
Macronutrient Intake in Pregnancy and Child Cognitive and Behavioural Outcomes

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
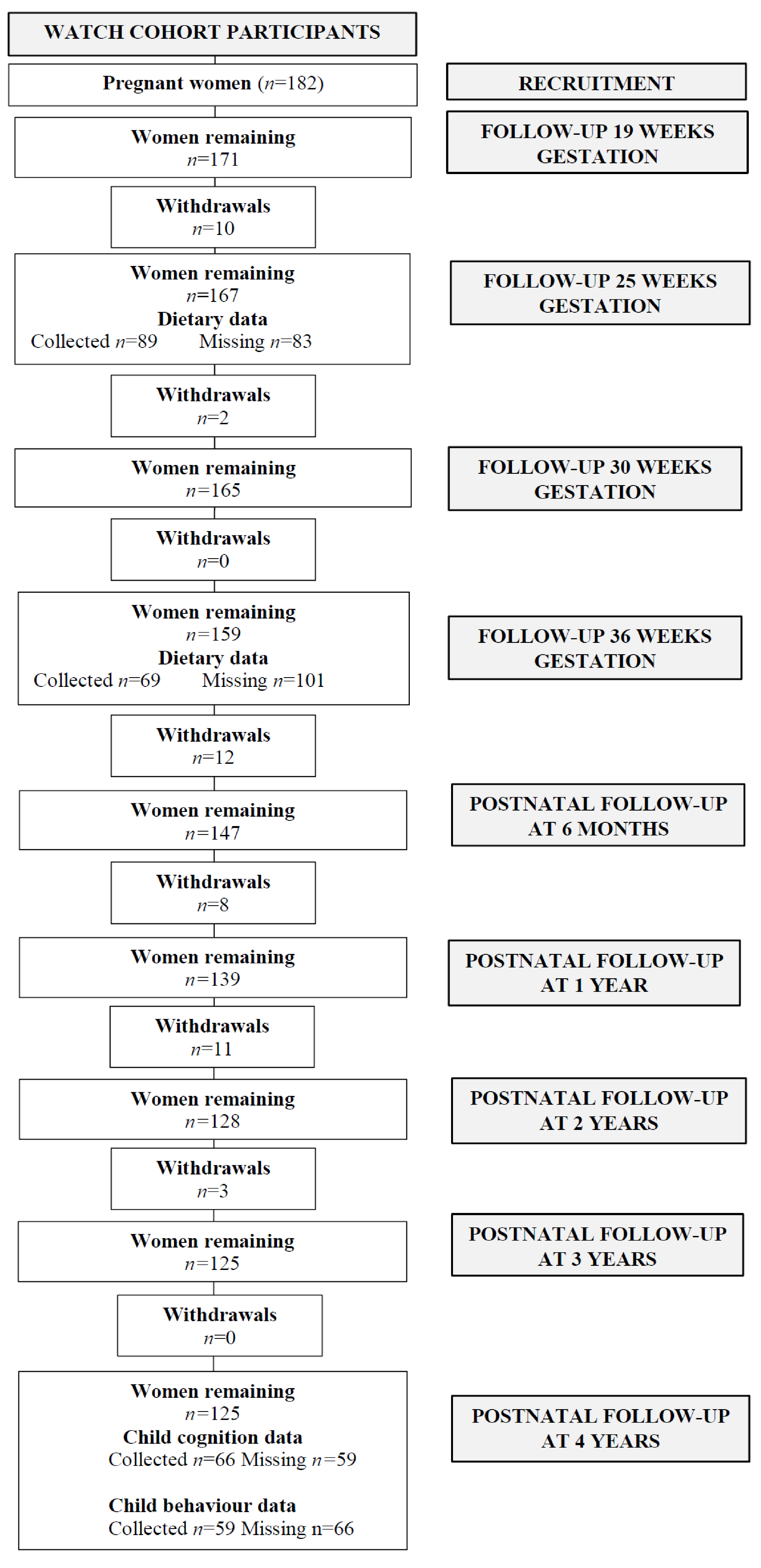
2.1. Study Population
2.2. Dietary Assessment
2.3. Dietary Analysis
2.4. Cognition and Behavioural Assessment
2.4.1. Cognition
2.4.2. Behaviour
2.4.3. Participant Characteristics
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gluckman, P.D.; Hanson, M.; Pinal, C. The developmental origins of adult disease. Matern. Child Nutr. 2005, 1, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Susser, E.; Neugebauer, R.; Hoek, H.W.; Brown, A.S.; Lin, S.; Labovitz, D.; Gorman, J.M. Schizophrenia After Prenatal Famine. Arch. Gen. Psychiatry 1996, 53, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Fox, S.E.; Levitt, P.; Nelson, C.A., III. How the Timing and Quality of Early Experiences Influence the Development of Brain Architecture. Child Dev. 2010, 81, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Dekaban, A.S.; Sadowsky, D. Changes in brain weights during the span of human life: Relation of brain weights to body heights and body weights. Ann. Neurol. 1978, 4, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Linderkamp, O.; Janus, L.; Linder, R.; Skoruppa, D.B. Time table of normal foetal brain development. Int. J. Prenat. Perinat. Psychol. Med. 2009, 21, 4–16. [Google Scholar]
- Prado, E.L.; Dewey, K.G. Nutrition and brain development in early life. Nutr. Rev. 2014, 72, 267–284. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.S.; Susser, E.S.; Lin, S.P.; Neugebauer, R.; Gorman, J.M. Increased Risk of Affective Disorders in Males after Second Trimester Prenatal Exposure to the Dutch Hunger Winter of 1944–45. Br. J. Psychiatry 1995, 166, 601–606. [Google Scholar] [CrossRef]
- Brown, A.S.; van Os, J.; Driessens, C.; Hoek, H.W.; Susser, E.S. Further evidence of relation between prenatal famine and major affective disorder. Am. J. Psychiatry 2000, 157, 190–195. [Google Scholar] [CrossRef]
- Rodriguez, A. Maternal pre-pregnancy obesity and risk for inattention and negative emotionality in children. J. Child Psychol. Psychiatry 2010, 51, 134–143. [Google Scholar] [CrossRef]
- Chen, Q.; Sjölander, A.; Långström, N.; Rodriguez, A.; Serlachius, E.; D’Onofrio, B.M.; Lichtenstein, P.; Larsson, H. Maternal pre-pregnancy body mass index and offspring attention deficit hyperactivity disorder: A population-based cohort study using a sibling-comparison design. Int. J. Epidemiol. 2014, 43, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.M.; Ou, J.-J.; Liu, L.; Zhang, D.; Zhao, J.-P.; Tang, S.Y. Association Between Maternal Obesity and Autism Spectrum Disorder in Offspring: A Meta-analysis. J. Autism Dev. Disord. 2016, 46, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Moody, L.; Chen, H.; Pan, Y.-X. Early-Life Nutritional Programming of Cognition—The Fundamental Role of Epigenetic Mechanisms in Mediating the Relation between Early-Life Environment and Learning and Memory Process. Adv. Nutr. 2017, 8, 337–350. [Google Scholar] [CrossRef]
