
Involvement of Gut Microbiota in Schizophrenia and Treatment Resistance to Antipsychotics

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sample
2.2. Sample Collection and DNA Extraction
2.3. Next-Generation Sequencing of Bacterial 16S rRNA Gene
2.4. Bioinformatic Analysis
2.5. Statistical Analysis
3. Results
3.1. Sample Characteristics
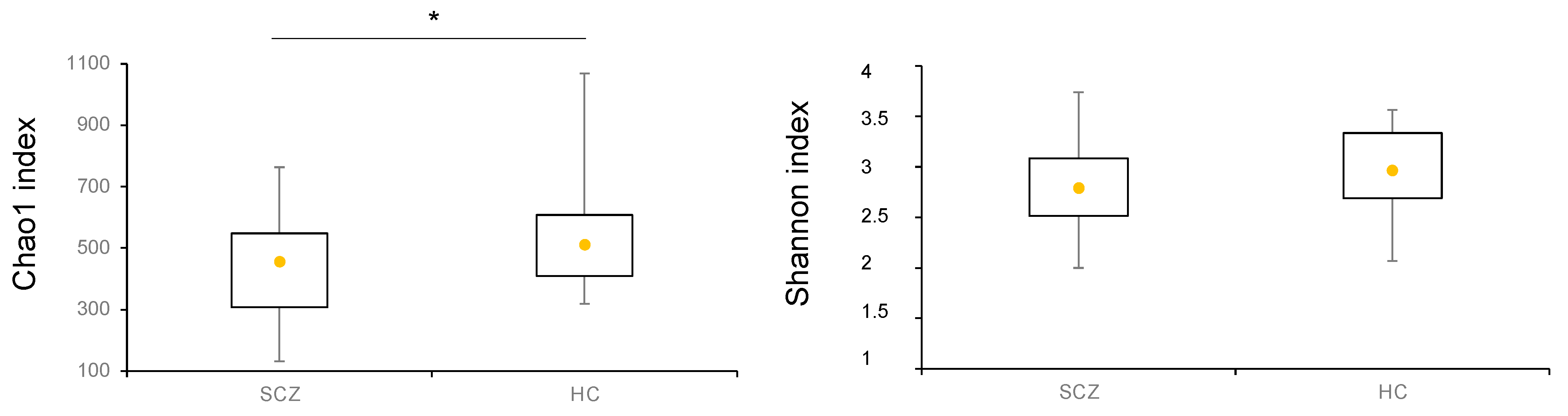
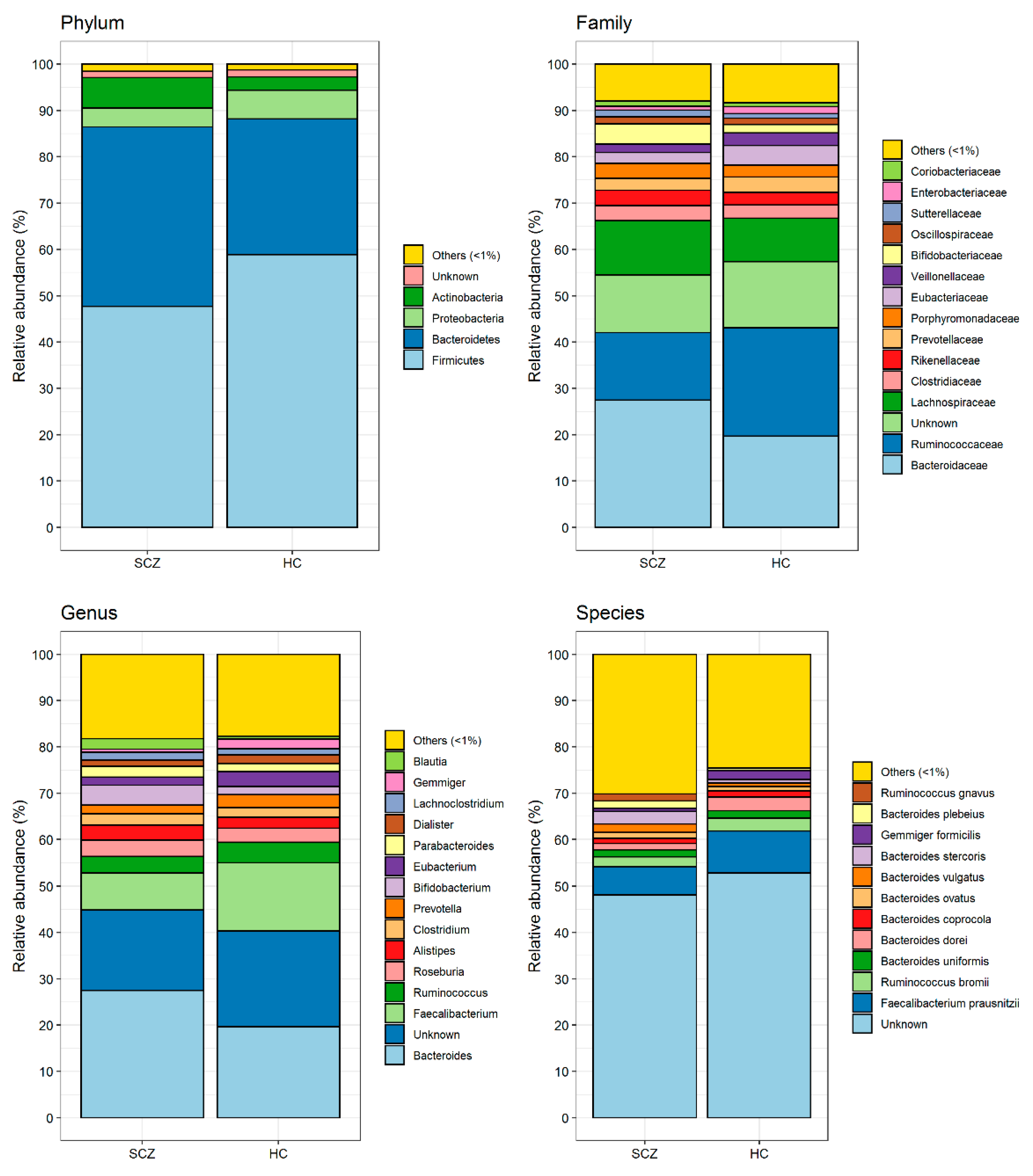
3.2. Comparison of Gut Microbiota Composition between Patients Affected by SCZ and HC
3.3. Comparison of Gut Microbiota Composition between TRS and R Patients and between TRS and HC, R and HC
3.4. Comparison of Gut Microbiota Composition between SCZ Patients Treated with Typical and Atypical Antipsychotics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Owen, M.J.; Sawa, A.; Mortensen, P.B. Schizophrenia. Lancet 2016, 388, 86–97. [Google Scholar] [CrossRef] [Green Version]
- Charlson, F.J.; Ferrari, A.J.; Santomauro, D.F.; Diminic, S.; Stockings, E.; Scott, J.G.; McGrath, J.J.; Whiteford, H.A. Global Epidemiology and Burden of Schizophrenia: Findings from the Global Burden of Disease Study 2016. Schizophr. Bull. 2018, 44, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Chant, D.; Welham, J.; McGrath, J. A Systematic Review of the Prevalence of Schizophrenia. PLoS Med. 2005, 2, e141. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Küstner, B.; Martín, C.; Pastor, L. Prevalence of Psychotic Disorders and Its Association with Methodological Issues: A Systematic Review and Meta-Analyses. PLoS ONE 2018, 13, e0195687. [Google Scholar] [CrossRef]
- Vos, T.; Abajobir, A.A.; Abate, K.H.; Abbafati, C.; Abbas, K.M.; Abd-Allah, F.; Abdulkader, R.S.; Abdulle, A.M.; Abebo, T.A.; Abera, S.F. Global, Regional, and National Incidence, Prevalence, and Years Lived with Disability for 328 Diseases and Injuries for 195 Countries, 1990–2016: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1211–1259. [Google Scholar] [CrossRef] [Green Version]
- Olfson, M.; Gerhard, T.; Huang, C.; Crystal, S.; Stroup, T.S. Premature Mortality among Adults with Schizophrenia in the United States. JAMA Psychiatry 2015, 72, 1172–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, M.J.; O’Donovan, M.C.; Thapar, A.; Craddock, N. Neurodevelopmental Hypothesis of Schizophrenia. Br. J. Psychiatry 2011, 198, 173–175. [Google Scholar] [CrossRef]
- Weinberger, D.R. The Pathogenesis of Schizophrenia: A Neurodevelopmental Theory. Neurol. Schizophr. 1986, 397–406. Available online: https://ci.nii.ac.jp/naid/10029853621/#cit (accessed on 21 July 2021).
