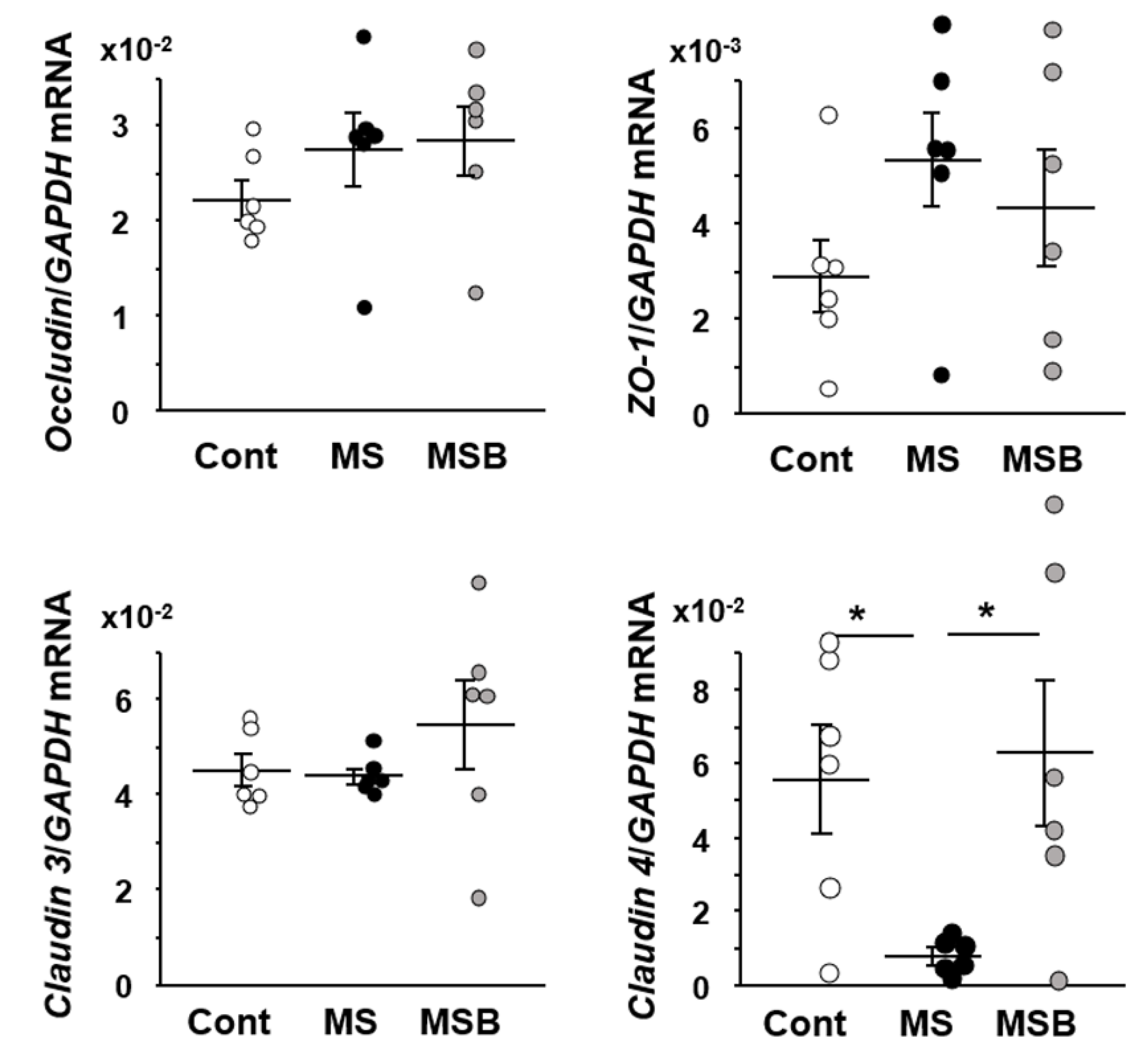
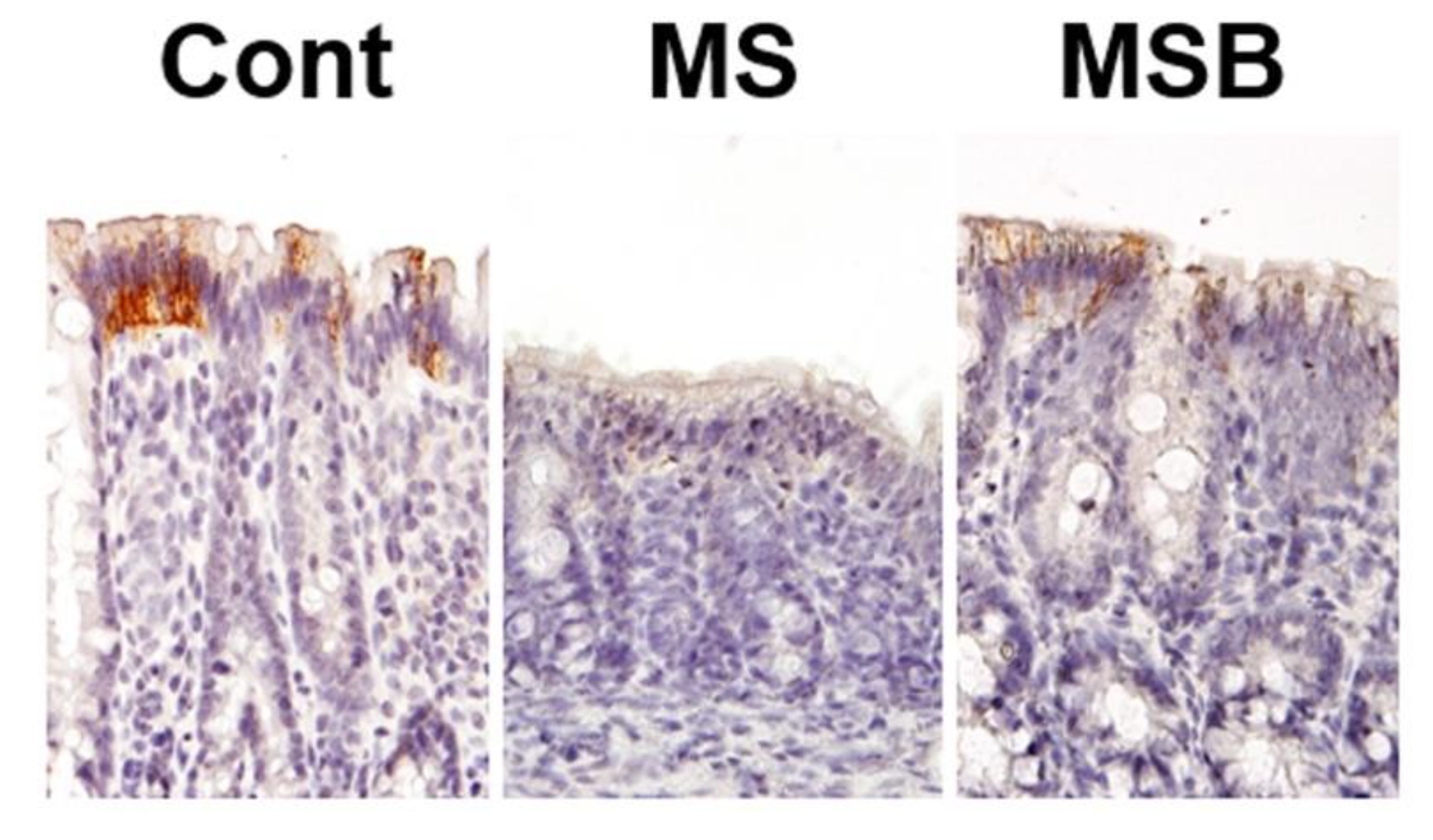
3.2. Effect of BBG9-1 on the Expression of Tight Junction Proteins in the Intestine of MS Rats
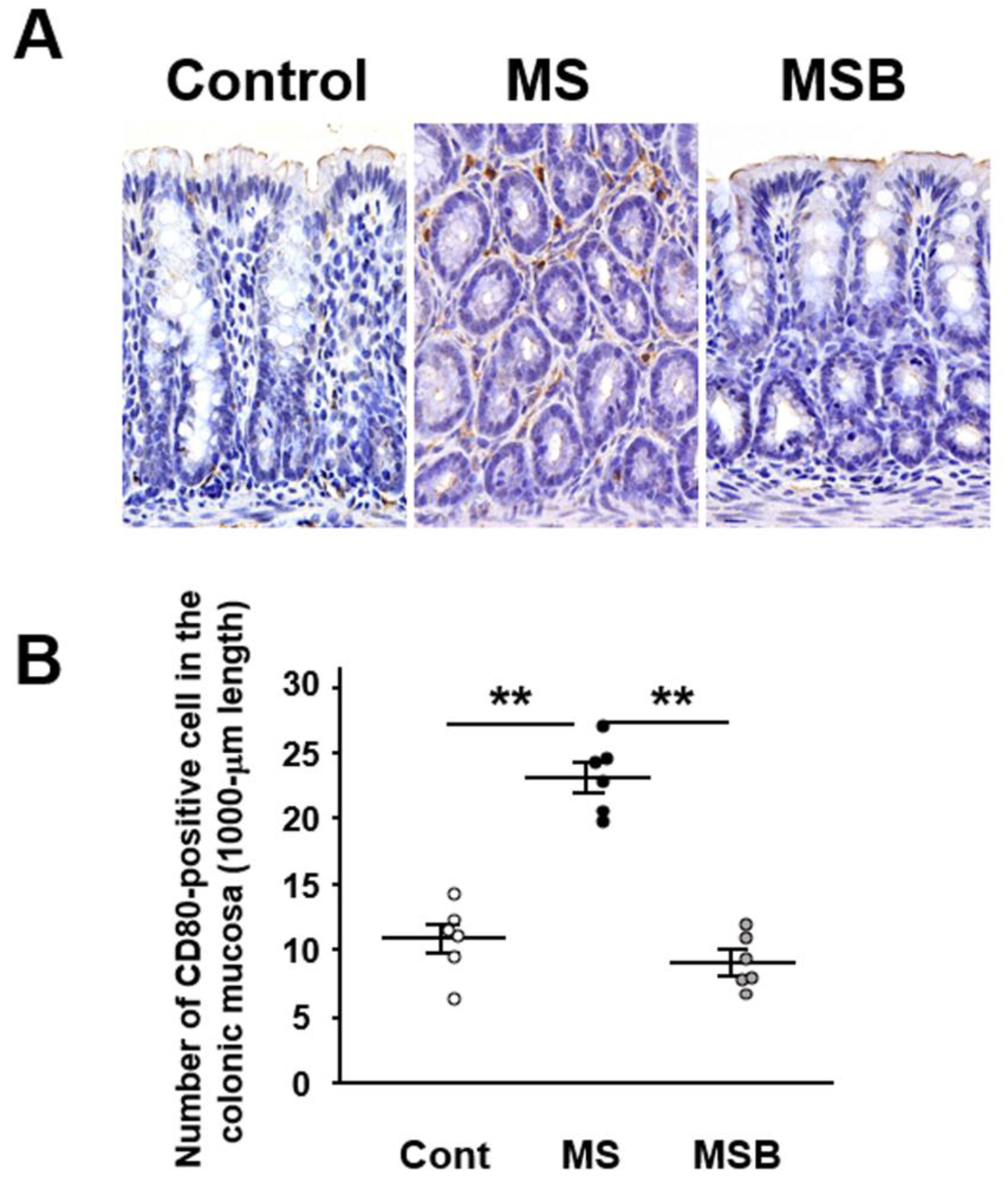
3.3. Effect of BBG9-1 on the Population of CD80-Positive Cells in the Intestinal Mucosa of MS Rats
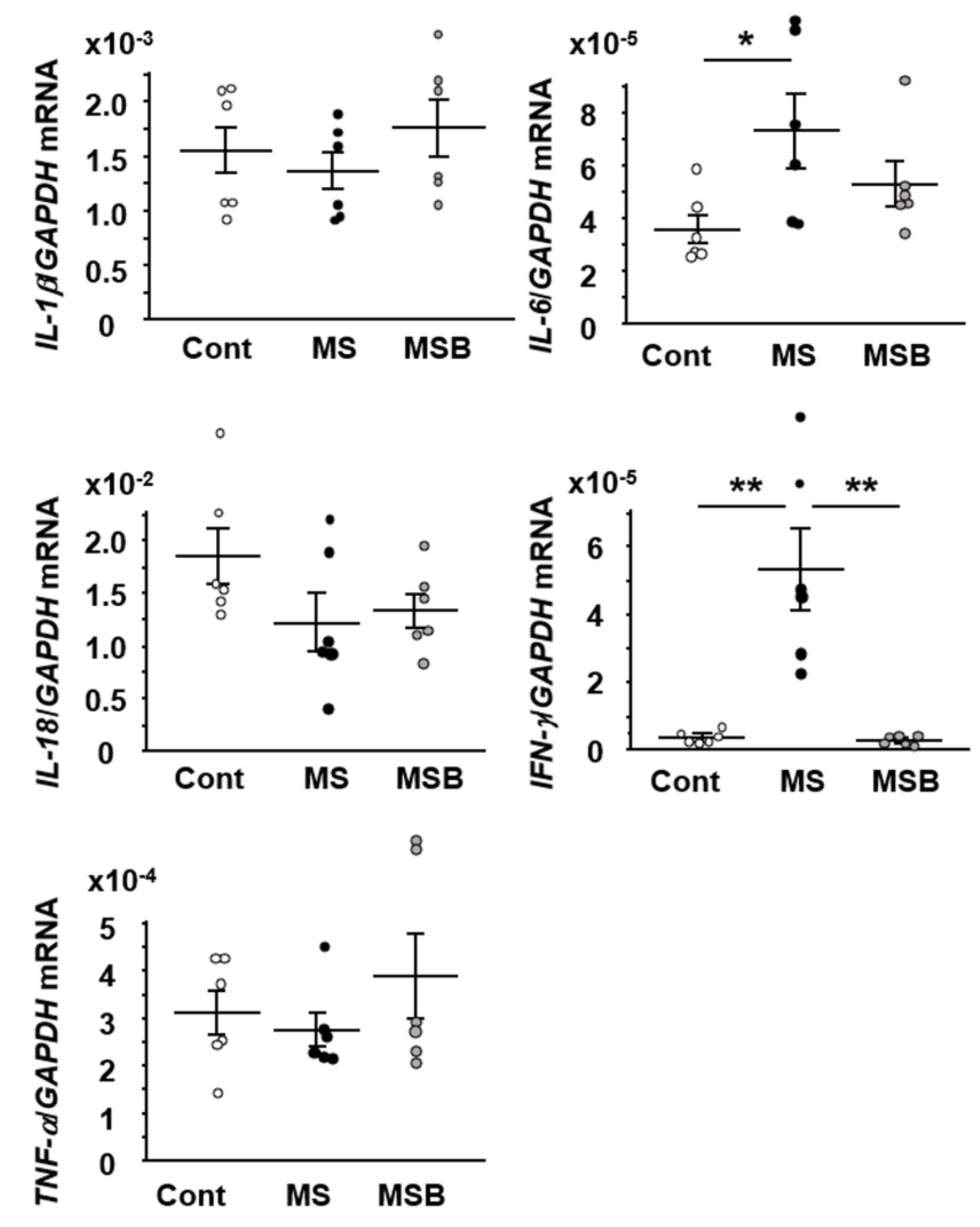
3.4. Effect of BBG9-1 on the Expression of Cytokines in the Intestine of MS Rats
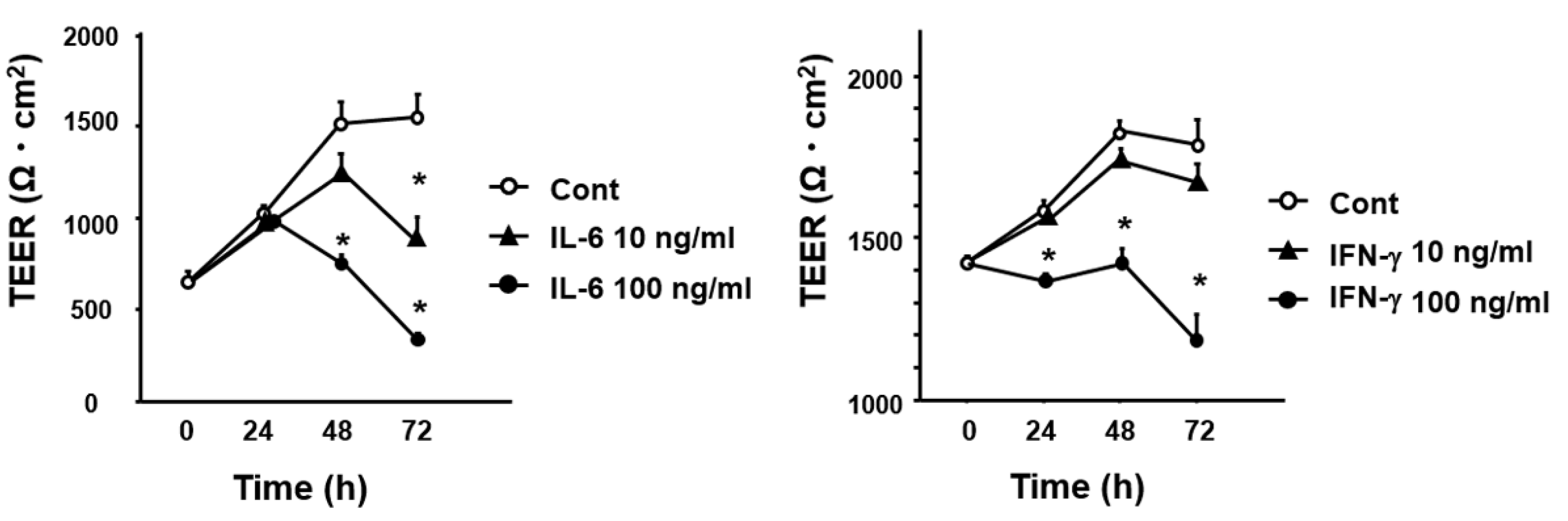
3.5. Effect of Cytokines IL-6 and IFN-γ on Intestinal Permeability In Vitro
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Longstreth, G.F.; Thompson, W.G.; Chey, W.D.; Houghton, L.A.; Mearin, F.; Spiller, R.C. Functional bowel disorders. Gastroenterology 2006, 130, 1480–1491. [Google Scholar] [CrossRef] [PubMed]
- Chey, W.D.; Kurlander, J.; Eswaran, S. Irritable bowel syndrome: A clinical review. JAMA 2015, 313, 949–958. [Google Scholar] [CrossRef] [PubMed]
- Gunnarsson, J.; Simren, M. Peripheral factors in the pathophysiology of irritable bowel syndrome. Dig. Liver Dis. 2009, 41, 788–793. [Google Scholar] [CrossRef]
- Cordaro, M.; Scuto, M.; Siracusa, R.; D’amico, R.; Filippo Peritore, A.; Gugliandolo, E.; Fusco, R.; Crupi, R.; Impellizzeri, D.; Pozzebon, M.; et al. Effect of N-palmitoylethanolamine-oxazoline on comorbid neuropsychiatric disturbance associated with inflammatory bowel disease. FASEB J. 2020, 34, 4085–4106. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, R.; Siracusa, R.; Fusco, R.; Cordaro, M.; Genovese, T.; Peritore, A.F.; Gugliandolo, E.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Protective effects of Colomast®, A new formulation of adelmidrol and sodium hyaluronate, in a mouse model of acute restraint stress. Int. J. Mol. Sci. 2020, 21, 8136. [Google Scholar] [CrossRef]