- Xu, J.; He, G.; Zhu, J.; Zhou, X.; Clair, D.S.; Wang, T.; Xiang, Y.; Zhao, Q.; Xing, Q.; Liu, Y.; et al. Prenatal Nutritional Deficiency Reprogrammed Postnatal Gene Expression in Mammal Brains: Implications for Schizophrenia. Int. J. Neuropsychopharmacol. 2014, 18. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Lupu, D.S.; Niculescu, M.D. Perinatal α-linolenic acid availability alters the expression of genes related to memory and to epigenetic machinery, and the Mecp2 DNA methylation in the whole brain of mouse offspring. Int. J. Dev. Neurosci. 2014, 36, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Dominguez-Salas, P.; Moore, S.E.; Baker, M.S.; Bergen, A.W.; Cox, S.E.; Dyer, R.A.; Fulford, A.J.; Guan, Y.; Laritsky, E.; Silver, M.J.; et al. Maternal nutrition at conception modulates DNA methylation of human metastable epialleles. Nat. Commun. 2014, 5, 3746. [Google Scholar] [CrossRef]
- Dominguez-Salas, P.; E Moore, S.; Cole, D.; Da Costa, K.-A.; E Cox, S.; A Dyer, R.; Fulford, A.J.C.; Innis, S.M.; A Waterland, R.; Zeisel, S.H.; et al. DNA methylation potential: Dietary intake and blood concentrations of one-carbon metabolites and cofactors in rural African women. Am. J. Clin. Nutr. 2013, 97, 1217–1227. [Google Scholar] [CrossRef] [Green Version]
- Switkowski, K.M.; Jacques, P.F.; Must, A.; Kleinman, K.P.; Gillman, M.W.; Oken, E. Maternal protein intake during pregnancy and linear growth in the offspring. Am. J. Clin. Nutr. 2016, 104, 1128–1136. [Google Scholar] [CrossRef] [Green Version]
- Sloan, N.L.; Lederman, S.A.; Leighton, J.; Himes, J.H.; Rush, D. The effect of prenatal dietary protein intake on birth weight. Nutr. Res. 2001, 21, 129–139. [Google Scholar] [CrossRef]
- Moore, V.M.; Davies, M.J.; Willson, K.J.; Worsley, A.; Robinson, J.S. Dietary Composition of Pregnant Women Is Related to Size of the Baby at Birth. J. Nutr. 2004, 134, 1820–1826. [Google Scholar] [CrossRef] [Green Version]
- E Watson, P.; McDonald, B.W. The association of maternal diet and dietary supplement intake in pregnant New Zealand women with infant birthweight. Eur. J. Clin. Nutr. 2010, 64, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Hayward, C.E.; Greenwood, S.L.; Sibley, C.P.; Baker, P.N.; Challis, J.R.G.; Jones, R.L. Effect of maternal age and growth on placental nutrient transport: Potential mechanisms for teenagers’ predisposition to small-for-gestational-age birth? Am. J. Physiol. Metab. 2012, 302, E233–E242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iruloh, C.G.; D’Souza, S.W.; Fergusson, W.D.; Baker, P.N.; Sibley, C.P.; Glazier, J.D. Amino Acid Transport Systems β and A in Fetal T Lymphocytes in Intrauterine Growth Restriction and With Tumor Necrosis Factor-α Treatment. Pediatr. Res. 2008, 65, 51–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battaglia, F.C.; Regnault, T.R.H. Placental Transport and Metabolism of Amino Acids. Placenta 2001, 22, 145–161. [Google Scholar] [CrossRef] [PubMed]
