
Perspective on Similarities and Possible Overlaps of Congenital Disease Formation—Exemplified on a Case of Congenital Diaphragmatic Hernia and Neuroblastoma in a Neonate

Abstract
:1. Introduction
2. Materials and Methods
2.1. Tumor Tissue
2.2. H&E and Immunostaining
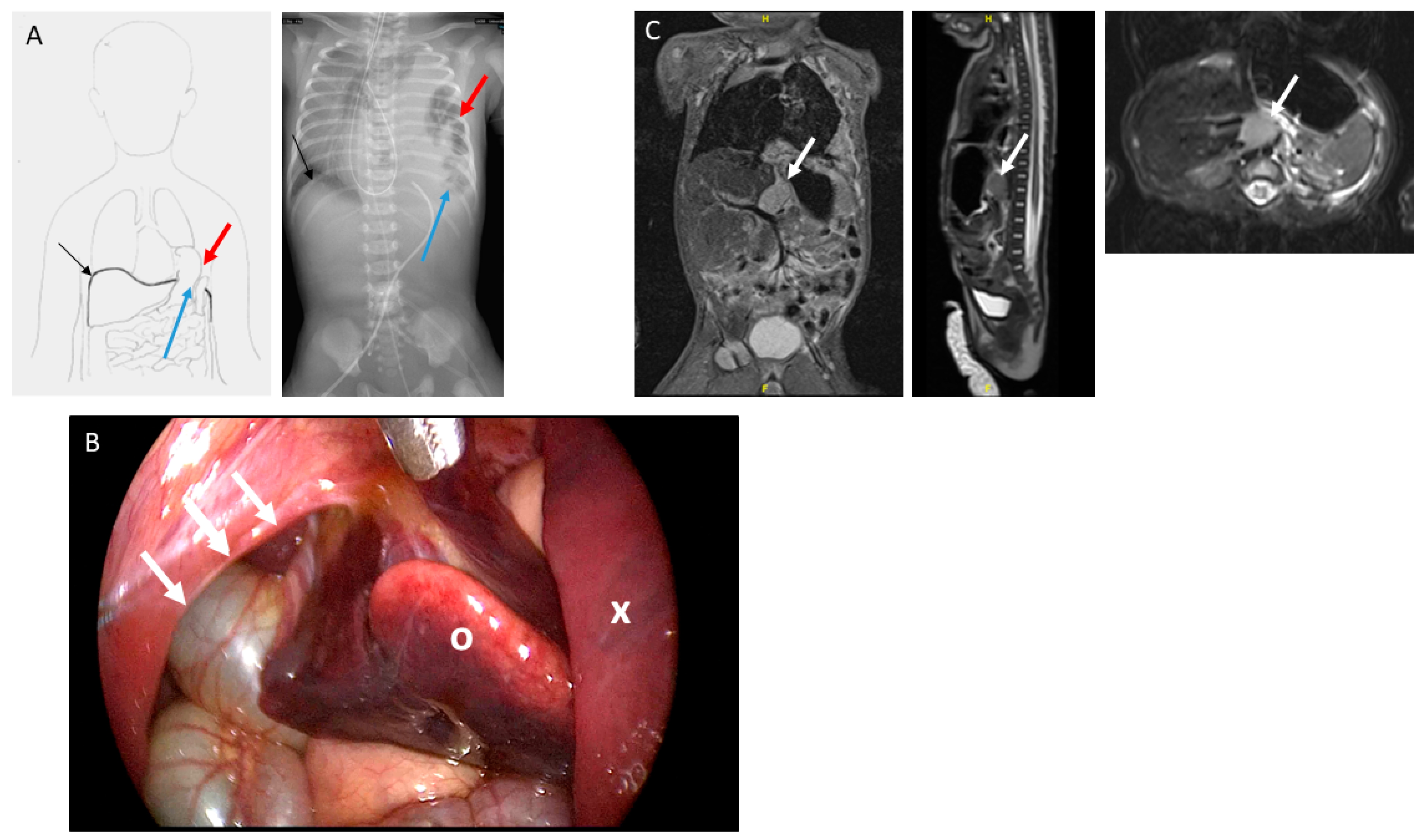
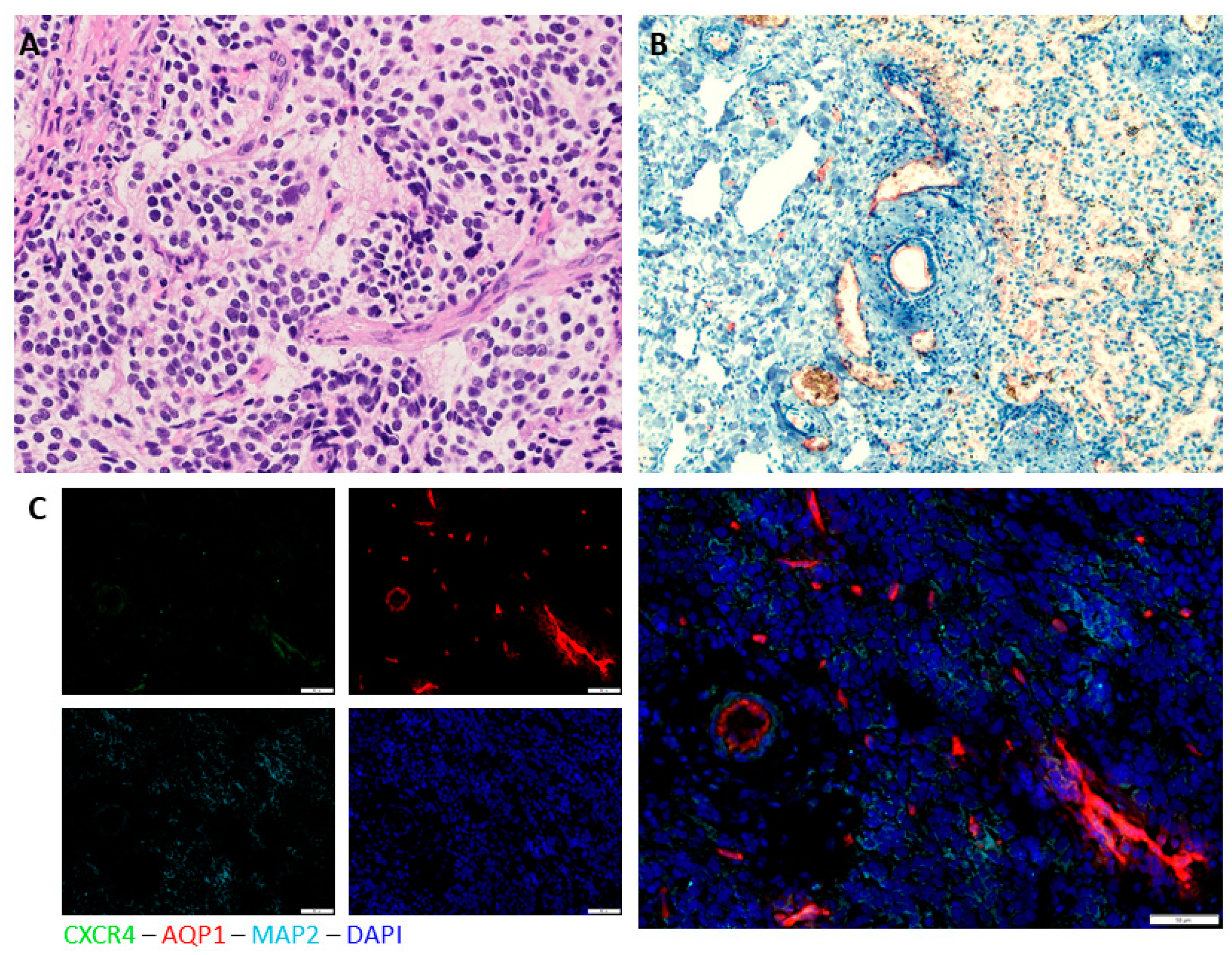