- Bradford, K.; Shih, W.; Videlock, E.J.; Presson, A.P.; Naliboff, B.D.; Mayer, E.A.; Chang, L. Association between early adverse life events and irritable bowel syndrome. Clin. Gastroenterol. Hepatol. 2012, 10, 385–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uusijärvi, A.; Bergström, A.; Simrén, M.; Ludvigsson, J.F.; Kull, I.; Wickman, M.; Alm, J.; Olén, O. Use of antibiotics in infancy and childhood and risk of recurrent abdominal pain—A Swedish birth cohort study. Neurogastroenterol. Motil. 2014, 26, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Fukui, H.; Oshima, T.; Tanaka, Y.; Oikawa, Y.; Makizaki, Y.; Ohno, H.; Tomita, T.; Watari, J.; Miwa, H. Effect of probiotic Bifidobacterium bifidum G9-1 on the relationship between gut microbiota profile and stress sensitivity in maternally separated rats. Sci. Rep. 2018, 8, 12384. [Google Scholar] [CrossRef]
- Kawahara, T.; Makizaki, Y.; Oikawa, Y.; Tanaka, Y.; Maeda, A.; Shimakawa, M.; Komoto, S.; Moriguchi, K.; Ohno, H.; Taniguchi, K. Oral administration of Bifidobacterium bifidum G9-1 alleviates rotavirus gastroenteritis through regulation of intestinal homeostasis by inducing mucosal protective factors. PLoS ONE 2017, 12, e0173979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makizaki, Y.; Maeda, A.; Oikawa, Y.; Tamura, S.; Tanaka, Y.; Nakajima, S.; Ohno, H.; Yamamura, H. Probiotic Bifidobacterium bifidum G9-1 ameliorates phytohemagglutinin-induced diarrhea caused by intestinal dysbiosis. Microbiol. Immunol. 