- Blumfield, M.L.; Hure, A.J.; MacDonald-Wicks, L.K.; Smith, R.; Simpson, S.J.; Giles, W.B.; Raubenheimer, D.; Collins, C.E. Dietary balance during pregnancy is associated with fetal adiposity and fat distribution. Am. J. Clin. Nutr. 2012, 96, 1032–1041. [Google Scholar] [CrossRef] [Green Version]
- Burdge, G.C.; Dunn, R.L.; Wootton, S.A.; Jackson, A.A. Effect of reduced dietary protein intake on hepatic and plasma essential fatty acid concentrations in the adult female rat: Effect of pregnancy and consequences for accumulation of arachidonic and docosahexaenoic acids in fetal liver and brain. Br. J. Nutr. 2002, 88, 379–387. [Google Scholar] [CrossRef] [Green Version]
- Resnick, O.; Miller, M.; Forbes, W.; Hall, R.; Kemper, T.; Bronzino, J.; Morgane, P.J. Developmental protein malnutrition: Influences on the central nervous system of the rat. Neurosci. Biobehav. Rev. 1979, 3, 233–246. [Google Scholar] [CrossRef]
- Diáz-Cintra, S.; Cintra, L.; Galván, A.; Aguilar, A.; Kemper, T.; Morgane, P.J. Effects of prenatal protein deprivation on postnatal development of granule cells in the fascia dentata. J. Comp. Neurol. 1991, 310, 356–364. [Google Scholar] [CrossRef]
- Granados-Rojas, L.; Larriva-Sahd, J.; Cintra, L.; Gutiérrez-Ospina, G.; Rondán, A.; Díaz-Cintra, S. Prenatal protein malnutrition decreases mossy fibers-CA3 thorny excrescences asymmetrical synapses in adult rats. Brain Res. 2002, 933, 164–171. [Google Scholar] [CrossRef]
- Andrade, J.; Cadete-Leite, A.; Madeira, M.; Paula-Barbosa, M.M. Long-term low-protein diet reduces the number of hippocampal mossy fiber synapses. Exp. Neurol. 1991, 112, 119–124. [Google Scholar] [CrossRef]
- Reyes-Castro, L.A.; Padilla-Gómez, E.; Parga-Martínez, N.J.; Castro-Rodríguez, D.C.; Quirarte, G.L.; Díaz-Cintra, S.; Nathanielsz, P.W.; Zambrano, E. Hippocampal mechanisms in impaired spatial learning and memory in male offspring of rats fed a low-protein isocaloric diet in pregnancy and/or lactation. Hippocampus 2018, 28, 18–30. [Google Scholar] [CrossRef]
- Torres, N.; Bautista, C.J.; Tovar, A.R.; Ordáz, G.; Rodríguez-Cruz, M.; Ortiz, V.; Granados, O.; Nathanielsz, P.W.; Larrea, F.; Zambrano, E. Protein restriction during pregnancy affects maternal liver lipid metabolism and fetal brain lipid composition in the rat. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E270–E277. [Google Scholar] [CrossRef] [Green Version]