3. Results
Immunohistochemical Analysis
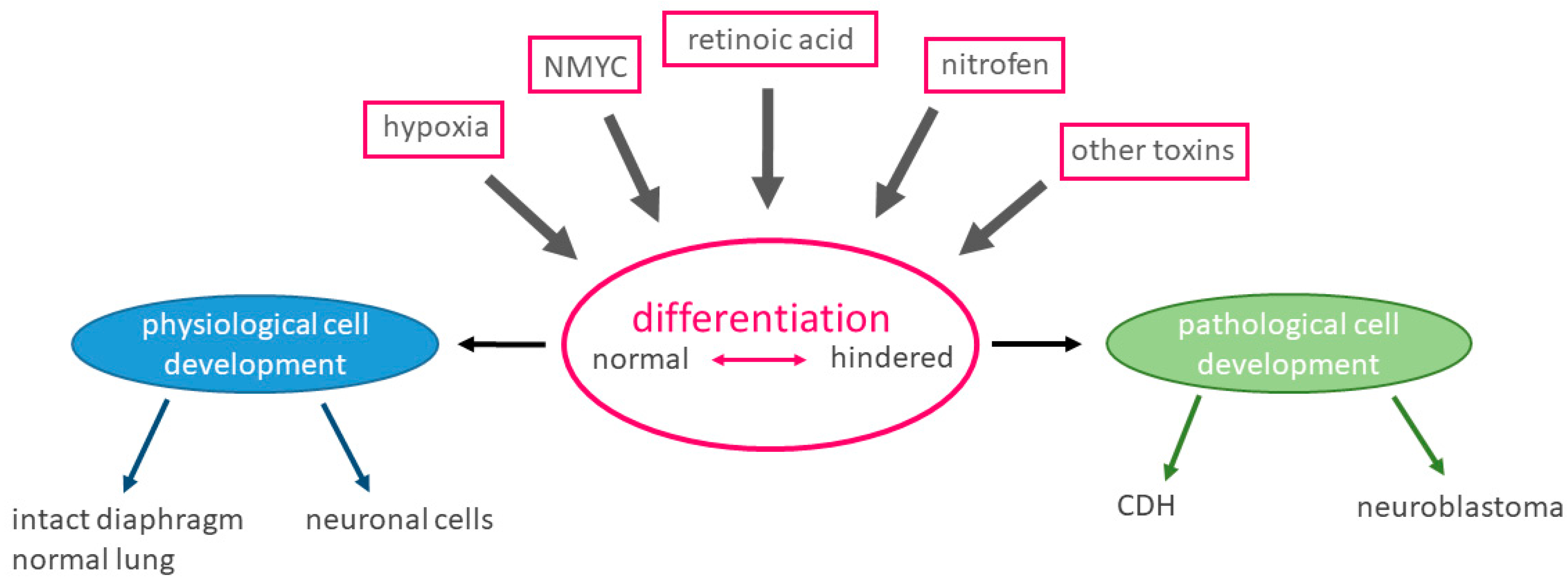
4. Discussion
4.1. Embryogenesis
4.2. Localization
4.3. Retinoic Acid Pathway
4.4. Influence of NMYC
4.5. Role of Hypoxia and Hypoxia-Associated Factors in Development
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Global Genes. Available online: https://globalgenes.org/rare-facts/ (accessed on 20 February 2020).
- Orphanet. Available online: www.orpha.net (accessed on 20 February 2020).
- Regulation (EC) No 141/2000 of the European Parliament and of the Council of 16 December 1999 on Orphan Medicinal Products. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32000R0141 (accessed on 20 February 2020).
