
MARCO+ Macrophage Dynamics in Regenerating Liver after 70% Liver Resection in Mice

 ,
, 
 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Animal Model
2.3. Isolation of Macrophages
2.4. Flow Cytometry Assay
2.5. Cell Culture of Macrophages
2.6. Activation of Macrophage Cell Cultures with LPS
2.7. Immunocytochemistry and Immunohistochemistry
2.8. Reverse Transcription Polymerase Chain Reaction (RT PCR) Gene Expression Assay
2.9. Immunoblotting
2.10. Statistics
3. Results
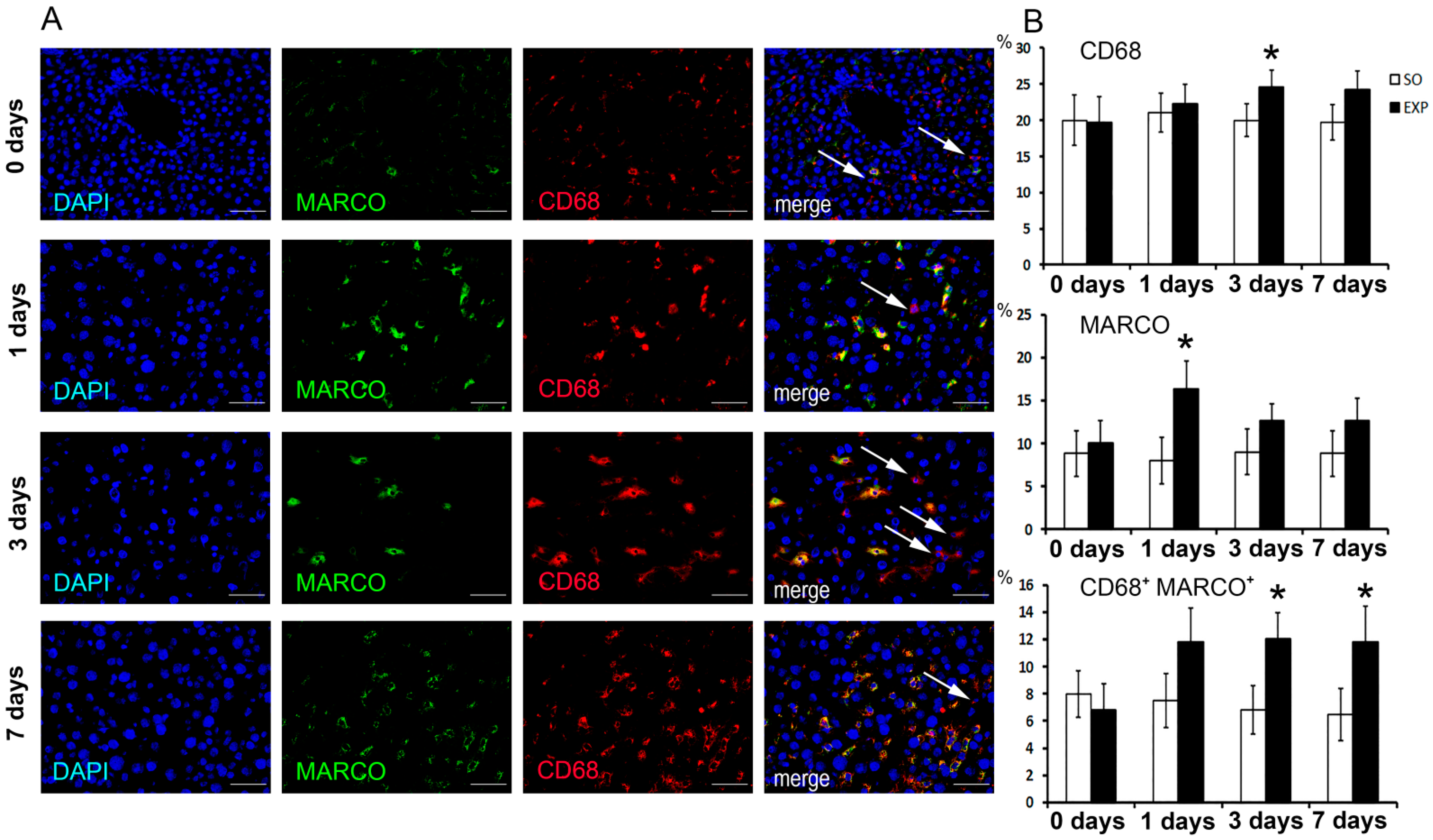
3.1. Macrophage Population Dynamics in Regenerating Livers
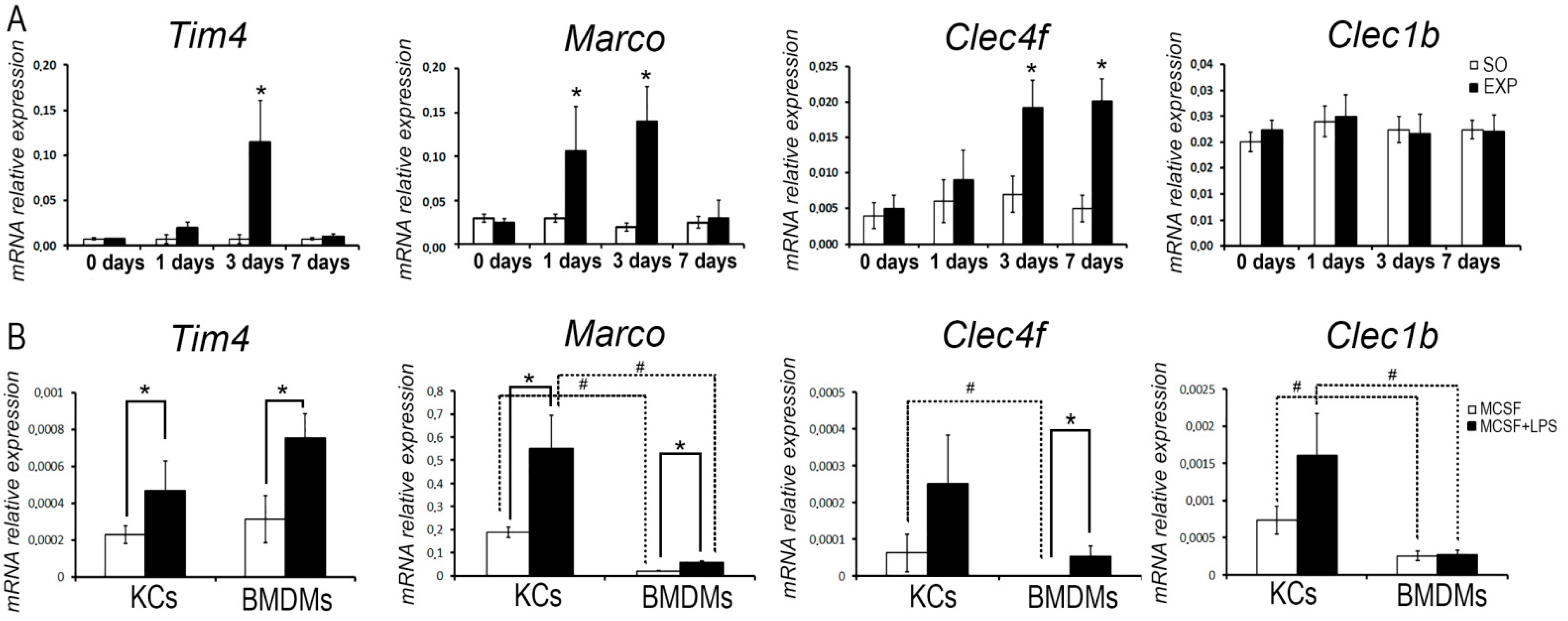
3.2. Transcriptional Dynamics of Macrophage Marker-Encoding Genes in Regenerating Livers
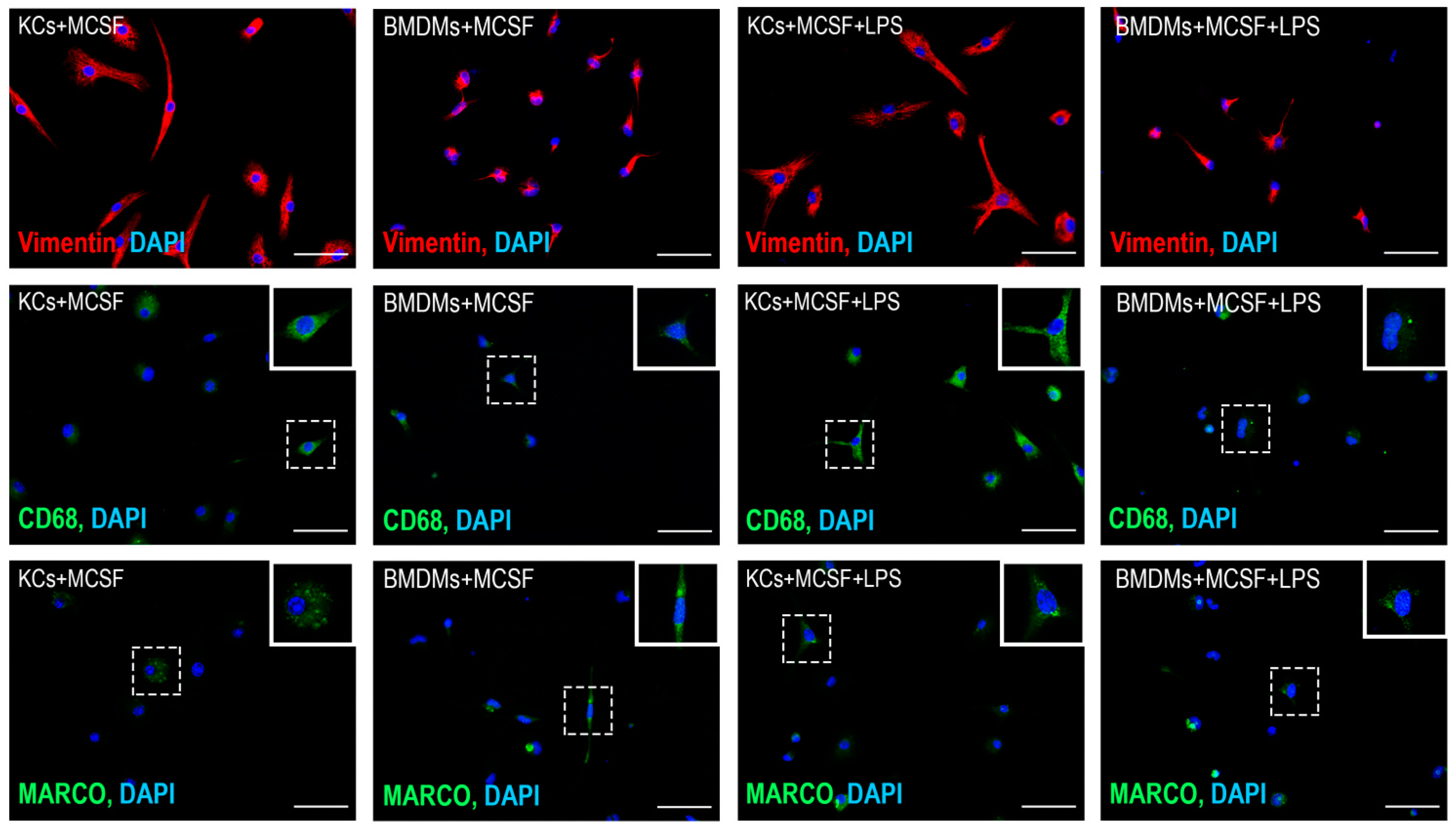
3.3. Comparative Analysis of Lipopolysaccharide (LPS) Effects on the Expression of Macrophage Markers in Bone Marrow-Derived Macrophages and Kupffer Cells
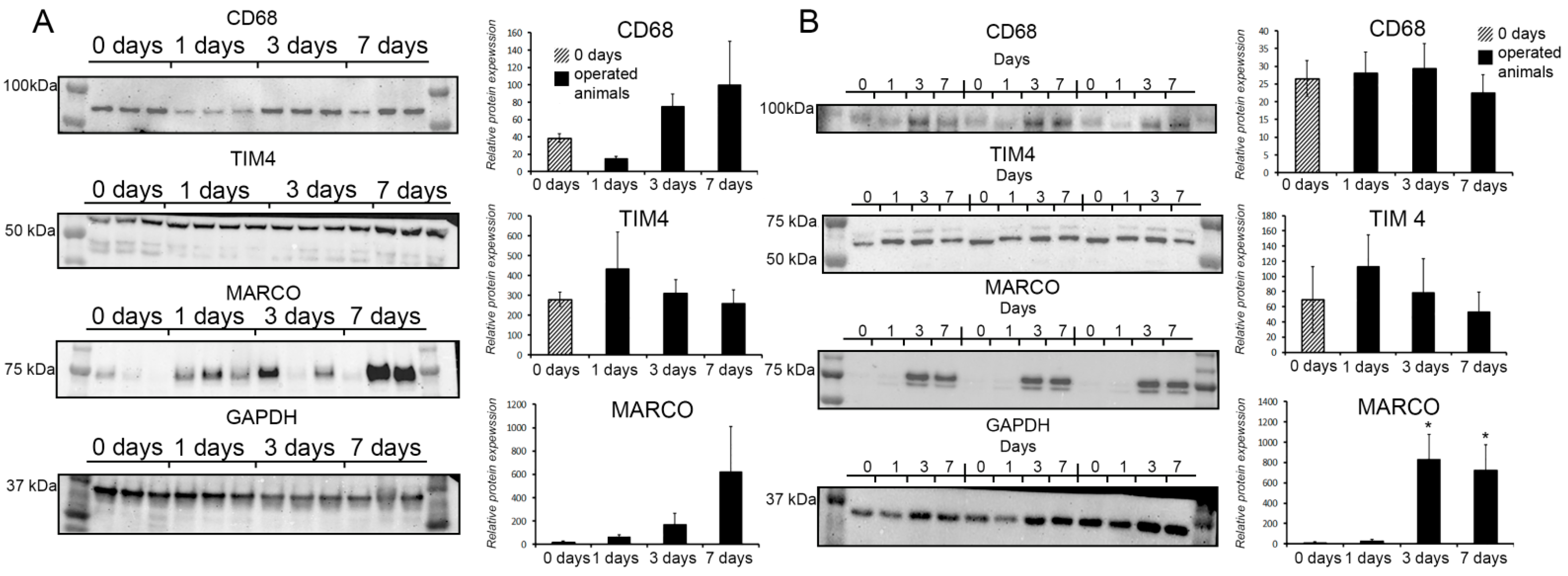
3.4. Protein Expression Dynamics for CD68, TIM4, and MARCO in Total Tissues and Macrophages of Regenerating Livers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liaskou, E.; Wilson, D.V.; Oo, Y.H. Innate immune cells in liver inflammation. Mediators Inflamm. 2012, 2012, 949157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campana, L.; Esser, H.; Huch, M.; Forbes, S. Liver regeneration and inflammation: From fundamental science to clinical applications. Nat. Rev. Mol. Cell Biol. 2021, 21, 608–624. [Google Scholar] [CrossRef]
- You, Q.; Holt, M.; Yin, H.; Li, G.; Hu, C.J.; Ju, C. Role of hepatic resident and infiltrating macrophages in liver repair after acute injury. Biochem. Pharmacol. 2013, 86, 836–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zigmond, E.; Samia-Grinberg, S.; Pasmanik-Chor, M.; Brazowski, E.; Shibolet, O.; Halpern, Z.; Varol, C. Infiltrating Monocyte-Derived Macrophages and Resident Kupffer Cells Display Different Ontogeny and Functions in Acute Liver Injury. J. Immunol. 2014, 193, 344–353. [Google Scholar] [CrossRef] [Green Version]