- Murray, R.M.; Lewis, S.W. Is Schizophrenia a Neurodevelopmental Disorder? Br. Med. J. 1987, 295, 681. [Google Scholar] [CrossRef] [Green Version]
- Demjaha, A.; Lappin, J.M.; Stahl, D.; Patel, M.X.; MacCabe, J.H.; Howes, O.D.; Heslin, M.; Reininghaus, U.A.; Donoghue, K.; Lomas, B. Antipsychotic Treatment Resistance in First-Episode Psychosis: Prevalence, Subtypes and Predictors. Psychol. Med. 2017, 47, 1981–1989. [Google Scholar] [CrossRef] [Green Version]
- Lally, J.; Ajnakina, O.; Di Forti, M.; Trotta, A.; Demjaha, A.; Kolliakou, A.; Mondelli, V.; Marques, T.R.; Pariante, C.; Dazzan, P. Two Distinct Patterns of Treatment Resistance: Clinical Predictors of Treatment Resistance in First-Episode Schizophrenia Spectrum Psychoses. Psychol. Med. 2016, 46, 3231–3240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howes, O.D.; McCutcheon, R.; Agid, O.; De Bartolomeis, A.; Van Beveren, N.J.; Birnbaum, M.L.; Bloomfield, M.A.; Bressan, R.A.; Buchanan, R.W.; Carpenter, W.T. Treatment-Resistant Schizophrenia: Treatment Response and Resistance in Psychosis (TRRIP) Working Group Consensus Guidelines on Diagnosis and Terminology. Am. J. Psychiatry 2017, 174, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Lehman, A.F.; Lieberman, J.A.; Dixon, L.B.; McGlashan, T.H.; Miller, A.L.; Perkins, D.O.; Kreyenbuhl, J.; McIntyre, J.S.; Charles, S.C.; Altshuler, K. Practice Guideline for the Treatment of Partients with Schizophrenia. Am. J. Psychiatry 2004, 161, 1–54. [Google Scholar]
- Kane, J.; Honigfeld, G.; Singer, J.; Meltzer, H. Clozapine for the Treatment-Resistant Schizophrenic: A Double-Blind Comparison with Chlorpromazine. Arch. Gen. Psychiatry 1988, 45, 789–796. [Google Scholar] [CrossRef]
- Siskind, D.; Siskind, V.; Kisely, S. Clozapine Response Rates among People with Treatment-Resistant Schizophrenia: Data from a Systematic Review and Meta-Analysis. Can. J. Psychiatry 2017, 62, 772–777. [Google Scholar] [CrossRef]
- Farooq, S.; Choudry, A.; Cohen, D.; Naeem, F.; Ayub, M. Barriers to Using Clozapine in Treatment-Resistant Schizophrenia: Systematic Review. BJPsych Bull. 2019, 43, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Lieberman, J.A.; Safferman, A.Z.; Pollack, S.; Szymanski, S.; Johns, C.; Howard, A.; Kronig, M.; Bookstein, P.; Kane, J.M. Clinical Effects of Clozapine in Chronic Schizophrenia: Response to Treatment and Predictors of Outcome. Am. J. Psychiatry 1994, 151, 1744–1752. [Google Scholar]
- McNabb, C.B.; Kydd, R.; Sundram, F.; Soosay, I.; Russell, B.R. Differences in White Matter Connectivity between Treatment-Resistant and Treatment-Responsive Subtypes of Schizophrenia. Psychiatry Res. Neuroimag. 2018, 282, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, A.L.; Samanaite, R.; Mill, J.; Egerton, A.; MacCabe, J.H. Is Treatment-Resistant Schizophrenia Categorically Distinct from Treatment-Responsive Schizophrenia? A Systematic Review. BMC Psychiatry 2017, 17, 12. [Google Scholar] [CrossRef] [Green Version]
- Potkin, S.G.; Kane, J.M.; Correll, C.U.; Lindenmayer, J.-P.; Agid, O.; Marder, S.R.; Olfson, M.; Howes, O.D. The Neurobiology of Treatment-Resistant Schizophrenia: Paths to Antipsychotic Resistance and a Roadmap for Future Research. NPJ Schizophr. 2020, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Farooq, S.; Agid, O.; Foussias, G.; Remington, G. Using Treatment Response to Subtype Schizophrenia: Proposal for a New Paradigm in Classification. Schizophr. Bull. 2013, 39, 1169–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, M.; Zimmermann-Kogadeeva, M.; Wegmann, R.; Goodman, A.L. Mapping Human Microbiome Drug Metabolism by Gut Bacteria and Their Genes. Nature 2019, 570, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Pisanu, C.; Squassina, A. We Are Not Alone in Our Body: Insights into the Involvement of Microbiota in the Etiopathogenesis and Pharmacology of Mental Illness. Curr. Drug Metab. 2018, 19, 688–694. [Google Scholar] [CrossRef]
- Javdan, B.; Lopez, J.G.; Chankhamjon, P.; Lee, Y.-C.J.; Hull, R.; Wu, Q.; Wang, X.; Chatterjee, S.; Donia, M.S. Personalized Mapping of Drug Metabolism by the Human Gut Microbiome. Cell 2020, 181, 1661–1679. [Google Scholar] [CrossRef] [PubMed]
- Golofast, B.; Vales, K. The Connection between Microbiome and Schizophrenia. Neurosci. Biobehav. Rev. 2020, 108, 712–731. [Google Scholar] [CrossRef]
- Seeman, M.V. The Gut Microbiome and Antipsychotic Treatment Response. Behav. Brain Res. 2021, 396, 112886. [Google Scholar] [CrossRef]
- Liu, J.C.; Gorbovskaya, I.; Hahn, M.K.; Müller, D.J. The Gut Microbiome in Schizophrenia and the Potential Benefits of Prebiotic and Probiotic Treatment. Nutrients 2021, 13, 1152. [Google Scholar] [CrossRef]
- Zhu, F.; Guo, R.; Wang, W.; Ju, Y.; Wang, Q.; Ma, Q.; Sun, Q.; Fan, Y.; Xie, Y.; Yang, Z. Transplantation of Microbiota from Drug-Free Patients with Schizophrenia Causes Schizophrenia-like Abnormal Behaviors and Dysregulated Kynurenine Metabolism in Mice. Mol. Psychiatry 2020, 25, 2905–2918. [Google Scholar] [CrossRef]