2019, 63, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, T.; Oikawa, Y.; Kato, T.; Kessoku, T.; Kobayashi, T.; Kato, S.; Misawa, N.; Ashikari, K.; Fuyuki, A.; Ohkubo, H.; et al. The protective effect of Bifidobacterium bifidum G9-1 against mucus degradation by Akkermansia muciniphila following small intestine injury caused by a proton pump inhibitor and aspirin. Gut Microbes 2020, 11, 1385–1404. [Google Scholar] [CrossRef]
- Castoldi, A.; Favero de Aguiar, C.; Moraes-Vieira, P.M.; Olsen Saraiva Câmara, N. They must hold tight: Junction proteins, microbiota and immunity in intestinal mucosa. Curr. Protein Pept. Sci. 2015, 16, 655–671. [Google Scholar] [CrossRef]
- Gareau, M.G.; Jury, J.; MacQueen, G.; Sherman, P.M.; Perdue, M.H. Probiotic treatment of rat pups normalises corticosterone release and ameliorates colonic dysfunction induced by maternal separation. Gut 2007, 56, 1522–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukui, H.; Zhang, X.; Sun, C.; Hara, K.; Kikuchi, S.; Yamasaki, T.; Kondo, T.; Tomita, T.; Oshima, T.; Watari, J.; et al. IL-22 produced by cancer-associated fibroblasts promotes gastric cancer cell invasion via STAT3 and ERK signaling. Br. J. Cancer 2014, 111, 763–771. [Google Scholar] [CrossRef]
- Yamagishi, H.; Fukui, H.; Sekikawa, A.; Kono, T.; Fujii, S.; Ichikawa, K.; Tomita, S.; Imura, J.; Hiraishi, H.; Chiba, T.; et al. Expression profile of REG family proteins REG Iα and REG IV in advanced gastric cancer: Comparison with mucin phenotype and prognostic markers. Mod. Pathol. 2009, 22, 906–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Fukui, H.; Eda, H.; Xu, X.; Kitayama, Y.; Hara, K.; Kodani, M.; Tomita, T.; Oshima, T.; Watari, J.; et al. Involvement of gut microbiota in association between GLP-1/GLP-1 receptor expression and gastrointestinal motility. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G367–G373. [Google Scholar] [CrossRef] [Green Version]