- Elchaninov, A.; Lokhonina, A.; Nikitina, M.; Vishnyakova, P.; Makarov, A.; Arutyunyan, I.; Poltavets, A.; Kananykhina, E.; Kovalchuk, S.; Karpulevich, E.; et al. Comparative analysis of the transcriptome, proteome, and mirna profile of kupffer cells and monocytes. Biomedicines 2020, 8, 627. [Google Scholar] [CrossRef] [PubMed]
- Beattie, L.; Sawtell, A.; Mann, J.; Frame, T.C.M.; Teal, B.; de Labastida Rivera, F.; Brown, N.; Walwyn-Brown, K.; Moore, J.W.J.; MacDonald, S.; et al. Bone marrow-derived and resident liver macrophages display unique transcriptomic signatures but similar biological functions. J. Hepatol. 2016, 65, 758–768. [Google Scholar] [CrossRef] [Green Version]
- Perdiguero, E.G.; Geissmann, F. The development and maintenance of resident macrophages. Nat. Immunol. 2016, 17, 2–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeffel, G.; Ginhoux, F. Fetal monocytes and the origins of tissue-resident macrophages. Cell. Immunol. 2018, 330, 5–15. [Google Scholar] [CrossRef]
- Gao, C.; Peng, J. All routes lead to Rome: Multifaceted origin of hepatocytes during liver regeneration. Cell Regen. 2021, 10, 2. [Google Scholar]
- Michalopoulos, G.K.; Bhushan, B. Liver regeneration: Biological and pathological mechanisms and implications. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 40–55. [Google Scholar] [CrossRef]
- Elchaninov, A.V.; Fatkhudinov, T.K.; Usman, N.Y.; Kananykhina, E.Y.; Arutyunyan, I.V.; Makarov, A.V.; Lokhonina, A.V.; Eremina, I.Z.; Surovtsev, V.V.; Goldshtein, D.V.; et al. Dynamics of macrophage populations of the liver after subtotal hepatectomy in rats. BMC Immunol. 2018, 19, 23. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, K.; Nakashima, H.; Ikarashi, M.; Kinoshita, M.; Nakashima, M.; Aosasa, S.; Seki, S.; Yamamoto, J. Mouse CD11b+Kupffer cells recruited from bone marrow accelerate liver regeneration after partial hepatectomy. PLoS ONE 2015, 10, e0136774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elchaninov, A.; Nikitina, M.; Vishnyakova, P.; Lokhonina, A.; Makarov, A.; Sukhikh, G.; Fatkhudinov, T. Macro- and microtranscriptomic evidence of the monocyte recruitment to regenerating liver after partial hepatectomy in mouse model. Biomed. Pharmacother. 2021, 138, 111516. [Google Scholar] [CrossRef] [PubMed]
