
Investigation of the lncRNA THOR in Mice Highlights the Importance of Noncoding RNAs in Mammalian Male Reproduction

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Ethical Statement
2.2. CRISPR/Cas9 sgRNA Preparation, Embryo Microinjection, and Embryo Transfer
2.3. Detection of THOR Deletion in Mouse Pups by PCR and Sequencing
2.4. T7E1 Cleavage Assay
2.5. Off-Target Analysis
2.6. RNA Isolation and qRT-PCR
2.7. Body Weight, Food Intake, Survival, and Statistical Analyses
2.8. Hormone Level Analysis
2.9. Histological Analysis
2.10. Morphological Analysis of Testes
2.11. Detection and Analysis of Offspring Fertility
2.12. Embryonic Development and Statistical Analyses
2.13. Sperm Motility and Morphological Analysis
2.14. In Vitro Fertilization (IVF)
2.15. Protein Isolation and Western Blotting
2.16. Statistical Analysis
3. Results
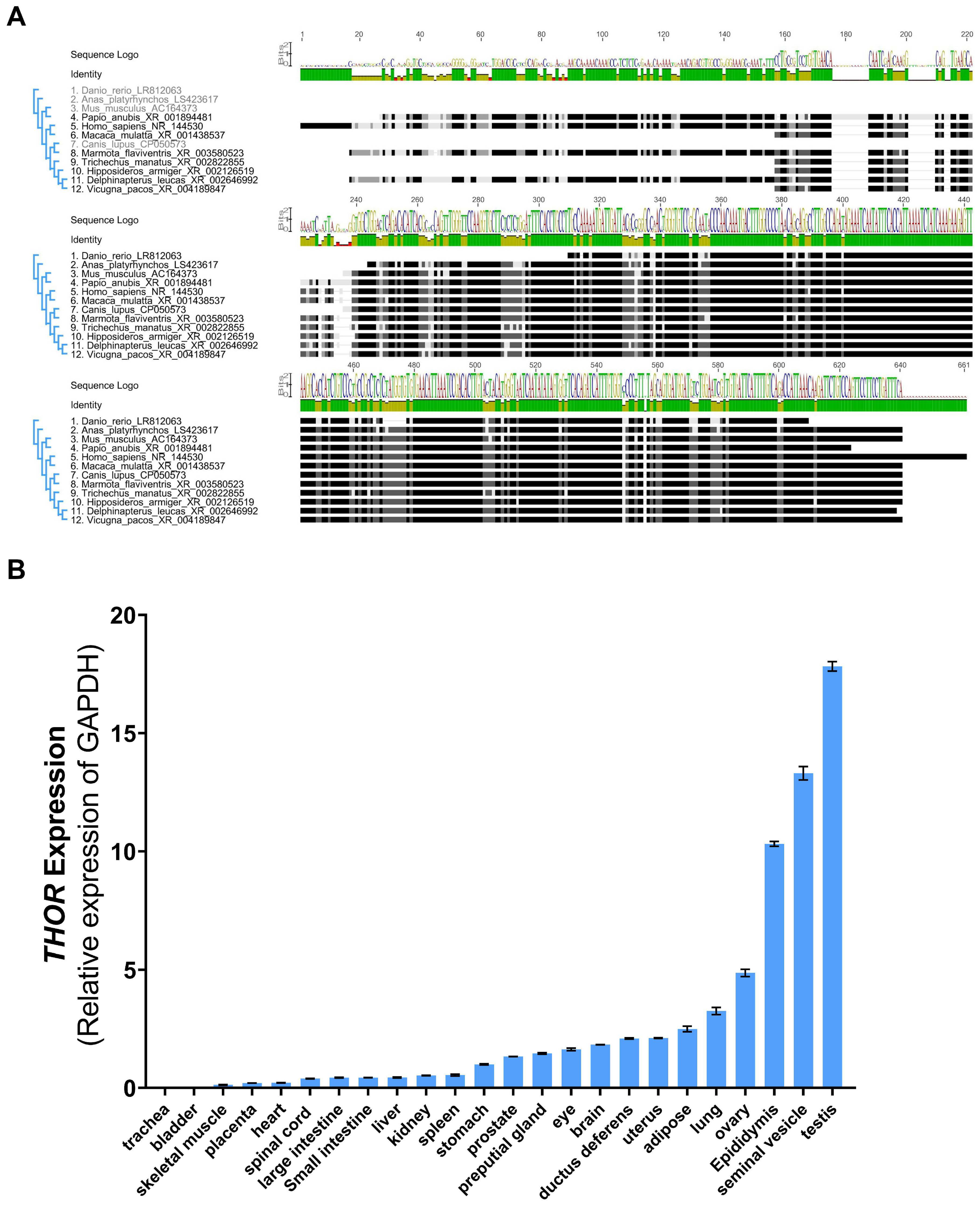
3.1. Evolutionary Relationship and Tissue Conservation of THOR in Vertebrates
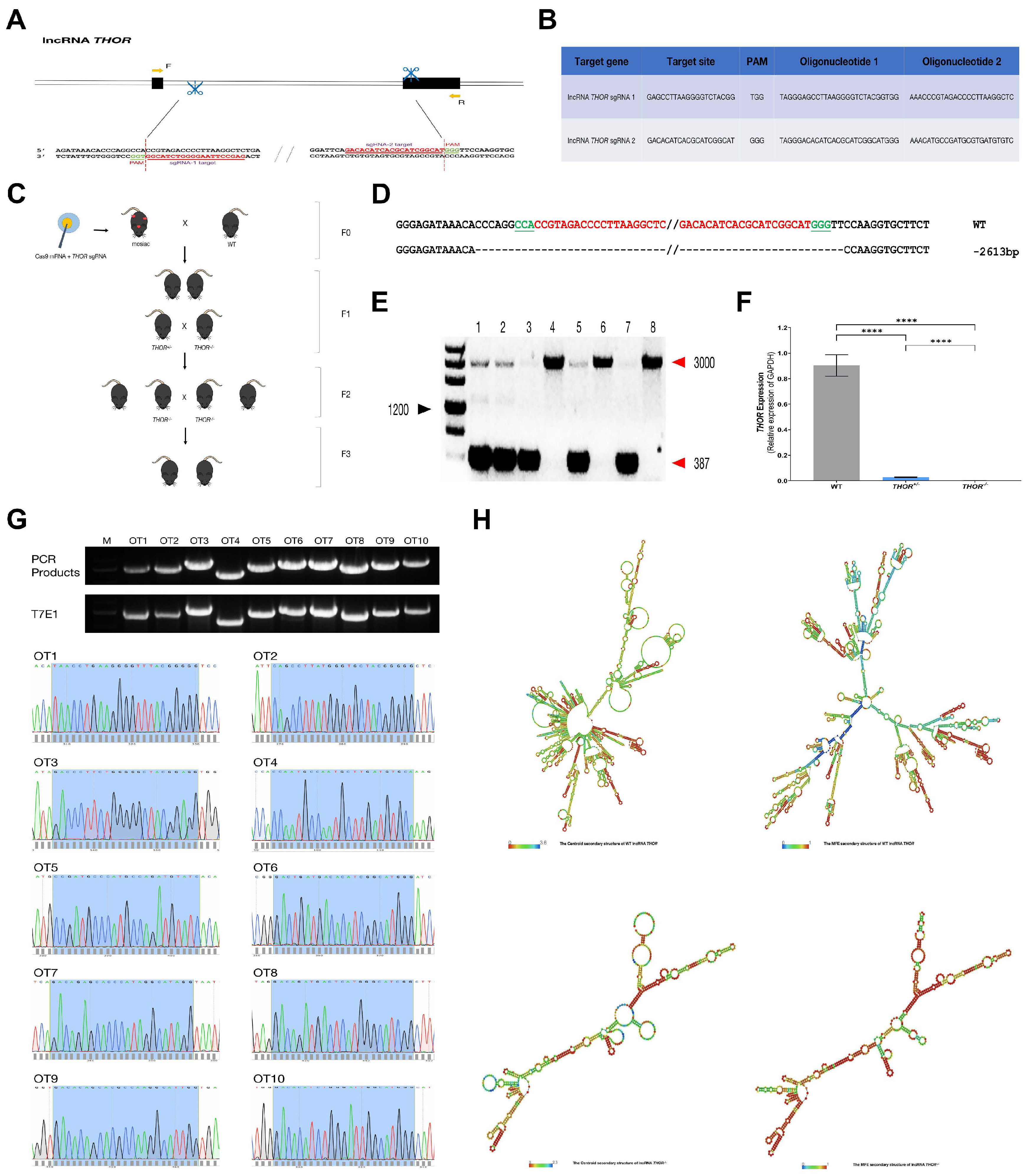
3.2. Generation of THOR-Deficient Mice
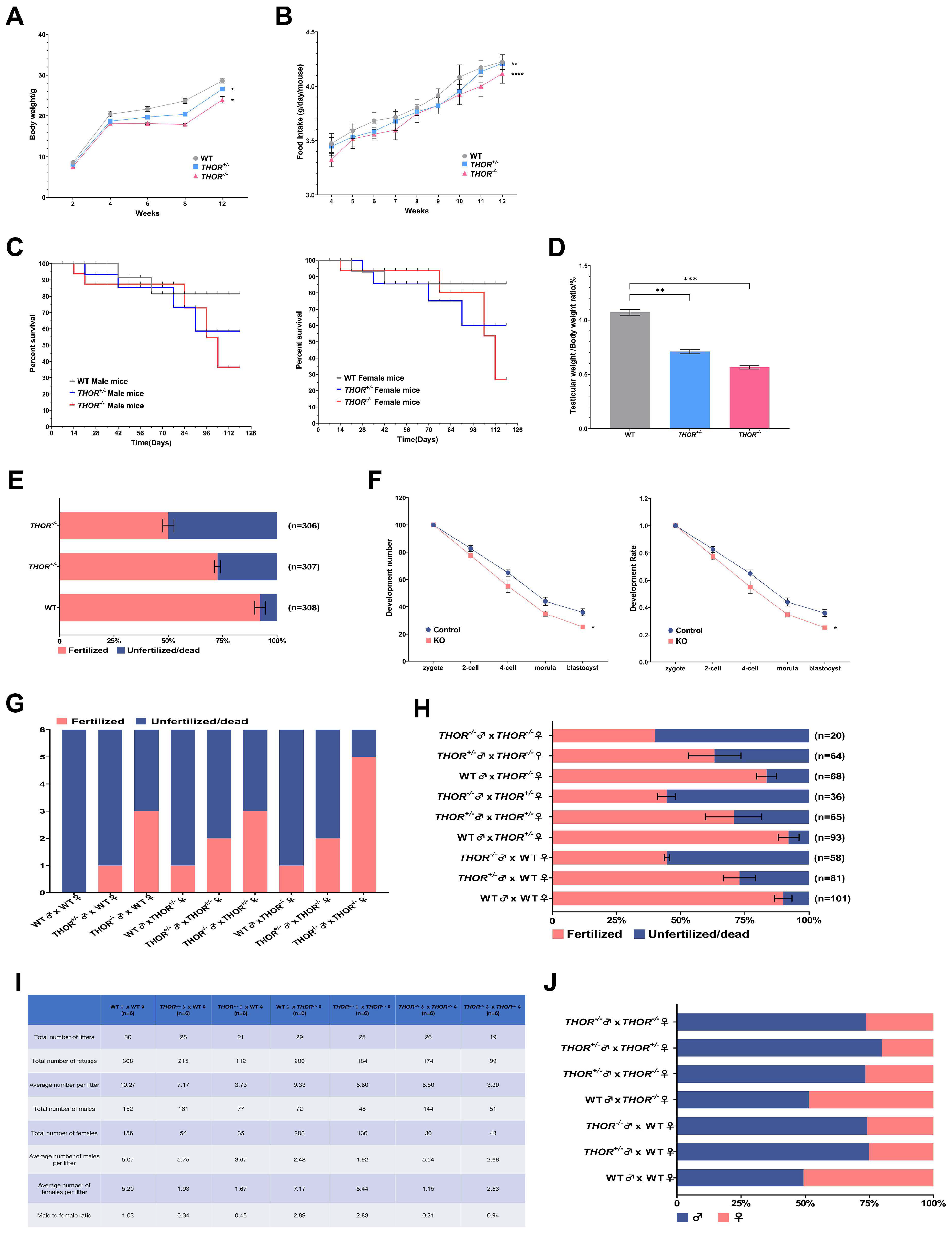
3.3. THOR-Deficient Mice Exhibit Decreased Fertility and a Female-to-Male Imbalance
3.4. THOR Deficiency Affects Testicular Development in Male Mice
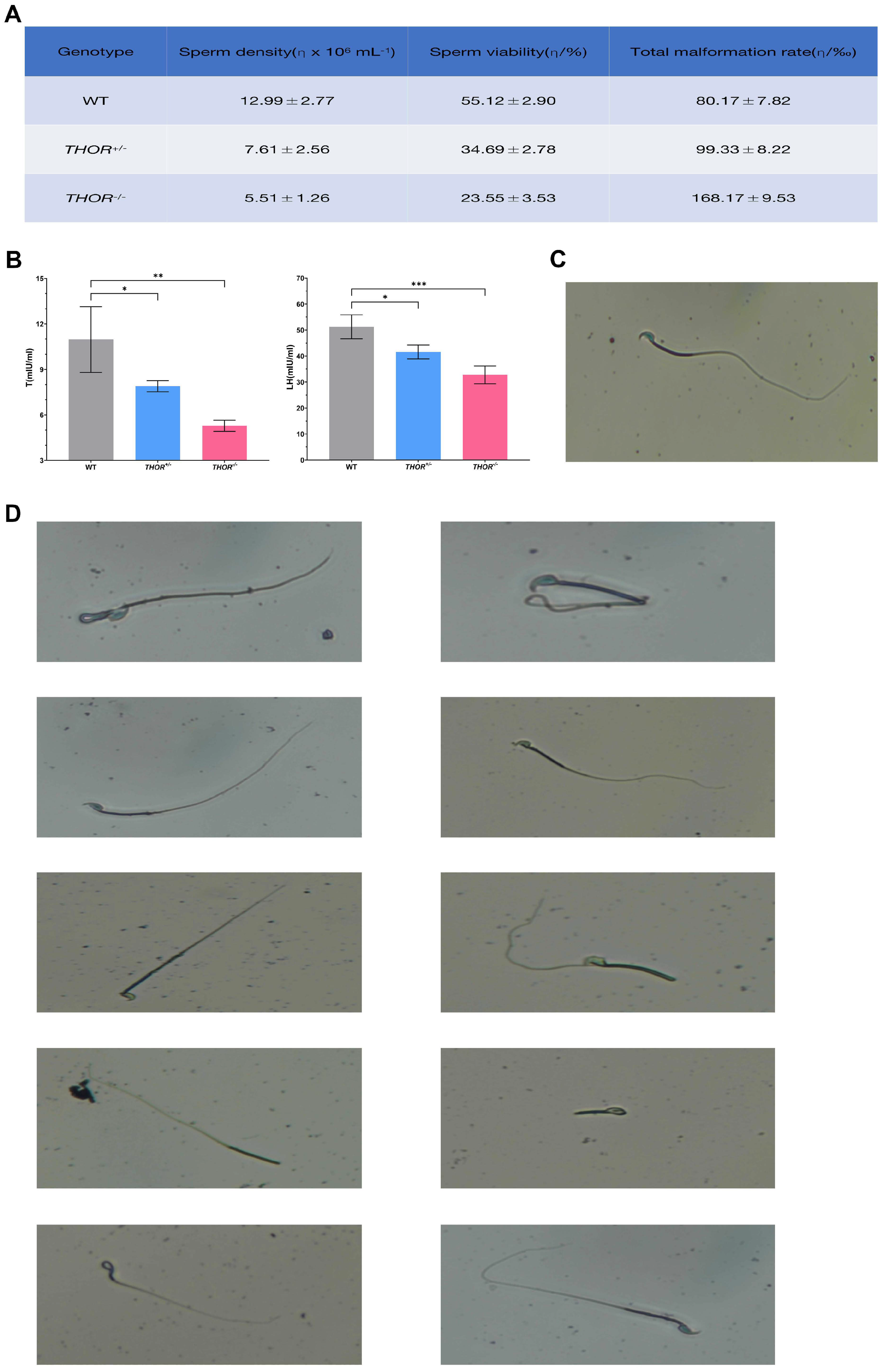
3.5. THOR Deficiency Affects Sperm Development
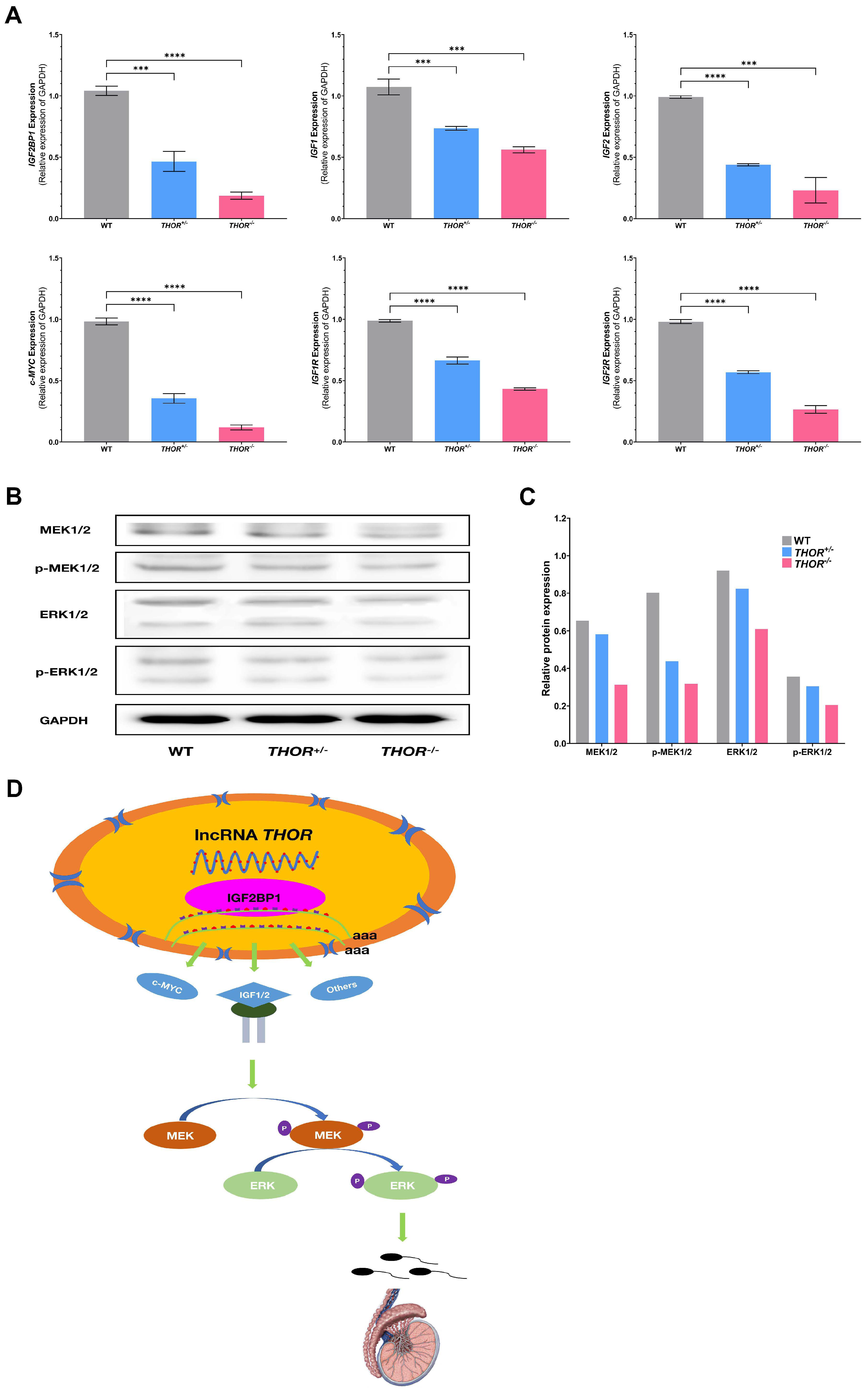
3.6. THOR Affects Sperm Development through the MEK Pathway