- Bastiaanssen, T.F.; Cryan, J.F. The Microbiota-Gut-Brain Axis in Mental Health and Medication Response: Parsing Directionality and Causality. Int. J. Neuropsychopharmacol. 2021, 24, 216–220. [Google Scholar] [CrossRef]
- Foster, J.A.; Neufeld, K.-A.M. Gut–Brain Axis: How the Microbiome Influences Anxiety and Depression. Trends Neurosci. 2013, 36, 305–312. [Google Scholar] [CrossRef]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The Gut-Brain Axis: Interactions between Enteric Microbiota, Central and Enteric Nervous Systems. Ann. Gastroenterol. Q. Publ. Hell. Soc. Gastroenterol. 2015, 28, 203. [Google Scholar]
- Stavropoulou, E.; Bezirtzoglou, E. Human Microbiota in Aging and Infection: A Review. Crit. Rev. Food Sci. Nutr. 2019, 59, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Kelsen, J.R.; Wu, G.D. The Gut Microbiota, Environment and Diseases of Modern Society. Gut Microbes 2012, 3, 374–382. [Google Scholar] [CrossRef]
- Cowan, C.S.; Dinan, T.G.; Cryan, J.F. Annual Research Review: Critical Windows–the Microbiota–Gut–Brain Axis in Neurocognitive Development. J. Child Psychol. Psychiatry 2020, 61, 353–371. [Google Scholar] [CrossRef]
- Manchia, M.; Paribello, P.; Arzedi, C.; Bocchetta, A.; Caria, P.; Cocco, C.; Congiu, D.; Cossu, E.; Dettori, T.; Frau, D.V.; et al. A Multidisciplinary Approach to Mental Illness: Do Inflammation, Telomere Length and Microbiota Form a Loop? A Protocol for a Cross-Sectional Study on the Complex Relationship between Inflammation, Telomere Length, Gut Microbiota and Psychiatric Disorders. BMJ Open 2020, 10, e032513. [Google Scholar] [CrossRef] [Green Version]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-IV-TR; American Psychiatric Association: Washington, DC, USA, 2000. [Google Scholar]
- Picchianti-Diamanti, A.; Panebianco, C.; Salemi, S.; Sorgi, M.L.; Di Rosa, R.; Tropea, A.; Sgrulletti, M.; Salerno, G.; Terracciano, F.; D’amelio, R. Analysis of Gut Microbiota in Rheumatoid Arthritis Patients: Disease-Related Dysbiosis and Modifications Induced by Etanercept. Int. J. Mol. Sci. 2018, 19, 2938. [Google Scholar] [CrossRef] [Green Version]
- Fontana, A.; Manchia, M.; Panebianco, C.; Paribello, P.; Arzedi, C.; Cossu, E.; Garzilli, M.; Montis, M.A.; Mura, A.; Pisanu, C. Exploring the Role of Gut Microbiota in Major Depressive Disorder and in Treatment Resistance to Antidepressants. Biomedicines 2020, 8, 311. [Google Scholar] [CrossRef]
- Dettling, M.; Bühlmann, P. Finding Predictive Gene Groups from Microarray Data. J. Multivar. Anal. 2004, 90, 106–131. [Google Scholar] [CrossRef]
- Team, R. Core. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Kelly, J.R.; Minuto, C.; Cryan, J.F.; Clarke, G.; Dinan, T.G. The Role of the Gut Microbiome in the Development of Schizophrenia. Schizophr. Res. 2020. [Google Scholar] [CrossRef]
- Li, S.; Song, J.; Ke, P.; Kong, L.; Lei, B.; Zhou, J.; Huang, Y.; Li, H.; Li, G.; Chen, J.; et al. The Gut Microbiome Is Associated with Brain Structure and Function in Schizophrenia. Sci. Rep. 2021, 11, 9743. [Google Scholar] [CrossRef]
- Li, J.; Ma, Y.; Bao, Z.; Gui, X.; Li, A.N.; Yang, Z.; Li, M.D. Clostridiales Are Predominant Microbes That Mediate Psychiatric Disorders. J. Psychiatr. Res. 2020, 130, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Kosciolek, T.; Maldonado, Y.; Daly, R.E.; Martin, A.S.; McDonald, D.; Knight, R.; Jeste, D.V. Differences in Gut Microbiome Composition between Persons with Chronic Schizophrenia and Healthy Comparison Subjects. Schizophr. Res. 2019, 204, 23–29. [Google Scholar] [CrossRef]
- Bojović, K.; Ignjatović, Ð.-D.I.; Soković Bajić, S.; Vojnović Milutinović, D.; Tomić, M.; Golić, N.; Tolinački, M. Gut Microbiota Dysbiosis Associated With Altered Production of Short Chain Fatty Acids in Children With Neurodevelopmental Disorders. Front. Cell Infect. Microbiol. 2020, 10, 223. [Google Scholar] [CrossRef]
- Naudé, P.J.W.; Claassen-Weitz, S.; Gardner-Lubbe, S.; Botha, G.; Kaba, M.; Zar, H.J.; Nicol, M.P.; Stein, D.J. Association of Maternal Prenatal Psychological Stressors and Distress with Maternal and Early Infant Faecal Bacterial Profile. Acta Neuropsychiatr. 2020, 32, 32–42. [Google Scholar] [CrossRef]
- Nakajima, M.; Arimatsu, K.; Kato, T.; Matsuda, Y.; Minagawa, T.; Takahashi, N.; Ohno, H.; Yamazaki, K. Oral Administration of P. gingivalis Induces Dysbiosis of Gut Microbiota and Impaired Barrier Function Leading to Dissemination of Enterobacteria to the Liver. PLoS ONE 2015, 10, e0134234. [Google Scholar] [CrossRef] [Green Version]
- Cook, R.R.; Fulcher, J.A.; Tobin, N.H.; Li, F.; Lee, D.J.; Woodward, C.; Javanbakht, M.; Brookmeyer, R.; Shoptaw, S.; Bolan, R.; et al. Alterations to the Gastrointestinal Microbiome Associated with Methamphetamine Use among Young Men Who Have Sex with Men. Sci. Rep. 2019, 9, 14840. [Google Scholar] [CrossRef] [PubMed]
