
High Resolution Episcopic Microscopy for Qualitative and Quantitative Data in Phenotyping Altered Embryos and Adult Mice Using the New “Histo3D” System

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
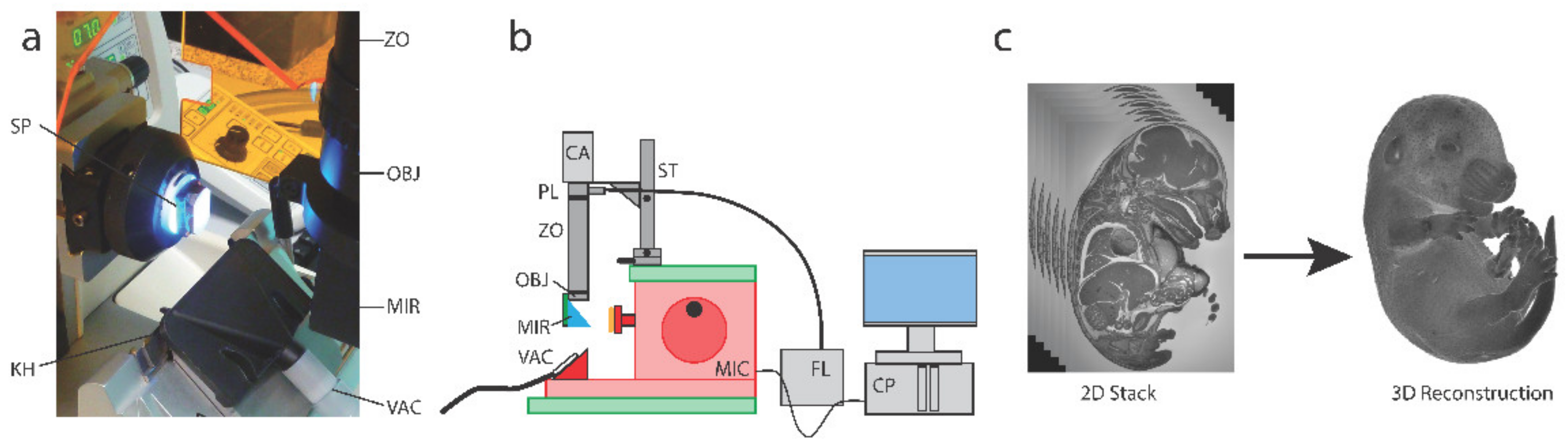
2.1. Sample Preparation and HREM Data Generation
2.2. Image Analysis and Quantification
2.2.1. Embryo Data Generation
2.2.2. Aorta Data Generation
2.2.3. Knee Joint Data Generation
3. Results
3.1. Qualitative and Quantitative Morphological Assessment in Mouse Developmental Pathology Studies
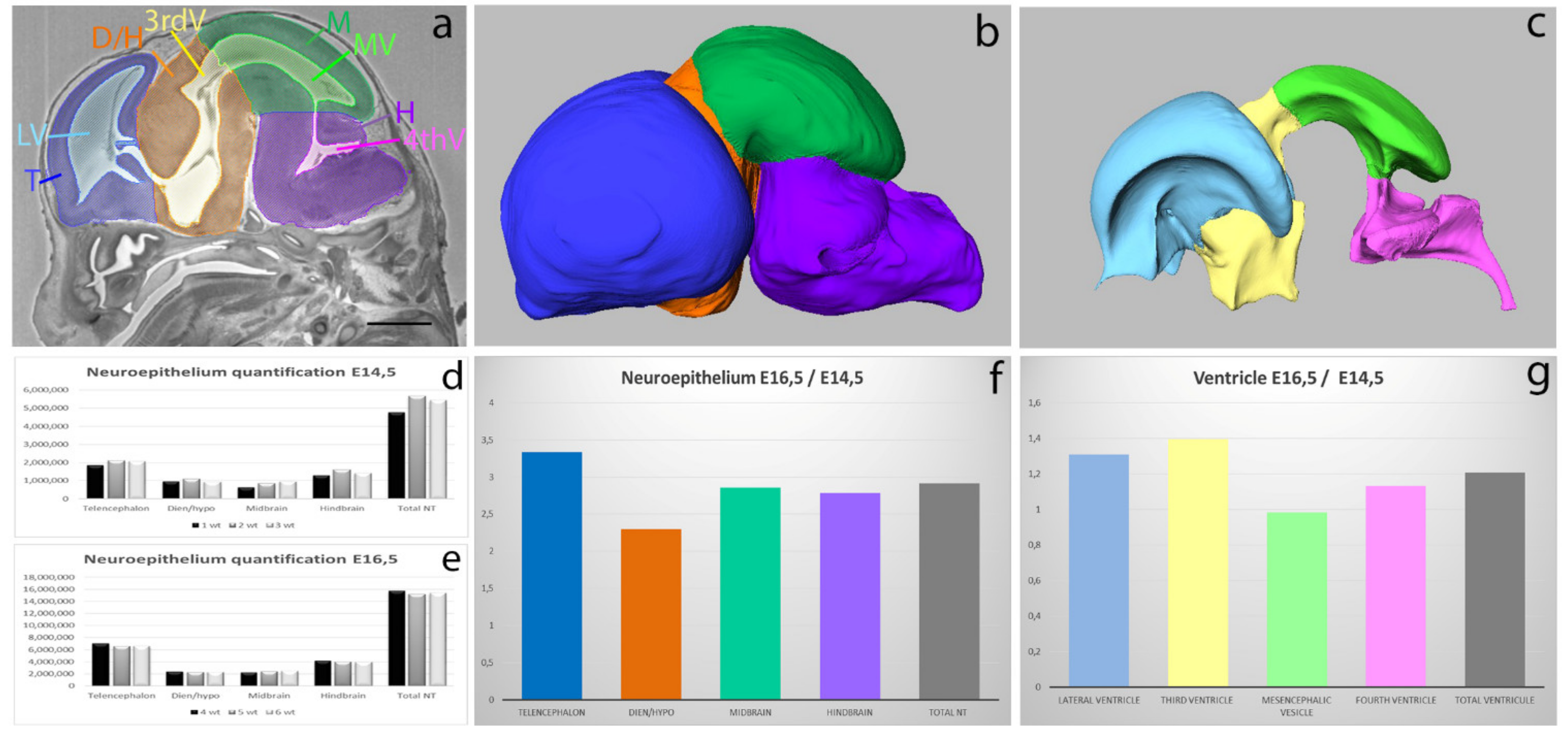
3.1.1. Neuroepithelium and Ventricular System Volume Quantification at Two Developmental Stages E14.5 and E16.5 in the Mouse
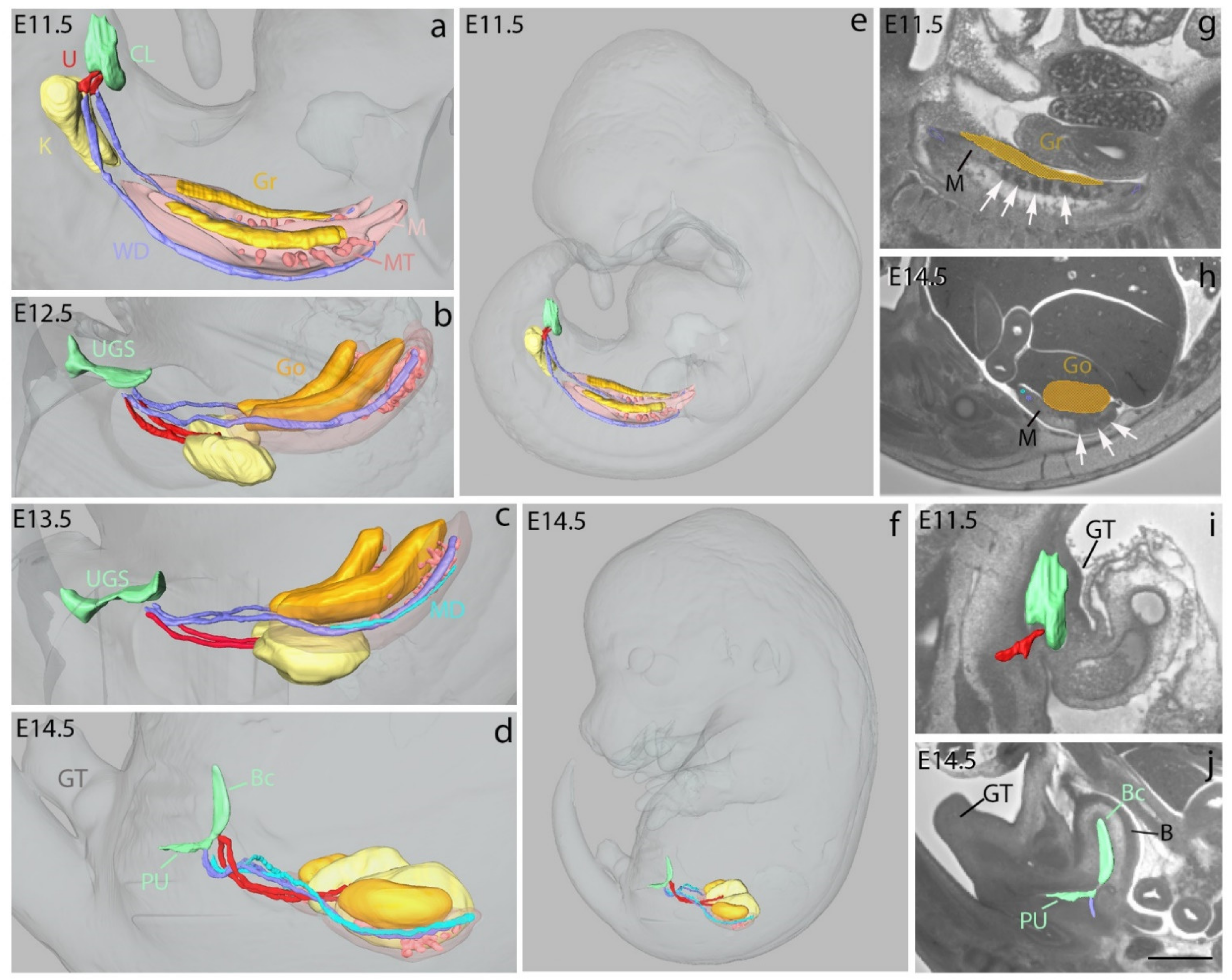
3.1.2. 3D Reconstructions of Cranial Nerves and Ganglia in Embryos and Fetuses
3.1.3. 3D Reconstructions of Urogenital System Development
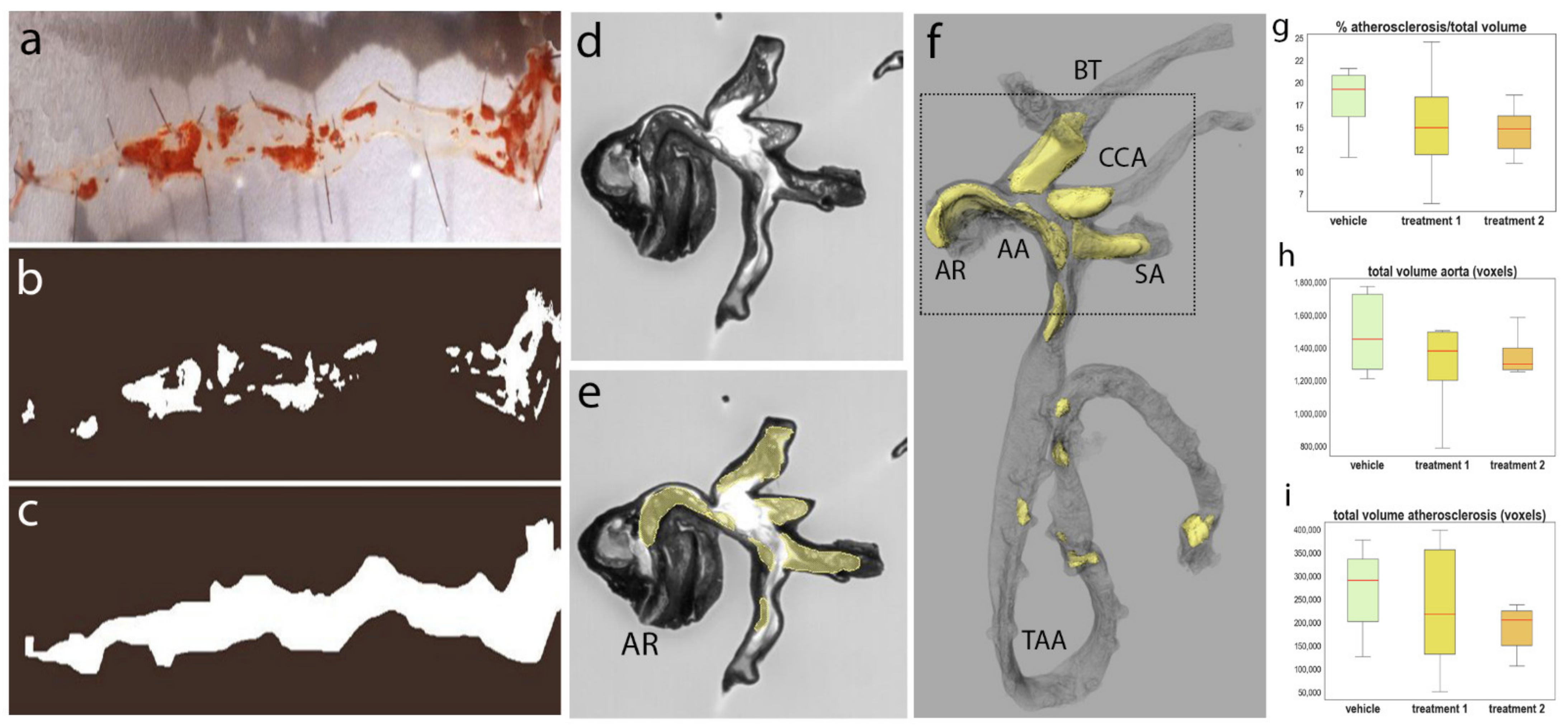
3.2. Quantification of Atherosclerotic Plaques in Adult Mouse Aortas
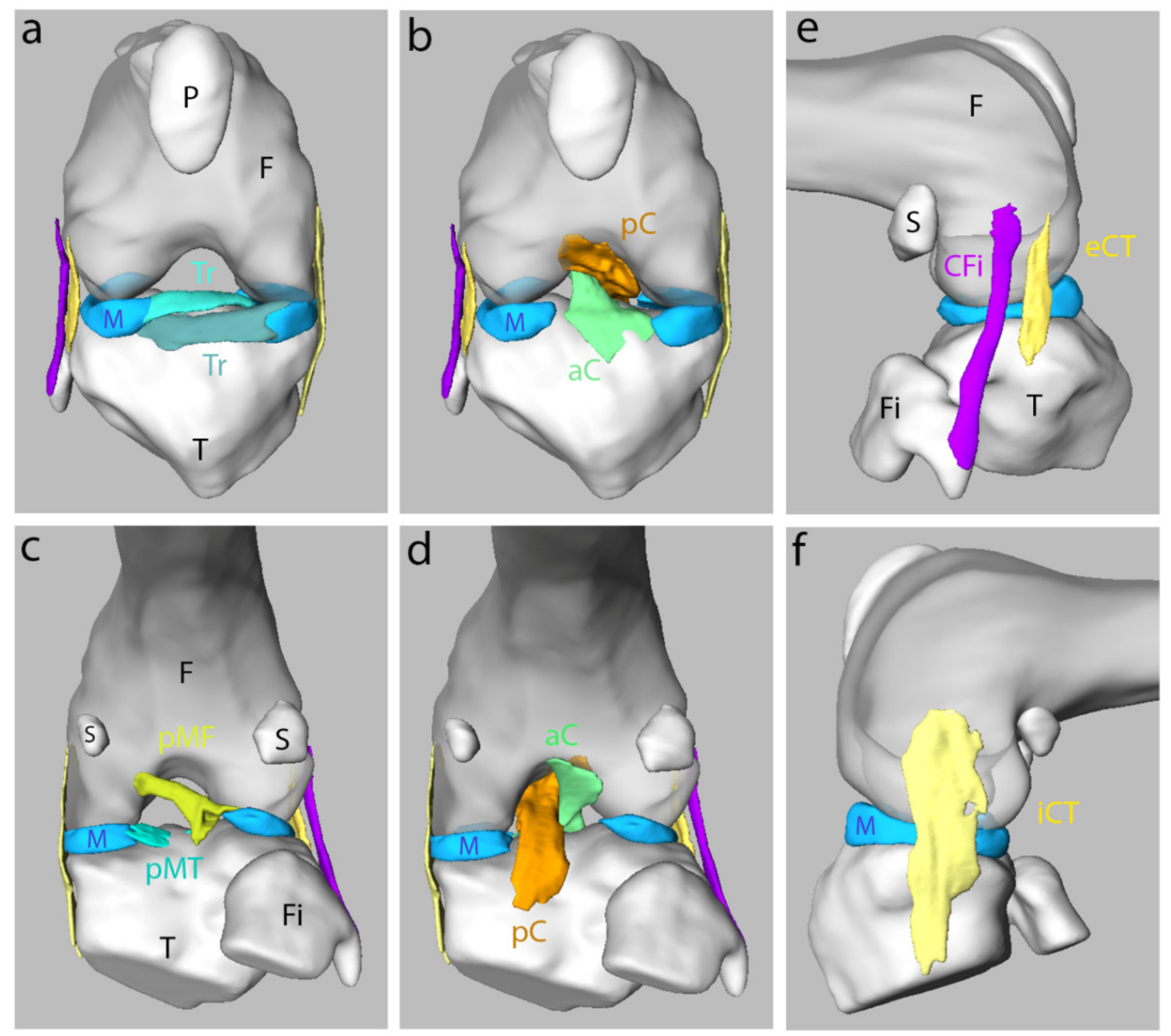
3.3. Morphology and Shape of Ligaments in the Knee Joint of the Adult Mouse
4. Discussion
5. Conclusions
6. Construction of Histo3D
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turgeon, B.; Meloche, S. Interpreting neonatal lethal phenotypes in mouse mutants: Insights into gene function and human diseases. Physiol. Rev. 2009, 89, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickinson, M.E.; Flenniken, A.M.; Ji, X.; Teboul, L.; Wong, M.D.; White, J.K.; Meehan, T.F.; Weninger, W.J.; Westerberg, H.; Adissu, H.; et al. High-throughput discovery of novel developmental phenotypes. Nature 2016, 537, 508–514. [Google Scholar] [CrossRef]
- Weninger, W.J.; Streicher, J.; Muller, G.B. 3-dimensional reconstruction of histological serial sections using a computer. Wien. Klin. Wochenschr. 1996, 108, 515–520. [Google Scholar]
- De Bakker, B.S.; de Jong, K.H.; Hagoort, J.; de Bree, K.; Besselink, C.T.; de Kanter, F.E.; Veldhuis, T.; Bais, B.; Schildmeijer, R.; Ruijter, J.M.; et al. An interactive three-dimensional digital atlas and quantitative database of human development. Science 2016, 354, 6315. [Google Scholar] [CrossRef]
- Young, D.M.; Fazel Darbandi, S.; Schwartz, G.; Bonzell, Z.; Yuruk, D.; Nojima, M.; Gole, L.C.; Rubenstein, J.L.; Yu, W.; Sanders, S.J. Constructing and optimizing 3D atlases from 2D data with application to the developing mouse brain. Elife 2021, 10, e61408. [Google Scholar] [CrossRef] [PubMed]
- Petiet, A.E.; Kaufman, M.H.; Goddeeris, M.M.; Brandenburg, J.; Elmore, S.A.; Johnson, G.A. High-resolution magnetic resonance histology of the embryonic and neonatal mouse: A 4D atlas and morphologic database. Proc. Natl. Acad. Sci. USA 2008, 105, 12331–12336. [Google Scholar] [CrossRef] [Green Version]