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ponting, C.P.; Belgard, T.G. Transcribed dark matter: Meaning or myth? Hum. Mol. Genet. 2010, 19, R162–R168. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Zhang, Z.; Krause, H.M. Long Noncoding RNAs and Repetitive Elements: Junk or Intimate Evolutionary Partners? Trends Genet. 2019, 35, 892–902. [Google Scholar] [CrossRef] [Green Version]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moraes, F.; Goes, A. A Decade of Human Genome Project Conclusion: Scientific Diffusion about Our Genome Knowledge. Biochem. Mol. Biol. Educ. 2016, 44, 215–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Yang, L.; Chen, L.L. The Diversity of Long Noncoding RNAs and Their Generation. Trends Genet. 2017, 33, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Glass, J.I.; Merryman, C.; Wise, K.S.; Hutchison, C.A.; Smith, H.O. Minimal Cells-Real and Imagined. Csh. Perspect Biol. 2017, 9, a023861. [Google Scholar] [CrossRef] [Green Version]
- Pang, Y.A.; Mao, C.B.; Liu, S.R. Encoding activities of non-coding RNAs. Theranostics 2018, 8, 2496–2507. [Google Scholar] [CrossRef]
- Uchida, S.; Dimmeler, S. Long noncoding RNAs in cardiovascular diseases. Circ. Res. 2015, 116, 737–750. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, A.M.; Chang, H.Y. Long Noncoding RNAs in Cancer Pathways. Cancer Cell 2016, 29, 452–463. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Chen, Y.; Qian, Q.; Wang, X.; Chang, Y.; Ju, S.; Xu, Y.; Zhang, C.; Qin, N.; Ding, H.; et al. Gene amplification derived a cancer-testis long noncoding RNA PCAT6 regulates cell proliferation and migration in hepatocellular carcinoma. Cancer Med. 2019, 8, 3017–3025. [Google Scholar] [CrossRef] [Green Version]
- Hosono, Y.; Niknafs, Y.S.; Prensner, J.R.; Iyer, M.K.; Dhanasekaran, S.M.; Mehra, R.; Pitchiaya, S.; Tien, J.; Escara-Wilke, J.; Poliakov, A.; et al. Oncogenic Role of THOR, a Conserved Cancer/Testis Long Non-coding RNA. Cell 2017, 171, 1559–1572.e20. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Zhang, H.; Guo, X.; Zhu, Z.; Cai, H.; Kong, X. Insulin-like growth factor 2 mRNA-binding protein 1 (IGF2BP1) in cancer. J. Hematol. Oncol. 2018, 11, 88. [Google Scholar] [CrossRef] [Green Version]
- Shang, Y.M. LncRNA THOR acts as a retinoblastoma promoter through enhancing the combination of c-myc mRNA and IGF2BP1 protein. Biomed. Pharm. 2018, 106, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.J.; Wu, G.Y.; Lin, C.; Guo, H.L.; Xu, J.J.; Zhao, T.L. IGF2BP1 over-expression in skin squamous cell carcinoma cells is essential for cell growth. Biochem. Biophys. Res. Commun. 2018, 501, 731–738. [Google Scholar] [CrossRef]
- Golyshev, V.M.; Abramova, T.V.; Pyshnyi, D.V.; Lomzov, A.A. Structure and Hybridization Properties of Glycine Morpholine Oligomers in Complexes with DNA and RNA: Experimental and Molecular Dynamics Studies. J. Phys. Chem. B 2019, 123, 10571–10581. [Google Scholar] [CrossRef]