- Paoletti, M.; Raffler, G.; Gaffi, M.S.; Antounians, L.; Lauriti, G.; Zani, A. Prevalence and risk factors for congenital diaphragmatic hernia: A global view. J. Pediatric Surg. 2020, 55, 2297–2307. [Google Scholar] [CrossRef]
- Dumpa, V.; Chandrasekharan, P. Congenital Diaphragmatic Hernia; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- McGivern, M.R.; Best, K.E.; Rankin, J.; Wellesley, D.; Greenlees, R.; Addor, M.C.; Arriola, L.; de Walle, H.; Barisic, I.; Beres, J.; et al. Epidemiology of congenital diaphragmatic hernia in Europe: A register-based study. Arch. Dis. Child Fetal. Neonatal. Ed. 2015, 100, F137–F144. [Google Scholar] [CrossRef] [PubMed]
- Iritani, I. Experimental study on embryogenesis of congenital diaphragmatic hernia. Anat. Embryol. 1984, 169, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Kluth, D.; Keijzer, R.; Hertl, M.; Tibboel, D. Embryology of congenital diaphragmatic hernia. Semin. Pediatric Surg. 1996, 5, 224–233. [Google Scholar]
- Greer, J.J.; Allan, D.W.; Babiuk, R.P.; Lemke, R.P. Recent advances in understanding the pathogenesis of nitrofen-induced congenital diaphragmatic hernia. Pediatric Pulmonol. 2000, 29, 394–399. [Google Scholar] [CrossRef]
- Liang, S.W.; Chen, G.; Luo, Y.G.; Chen, P.; Gu, J.H.; Xu, Q.Q.; Dang, Y.-W.; Qin, L.-T.; Lu, H.-P.; Huang, W.-T.; et al. Nomogram for predicting overall survival in children with neuroblastoma based on SEER database. Ann. Surg. Treat Res. 2020, 99, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Bosse, K.R.; Maris, J.M. Advances in the translational genomics of neuroblastoma: From improving risk stratification and revealing novel biology to identifying actionable genomic alterations. Cancer 2016, 122, 20–33. [Google Scholar] [CrossRef]
- Boeva, V.; Louis-Brennetot, C.; Peltier, A.; Durand, S.; Pierre-Eugene, C.; Raynal, V.; Etchevers, H.C.; Thomas, S.; Lermine, A.; Daudigeos-Dubus, E.; et al. Heterogeneity of neuroblastoma cell identity defined by transcriptional circuitries. Nat. Genet. 2017, 49, 1408–1413. [Google Scholar] [CrossRef]
- Ohira, M.; Morohashi, A.; Inuzuka, H.; Shishikura, T.; Kawamoto, T.; Kageyama, H.; Nakamura, Y.; Isogai, E.; Takayasu, H.; Sakiyama, S.; et al. Expression profiling and characterization of 4200 genes cloned from primary neuroblastomas: Identification of 305 genes differentially expressed between favorable and unfavorable subsets. Oncogene 2003, 22, 5525–5536. [Google Scholar] [CrossRef] [Green Version]
- Shipley, M.M.; Mangold, C.A.; Szpara, M.L. Differentiation of the SH-SY5Y Human Neuroblastoma Cell Line. J. Vis. Exp. JoVE 2016, 108, 53193. [Google Scholar] [CrossRef]
- Westermark, U.K.; Wilhelm, M.; Frenzel, A.; Henriksson, M.A. The MYCN oncogene and differentiation in neuroblastoma. Semin. Cancer Biol. 2011, 21, 256–266. [Google Scholar] [CrossRef]
- Wakamatsu, Y.; Watanabe, Y.; Nakamura, H.; Kondoh, H. Regulation of the neural crest cell fate by N-myc: Promotion of ventral migration and neuronal differentiation. Development 1997, 124, 1953–1962. [Google Scholar] [PubMed]
- Kang, J.H.; Rychahou, P.G.; Ishola, T.A.; Qiao, J.; Evers, B.M.; Chung, D.H. MYCN silencing induces differentiation and apoptosis in human neuroblastoma cells. Biochem. Biophys. Res. Commun. 2006, 351, 192–197. [Google Scholar] [CrossRef] [Green Version]
- Kaifi, J.T.; Yekebas, E.F.; Schurr, P.; Obonyo, D.; Wachowiak, R.; Busch, P.; Heinecke, A.; Pantel, K.; Izbicki, J.R. Tumor-cell homing to lymph nodes and bone marrow and CXCR4 expression in esophageal cancer. J. Natl. Cancer Inst. 2005, 97, 1840–1847. [Google Scholar] [CrossRef] [PubMed]
- Russell, H.V.; Hicks, J.; Okcu, M.F.; Nuchtern, J.G. CXCR4 expression in neuroblastoma primary tumors is associated with clinical presentation of bone and bone marrow metastases. J. Pediatric Surg. 2004, 39, 1506–1511. [Google Scholar] [CrossRef] [PubMed]