- Müller, N.; Weidinger, E.; Leitner, B.; Schwarz, M.J. The Role of Inflammation in Schizophrenia. Front. Neurosci. 2015, 9, 372. [Google Scholar] [CrossRef] [Green Version]
- Petrov, V.A.; Saltykova, I.V.; Zhukova, I.A.; Alifirova, V.M.; Zhukova, N.G.; Dorofeeva, Y.B.; Tyakht, A.V.; Kovarsky, B.A.; Alekseev, D.G.; Kostryukova, E.S.; et al. Analysis of Gut Microbiota in Patients with Parkinson’s Disease. Bull. Exp. Biol. Med. 2017, 162, 734–737. [Google Scholar] [CrossRef]
- Mandić, A.D.; Woting, A.; Jaenicke, T.; Sander, A.; Sabrowski, W.; Rolle-Kampcyk, U.; von Bergen, M.; Blaut, M. Clostridium Ramosum Regulates Enterochromaffin Cell Development and Serotonin Release. Sci. Rep. 2019, 9, 1177. [Google Scholar] [CrossRef] [Green Version]
- Rogers, G.B.; Keating, D.J.; Young, R.L.; Wong, M.-L.; Licinio, J.; Wesselingh, S. From Gut Dysbiosis to Altered Brain Function and Mental Illness: Mechanisms and Pathways. Mol. Psychiatry 2016, 21, 738–748. [Google Scholar] [CrossRef] [Green Version]
- Sani, G.; Manchia, M.; Simonetti, A.; Janiri, D.; Paribello, P.; Pinna, F.; Carpiniello, B. The Role of Gut Microbiota in the High-Risk Construct of Severe Mental Disorders: A Mini Review. Front. Psychiatry. 2020, 11, 585769. [Google Scholar] [CrossRef] [PubMed]
- Sandstrom, A.; Sahiti, Q.; Pavlova, B.; Uher, R. Offspring of Parents with Schizophrenia, Bipolar Disorder, and Depression: A Review of Familial High-Risk and Molecular Genetics Studies. Psychiatr. Genet. 2019, 29, 160–169. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Category | TRS (N = 18) | R (N = 20) | HC (N = 20) | p-Value |
|---|---|---|---|---|---|
| Age (years) | Median [IQR] | 44.0 [41.6–49.8] | 50.0 [40.3–60.9] | 37.7 [30.6–58.0] | 0.277 * |
| Gender—N(%) | Males | 16 (88.9) | 18 (90.0) | 13 (65.0) | 0.102 # |
| Females | 2 (11.1) | 2 (10.0) | 7 (35.0) | ||
| BMI (Kg/m2) | Median [IQR] | 27.3 [25.5–29.2] | 26.9 [25.5–28.5] | 22.7 [21.2–23.8] | <0.001 * |
| Family history for mental disorders—N(%) | No | 9 (50.0) | 11 (55.0) | 11 (68.8) | 0.542 # |
| Yes | 9 (50.0) | 9 (45.0) | 5 (31.2) | ||
| Diet—N(%) | Mediterranean only | 10 (58.8) | 13 (65.0) | 18 (90.0) | 0.445 # |
| Carbohydrates only | 1 (5.9) | 0 (0.0) | 0 (0.0) | ||
| Vegetarian/Vegan only | 1 (5.9) | 0 (0.0) | 0 (0.0) | ||
| Mediterran + iperproteic | 1 (5.9) | 2 (10.0) | 0 (0.0) | ||
| Mediterran + ipercaloric | 0 (0.0) | 0 (0.0) | 1 (5.0) | ||
| Mediterran + carbohydrates | 2 (11.8) | 1 (5.0) | 1 (5.0) | ||
| Mediterran + iperproteic + carbohydrates | 1 (5.9) | 1 (5.0) | 0 (0.0) | ||
| Mediterran + ipercaloric + carbohydrates | 0 (0.0) | 1 (5.0) | 0 (0.0) | ||
| Iperproteic + carbohydrates | 1 (5.9) | 2 (10.0) | 0 (0.0) | ||
| Smoking habits—N(%) | Non-smoker | 5 (27.8) | 4 (20.0) | 13 (65.0) | 0.021 # |
| Smoker | 11 (61.1) | 11 (55.0) | 4 (20.0) | ||
| Ex-smoker | 2 (11.1) | 5 (25.0) | 3 (15.0) | ||
| Drink habits—N(%) | None | 12 (66.7) | 13 (65.0) | 3 (15.8) | 0.003 # |
| One occasional drink | 6 (33.3) | 6 (30.0) | 12 (63.2) | ||
| 1-2 drinks per day | 0 (0.0) | 1 (5.0) | 4 (21.1) | ||
| Physical activity—N(%) | No | 11 (61.1) | 14 (70.0) | 5 (25.0) | 0.012 # |
| Yes | 7 (38.9) | 6 (30.0) | 15 (75.0) | ||
| Cardiometabolic comorbidities—N(%) | No | 11 (61.1) | 13 (65.0) | 16 (80.0) | 0.458 # |
| Yes | 7 (38.9) | 7 (35.0) | 4 (20.0) | ||
| Age at onset (years) | Median [IQR] | 23.0 [20.0–28.0] | 24.0 [21.8–30.0] | NA | 0.318 § |
| Disease duration (years) | Median [IQR] | 19.6 [14.7–24.2] | 19.2 [10.4–32.7] | NA | 0.770 § |
| History of suicide attempt—N(%) | No | 13 (72.2) | 17 (85.0) | NA | 0.438 # |
| Yes | 5 (27.8) | 3 (15.0) | |||
| Length of treatment with APs (months) | Median [IQR] | 66.0 [36.0–165.0] | 66.0 [34.0–87.0] | NA | 0.348 § |
| Treatment at sample collection—N(%) | Typical APs (first generation) | 3 (16.7) | 4 (20.0) | NA | 0.261 # |
| Atypical APs (second generation) | 15 (83.3) | 13 (65.0) | |||
| Aripiprazole (third generation) | 0 (0.0) | 3 (15.0) | |||
| Mood stabilizers—N(%) | No | 11 (61.1) | 16 (80.0) | NA | 0.288 # |
| Yes | 7 (38.9) | 4 (20.0) | |||
| Antidepressant—N(%) | No | 13 (72.2) | 14 (70.0) | NA | 1.000 # |
| Yes | 5 (27.8) | 6 (30.0) | |||
| Any concomitant drugs—N(%) | No | 10 (55.6) | 11 (55.0) | NA | 1.000 # |
| Yes | 8 (44.4) | 9 (45.0) |
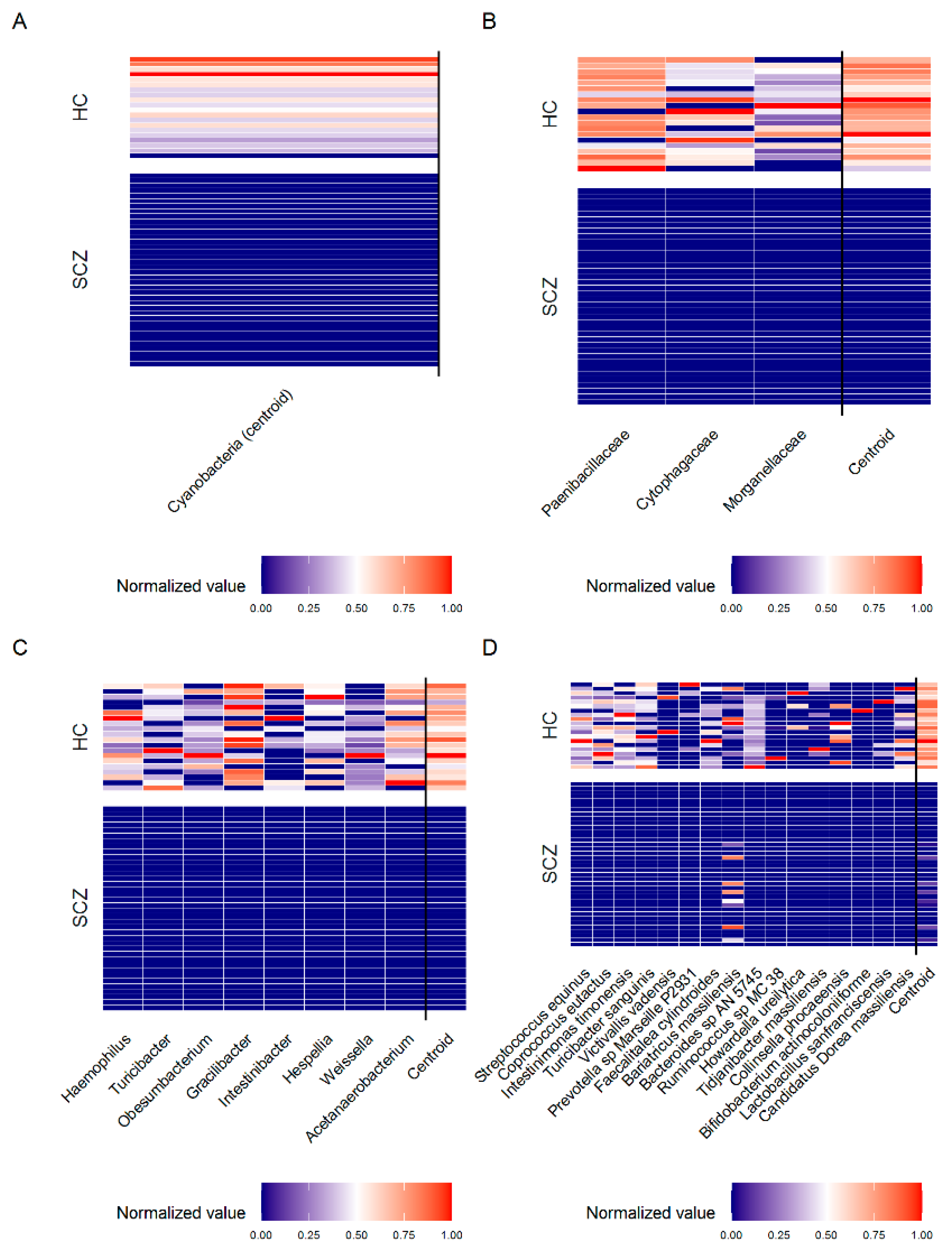
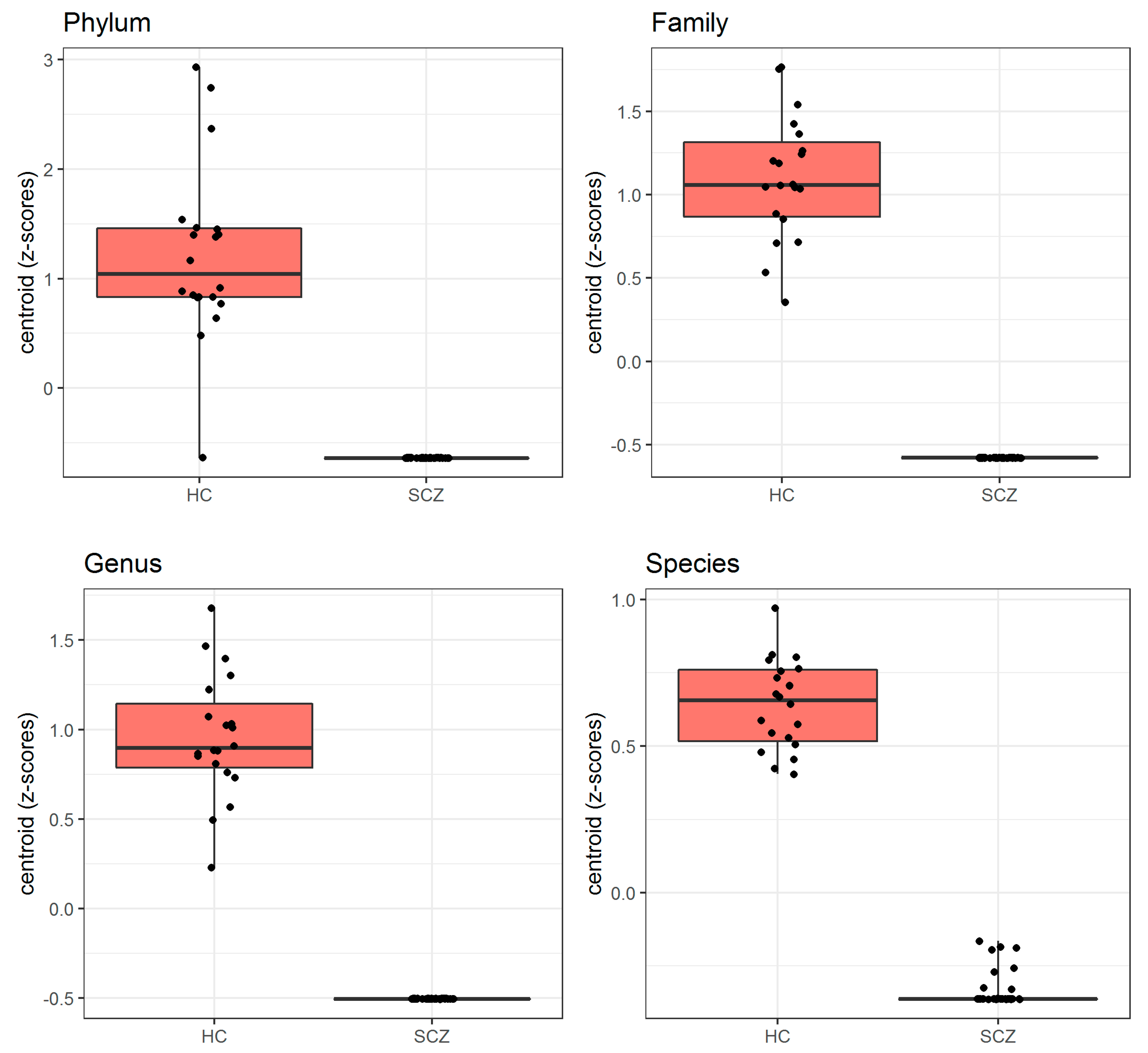
| Taxa Level | Bacteria Selected by PELORA | Quantity | Statistics | SCZ (N = 38) | HC (N = 20) | p-Value |
|---|---|---|---|---|---|---|
| Phylum | Cyanobacteria [Cluster centroid] | Relative abundance (%) | Median [IQR] | Absent | 0.004 [0.002–0.009] | <0.001 § |
| Z-score (means) | Mean ± SD | −0.638 ± 0.000 | 1.211 ± 0.802 | |||
| Family | Paenibacillaceae | Relative abundance (%) | Median [IQR] | Absent | 0.011 [0.006–0.014] | <0.001 § |