- Cui, H.M. Loss of imprinting of IGF2 as an epigenetic marker for the risk of human cancer. Dis. Markers 2007, 23, 105–112. [Google Scholar] [CrossRef] [Green Version]
- DeChiara, T.M.; Robertson, E.J.; Efstratiadis, A. Parental imprinting of the mouse insulin-like growth factor II gene. Cell 1991, 64, 849–859. [Google Scholar] [CrossRef]
- Rainier, S.; Johnson, L.A.; Dobry, C.J.; Ping, A.J.; Grundy, P.E.; Feinberg, A.P. Relaxation of imprinted genes in human cancer. Nature 1993, 362, 747–749. [Google Scholar] [CrossRef] [PubMed]
- Ni, W.; Pan, C.; Pan, Q.; Fei, Q.; Huang, X.; Zhang, C. Methylation levels of IGF2 and KCNQ1 in spermatozoa from infertile men are associated with sperm DNA damage. Andrologia 2019, 51, e13239. [Google Scholar] [CrossRef] [PubMed]
- Neirijnck, Y.; Kuhne, F.; Mayere, C.; Pavlova, E.; Sararols, P.; Foti, M.; Atanassova, N.; Nef, S. Tumor Suppressor PTEN Regulates Negatively Sertoli Cell Proliferation, Testis Size, and Sperm Production In Vivo. Endocrinology 2019, 160, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, M.; Li, J.; Wan, X.; Huang, Y.; Wang, C.; Zhang, P.; Xu, Y.; Kong, Z.; Lu, Y.; et al. Comprehensive Characterization of Androgen-Responsive lncRNAs Mediated Regulatory Network in Hormone-Related Cancers. Dis. Markers 2020, 2020, 8884450. [Google Scholar] [CrossRef]
- Nickols, N.G.; Nazarian, R.; Zhao, S.G.; Tan, V.; Uzunangelov, V.; Xia, Z.; Baertsch, R.; Neeman, E.; Gao, A.C.; Thomas, G.V.; et al. MEK-ERK signaling is a therapeutic target in metastatic castration resistant prostate cancer. Prostate Cancer Prostatic Dis. 2019, 22, 531–538. [Google Scholar] [CrossRef]