- Jing, J.; Yang, I.V.; Hui, L.; Patel, J.A.; Evans, C.M.; Prikeris, R.; Kobzik, L.; O’Connor, B.P.; Schwartz, D.A. Role of Macrophage Receptor with Collagenous Structure in Innate Immune Tolerance. J. Immunol. 2013, 190, 6360–6367. [Google Scholar] [CrossRef] [Green Version]
- Kanno, S.; Hirano, S.; Sakamoto, T.; Furuyama, A.; Takase, H.; Kato, H.; Fukuta, M.; Aoki, Y. Scavenger receptor MARCO contributes to cellular internalization of exosomes by dynamin-dependent endocytosis and macropinocytosis. Sci. Rep. 2020, 10, 21795. [Google Scholar] [CrossRef]
- Scott, C.L.; Zheng, F.; De Baetselier, P.; Martens, L.; Saeys, Y.; De Prijck, S.; Lippens, S.; Abels, C.; Schoonooghe, S.; Raes, G.; et al. Bone marrow-derived monocytes give rise to self-renewing and fully differentiated Kupffer cells. Nat. Commun. 2016, 7, 10321. [Google Scholar] [CrossRef]
- Bonnardel, J.; T’Jonck, W.; Gaublomme, D.; Browaeys, R.; Scott, C.L.; Martens, L.; Vanneste, B.; De Prijck, S.; Nedospasov, S.A.; Kremer, A.; et al. Stellate Cells, Hepatocytes, and Endothelial Cells Imprint the Kupffer Cell Identity on Monocytes Colonizing the Liver Macrophage Niche. Immunity 2019, 51, 638–654.e9. [Google Scholar] [CrossRef] [Green Version]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage Activation and Polarization: Nomenclature and Experimental Guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Nevzorova, Y.; Tolba, R.; Trautwein, C.; Liedtke, C. Partial hepatectomy in mice. Lab. Anim. 2015, 49, 81–88. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Elchaninov, A.; Sukhikh, G.; Fatkhudinov, T. Evolution of Regeneration in Animals: A Tangled Story. Front. Ecol. Evol. 2021, 9, 121. [Google Scholar] [CrossRef]