- Cleary, J.O.; Modat, M.; Norris, F.C.; Price, A.N.; Jayakody, S.A.; Martinez-Barbera, J.P.; Greene, N.D.; Hawkes, D.J.; Ordidge, R.J.; Scambler, P.J.; et al. Magnetic resonance virtual histology for embryos: 3D atlases for automated high-throughput phenotyping. Neuroimage 2011, 54, 769–778. [Google Scholar] [CrossRef]
- Johnson, J.T.; Hansen, M.S.; Wu, I.; Healy, L.J.; Johnson, C.R.; Jones, G.M.; Capecchi, M.R.; Keller, C. Virtual histology of transgenic mouse embryos for high-throughput phenotyping. PLoS Genet. 2006, 2, e61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degenhardt, K.; Wright, A.C.; Horng, D.; Padmanabhan, A.; Epstein, J.A. Rapid 3D phenotyping of cardiovascular development in mouse embryos by micro-CT with iodine staining. Circ. Cardiovasc. Imaging 2010, 3, 314–322. [Google Scholar] [CrossRef] [Green Version]
- Metscher, B.D.; Muller, G.B. MicroCT for molecular imaging: Quantitative visualization of complete three-dimensional distributions of gene products in embryonic limbs. Dev. Dyn. 2011, 240, 2301–2308. [Google Scholar] [CrossRef]
- Wong, M.D.; Dorr, A.E.; Walls, J.R.; Lerch, J.P.; Henkelman, R.M. A novel 3D mouse embryo atlas based on micro-CT. Development 2012, 139, 3248–3256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.W.; Wong, L.; Rasmussen, T.L.; Kalaga, S.; McElwee, M.L.; Keith, L.C.; Bohat, R.; Seavitt, J.R.; Beaudet, A.L.; Dickinson, M.E. Three-dimensional microCT imaging of mouse development from early post-implantation to early postnatal stages. Dev. Biol. 2016, 419, 229–236. [Google Scholar] [CrossRef] [Green Version]
- Ragan, T.; Kadiri, L.R.; Venkataraju, K.U.; Bahlmann, K.; Sutin, J.; Taranda, J.; Arganda-Carreras, I.; Kim, Y.; Seung, H.S.; Osten, P. Serial two-photon tomography for automated ex vivo mouse brain imaging. Nat. Methods 2012, 9, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Dodt, H.U.; Leischner, U.; Schierloh, A.; Jahrling, N.; Mauch, C.P.; Deininger, K.; Deussing, J.M.; Eder, M.; Zieglgansberger, W.; Becker, K. Ultramicroscopy: Three-dimensional visualization of neuronal networks in the whole mouse brain. Nat. Methods 2007, 4, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Renier, N.; Wu, Z.; Simon, D.J.; Yang, J.; Ariel, P.; Tessier-Lavigne, M. iDISCO: A simple, rapid method to immunolabel large tissue samples for volume imaging. Cell 2014, 159, 896–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belle, M.; Godefroy, D.; Dominici, C.; Heitz-Marchaland, C.; Zelina, P.; Hellal, F.; Bradke, F.; Chedotal, A. A simple method for 3D analysis of immunolabeled axonal tracts in a transparent nervous system. Cell Rep. 2014, 9, 1191–1201. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, J.; Ahlgren, U.; Perry, P.; Hill, B.; Ross, A.; Hecksher-Sorensen, J.; Baldock, R.; Davidson, D. Optical projection tomography as a tool for 3D microscopy and gene expression studies. Science 2002, 296, 541–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharpe, J. Optical projection tomography as a new tool for studying embryo anatomy. J. Anat. 2003, 202, 175–181. [Google Scholar] [CrossRef]