- Pui, H.P.; Saga, Y. Gonocytes-to-spermatogonia transition initiates prior to birth in murine testes and it requires FGF signaling. Mech. Dev. 2017, 144, 125–139. [Google Scholar] [CrossRef]
- Bley, N.; Schott, A.; Muller, S.; Misiak, D.; Lederer, M.; Fuchs, T.; Assmann, C.; Glass, M.; Ihling, C.; Sinz, A.; et al. IGF2BP1 is a targetable SRC/MAPK-dependent driver of invasive growth in ovarian cancer. RNA Biol. 2021, 18, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Yang, X.; Wang, L. Effect of lncrna thor on proliferation and migration of colon cancer cells. Oncol Lett 2019, 18, 2518–2522. [Google Scholar] [PubMed] [Green Version]
- Wang, S.S.; Lv, Y.; Xu, X.C.; Zuo, Y.; Song, Y.; Wu, G.P.; Lu, P.H.; Zhang, Z.Q.; Chen, M.B. Triptonide inhibits human nasopharyngeal carcinoma cell growth via disrupting Lnc-RNA THOR-IGF2BP1 signaling. Cancer Lett. 2019, 443, 13–24. [Google Scholar] [CrossRef]
- Liu, H.; Xu, Y.; Yao, B.; Sui, T.; Lai, L.; Li, Z. A novel N6-methyladenosine (m6A)-dependent fate decision for the lncRNA THOR. Cell Death Dis. 2020, 11, 613. [Google Scholar] [CrossRef]
- El Refaey, M.; Xu, L.; Gao, Y.; Canan, B.D.; Adesanya, T.M.A.; Warner, S.C.; Akagi, K.; Symer, D.E.; Mohler, P.J.; Ma, J.; et al. In Vivo Genome Editing Restores Dystrophin Expression and Cardiac Function in Dystrophic Mice. Circ. Res. 2017, 121, 923–929. [Google Scholar] [CrossRef]