- Inoue, Y.; Fukui, H.; Xu, X.; Ran, Y.; Tomita, T.; Oshima, T.; Watari, J.; Miwa, H. Colonic M1 macrophage is associated with the prolongation of gastrointestinal motility and obesity in mice treated with vancomycin. Mol. Med. Rep. 2019, 19, 2591–2598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosztoczy, A.; Fioramonti, J.; Jármay, K.; Barreau, F.; Wittmann, T.; Buéno, L. Influence of sex and experimental protocol on the effect of maternal deprivation on rectal sensitivity to distension in the adult rat. Neurogastroenterol. Motil. 2003, 15, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Kitayama, Y.; Fukui, H.; Hara, K.; Eda, H.; Kodani, M.; Yang, M.; Sun, C.; Yamagishi, H.; Tomita, T.; Oshima, T.; et al. Role of regenerating gene I in claudin expression and barrier function in the small intestine. Transl. Res. 2016, 173, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Ran, Y.; Fukui, H.; Xu, X.; Wang, X.; Ebisutani, N.; Tanaka, Y.; Maeda, A.; Makizaki, Y.; Ohno, H.; Kondo, T.; et al. Alteration of colonic mucosal permeability during antibiotic-induced dysbiosis. Int. J. Mol. Sci. 2020, 21, 6108. [Google Scholar] [CrossRef]
- Cho, H.S.; Park, J.M.; Lim, C.H.; Cho, Y.K.; Lee, I.S.; Kim, S.W.; Choi, M.G.; Chung, I.S.; Chung, Y.K. Anxiety, depression and quality of life in patients with irritable bowel syndrome. Gut Liver 2011, 5, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, S.W.; Auyeung, K.K.; Bian, Z.X.; Ko, J.K. Pathogenesis, experimental models and contemporary pharmacotherapy of irritable bowel syndrome: Story about the brain-gut axis. Curr. Neuropharmacol. 