- Anderson, G.A.; Wong, M.D.; Yang, J.; Henkelman, R.M. 3D imaging, registration, and analysis of the early mouse embryonic vasculature. Dev. Dyn. 2013, 242, 527–538. [Google Scholar] [CrossRef]
- Weninger, W.J.; Geyer, S.H.; Mohun, T.J.; Rasskin-Gutman, D.; Matsui, T.; Ribeiro, I.; Costa Lda, F.; Izpisua-Belmonte, J.C.; Muller, G.B. High-resolution episcopic microscopy: A rapid technique for high detailed 3D analysis of gene activity in the context of tissue architecture and morphology. Anat. Embryol. 2006, 211, 213–221. [Google Scholar] [CrossRef]
- Mohun, T.J.; Weninger, W.J. Episcopic three-dimensional imaging of embryos. Cold Spring Harb. Protoc. 2012, 2012, 641–646. [Google Scholar] [CrossRef]
- Geyer, S.H.; Mohun, T.J.; Weninger, W.J. Visualizing vertebrate embryos with episcopic 3D imaging techniques. Sci. World J. 2009, 9, 1423–1437. [Google Scholar] [CrossRef]
- Weninger, W.J.; Maurer-Gesek, B.; Reissig, L.F.; Prin, F.; Wilson, R.; Galli, A.; Adams, D.J.; White, J.K.; Mohun, T.J.; Geyer, S.H. Visualising the Cardiovascular System of Embryos of Biomedical Model Organisms with High Resolution Episcopic Microscopy (HREM). J. Cardiovasc. Dev. Dis. 2018, 5, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geyer, S.H.; Weninger, W.J. High-Resolution Episcopic Microscopy (HREM): Looking Back on 13 Years of Successful Generation of Digital Volume Data of Organic Material for 3D Visualisation and 3D Display. Appl. Sci. 2019, 9, 3826. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.; Geyer, S.H.; Reissig, L.; Rose, J.; Szumska, D.; Hardman, E.; Prin, F.; McGuire, C.; Ramirez-Solis, R.; White, J.; et al. Highly variable penetrance of abnormal phenotypes in embryonic lethal knockout mice. Wellcome Open Res. 2016, 1, 1. [Google Scholar] [CrossRef] [Green Version]
- Weninger, W.J.; Geyer, S.H.; Martineau, A.; Galli, A.; Adams, D.J.; Wilson, R.; Mohun, T.J. Phenotyping structural abnormalities in mouse embryos using high-resolution episcopic microscopy. Dis. Model. Mech. 2014, 7, 1143–1152. [Google Scholar] [CrossRef] [Green Version]
- Reissig, L.F.; Seyedian Moghaddam, A.; Prin, F.; Wilson, R.; Galli, A.; Tudor, C.; White, J.K.; Geyer, S.H.; Mohun, T.J.; Weninger, W.J. Hypoglossal Nerve Abnormalities as Biomarkers for Central Nervous System Defects in Mouse Lines Producing Embryonically Lethal Offspring. Front. Neuroanat. 2021, 15, 625716. [Google Scholar] [CrossRef]