- Shen, B.; Zhang, J.; Wu, H.; Wang, J.; Ma, K.; Li, Z.; Zhang, X.; Zhang, P.; Huang, X. Generation of gene-modified mice via Cas9/RNA-mediated gene targeting. Cell Res. 2013, 23, 720–723. [Google Scholar] [CrossRef]
- Sui, T.; Yuan, L.; Liu, H.; Chen, M.; Deng, J.; Wang, Y.; Li, Z.; Lai, L. CRISPR/Cas9-mediated mutation of PHEX in rabbit recapitulates human X-linked hypophosphatemia (XLH). Hum. Mol. Genet. 2016, 25, 2661–2671. [Google Scholar]
- Xu, L.; Park, K.H.; Zhao, L.; Xu, J.; El Refaey, M.; Gao, Y.; Zhu, H.; Ma, J.; Han, R. CRISPR-mediated Genome Editing Restores Dystrophin Expression and Function in mdx Mice. Mol. Ther. 2016, 24, 564–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weidensdorfer, D.; Stöhr, N.; Baude, A.; Lederer, M.; Köhn, M.; Schierhorn, A.; Buchmeier, S.; Wahle, E.; Hüttelmaier, S. Control of c-myc mrna stability by igf2bp1-associated cytoplasmic rnps. Rna 2009, 15, 104–115. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.T.; Huang, H.; Huang, W.P.; Hu, W.L. LncRNA THOR promotes human renal cell carcinoma cell growth. Biochem. Biophys. Res. Commun. 2018, 501, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.W.; Xue, M.; Zhu, B.X.; Yue, C.L.; Chen, M.; Qin, H.H. microRNA-4500 inhibits human glioma cell progression by targeting IGF2BP1. Biochem. Biophys. Res. Commun. 2019, 513, 800–806. [Google Scholar] [CrossRef] [PubMed]