- Carlson, B.M. Some principles of regeneration in mammalian systems. Anat. Rec. B New Anat. 2005, 287, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Maden, M. The evolution of regeneration-Where does that leave mammals? Int. J. Dev. Biol. 2018, 62, 369–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mescher, A.L.; Neff, A.W.; King, M.W. Inflammation and immunity in organ regeneration. Dev. Comp. Immunol. 2017, 66, 98–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Hua, J. Immune cells in liver regeneration. Oncotarget 2017, 8, 3628–3639.mune cells in liver regeneration. Oncotarget 2017, 8, 3628–3639. [Google Scholar] [CrossRef] [Green Version]
- Gao, B.; Radaeva, S.; Park, O. Liver natural killer and natural killer T cells: Immunobiology and emerging roles in liver diseases. J. Leukoc. Biol. 2009, 86, 513–528. [Google Scholar] [CrossRef]
- Michalopoulos, G.K. Advances in liver regeneration. Expert Rev. Gastroenterol. Hepatol. 2014, 8, 897–907. [Google Scholar] [CrossRef]
- Michalopoulos, G.K. Principles of liver regeneration and growth homeostasis. Compr. Physiol. 2013, 3, 485–513. [Google Scholar] [CrossRef]
- Goh, Y.P.S.; Henderson, N.C.; Heredia, J.E.; Eagle, A.R.; Odegaard, J.I.; Lehwald, N.; Nguyen, K.D.; Sheppard, D.; Mukundan, L.; Locksley, R.M.; et al. Eosinophils secrete IL-4 to facilitate liver regeneration. Proc. Natl. Acad. Sci. USA 2013, 110, 9914–9919. [Google Scholar] [CrossRef] [Green Version]
- Nachmany, I.; Bogoch, Y.; Sivan, A.; Amar, O.; Bondar, E.; Zohar, N.; Yakubovsky, O.; Fainaru, O.; Klausner, J.M.; Pencovich, N. CD11b + Ly6G + myeloid-derived suppressor cells promote liver regeneration in a murine model of major hepatectomy. FASEB J. 2019, 33, 5967–5978. [Google Scholar] [CrossRef] [PubMed]
- Fausto, N.; Campbell, J.S.; Riehle, K.J. Liver regeneration. J. Hepatol. 2012, 57, 692–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teh, Y.C.; Ding, J.L.; Ng, L.G.; Chong, S.Z. Capturing the Fantastic Voyage of Monocytes Through Time and Space. Front. Immunol. 2019, 10, 834. [Google Scholar] [CrossRef] [PubMed]