- Ameis, H.M.; Drenckhan, A.; von Loga, K.; Escherich, G.; Wenke, K.; Izbicki, J.R.; Reinshagen, K.; Gros, S.J. PGK1 as predictor of CXCR4 expression, bone marrow metastases and survival in neuroblastoma. PLoS ONE 2013, 8, e83701. [Google Scholar] [CrossRef]
- Pini, N.; Huo, Z.; Kym, U.; Holland-Cunz, S.; Gros, S.J. AQP1-Driven Migration Is Independent of Other Known Adverse Factors but Requires a Hypoxic Undifferentiated Cell Profile in Neuroblastoma. Children 2021, 8, 48. [Google Scholar] [CrossRef]
- Cruz, S.M.; Akinkuotu, A.C.; Cass, D.L.; Lee, T.C.; Cassady, C.I.; Mehollin-Ray, A.R.; Ruano, R.; Welty, S.E.; Olutoye, O.O. Space occupying lesions in the presence of congenital diaphragmatic hernia. J. Pediatric Surg. 2016, 51, 710–713. [Google Scholar] [CrossRef]
- Altmann, A.E.; Halliday, J.L.; Giles, G.G. Associations between congenital malformations and childhood cancer. A register-based case-control study. Br. J. Cancer 1998, 78, 1244–1249. [Google Scholar] [CrossRef] [Green Version]
- Berbel Tornero, O.; Ortega Garcia, J.A.; Ferrisi Tortajada, J.; Garcia Castell, J.; Donati Colomer, J.; Soldin, O.P.; Fuster Soler, J.L. Neonatal tumours and congenital malformations. An. Pediatric 2008, 68, 589–595. [Google Scholar] [CrossRef]
- Mito, K.; Amano, Y.; Oshiro, H.; Matsubara, D.; Fukushima, N.; Ono, S. Liver heterotopia associated with congenital diaphragmatic hernia: Two case reports and a review of the literature. Med. Baltim. 2019, 98, e14211. [Google Scholar] [CrossRef] [PubMed]
- Babiuk, R.P.; Zhang, W.; Clugston, R.; Allan, D.W.; Greer, J.J. Embryological origins and development of the rat diaphragm. J. Comp. Neurol. 2003, 455, 477–487. [Google Scholar] [CrossRef]
- Sefton, E.M.; Gallardo, M.; Kardon, G. Developmental origin and morphogenesis of the diaphragm, an essential mammalian muscle. Dev. Biol. 2018, 440, 64–73. [Google Scholar] [CrossRef]
- Kardon, G.; Ackerman, K.G.; McCulley, D.J.; Shen, Y.; Wynn, J.; Shang, L.; Bogenschutz, E.; Sun, X.; Chung, W.K. Congenital diaphragmatic hernias: From genes to mechanisms to therapies. Dis. Model Mech. 2017, 10, 955–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, Y.I.; Smith, J.E.; Goodman, D.S. Vitamin A and retinol-binding protein metabolism during fetal development in the rat. Am. J. Physiol. 1977, 233, E263–E272. [Google Scholar] [CrossRef] [PubMed]
- Sidell, N.; Altman, A.; Haussler, M.R.; Seeger, R.C. Effects of retinoic acid (RA) on the growth and phenotypic expression of several human neuroblastoma cell lines. Exp. Cell Res. 1983, 148, 21–30. [Google Scholar] [CrossRef]
- Sidell, N.; Sarafian, T.; Kelly, M.; Tsuchida, T.; Haussler, M. Retinoic acid-induced differentiation of human neuroblastoma: A cell variant system showing two distinct responses. Exp. Cell Biol. 1986, 54, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Wada, R.K.; Seeger, R.C.; Reynolds, C.P.; Alloggiamento, T.; Yamashiro, J.M.; Ruland, C.; Black, A.C.; Rosenblatt, J.D. Cell type-specific expression and negative regulation by retinoic acid of the human N-myc promoter in neuroblastoma cells. Oncogene 1992, 7, 711–717. [Google Scholar]
- Reynolds, C.P.; Kane, D.J.; Einhorn, P.A.; Matthay, K.K.; Crouse, V.L.; Wilbur, J.R.; Shurin, S.B.; Seeger, R.C. Response of neuroblastoma to retinoic acid in vitro and in vivo. Prog. Clin. Biol. Res. 1991, 366, 203–211. [Google Scholar]
- Ladenstein, R.; Potschger, U.; Pearson, A.D.J.; Brock, P.; Luksch, R.; Castel, V.; Yaniv, I.; Papadakis, V.; Laureys, G.; Malis, J.; et al. Busulfan and melphalan versus carboplatin, etoposide, and melphalan as high-dose chemotherapy for high-risk neuroblastoma (HR-NBL1/SIOPEN): An international, randomised, multi-arm, open-label, phase 3 trial. Lancet Oncol. 2017, 18, 500–514. [Google Scholar] [CrossRef]