| Z-score ° | Mean ± SD | 1.236 ± 0.739 | ||||
| Cytophagaceae | Relative abundance (%) | Median [IQR] | Absent | 0.002 [0.001–0.006] | <0.001 § | |
| Z-score ° | Mean ± SD | 1.069 ± 1.079 | ||||
| Morganellaceae | Relative abundance (%) | Median [IQR] | Absent | 0.001 [0.000–0.004] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.998 ± 1.183 | ||||
| Cluster centroid | Z-score (means) | Mean ± SD | −0.579 ± 0.000 | 1.101 ± 0.371 | <0.001 # | |
| Genus | Acetanaerobacterium | Relative abundance (%) | Median [IQR] | Absent | 0.004 [0.001–0.015] | <0.001 § |
| Z-score ° | Mean ± SD | 1.124 ± 0.985 | ||||
| Haemophilus | Relative abundance (%) | Median [IQR] | Absent | 0.003 [0.001–0.018] | <0.001 § | |
| Z-score ° | Mean ± SD | 1.041 ± 1.122 | ||||
| Turicibacter | Relative abundance (%) | Median [IQR] | Absent | 0.002 [0.000–0.003] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.971 ± 1.219 | ||||
| Obesumbacterium | Relative abundance (%) | Median [IQR] | Absent | 0.002 [0.000–0.005] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.944 ± 1.253 | ||||
| Gracilibacter | Relative abundance (%) | Median [IQR] | Absent | 0.010 [0.001–0.032] | <0.001 § | |
| Z-score ° | Mean ± SD | 1.064 ± 1.087 | ||||
| Intestinibacter | Relative abundance (%) | Median [IQR] | Absent | 0.000 [0.000–0.002] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.687 ± 1.498 | ||||
| Hespellia | Relative abundance (%) | Median [IQR] | Absent | 0.006 [0.000–0.017] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.974 ± 1.214 | ||||
| Weissella | Relative abundance (%) | Median [IQR] | Absent | 0.001 [0.000–0.001] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.872 ± 1.334 | ||||
| Cluster centroid | Z-score (means) | Mean ± SD | −0.505 ± 0.000 | 0.959 ± 0.343 | <0.001 # | |
| Species | Streptococcus equinus | Relative abundance (%) | Median [IQR] | Absent | 0.003 [0.002–0.012] | <0.001 § |
| Z-score ° | Mean ± SD | 1.112 ± 1.007 | ||||
| Coprococcus eutactus | Relative abundance (%) | Median [IQR] | Absent | 0.002 [0.000–0.008] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.993 ± 1.190 | ||||
| Turicibacter sanguinis | Relative abundance (%) | Median [IQR] | Absent | 0.002 [0.000–0.003] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.965 ± 1.226 | ||||
| Victivallis vadensis | Relative abundance (%) | Median [IQR] | Absent | 0.000 [0.000–0.000] | 0.049 § | |
| Z-score ° | Mean ± SD | 0.356 ± 1.672 | ||||
| Bariatricus massiliensis | Relative abundance (%) | Median [IQR] | 0.000 [0.000-0.000] | 0.000 [0.000–0.001] | 0.146 § | |
| Z-score ° | Mean ± SD | −0.087 ± 0.933 | 0.166 ± 1.123 | |||
| Prevotella sp. Marseille P2931 | Relative abundance (%) | Median [IQR] | Absent | 0.000 [0.000–0.000] | 0.005 § | |
| Z-score ° | Mean ± SD | 0.430 ± 1.644 | ||||
| Faecalitalea cylindroides | Relative abundance (%) | Median [IQR] | Absent | 0.001 [0.000–0.002] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.821 ± 1.385 | ||||
| Intestinimonas timonensis | Relative abundance (%) | Median [IQR] | Absent | 0.001 [0.001–0.006] | <0.001 § | |
| Z-score ° | Mean ± SD | 1.052 ± 1.106 | ||||
| Bacteroides sp. AN 5745 | Relative abundance (%) | Median [IQR] | Absent | 0.003 [0.001–0.005] | <0.001 § | |
| Z-score ° | Mean ± SD | 1.054 ± 1.103 | ||||
| Collinsella phocaeensis | Relative abundance (%) | Median [IQR] | Absent | 0.000 [0.000–0.000] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.626 ± 1.540 | ||||
| Candidatus Dorea massiliensis | Relative abundance (%) | Median [IQR] | Absent | 0.001 [0.000–0.004] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.857 ± 1.349 | ||||
| Ruminococcus sp. MC 38 | Relative abundance (%) | Median [IQR] | Absent | 0.000 [0.000–0.000] | 0.049 § | |
| Z-score ° | Mean ± SD | 0.293 ± 1.692 | ||||
| Bifidobacterium actinocoloniiforme | Relative abundance (%) | Median [IQR] | Absent | 0.000 [0.000, 0.000] | 0.168 § | |
| Z-score ° | Mean ± SD | 0.249 ± 1.703 | ||||
| Tidjanibacter massiliensis | Relative abundance (%) | Median [IQR] | Absent | 0.000 [0.000–0.000] | 0.001 § | |
| Z-score ° | Mean ± SD | 0.478 ± 1.623 | ||||
| Howardella ureilytica | Relative abundance (%) | Median [IQR] | Absent | 0.000 [0.000–0.000] | 0.015 § | |
| Z-score ° | Mean ± SD | 0.427 ± 1.645 | ||||
| Lactobacillus sanfranciscensis | Relative abundance (%) | Median [IQR] | Absent | 0.000 [0.000–0.000] | 0.015 § | |
| Z-score ° | Mean ±SD | 0.389 ± 1.660 | ||||
| Cluster centroid | Z-score (means) | Mean ± SD | −0.338 ± 0.058 | 0.642 ± 0.152 | <0.001 # |
| Taxa Level | Bacteria Selected by PELORA | Quantity | Statistics | TRS (N = 18) | R (N = 20) | p-Value |
|---|---|---|---|---|---|---|
| Phylum | Candidatus Saccharibacteria | Relative abundance (%) | Median [IQR] | 0.006 [0.005-0.012] | Absent | <0.001 § |
| Z-score ° | Mean ± SD | 0.940 ± 0.632 | ||||