- Guilliams, M.; Mildner, A.; Yona, S. Developmental and Functional Heterogeneity of Monocytes. Immunity 2018, 49, 595–613. [Google Scholar] [CrossRef] [Green Version]
- Kangas, M.; Brännström, A.; Elomma, O.; Matsuda, Y.; Eddy, R.; Shows, T.B.; Tryggvason, K. Structure and chromosomal localization of the human and murine genes for the macrophage MARCO receptor. Genomics 1999, 58, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Kraal, G.; Van Der Laan, L.J.W.; Elomaa, O.; Tryggvason, K. The macrophage receptor MARCO. Microbes Infect. 2000, 2, 313–316. [Google Scholar] [CrossRef]
- Gensel, J.C.; Kopper, T.J.; Zhang, B.; Orr, M.B.; Bailey, W.M. Predictive screening of M1 and M2 macrophages reveals the immunomodulatory effectiveness of post spinal cord injury azithromycin treatment. Sci. Rep. 2017, 7, 40144. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, S.; Varin, A.; Chen, Y.; Liu, B.; Tryggvason, K.; Gordon, S. SR-A/MARCO-mediated ligand delivery enhances intracellular TLR and NLR function, but ligand scavenging from cell surface limits TLR4 response to pathogens. Blood 2011, 117, 1319–1328. [Google Scholar] [CrossRef] [PubMed]
- Georgoudaki, A.M.; Prokopec, K.; Boura, V.; Hellqvist, E.; Sohn, S.; Östling, J.; Dahan, R.; Harris, R.; Rantalainen, M.; Klevebring, D.; et al. Reprogramming Tumor-Associated Macrophages by Antibody Targeting Inhibits Cancer Progression and Metastasis. Cell Rep. 2016, 15, 2000–2011. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Bao, Q.; Kumar, S.; Hu, M.; Wang, G.Y.; Dai, G. Four waves of hepatocyte proliferation linked with three waves of hepatic fat accumulation during partial hepatectomy-induced liver regeneration. PLoS ONE 2012, 7, e30675. [Google Scholar] [CrossRef]
- La Fleur, L.; Boura, V.F.; Alexeyenko, A.; Berglund, A.; Pontén, V.; Mattsson, J.S.M.; Djureinovic, D.; Persson, J.; Brunnström, H.; Isaksson, J.; et al. Expression of scavenger receptor MARCO defines a targetable tumor-associated macrophage subset in non-small cell lung cancer. Int. J. Cancer 2018, 143, 1741–1752. [Google Scholar] [CrossRef] [Green Version]