- Tatsumi, N.; Miki, R.; Katsu, K.; Yokouchi, Y. Neurturin-GFRalpha2 signaling controls liver bud migration along the ductus venosus in the chick embryo. Dev. Biol. 2007, 307, 14–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clagett-Dame, M.; DeLuca, H.F. The role of vitamin A in mammalian reproduction and embryonic development. Ann. Rev. Nutr. 2002, 22, 347–381. [Google Scholar] [CrossRef] [PubMed]
- Marlétaz, F.; Holland, L.Z.; Laudet, V.; Schubert, M. Retinoic acid signaling and the evolution of chordates. Int. J. Biol. Sci. 2006, 2, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Ghyselinck, N.B.; Duester, G. Retinoic acid signaling pathways. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, D.H. Effect of diet during pregnancy upon the incidence of congenital hereditary diaphragmatic hernia in the rat; failure to produce cystic fibrosis of the pancreas by maternal vitamin A deficiency. Am. J. Pathol. 1949, 25, 163–185. [Google Scholar] [PubMed]
- Mendelsohn, C.; Lohnes, D.; Decimo, D.; Lufkin, T.; LeMeur, M.; Chambon, P.; Mark, M. Function of the retinoic acid receptors (RARs) during development (II). Multiple abnormalities at various stages of organogenesis in RAR double mutants. Development 1994, 120, 2749–2771. [Google Scholar]
- Greer, J.J.; Babiuk, R.P.; Thebaud, B. Etiology of congenital diaphragmatic hernia: The retinoid hypothesis. Pediatric Res. 2003, 53, 726–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurlin, P.J. N-Myc functions in transcription and development. Birth Defects Res. C Embryo Today 2005, 75, 340–352. [Google Scholar] [CrossRef]
- Zimmerman, K.A.; Yancopoulos, G.D.; Collum, R.G.; Smith, R.K.; Kohl, N.E.; Denis, K.A.; Nau, M.M.; Witte, O.N.; Toran-Allerand, D.; Gee, C.E.; et al. Differential expression of myc family genes during murine development. Nature 1986, 319, 780–783. [Google Scholar] [CrossRef]
- Stanton, B.R.; Perkins, A.S.; Tessarollo, L.; Sassoon, D.A.; Parada, L.F. Loss of N-myc function results in embryonic lethality and failure of the epithelial component of the embryo to develop. Genes Dev. 1992, 6, 2235–2247. [Google Scholar] [CrossRef]
- Hirvonen, H.; Makela, T.P.; Sandberg, M.; Kalimo, H.; Vuorio, E.; Alitalo, K. Expression of the myc proto-oncogenes in developing human fetal brain. Oncogene 1990, 5, 1787–1797. [Google Scholar]
- Hirvonen, H.; Sandberg, M.; Kalimo, H.; Hukkanen, V.; Vuorio, E.; Salmi, T.T.; Alitalo, T. The N-myc proto-oncogene and IGF-II growth factor mRNAs are expressed by distinct cells in human fetal kidney and brain. J. Cell Biol. 1989, 108, 1093–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mugrauer, G.; Alt, F.W.; Ekblom, P. N-myc proto-oncogene expression during organogenesis in the developing mouse as revealed by in situ hybridization. J. Cell Biol. 1988, 107, 1325–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grady, E.F.; Schwab, M.; Rosenau, W. Expression of N-myc and c-src during the development of fetal human brain. Cancer Res. 1987, 47, 2931–2936. [Google Scholar] [PubMed]
- Hirning, U.; Schmid, P.; Schulz, W.A.; Rettenberger, G.; Hameister, H. A comparative analysis of N-myc and c-myc expression and cellular proliferation in mouse organogenesis. Mech. Dev. 1991, 33, 119–125. [Google Scholar] [CrossRef]
- Moens, C.B.; Stanton, B.R.; Parada, L.F.; Rossant, J. Defects in heart and lung development in compound heterozygotes for two different targeted mutations at the N-myc locus. Development 1993, 119, 485–499. [Google Scholar]