- Geyer, S.H.; Nohammer, M.M.; Tinhofer, I.E.; Weninger, W.J. The dermal arteries of the human thumb pad. J. Anat. 2013, 223, 603–609. [Google Scholar] [CrossRef]
- Geyer, S.H.; Nohammer, M.M.; Matha, M.; Reissig, L.; Tinhofer, I.E.; Weninger, W.J. High-resolution episcopic microscopy (HREM): A tool for visualizing skin biopsies. Microsc. Microanal. 2014, 20, 1356–1364. [Google Scholar] [CrossRef] [PubMed]
- Geyer, S.H.; Maurer-Gesek, B.; Reissig, L.F.; Weninger, W.J. High-resolution episcopic microscopy (HREM)-simple and robust protocols for processing and visualizing organic materials. J. Vis. Exp. JoVE 2017, 56071. [Google Scholar] [CrossRef] [Green Version]
- Mohun, T.J.; Weninger, W.J. Embedding embryos for high-resolution episcopic microscopy (HREM). Cold Spring Harb. Protoc. 2012, 2012, 678–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gato, A.; Desmond, M.E. Why the embryo still matters: CSF and the neuroepithelium as interdependent regulators of embryonic brain growth, morphogenesis and histiogenesis. Dev. Biol. 2009, 327, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Gato, A.; Alonso, M.I.; Martin, C.; Carnicero, E.; Moro, J.A.; De la Mano, A.; Fernandez, J.M.; Lamus, F.; Desmond, M.E. Embryonic cerebrospinal fluid in brain development: Neural progenitor control. Croat. Med. J. 2014, 55, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Aristizabal, O.; Mamou, J.; Ketterling, J.A.; Turnbull, D.H. High-throughput, high-frequency 3-D ultrasound for in utero analysis of embryonic mouse brain development. Ultrasound Med. Biol. 2013, 39, 2321–2332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Autuori, M.C.; Pai, Y.J.; Stuckey, D.J.; Savery, D.; Marconi, A.M.; Massa, V.; Lythgoe, M.F.; Copp, A.J.; David, A.L.; Greene, N.D. Use of high-frequency ultrasound to study the prenatal development of cranial neural tube defects and hydrocephalus in Gldc-deficient mice. Prenat. Diagn. 2017, 37, 273–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parasoglou, P.; Berrios-Otero, C.A.; Nieman, B.J.; Turnbull, D.H. High-resolution MRI of early-stage mouse embryos. NMR Biomed. 2013, 26, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Ishihara, K.; Amano, K.; Takaki, E.; Shimohata, A.; Sago, H.; Epstein, C.J.; Yamakawa, K. Enlarged brain ventricles and impaired neurogenesis in the Ts1Cje and Ts2Cje mouse models of Down syndrome. Cereb. Cortex 2010, 20, 1131–1143. [Google Scholar] [CrossRef] [Green Version]
- Peterson, B.S.; Staib, L.; Scahill, L.; Zhang, H.; Anderson, C.; Leckman, J.F.; Cohen, D.J.; Gore, J.C.; Albert, J.; Webster, R. Regional brain and ventricular volumes in Tourette syndrome. Arch. Gen. Psychiatry 2001, 58, 427–440. [Google Scholar] [CrossRef] [Green Version]