- La Fleur, L.; Botling, J.; He, F.; Pelicano, C.; Zhou, C.; He, C.; Palano, G.; Mezheyeuski, A.; Micke, P.; Ravetch, J.V.; et al. Targeting MARCO and IL37R on Immunosuppressive Macrophages in Lung Cancer Blocks Regulatory T Cells and Supports Cytotoxic Lymphocyte Function. Cancer Res. 2021, 81, 956–967. [Google Scholar] [CrossRef]
- Xiao, Y.; Chen, B.; Yang, K.; Wang, Q.; Liu, P.; Gu, Y.; Zhong, Q.; Liu, Z.; He, Y.; Liu, Q. Down-regulation of MARCO associates with tumor progression in hepatocellular carcinoma. Exp. Cell Res. 2019, 383, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Qian, X.; Wang, N.; Ding, Y.; Li, H.; Zhao, Y.; Yao, S. Inhibition of MARCO ameliorates silica-induced pulmonary fibrosis by regulating epithelial-mesenchymal transition. Toxicol. Lett. 2019, 301, 64–72. [Google Scholar] [CrossRef]
- Wu, M.; Gibbons, J.G.; DeLoid, G.M.; Bedugnis, A.S.; Thimmulappa, R.K.; Biswal, S.; Kobzik, L. Immunomodulators targeting MARCO expression improve resistance to postinfluenza bacterial pneumonia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2017, 313, L138–L153. [Google Scholar] [CrossRef]
- Larionova, I.; Tuguzbaeva, G.; Ponomaryova, A.; Stakheyeva, M.; Cherdyntseva, N.; Pavlov, V.; Choinzonov, E.; Kzhyshkowska, J. Tumor-Associated Macrophages in Human Breast, Colorectal, Lung, Ovarian and Prostate Cancers. Front. Oncol. 2020, 10, 566511. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Ye, R.D. Microglial Aβ Receptors in Alzheimer’s Disease. Cell. Mol. Neurobiol. 2014, 35, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Miyanishi, M.; Tada, K.; Koike, M.; Uchiyama, Y.; Kitamura, T.; Nagata, S. Identification of Tim4 as a phosphatidylserine receptor. Nature 2007, 450, 435–439. [Google Scholar] [CrossRef] [Green Version]