2016, 14, 842–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, Q.X.; Soh, A.Y.S.; Loke, W.; Lim, D.Y.; Yeo, W.S. The role of inflammation in irritable bowel syndrome (IBS). J. Inflamm. Res. 2018, 11, 345–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piche, T. Tight junctions and IBS—The link between epithelial permeability, low-grade inflammation, and symptom generation? Neurogastroenterol. Motil. 2014, 26, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Fukui, H.; Nishida, A.; Matsuda, S.; Kira, F.; Watanabe, S.; Kuriyama, M.; Kawakami, K.; Aikawa, Y.; Oda, N.; Arai, K.; et al. Usefulness of machine learning-based gut microbiome analysis for identifying patients with irritable bowels syndrome. J. Clin. Med. 2020, 9, 2403. [Google Scholar] [CrossRef]
- Coutinho, S.V.; Plotsky, P.M.; Sablad, M.; Miller, J.C.; Zhou, H.; Bayati, A.I.; McRoberts, J.A.; Mayer, E.A. Neonatal maternal separation alters stress-induced responses to viscerosomatic nociceptive stimuli in rat. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 282, G307–G316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreau, F.; Ferrier, L.; Fioramonti, J.; Bueno, L. New insights in the etiology and pathophysiology of irritable bowel syndrome: Contribution of neonatal stress models. Pediatr. Res. 2007, 62, 240–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, J.; Feldon, J. Long-term biobehavioral effects of maternal separation in the rat: Consistent or confusing? Rev. Neurosci. 2000, 11, 383–408. [Google Scholar] [CrossRef]