- Le Douarin, N.M. Cell line segregation during peripheral nervous system ontogeny. Science 1986, 231, 1515–1522. [Google Scholar] [CrossRef]
- Vogel, K.S. Sensory Neurons: Diversity, Development and Plasticity in Origins and Early Development of Vertebrate Cranial Sensory Neurons; Oxford University Press: Oxford, UK, 1992. [Google Scholar]
- Webb, J.F.; Noden, D.M. Ectodermal placodes contributions to the development of the vertebrate head. Am. Zool. 1993, 33, 434–447. [Google Scholar] [CrossRef]
- Swiatek, P.J.; Gridley, T. Perinatal lethality and defects in hindbrain development in mice homozygous for a targeted mutation of the zinc finger gene Krox20. Genes Dev. 1993, 7, 2071–2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider-Maunoury, S.; Topilko, P.; Seitandou, T.; Levi, G.; Cohen-Tannoudji, M.; Pournin, S.; Babinet, C.; Charnay, P. Disruption of Krox-20 results in alteration of rhombomeres 3 and 5 in the developing hindbrain. Cell 1993, 75, 1199–1214. [Google Scholar] [CrossRef]
- Ma, Q.; Chen, Z.; del Barco Barrantes, I.; de la Pompa, J.L.; Anderson, D.J. neurogenin1 is essential for the determination of neuronal precursors for proximal cranial sensory ganglia. Neuron 1998, 20, 469–482. [Google Scholar] [CrossRef] [Green Version]
- McEvilly, R.J.; Erkman, L.; Luo, L.; Sawchenko, P.E.; Ryan, A.F.; Rosenfeld, M.G. Requirement for Brn-3.0 in differentiation and survival of sensory and motor neurons. Nature 1996, 384, 574–577. [Google Scholar] [CrossRef]
- Eng, S.R.; Gratwick, K.; Rhee, J.M.; Fedtsova, N.; Gan, L.; Turner, E.E. Defects in sensory axon growth precede neuronal death in Brn3a-deficient mice. J. Neurosci. 2001, 21, 541–549. [Google Scholar] [CrossRef]
- Huang, E.J.; Zang, K.; Schmidt, A.; Saulys, A.; Xiang, M.; Reichardt, L.F. POU domain factor Brn-3a controls the differentiation and survival of trigeminal neurons by regulating Trk receptor expression. Development 1999, 126, 2869–2882. [Google Scholar] [CrossRef] [PubMed]
- Farinas, I.; Reichardt, L.F. Neurotrophic factors and their receptors: Implications of genetic studies. Semin. Neurosci. 1996, 8, 133–143. [Google Scholar] [CrossRef] [Green Version]
- McNeill, E.M.; Roos, K.P.; Moechars, D.; Clagett-Dame, M. Nav2 is necessary for cranial nerve development and blood pressure regulation. Neural Dev. 2010, 5, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fei, D.; Huang, T.; Krimm, R.F. The neurotrophin receptor p75 regulates gustatory axon branching and promotes innervation of the tongue during development. Neural Dev. 2014, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Vermeiren, S.; Bellefroid, E.J.; Desiderio, S. Vertebrate Sensory Ganglia: Common and Divergent Features of the Transcriptional Programs Generating Their Functional Specialization. Front. Cell Dev. Biol. 2020, 8, 1026. [Google Scholar] [CrossRef] [PubMed]