| Tenericutes | Relative abundance (%) | Median [IQR] | 0.003 [0.001–0.005] | Absent | <0.001 § | |
| Z-score ° | Mean ± SD | 0.686 ± 1.109 | ||||
| Cluster centroid | Z-score (means) | Mean ± SD | 0.813 ± 0.463 | −0.732 ± 0.000 | <0.001 # | |
| Family | Flavobacteriaceae | Relative abundance (%) | Median [IQR] | Absent | 0.005 [0.003–0.009] | <0.001 § |
| Z-score ° | Mean ± SD | 0.830 ± 0.646 | ||||
| Enterococcaceae | Relative abundance (%) | Median [IQR] | Absent | 0.003 [0.001–0.010] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.692 ± 0.940 | ||||
| Cluster centroid | Z-score (means) | Mean ± SD | −0.845 ± 0.000 | 0.761 ± 0.512 | <0.001 # | |
| Genus | Actinomyces | Relative abundance (%) | Median [IQR] | 0.005 [0.002–0.016] | Absent | <0.001 § |
| Z-score ° | Mean ± SD | 0.906 ± 0.724 | ||||
| Porphyromonas | Relative abundance (%) | Median [IQR] | 0.003 [0.001–0.008] | Absent | <0.001 § | |
| Z-score ° | Mean ± SD | 0.772 ± 0.988 | ||||
| Cluster centroid | Z-score (means) | Mean ± SD | 0.839 ± 0.483 | −0.755 ± 0.000 | <0.001 # | |
| Species | Flintibacter butyricus [cluster centroid] | Relative abundance (%) | Median [IQR] | Absent | 0.017 [0.008–0.031] | <0.001 § |
| Z-score ° | Mean ± SD | −0.989 ± 0.000 | 0.890 ± 0.430 |
| Taxa Level | Bacteria Selected by PELORA | Quantity | Statistics | R (N = 20) | HC (N = 20) | p-Value |
|---|---|---|---|---|---|---|
| Phylum | Candidatus Saccharibacteria | Relative abundance (%) | Median [IQR] | Absent | 0.006 [0.004–0.014] | <0.001 § |
| Z-score ° | Mean ± SD | 0.869 ± 0.681 | ||||
| Cyanobacteria | Relative abundance (%) | Median [IQR] | Absent | 0.004 [0.002–0.009] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.848 ± 0.735 | ||||
| Tenericutes | Relative abundance (%) | Median [IQR] | Absent | 0.024 [0.003–0.111] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.825 ± 0.787 | ||||
| Cluster centroid | Z-score (means) | Mean ± SD | −0.847 ± 0.000 | 0.847 ± 0.415 | <0.001 # | |
| Family | Flavobacteriaceae | Relative abundance (%) | Median [IQR] | 0.005 [0.003–0.009] | Absent | <0.001 § |
| Z-score ° | Mean ± SD | 0.880 ± 0.650 | ||||
| Desulfobacteraceae | Relative abundance (%) | Median [IQR] | 0.000 [0.000-0.003] | Absent | <0.001 § | |
| Z-score ° | Mean ± SD | 0.462 ± 1.266 | ||||
| Cluster centroid | Z-score (means) | Mean ± SD | 0.671 ± 0.675 | −0.671 ± 0.000 | <0.001 # | |
| Genus | Fenollaria | Relative abundance (%) | Median [IQR] | 0.007 [0.004–0.012] | Absent | <0.001 § |
| Z-score ° | Mean ± SD | 0.845 ± 0.740 | ||||
| Mitsuokella | Relative abundance (%) | Median [IQR] | 0.000 [0.000–0.002] | Absent | 0.002 § | |
| Z-score ° | Mean ± SD | 0.432 ± 1.288 | ||||
| Harryflintia | Relative abundance (%) | Median [IQR] | 0.001 [0.000–0.007] | Absent | <0.001 § | |
| Z-score ° | Mean ± SD | 0.573 ± 1.167 | ||||
| Mailhella | Relative abundance (%) | Median [IQR] | 0.000 [0.000–0.002] | Absent | 0.002 § | |
| Z-score ° | Mean ± SD | 0.432 ± 1.289 | ||||
| Cluster centroid | Z-score (means) | Mean ± SD | 0.570 ± 0.341 | −0.570 ± 0.000 | <0.001 # | |
| Species | Flintibacter butyricus [cluster centroid] | Relative abundance (%) | Median [IQR] | 0.017 [0.008–0.031] | Absent | <0.001 # |
| Z-score ° | Mean ± SD | 0.942 ± 0.431 | −0.942 ± 0.000 |
| Taxa Level | Bacteria Selected by PELORA | Quantity | Statistics | TRS (N = 18) | HC (N = 20) | p-Value |
|---|---|---|---|---|---|---|
| Phylum | Cyanobacteria [cluster centroid] | Relative abundance (%) | Median [IQR] | Absent | 0.004 [0.002–0.009] | <0.001 § |
| Z-score ° | Mean ± SD | −0.886 ± 0.000 | 0.798 ± 0.730 | |||
| Family | Enterococcaceae | Relative abundance (%) | Median [IQR] | Absent | 0.007 [0.002–0.012] | <0.001 § |
| Z-score ° | Mean ± SD | 0.744 ± 0.847 | ||||
| Paenibacillaceae | Relative abundance (%) | Median [IQR] | Absent | 0.011 [0.006–0.014] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.819 ± 0.677 | ||||
| Cytophagaceae | Relative abundance (%) | Median [IQR] | Absent | 0.002 [0.001–0.006] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.683 ± 0.954 | ||||
| Hafniaceae | Relative abundance (%) | Median [IQR] | Absent | 0.003 [0.001–0.010] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.660 ± 0.989 | ||||
| Pasteurellaceae | Relative abundance (%) | Median [IQR] | Absent | 0.006 [0.003–0.023] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.745 ± 0.844 | ||||
| Cluster centroid | Z-score (means) | Mean ± SD | −0.811 ± 0.000 | 0.730 ± 0.320 | <0.001 # | |
| Genus | Murimonas | Relative abundance (%) | Median [IQR] | Absent | 0.019 [0.008–0.037] | <0.001 § |
| Z-score ° | Mean ± SD | 0.849 ± 0.587 | ||||
| Haemophilus | Relative abundance (%) | Median [IQR] | Absent | 0.003 [0.001–0.018] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.662 ± 0.987 | ||||
| Peptococcus | Relative abundance (%) | Median [IQR] | Absent | 0.006 [0.000–0.010] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.615 ± 1.052 | ||||