- Moens, C.B.; Auerbach, A.B.; Conlon, R.A.; Joyner, A.L.; Rossant, J. A targeted mutation reveals a role for N-myc in branching morphogenesis in the embryonic mouse lung. Genes Dev. 1992, 6, 691–704. [Google Scholar] [CrossRef] [Green Version]
- Sawai, S.; Shimono, A.; Hanaoka, K.; Kondoh, H. Embryonic lethality resulting from disruption of both N-myc alleles in mouse zygotes. New Biol. 1991, 3, 861–869. [Google Scholar]
- Sawai, S.; Shimono, A.; Wakamatsu, Y.; Palmes, C.; Hanaoka, K.; Kondoh, H. Defects of embryonic organogenesis resulting from targeted disruption of the N-myc gene in the mouse. Development 1993, 117, 1445–1455. [Google Scholar]
- Charron, J.; Malynn, B.A.; Fisher, P.; Stewart, V.; Jeannotte, L.; Goff, S.P.; Robertson, E.J.; Alt, F.W. Embryonic lethality in mice homozygous for a targeted disruption of the N-myc gene. Genes Dev. 1992, 6, 2248–2257. [Google Scholar] [CrossRef]
- Cotterman, R.; Knoepfler, P.S. N-Myc regulates expression of pluripotency genes in neuroblastoma including lif, klf2, klf4, and lin28b. PLoS ONE 2009, 4, e5799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knoepfler, P.S.; Cheng, P.F.; Eisenman, R.N. N-myc is essential during neurogenesis for the rapid expansion of progenitor cell populations and the inhibition of neuronal differentiation. Genes Dev. 2002, 16, 2699–2712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, G.; Cui, H.; Shi, H.; Yang, L.; Ding, J.; Mao, L.; Maltese, W.A.; Ding, H.-F. MYCN promotes the expansion of Phox2B-positive neuronal progenitors to drive neuroblastoma development. Am. J. Pathol. 2009, 175, 856–866. [Google Scholar] [CrossRef] [Green Version]
- Thiele, C.J.; Reynolds, C.P.; Israel, M.A. Decreased expression of N-myc precedes retinoic acid-induced morphological differentiation of human neuroblastoma. Nature 1985, 313, 404–406. [Google Scholar] [CrossRef]
- Abemayor, E.; Sidell, N. Human neuroblastoma cell lines as models for the in vitro study of neoplastic and neuronal cell differentiation. Environ. Health Perspect 1989, 80, 3–15. [Google Scholar] [CrossRef]
- Buechner, J.; Henriksen, J.R.; Haug, B.H.; Tomte, E.; Flaegstad, T.; Einvik, C. Inhibition of mir-21, which is up-regulated during MYCN knockdown-mediated differentiation, does not prevent differentiation of neuroblastoma cells. Differentiation 2011, 81, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negroni, A.; Scarpa, S.; Romeo, A.; Ferrari, S.; Modesti, A.; Raschella, G. Decrease of proliferation rate and induction of differentiation by a MYCN antisense DNA oligomer in a human neuroblastoma cell line. Cell Growth Differ. 1991, 2, 511–518. [Google Scholar]
- Henriksen, J.R.; Haug, B.H.; Buechner, J.; Tomte, E.; Lokke, C.; Flaegstad, T.; Einvik, C. Conditional expression of retrovirally delivered anti-MYCN shRNA as an in vitro model system to study neuronal differentiation in MYCN-amplified neuroblastoma. BMC Dev. Biol. 2011, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. Hypoxia-inducible factors: Mediators of cancer progression and targets for cancer therapy. Trends Pharmacol. Sci. 2012, 33, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Pahlman, S.; Mohlin, S. Hypoxia and hypoxia-inducible factors in neuroblastoma. Cell Tissue Res. 2018, 372, 269–275. [Google Scholar] [CrossRef] [Green Version]
- Bhaskara, V.K.; Mohanam, I.; Rao, J.S.; Mohanam, S. Intermittent hypoxia regulates stem-like characteristics and differentiation of neuroblastoma cells. PLoS ONE 2012, 7, e30905. [Google Scholar] [CrossRef] [Green Version]