- Jährling, N.; Becker, K.; Saghafi, S.; Dodt, H.-U. Light-sheet fluorescence microscopy: Chemical clearing and labeling protocols for ultramicroscopy. In Light Microscopy; Springer: Berlin/Heidelberg, Germany, 2017; pp. 33–49. [Google Scholar]
- Erturk, A.; Becker, K.; Jahrling, N.; Mauch, C.P.; Hojer, C.D.; Egen, J.G.; Hellal, F.; Bradke, F.; Sheng, M.; Dodt, H.U. Three-dimensional imaging of solvent-cleared organs using 3DISCO. Nat. Protoc. 2012, 7, 1983–1995. [Google Scholar] [CrossRef]
- Belle, M.; Godefroy, D.; Couly, G.; Malone, S.A.; Collier, F.; Giacobini, P.; Chedotal, A. Tridimensional Visualization and Analysis of Early Human Development. Cell 2017, 169, 161–173.e112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meziane, H.; Birling, M.C.; Wendling, O.; Leblanc, S.; Dubos, A.; Selloum, M.; Vasseur, L.; Pavlovic, G.; Sorg, T.; Kalscheuer, V.M.; et al. Large Scale functional evaluation of genes involved in rare diseases with intellectual disabilities unravelled unique behavioural profiles in mouse. Nat. Commun. 2021, submitted. [Google Scholar]
- Georgas, K.M.; Armstrong, J.; Keast, J.R.; Larkins, C.E.; McHugh, K.M.; Southard-Smith, E.M.; Cohn, M.J.; Batourina, E.; Dan, H.; Schneider, K.; et al. An illustrated anatomical ontology of the developing mouse lower urogenital tract. Development 2015, 142, 1893–1908. [Google Scholar] [CrossRef] [Green Version]
- Davidson, A.J. Mouse kidney development. Afr. Sci. 2019, 16, 171–204. [Google Scholar] [CrossRef]
- Elmore, S.A.; Kavari, S.L.; Hoenerhoff, M.J.; Mahler, B.; Scott, B.E.; Yabe, K.; Seely, J.C. Histology Atlas of the Developing Mouse Urinary System with Emphasis on Prenatal Days E10.5–E18.5. Toxicol. Pathol. 2019, 47, 865–886. [Google Scholar] [CrossRef] [PubMed]
- Mark, M.; Teletin, M.; Wendling, O.; Vonesch, J.L.; Ferret, B.; Herault, Y.; Ghyselinck, N.B. Pathogenesis of anorectal malformations in Retinoic Acid Receptor knockout mice studied by HREM. Biomedicines 2021, 46, 1396–1399. [Google Scholar]
- Nakashima, Y.; Plump, A.S.; Raines, E.W.; Breslow, J.L.; Ross, R. ApoE-deficient mice develop lesions of all phases of atherosclerosis throughout the arterial tree. Arterioscler. Thromb. 1994, 14, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Andres-Manzano, M.J.; Andres, V.; Dorado, B. Oil Red O and Hematoxylin and Eosin Staining for Quantification of Atherosclerosis Burden in Mouse Aorta and Aortic Root. Methods Mol. Biol. 2015, 1339, 85–99. [Google Scholar] [CrossRef]