- De Palma, G.; Blennerhassett, P.; Lu, J.; Deng, Y.; Park, A.J.; Green, W.; Denou, E.; Silva, M.A.; Santacruz, A.; Sanz, Y.; et al. Microbiota and host determinants of behavioural phenotype in maternally separated mice. Nat. Commun. 2015, 6, 7735. [Google Scholar] [CrossRef] [Green Version]
- Capaldo, C.T.; Powell, D.N.; Kalman, D. Layered defense: How mucus and tight junctions seal the intestinal barrier. J. Mol. Med. 2017, 95, 927–934. [Google Scholar] [CrossRef] [Green Version]
- Rodiño-Janeiro, B.K.; Alonso-Cotoner, C.; Pigrau, M.; Lobo, B.; Vicario, M.; Santos, J. Role of corticotropin-releasing factor in gastrointestinal permeability. J. Neurogastroenterol. Motil. 2015, 21, 33–50. [Google Scholar] [CrossRef] [Green Version]
- Chichlowski, M.; Hale, L.P. Bacterial-mucosal interactions in inflammatory bowel disease: An alliance gone bad. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G1139–G1149. [Google Scholar] [CrossRef] [Green Version]
- Ling, X.; Linglong, P.; Weixia, D.; Hong, W. Protective effects of Bifidobacterium on intestinal barrier function in LPS-induced enterocyte barrier injury of Caco-2 monolayers and in a rat NEC model. PLoS ONE 2016, 11, e0161635. [Google Scholar] [CrossRef]
- Khailova, L.; Mount Patrick, S.K.; Arganbright, K.M.; Halpern, M.D.; Kinouchi, T.; Dvorak, B. Bifidobacterium bifidum reduces apoptosis in the intestinal epithelium in necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G1118–G1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroeder, B.O.; Birchenough, G.M.H.; Ståhlman, M.; Arike, L.; Johansson, M.E.V.; Hansson, G.C.; Bäckhed, F. Bifidobacteria or fiber protects against diet-induced microbiota-mediated colonic mucus deterioration. Cell Host Microbe 2018, 23, 27–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, M.; Wasan, A.; Sharma, R.K. Recent developments in probiotics: An emphasis on Bifidobacterium. Food Biosci. 2021, 41, 100993. [Google Scholar] [CrossRef]