| Weissella | Relative abundance (%) | Median [IQR] | Absent | 0.001 [0.000–0.001] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.539 ± 1.141 | ||||
| Enterobacter | Relative abundance (%) | Median [IQR] | Absent | 0.001 [0.000–0.040] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.538 ± 1.142 | ||||
| Hespellia | Relative abundance (%) | Median [IQR] | Absent | 0.006 [0.000–0.017] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.612 ± 1.056 | ||||
| Turicibacter | Relative abundance (%) | Median [IQR] | Absent | 0.002 [0.000–0.003] | <0.001 § | |
| Z-score ° | Mean ± SD | 0.610 ± 1.059 | ||||
| Fusicatenibacter | Relative abundance (%) | Median [IQR] | 0.005 [0.003–0.018] | 0.010 [0.005–0.038] | 0.334 # | |
| Z-score ° | Mean ± SD | −0.167 ± 1.062 | 0.151 ± 0.942 | |||
| Eggerthella | Relative abundance (%) | Median [IQR] | 0.013 [0.007–0.079] | 0.017 [0.010–0.043] | 0.920 # | |
| Z-score ° | Mean ± SD | 0.018 ± 1.024 | −0.016 ± 1.004 | |||
| Cluster centroid | Z-score (means) | Mean ± SD | −0.563 ± 0.125 | 0.507 ± 0.168 | <0.001 # | |
| Species | Bacteroides sp. Marseille P3108 | Relative abundance (%) | Median [IQR] | 0.005 [0.003–0.016] | Absent | <0.001 § |
| Z-score ° | Mean ± SD | 0.888 ± 0.767 | ||||
| Pseudoflavonifractor capillosus | Relative abundance (%) | Median [IQR] | 0.009 [0.002–0.028] | Absent | <0.001 § | |
| Z-score ° | Mean ± SD | 0.834 ± 0.881 | ||||
| Erysipelatoclostridium ramosum | Relative abundance (%) | Median [IQR] | 0.001 [0.000–0.019] | Absent | <0.001 § | |
| Z-score ° | Mean ± SD | 0.613 ± 1.192 | ||||
| Papillibacter cinnamivorans | Relative abundance (%) | Median [IQR] | 0.006 [0.000–0.018] | Absent | <0.001 § | |
| Z-score ° | Mean ± SD | 0.735 ± 1.044 | ||||
| Clostridium sp. BPY5 | Relative abundance (%) | Median [IQR] | 0.007 [0.001–0.016] | Absent | <0.001 § | |
| Z-score ° | Mean ± SD | 0.784 ± 0.969 | ||||
| Cluster centroid | Z-score (means) | Mean ± SD | 0.771 ± 0.204 | −0.694 ± 0.000 | <0.001 # |
| Taxa Level | Bacteria Selected by PELORA | Quantity | Statistics | T SCZ (N = 7) | AT SCZ (N = 31) | p-Value |
|---|---|---|---|---|---|---|
| Phylum | Fusobacteria | Relative abundance (%) | Median [IQR] | 0.012 [0.001-0.039] | 0.000 [0.000–0.001] | 0.001 # |
| Z-score ° | Mean ± SD | 1.103 ± 1.223 | −0.249 ± 0.766 | |||
| Tenericutes * | Relative abundance (%) | Median [IQR] | 0.000 [0.000–0.000] | 0.000 [0.000–0.003] | 0.153 # | |
| Z-score ° | Mean ± SD | −0.491 ± 0.335 | 0.111 ± 1.069 | |||
| Bacteroidetes * | Relative abundance (%) | Median [IQR] | 42.397 [26.873–44.399] | 37.181 [28.697–49.898] | 0.543 # | |
| Z-score ° | Mean ± SD | −0.211 ± 0.951 | 0.048 ± 1.020 | |||
| Cluster centroid | Z-score (means) | Mean ± SD | 0.602 ± 0.314 | –0.136 ± 0.434 | <0.001 # | |
| Family | Fusobacteriaceae | Relative abundance (%) | Median [IQR] | 0.012 [0.000–0.039] | 0.000 [0.000–0.001] | 0.001 # |
| Z-score ° | Mean ± SD | 1.036 ± 1.351 | −0.234 ± 0.749 | |||
| Streptomycetaceae | Relative abundance (%) | Median [IQR] | 0.011 [0.009–0.015] | 0.008 [0.004–0.018] | 0.438 # | |
| Z-score ° | Mean ± SD | 0.269 ± 0.876 | −0.061 ± 1.029 | |||
| Helicobacteraceae | Relative abundance (%) | Median [IQR] | 0.001 [0.000–0.094] | 0.000 [0.000–0.002] | 0.085 # | |
| Z-score ° | Mean ± SD | 0.588 ± 1.609 | −0.133 ± 0.785 | |||
| Clostridiales Family XII. Incertae Sedis * | Relative abundance (%) | Median [IQR] | Absent | 0.000 [0.000–0.001] | 0.070 § | |
| Z-score ° | Mean ± SD | 0.125 ± 1.071 | ||||
| Bacillaceae | Relative abundance (%) | Median [IQR] | 0.012 [0.011–0.017] | 0.007 [0.005–0.013] | 0.122 # | |
| Z-score ° | Mean ± SD | 0.530 ± 0.798 | −0.120 ± 1.013 | |||
| Oxalobacteraceae | Relative abundance (%) | Median [IQR] | 0.008 [0.002–0.020] | 0.002 [0.000–0.018] | 0.366 # | |
| Z-score ° | Mean ± SD | 0.314 ± 0.635 | −0.071 ± 1.060 | |||
| Synergistaceae * | Relative abundance (%) | Median [IQR] | 0.001 [0.000–0.013] | 0.002 [0.000–0.006] | 0.603 # | |
| Z-score ° | Mean ± SD | −0.181 ± 0.995 | 0.041 ± 1.013 | |||
| Erysipelotrichaceae | Relative abundance (%) | Median [IQR] | 0.138 [0.090–1.438] | 0.092 [0.046–0.450] | 0.338 # | |
| Z-score ° | Mean ± SD | 0.332 ± 1.111 | −0.075 ± 0.977 | |||
| Clostridiales Family XIII Incertae Sedis * | Relative abundance (%) | Median [IQR] | 0.006 [0.000–0.017] | 0.013 [0.002–0.022] | 0.107 # | |
| Z-score ° | Mean ± SD | −0.551 ± 1.302 | 0.124 ± 0.898 | |||
| Veillonellaceae * | Relative abundance (%) | Median [IQR] | 0.146 [0.044–1.892] | 0.836 [0.125–3.110] | 0.590 # | |
| Z-score ° | Mean ± SD | −0.188 ± 1.189 | 0.042 ± 0.970 | |||
| Cluster centroid | Z-score (means) | Mean ± SD | 0.454 ± 0.159 | –0.103 ± 0.147 | <0.001 # | |
| Genus | Fusobacterium | Relative abundance (%) | Median [IQR] | 0.012 [0.000–0.039] | 0.000 [0.000–0.000] | 0.001 # |
| Z-score ° | Mean ± SD | 1.067 ± 1.351 | −0.241 ± 0.736 | |||