- Lin, Y.; Bai, L.; Chen, Y.; Zhu, N.; Bai, Y.; Li, Q.; Zhao, S.; Fan, J.; Liu, E. Practical assessment of the quantification of atherosclerotic lesions in apoE(-)/(-) mice. Mol. Med. Rep. 2015, 12, 5298–5306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhury, R.P.; Aguinaldo, J.G.; Rong, J.X.; Kulak, J.L.; Kulak, A.R.; Reis, E.D.; Fallon, J.T.; Fuster, V.; Fisher, E.A.; Fayad, Z.A. Atherosclerotic lesions in genetically modified mice quantified in vivo by non-invasive high-resolution magnetic resonance microscopy. Atherosclerosis 2002, 162, 315–321. [Google Scholar] [CrossRef]
- McAteer, M.A.; Schneider, J.E.; Clarke, K.; Neubauer, S.; Channon, K.M.; Choudhury, R.P. Quantification and 3D reconstruction of atherosclerotic plaque components in apolipoprotein E knockout mice using ex vivo high-resolution MRI. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 2384–2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, C.; Christiansen, S.; Kaul, M.G.; Koziolek, E.; Reimer, R.; Heeren, J.; Adam, G.; Heine, M.; Ittrich, H. Quantitative and qualitative estimation of atherosclerotic plaque burden in vivo at 7T MRI using Gadospin F in comparison to en face preparation evaluated in ApoE KO mice. PLoS ONE 2017, 12, e0180407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd, D.J.; Helmering, J.; Kaufman, S.A.; Turk, J.; Silva, M.; Vasquez, S.; Weinstein, D.; Johnston, B.; Hale, C.; Veniant, M.M. A volumetric method for quantifying atherosclerosis in mice by using microCT: Comparison to en face. PLoS ONE 2011, 6, e18800. [Google Scholar] [CrossRef] [Green Version]
- Treuting, P.M.; Dintzis, S.M.; Montine, K.S. Comparative Anatomy and Histology, A Mouse, Rat and Human Atlas; Academic Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Ruberte, J.; Carretero, A.; Navarro, M. Morphological Mouse Phenotyping; Academic Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Weninger, W.; Maurer, B.; Zendron, B.; Dorfmeister, K.; Geyer, S. Measurements of the diameters of the great arteries and semi-lunar valves of chick and mouse embryos. J. Microsc. 2009, 234, 173–190. [Google Scholar] [CrossRef] [PubMed]
- Weninger, W.J.; Mohun, T.J. Three-dimensional analysis of molecular signals with episcopic imaging techniques. Methods Mol. Biol. 2007, 411, 35–46. [Google Scholar] [CrossRef]
- Walsh, C.; Holroyd, N.A.; Finnerty, E.; Ryan, S.G.; Sweeney, P.W.; Shipley, R.J.; Walker-Samuel, S. Multi-Fluorescence High-Resolution Episcopic Microscopy (MF-HREM) for Three-Dimensional Imaging of Adult Murine Organs. bioRxiv 2021. [Google Scholar] [CrossRef] [Green Version]
- Karl, T. The House Mouse: Atlas of Embryonic Development; Springer: New York, NY, USA, 1989. [Google Scholar]
- Kaufman, M.H. Atlas of Mouse Development; Academic Press: Cambridge, MA, USA, 1992. [Google Scholar]