- Tietjen, G.T.; Gong, Z.; Chen, C.H.; Vargas, E.; Crooks, J.E.; Cao, K.D.; Heffern, C.T.; Henderson, J.M.; Meron, M.; Lin, B.; et al. Molecular mechanism for differential recognition of membrane phosphatidylserine by the immune regulatory receptor Tim4. Proc. Natl. Acad. Sci. USA 2014, 111, E1463–E1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGrath, M.M. Diverse roles of TIM4 in immune activation: Implications for alloimmunity. Curr. Opin. Organ. Transplant. 2018, 23, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Xu, X.; Li, J.; Gong, J.; Li, M. TIM-4 blockade of KCs combined with exogenous TGF-β injection helps to reverse acute rejection and prolong the survival rate of mice receiving liver allografts. Int. J. Mol. Med. 2018, 42, 346–358. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Tang, Y.; Hoover, C.; Kondo, Y.; Huang, D.; Restagno, D.; Shao, B.; Gao, L.; Michael McDaniel, J.; Zhou, M.; et al. Kupffer cell receptor CLEC4F is important for the destruction of desialylated platelets in mice. Cell Death Differ. 2021, 1–13. [Google Scholar] [CrossRef]
- Yang, C.Y.; Chen, J.B.; Tsai, T.F.; Tsai, Y.C.; Tsai, C.Y.; Liang, P.H.; Hsu, T.L.; Wu, C.Y.; Netea, M.G.; Wong, C.H.; et al. CLEC4F is an inducible C-type lectin in F4/80-positive cells and is involved in alpha-galactosylceramide presentation in liver. PLoS ONE 2013, 8, e65070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, D.; Luo, M.; Liu, B. The Role of CLEC-2 and Its Ligands in Thromboinflammation. Front. Immunol. 2021, 12, 688643. [Google Scholar] [CrossRef] [PubMed]
- Lowe, K.L.; Navarro-Núñez, L.; Bénézech, C.; Nayar, S.; Kingston, B.L.; Nieswandt, B.; Barone, F.; Watson, S.P.; Buckley, C.D.; Desanti, G.E. The expression of mouse CLEC-2 on leucocyte subsets varies according to their anatomical location and inflammatory state. Eur. J. Immunol. 2015, 45, 2484. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.H.; Chung, C.H.; Hsu, C.C.; Huang, T.Y.; Huang, T.F. A novel mechanism of cytokine release in phagocytes induced by aggretin, a snake venom C-type lectin protein, through CLEC-2 ligation. J. Thromb. Haemost. 2010, 8, 2563–2570. [Google Scholar] [CrossRef]
- Gottfried, E.; Kunz-Schughart, L.A.; Weber, A.; Rehli, M.; Peuker, A.; Müller, A.; Kastenberger, M.; Brockhoff, G.; Andreesen, R.; Kreutz, M. Expression of CD68 in Non-Myeloid Cell Types. Scand. J. Immunol. 2008, 67, 453–463. [Google Scholar] [CrossRef]