| Butyricimonas | Relative abundance (%) | Median [IQR] | 0.800 [0.341–0.895] | 0.229 [0.103–0.484] | 0.201 # | |
| Z-score ° | Mean ± SD | 0.440 ± 0.751 | −0.099 ± 1.032 | |||
| Blautia | Relative abundance (%) | Median [IQR] | 1.794 [0.639–6.945] | 0.544 [0.323–0.831] | 0.018 # | |
| Z-score ° | Mean ± SD | 0.791 ± 1.066 | −0.179 ± 0.909 | |||
| Paraprevotella | Relative abundance (%) | Median [IQR] | 0.053 [0.007–1.231] | 0.007 [0.002–0.562] | 0.395 # | |
| Z-score ° | Mean ± SD | 0.295 ± 1.130 | −0.067 ± 0.976 | |||
| Klebsiella * | Relative abundance (%) | Median [IQR] | 0.001 [0.000–0.003] | 0.000 [0.000–0.003] | 0.889 # | |
| Z-score ° | Mean ± SD | −0.049 ± 0.674 | 0.011 ± 1.069 | |||
| Olsenella * | Relative abundance (%) | Median [IQR] | 0.000 [0.000–0.000] | 0.000 [0.000–0.000] | 0.735 # | |
| Z-score ° | Mean ± SD | −0.118 ± 0.944 | 0.027 ± 1.025 | |||
| Cronobacter | Relative abundance (%) | Median [IQR] | 0.000 [0.000–0.000] | 0.000 [0.000–0.000] | 0.142 # | |
| Z-score ° | Mean ± SD | 0.504 ± 1.783 | −0.114 ± 0.725 | |||
| Cluster centroid | Z-score (means) | Mean ± SD | 0.466 ± 0.264 | –0.105 ± 0.156 | <0.001 # | |
| Species | Parabacteroides merdae | Relative abundance (%) | Median [IQR] | 0.000 [0.000–0.001] | 0.000 [0.000–0.000] | 0.006 # |
| Z-score ° | Mean ± SD | 0.910 ± 2.108 | −0.205 ± 0.329 | |||
| Parabacteroides sp. DJF B084 | Relative abundance (%) | Median [IQR] | 0.000 [0.000–0.109] | 0.000 [0.000–0.000] | 0.025 # | |
| Z-score ° | Mean ± SD | 0.753 ± 1.805 | −0.170 ± 0.648 | |||
| Collinsella massiliensis | Relative abundance (%) | Median [IQR] | 0.000 [0.000–0.000] | 0.000 [0.000–0.000] | 0.069 # | |
| Z-score ° | Mean ± SD | 0.620 ± 2.198 | −0.140 ± 0.397 | |||
| Odoribacter sp. S90 | Relative abundance (%) | Median [IQR] | 0.000 [0.000–0.000] | Absent | 0.035 0.035 § | |
| Z-score ° | Mean ± SD | 0.718 ± 2.330 | ||||
| Ruminococcus sp. AT10 | Relative abundance (%) | Median [IQR] | 0.000 [0.000–0.003] | 0.000 [0.000–0.000] | 0.132 # | |
| Z-score ° | Mean ± SD | 0.517 ± 1.828 | −0.117 ± 0.699 | |||
| Clostridium piliforme | Relative abundance (%) | Median [IQR] | 0.000 [0.000–0.002] | 0.000 [0.000–0.000] | 0.051 # | |
| Z-score ° | Mean ± SD | 0.663 ± 1.848 | −0.150 ± 0.651 | |||
| Enorma massiliensis | Relative abundance (%) | Median [IQR] | 0.000 [0.000–0.091] | 0.000 [0.000–0.000] | 0.109 # | |
| Z-score ° | Mean ± SD | 0.549 ± 1.632 | −0.124 ± 0.784 | |||
| Megamonas rupellensis | Relative abundance (%) | Median [IQR] | 0.000 [0.000–0.187] | 0.000 [0.000–0.000] | 0.061 # | |
| Z-score ° | Mean ± SD | 0.637 ± 1.703 | −0.144 ± 0.733 | |||
| Bacteroides ovatus | Relative abundance (%) | Median [IQR] | 0.902 [0.736–1.158] | 0.644 [0.004–1.191] | 0.077 # | |
| Z-score ° | Mean ± SD | 0.604 ± 0.351 | −0.136 ± 1.051 | |||
| Butyricimonas sp. GD2 | Relative abundance (%) | Median [IQR] | 0.042 [0.000–0.185] | 0.000 [0.000–0.068] | 0.352 # | |
| Z-score ° | Mean ± SD | 0.322 ± 1.170 | −0.073 ± 0.964 | |||
| Faecalibacterium prausnitzii * | Relative abundance (%) | Median [IQR] | 0.920 [0.450–2.506] | 5.561 [2.425–9.540] | 0.061 # | |
| Z-score ° | Mean ± SD | −0.636 ± 0.900 | 0.144 ± 0.978 | |||
| Ruminococcus sp. DJF VR70k1 | Relative abundance (%) | Median [IQR] | 0.000 [0.000–0.004] | 0.000 [0.000–0.003] | 0.910 # | |
| Z-score ° | Mean ± SD | 0.039 ± 0.861 | −0.009 ± 1.042 | |||
| Clostridium aldenense * | Relative abundance (%) | Median [IQR] | 0.006 [0.006–0.118] | 0.172 [0.035–0.495] | 0.017 # | |
| Z-score ° | Mean ± SD | −0.802 ± 1.265 | 0.181 ± 0.854 | |||
| Tetragenococcus halophilus * | Relative abundance (%) | Median [IQR] | Absent | 0.000 [0.000–0.000] | 0.322 § | |
| Z-score ° | Mean ± SD | 0.064 ± 1.100 | ||||
| Cluster centroid | Z-score (means) | Mean ± SD | 0.575 ± 0.093 | −0.130 ± 0.074 | <0.001 # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manchia, M.; Fontana, A.; Panebianco, C.; Paribello, P.; Arzedi, C.; Cossu, E.; Garzilli, M.; Montis, M.A.; Mura, A.; Pisanu, C.; et al. Involvement of Gut Microbiota in Schizophrenia and Treatment Resistance to Antipsychotics. Biomedicines 2021, 9, 875. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9080875
Manchia M, Fontana A, Panebianco C, Paribello P, Arzedi C, Cossu E, Garzilli M, Montis MA, Mura A, Pisanu C, et al. Involvement of Gut Microbiota in Schizophrenia and Treatment Resistance to Antipsychotics. Biomedicines. 2021; 9(8):875. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9080875
Chicago/Turabian StyleManchia, Mirko, Andrea Fontana, Concetta Panebianco, Pasquale Paribello, Carlo Arzedi, Eleonora Cossu, Mario Garzilli, Maria Antonietta Montis, Andrea Mura, Claudia Pisanu, and et al. 2021. "Involvement of Gut Microbiota in Schizophrenia and Treatment Resistance to Antipsychotics" Biomedicines 9, no. 8: 875. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9080875