- Geyer, S.H.; Reissig, L.; Rose, J.; Wilson, R.; Prin, F.; Szumska, D.; Ramirez-Solis, R.; Tudor, C.; White, J.; Mohun, T.J.; et al. A staging system for correct phenotype interpretation of mouse embryos harvested on embryonic day 14 (E14.5). J. Anat. 2017, 230, 710–719. [Google Scholar] [CrossRef] [Green Version]
- Karner, C.M.; Dietrich, M.F.; Johnson, E.B.; Kappesser, N.; Tennert, C.; Percin, F.; Wollnik, B.; Carroll, T.J.; Herz, J. Lrp4 regulates initiation of ureteric budding and is crucial for kidney formation—A mouse model for Cenani-Lenz syndrome. PLoS ONE 2010, 5, e10418. [Google Scholar] [CrossRef] [PubMed]
- Centa, M.; Ketelhuth, D.F.J.; Malin, S.; Gistera, A. Quantification of Atherosclerosis in Mice. J. Vis. Exp. 2019. [Google Scholar] [CrossRef]
- Antal, C.; Teletin, M.; Wendling, O.; Dgheem, M.; Auwerx, J.; Mark, M. Tissue collection for systematic phenotyping in the mouse. Curr. Protoc. Mol. Biol. 2007. [Google Scholar] [CrossRef] [PubMed]
- Brayton, C.; Justice, M.; Montgomery, C.A. Evaluating mutant mice: Anatomic pathology. Vet. Pathol. 2001, 38, 1–19. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zoom Position | Pixel Size (µm) | x-Axis FOV (mm) | y-Axis FOV (mm) | FOV (mm2) |
|---|---|---|---|---|
| 0.6 | 11.7 | 24 | 24 | 574.1 |
| 0.8 | 8 | 16.4 | 16.4 | 268.4 |
| 1 | 6.5 | 13.3 | 13.3 | 177.2 |
| 1.2 | 5.2 | 10.6 | 10.6 | 113.4 |
| 1.6 | 4 | 8.2 | 8.2 | 67.1 |
| 2 | 3.2 | 6.5 | 6.5 | 42.9 |
| 2.5 | 2.6 | 5.3 | 5.3 | 28.3 |
| 3.2 | 2 | 4.1 | 4.1 | 16.8 |
| 4 | 1.6 | 3.3 | 3.3 | 10.7 |
| 5 | 1.3 | 2.7 | 2.7 | 7.1 |
| 6.3 | 1.1 | 2.2 | 2.2 | 5.1 |
| 8 | 0.8 | 1.6 | 1.6 | 2.7 |
| 9.2 | 0.7 | 1.4 | 1.4 | 2.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wendling, O.; Hentsch, D.; Jacobs, H.; Lemercier, N.; Taubert, S.; Pertuy, F.; Vonesch, J.-L.; Sorg, T.; Di Michele, M.; Le Cam, L.; et al. High Resolution Episcopic Microscopy for Qualitative and Quantitative Data in Phenotyping Altered Embryos and Adult Mice Using the New “Histo3D” System. Biomedicines 2021, 9, 767. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9070767
Wendling O, Hentsch D, Jacobs H, Lemercier N, Taubert S, Pertuy F, Vonesch J-L, Sorg T, Di Michele M, Le Cam L, et al. High Resolution Episcopic Microscopy for Qualitative and Quantitative Data in Phenotyping Altered Embryos and Adult Mice Using the New “Histo3D” System. Biomedicines. 2021; 9(7):767. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9070767
Chicago/Turabian StyleWendling, Olivia, Didier Hentsch, Hugues Jacobs, Nicolas Lemercier, Serge Taubert, Fabien Pertuy, Jean-Luc Vonesch, Tania Sorg, Michela Di Michele, Laurent Le Cam, and et al. 2021. "High Resolution Episcopic Microscopy for Qualitative and Quantitative Data in Phenotyping Altered Embryos and Adult Mice Using the New “Histo3D” System" Biomedicines 9, no. 7: 767. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9070767