- Bush, S.J.; McCulloch, M.E.B.; Lisowski, Z.M.; Muriuki, C.; Clark, E.L.; Young, R.; Pridans, C.; Prendergast, J.G.D.; Summers, K.M.; Hume, D.A. Species-Specificity of Transcriptional Regulation and the Response to Lipopolysaccharide in Mammalian Macrophages. Front. Cell Dev. Biol. 2020, 8, 661. [Google Scholar] [CrossRef] [PubMed]
- Michalopoulos, G.K.; DeFrances, M.C. Liver Regeneration. Science 1997, 276, 60–66. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tim4 | for | ATTAACAACAAGTGAGTCTTTGCAG |
| rev | CTTCAGTGTTGTCTGGCCTCT | |
| Marco | for | CCAGGACTTTCAGGTGCCAA |
| rev | TGGCCAGAAGACCCTTTCAT | |
| Clec4f | for | TGCAGGACAGGCTTTAGGAA |
| rev | TGACTTAGGCCCCAGTCCTTG | |
| Clec1b | for | GTTGGATTGGATTATCACGCCAG |
| rev | TCCCAGAAAGATTAATCCCGTT | |
| Gapdh | for | AGGCCGGTGCTGAGTATGTC |
| rev | TGCCTGCTTCACCACCTTCT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elchaninov, A.; Lokhonina, A.; Vishnyakova, P.; Soboleva, A.; Poltavets, A.; Artemova, D.; Makarov, A.; Glinkina, V.; Goldshtein, D.; Bolshakova, G.; et al. MARCO+ Macrophage Dynamics in Regenerating Liver after 70% Liver Resection in Mice. Biomedicines 2021, 9, 1129. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9091129
Elchaninov A, Lokhonina A, Vishnyakova P, Soboleva A, Poltavets A, Artemova D, Makarov A, Glinkina V, Goldshtein D, Bolshakova G, et al. MARCO+ Macrophage Dynamics in Regenerating Liver after 70% Liver Resection in Mice. Biomedicines. 2021; 9(9):1129. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9091129
Chicago/Turabian StyleElchaninov, Andrey, Anastasia Lokhonina, Polina Vishnyakova, Anna Soboleva, Anastasiya Poltavets, Daria Artemova, Andrey Makarov, Valeria Glinkina, Dmitry Goldshtein, Galina Bolshakova, and et al. 2021. "MARCO+ Macrophage Dynamics in Regenerating Liver after 70% Liver Resection in Mice" Biomedicines 9, no. 9: 1129. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9091129