- Luo, Y.; Xiao, Y.; Zhao, J.; Zhang, H.; Chen, W.; Zhai, Q. The role of mucin and oligosaccharides via cross-feeding activities by Bifidobacterium. Int. J. Biol. Macromol. 2021, 167, 1329–1337. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Jeong, Y.; Kang, S.; You, H.J.; Ji, G.E. Co-culture with Bifidobacterium catenulatum improves the growth, gut colonization, and butyrate production of Faecalibacterium prausnitzii: In vitro and in vivo studies. Microorganisms 2020, 8, 788. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Ran, X.; Li, B.; Li, Y.; He, D.; Huang, B.; Fu, S.; Liu, J.; Wang, W. Sodium butyrate inhibits inflammation and maintains epithelium barrier integrity in a TNBS-induced inflammatory bowel disease mice model. EBioMedicine 2018, 30, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, N.; Bai, C.; Song, S.; Zhang, Y.; Wang, B.; Li, Z. Bifidobacterium plays a protective role in TNF-α-induced inflammatory response in Caco-2 cell through NF-κB and p38MAPK pathways. Mol. Cell Biochem. 2020, 464, 83–91. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Direction | Primer Sequence |
|---|---|---|
| IL-1β | Forward Reverse | 5′-AATGCCTCGTGCTGTCTGACC-3′ 5′-GGGTGGGTGTGCCGTCTTTC-3′ |
| IL-6 | Forward Reverse | 5′-TCCTACCCCAACTTCCAATGCTC-3′ 5′-TTGGATGGTCTTGGTCCTTAGCC-3′ |
| IL-18 | Forward Reverse | 5′-AAACCCGCCTGTGTTCGA-3′ 5′-TCAGTCTGGTCTGGGATTCGT-3′ |
| IFN-γ | Forward Reverse | 5′-AGGTGAACAACCCACAGAT-3′ 5′-CTTCTTATTGGCACACTCTCTAC-3′ |