- Moreira, D.P.; Melo, R.M.C.; Weber, A.A.; Rizzo, E. Insulin-like growth factors 1 and 2 are associated with testicular germ cell proliferation and apoptosis during fish reproduction. Reprod. Fertil. Dev. 2020, 32, 988–998. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, J.; Gong, C.; Lan, K.; Shen, Y.; Ding, X. Development of an immunogenomic landscape for the competing endogenous RNAs network of peri-implantitis. BMC Med. Genet. 2020, 21, 208. [Google Scholar] [CrossRef]
- Chang, Y.; Wang, X.; Xu, Y.; Yang, L.; Qian, Q.; Ju, S.; Chen, Y.; Chen, S.; Qin, N.; Ma, Z.; et al. Comprehensive characterization of cancer-testis genes in testicular germ cell tumor. Cancer Med. 2019, 8, 3511–3519. [Google Scholar] [CrossRef]
- Xue, J.; Zhong, S.; Sun, B.M.; Sun, Q.F.; Hu, L.Y.; Pan, S.J. Lnc-THOR silencing inhibits human glioma cell survival by activating MAGEA6-AMPK signaling. Cell Death Dis. 2019, 10, 866. [Google Scholar] [CrossRef]
- Cheng, Z.; Lei, Z.; Yang, P.; Si, A.; Xiang, D.; Zhou, J.; Huser, N. Long non-coding RNA THOR promotes cell proliferation and metastasis in hepatocellular carcinoma. Gene 2018, 678, 129–136. [Google Scholar] [CrossRef]
- Song, H.; Wei, M.; Liu, W.; Shen, S.; Li, J.; Wang, L. Cisplatin induced apoptosis of ovarian cancer A2780s cells by activation of ERK/p53/PUMA signals. Histol. Histopathol. 2018, 33, 73–79. [Google Scholar]