- Cimmino, F.; Pezone, L.; Avitabile, M.; Acierno, G.; Andolfo, I.; Capasso, M.; Iolascon, A. Inhibition of hypoxia inducible factors combined with all-trans retinoic acid treatment enhances glial transdifferentiation of neuroblastoma cells. Sci. Rep. 2015, 5, 11158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerlund, I.; Shi, Y.; Toskas, K.; Fell, S.M.; Li, S.; Surova, O.; Södersten, E.; Kogner, P.; Nyman, U.; Schlisio, S.; et al. Combined epigenetic and differentiation-based treatment inhibits neuroblastoma tumor growth and links HIF2alpha to tumor suppression. Proc. Natl. Acad. Sci. USA 2017, 114, E6137–E6146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmquist, L.; Jogi, A.; Pahlman, S. Phenotypic persistence after reoxygenation of hypoxic neuroblastoma cells. Int. J. Cancer 2005, 116, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Jogi, A.; Ora, I.; Nilsson, H.; Lindeheim, A.; Makino, Y.; Poellinger, L.; Axelson, H.; Påhlman, S. Hypoxia alters gene expression in human neuroblastoma cells toward an immature and neural crest-like phenotype. Proc. Natl. Acad. Sci. USA 2002, 99, 7021–7026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jogi, A.; Vallon-Christersson, J.; Holmquist, L.; Axelson, H.; Borg, A.; Pahlman, S. Human neuroblastoma cells exposed to hypoxia: Induction of genes associated with growth, survival, and aggressive behavior. Exp. Cell Res. 2004, 295, 469–487. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, M.C.; Saadoun, S.; Verkman, A.S. Aquaporins and cell migration. Pflug. Arch. Eur. J. Physiol. 2008, 456, 693–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saadoun, S.; Papadopoulos, M.C.; Hara-Chikuma, M.; Verkman, A.S. Impairment of angiogenesis and cell migration by targeted aquaporin-1 gene disruption. Nature 2005, 434, 786–792. [Google Scholar] [CrossRef] [PubMed]
- Van der Horst, I.W.; Rajatapiti, P.; van der Voorn, P.; van Nederveen, F.H.; Tibboel, D.; Rottier, R.; Reiss, I.; de Krijger, R.R. Expression of hypoxia-inducible factors, regulators, and target genes in congenital diaphragmatic hernia patients. Pediatric Dev. Pathol. 2011, 14, 384–390. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Biopsy | Location | Tissue | Immunohistochemical | FISH | MGMT-STP27 | Hughes Classification | Evans Classification | Evaluation |
|---|---|---|---|---|---|---|---|---|
| 1 | Thoracically herniated mass | Liver metastasis | Synaptophysin + Tyrosinhydroxylase + NB84 + | NMYC not amplified | Unmethylated | Grade 3 | Poorly differentiated, stromal-poor neuroblastoma | |
| 2 | Retroperitoneal | Primary tumor | Synaptophysin + Chromogranin A + Neurofilament + | NMYC not amplified | Stage 4-S | poorly differentiated, stromal-poor neuroblastoma | ||
| 3 | Retroperitoneal | Primary tumor | Chromogranin A + | NMYC not amplified | Stage 4-S | Poorly differentiated, stromal-poor neuroblastoma |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huo, Z.; Bilang, R.; Brantner, B.; von der Weid, N.; Holland-Cunz, S.G.; Gros, S.J. Perspective on Similarities and Possible Overlaps of Congenital Disease Formation—Exemplified on a Case of Congenital Diaphragmatic Hernia and Neuroblastoma in a Neonate. Children 2021, 8, 163. https://0-doi-org.brum.beds.ac.uk/10.3390/children8020163
Huo Z, Bilang R, Brantner B, von der Weid N, Holland-Cunz SG, Gros SJ. Perspective on Similarities and Possible Overlaps of Congenital Disease Formation—Exemplified on a Case of Congenital Diaphragmatic Hernia and Neuroblastoma in a Neonate. Children. 2021; 8(2):163. https://0-doi-org.brum.beds.ac.uk/10.3390/children8020163
Chicago/Turabian StyleHuo, Zihe, Remo Bilang, Benedikt Brantner, Nicolas von der Weid, Stefan G. Holland-Cunz, and Stephanie J. Gros. 2021. "Perspective on Similarities and Possible Overlaps of Congenital Disease Formation—Exemplified on a Case of Congenital Diaphragmatic Hernia and Neuroblastoma in a Neonate" Children 8, no. 2: 163. https://0-doi-org.brum.beds.ac.uk/10.3390/children8020163