| TNF-α | Forward Reverse | 5′-TGGCGTGTTCATCCGTTCTCTACC-3′ 5′-CCCGCAATCCAGGCCACTACTT-3′ |
| ZO-1 | Forward Reverse | 5′-GGAAACCCGAAACTGATGCTATGG-3′ 5′-AACTGGCTGGCTGTACTGTGAG-3′ |
| occludin | Forward Reverse | 5′-AGCAACGATAACCTAGAGACA-3′ 5′-TGTCTCTGTTGATCTGAAGTG-3′ |
| claudin 3 | Forward Reverse | 5′-GGGTTGTACGTGGGCTGGGC-3′ 5′-GTGGATCGCGGCGCGGAATA-3′ |
| claudin 4 | Forward Reverse | 5′-GCCAGCAACTATGTGTAAG-3′ 5′-GCCGTTATGAGTTCAATCC-3′ |
| GAPDH | Forward Reverse | 5′-CTTGGGCTACACTGAGGACC-3′ 5′-CTGTTGCTGTAGCCGTATTC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Fukui, H.; Ran, Y.; Xu, X.; Ebisutani, N.; Nakanishi, T.; Tanaka, Y.; Maeda, A.; Makizaki, Y.; Tomita, T.; et al. Probiotic Bifidobacterium bifidum G9-1 Has a Preventive Effect on the Acceleration of Colonic Permeability and M1 Macrophage Population in Maternally Separated Rats. Biomedicines 2021, 9, 641. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9060641
Wang X, Fukui H, Ran Y, Xu X, Ebisutani N, Nakanishi T, Tanaka Y, Maeda A, Makizaki Y, Tomita T, et al. Probiotic Bifidobacterium bifidum G9-1 Has a Preventive Effect on the Acceleration of Colonic Permeability and M1 Macrophage Population in Maternally Separated Rats. Biomedicines. 2021; 9(6):641. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9060641
Chicago/Turabian StyleWang, Xuan, Hirokazu Fukui, Ying Ran, Xin Xu, Nobuhiko Ebisutani, Takashi Nakanishi, Yoshiki Tanaka, Ayako Maeda, Yutaka Makizaki, Toshihiko Tomita, and et al. 2021. "Probiotic Bifidobacterium bifidum G9-1 Has a Preventive Effect on the Acceleration of Colonic Permeability and M1 Macrophage Population in Maternally Separated Rats" Biomedicines 9, no. 6: 641. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9060641