- Fang, C.Y.; Wu, C.Z.; Chen, P.N.; Chang, Y.C.; Chuang, C.Y.; Lai, C.T.; Yang, S.F.; Tsai, L.L. Antimetastatic potentials of salvianolic acid A on oral squamous cell carcinoma by targeting MMP-2 and the c-Raf/MEK/ERK pathway. Environ. Toxicol. 2018, 33, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Schmeisser, M.J.; Baumann, B.; Johannsen, S.; Vindedal, G.F.; Jensen, V.; Hvalby, O.C.; Sprengel, R.; Seither, J.; Maqbool, A.; Magnutzki, A.; et al. IkappaB kinase/nuclear factor kappaB-dependent insulin-like growth factor 2 (Igf2) expression regulates synapse formation and spine maturation via Igf2 receptor signaling. J. Neurosci. 2012, 32, 5688–5703. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, L.; Li, J.; Liu, J.; Wang, A.; Liu, Y.; Yu, H.; Ouyang, H.; Pang, D. Investigation of the lncRNA THOR in Mice Highlights the Importance of Noncoding RNAs in Mammalian Male Reproduction. Biomedicines 2021, 9, 859. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9080859
Zhou L, Li J, Liu J, Wang A, Liu Y, Yu H, Ouyang H, Pang D. Investigation of the lncRNA THOR in Mice Highlights the Importance of Noncoding RNAs in Mammalian Male Reproduction. Biomedicines. 2021; 9(8):859. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9080859
Chicago/Turabian StyleZhou, Lin, Jianing Li, Jinsong Liu, Anbei Wang, Ying Liu, Hao Yu, Hongsheng Ouyang, and Daxin Pang. 2021. "Investigation of the lncRNA THOR in Mice Highlights the Importance of Noncoding RNAs in Mammalian Male Reproduction" Biomedicines 9, no. 8: 859. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9080859