- Burdge, G.C.; Delange, E.; Dubois, L.; Dunn, R.L.; Hanson, M.A.; Jackson, A.A.; Calder, P.C. Effect of reduced maternal protein intake in pregnancy in the rat on the fatty acid composition of brain, liver, plasma, heart and lung phospholipids of the offspring after weaning. Br. J. Nutr. 2003, 90, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Castro, L.; Rodriguez, J.; Rodríguez-González, G.; Wimmer, R.; McDonald, T.; Larrea, F.; Nathanielsz, P.; Zambrano, E. Pre- and/or postnatal protein restriction in rats impairs learning and motivation in male offspring. Int. J. Dev. Neurosci. 2011, 29, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Furuse, T.; Miyake, K.; Kohda, T.; Kaneda, H.; Hirasawa, T.; Yamada, I.; Kushida, T.; Kashimura, M.; Kobayashi, K.; Ishino, F.; et al. Protein-restricted diet during pregnancy after insemination alters behavioral phenotypes of the progeny. Genes Nutr. 2017, 12, 1. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Castro, L.; Rodriguez, J.; Charco, R.; Bautista, C.; Larrea, F.; Nathanielsz, P.; Zambrano, E. Maternal protein restriction in the rat during pregnancy and/or lactation alters cognitive and anxiety behaviors of female offspring. Int. J. Dev. Neurosci. 2012, 30, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Jacka, F.N.; Ystrom, E.; Brantsaeter, A.L.; Karevold, E.; Roth, C.; Haugen, M.; Meltzer, H.M.; Schjolberg, S.; Berk, M. Maternal and Early Postnatal Nutrition and Mental Health of Offspring by Age 5 Years: A Prospective Cohort Study. J. Am. Acad. Child Adolesc. Psychiatry 2013, 52, 1038–1047. [Google Scholar] [CrossRef]
- Taylor, R.M.; Fealy, S.M.; Bisquera, A.; Smith, R.; Collins, C.E.; Evans, T.-J.; Hure, A.J. Effects of Nutritional Interventions during Pregnancy on Infant and Child Cognitive Outcomes: A Systematic Review and Meta-Analysis. Nutrients 2017, 9, 1265. [Google Scholar] [CrossRef] [Green Version]
- Larson, L.M.; Yousafzai, A.K. A meta-analysis of nutrition interventions on mental development of children under-two in low- and middle-income countries. Matern. Child Nutr. 2017, 13, e12229. [Google Scholar] [CrossRef]
- Bourre, J.-M. Effects of nutrients (in food) on the structure and function of the nervous system: Update on dietary requirements for brain. Part 1: Micronutrients. J. Nutr. Health Aging 2006, 10, 377–385. [Google Scholar]
- Paulson, O.B. Blood–brain barrier, brain metabolism and cerebral blood flow. Eur. Neuropsychopharmacol. 2002, 12, 495–501. [Google Scholar] [CrossRef]
- Hure, A.J.; Collins, C.E.; Giles, W.B.; Wright, I.; Smith, R. Protocol for the Women and Their Children’s Health (WATCH) Study: A Cohort of Pregnancy and Beyond. J. Epidemiology 2012, 22, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Hure, A.J.; Smith, R.; Collins, C.E. A recruiting failure turned success. BMC Health Serv. Res. 2008, 8, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hure, A.J.; Collins, C.E.; Giles, W.B.; Paul, J.W.; Smith, R. Greater Maternal Weight Gain During Pregnancy Predicts a Large but Lean Fetal Phenotype: A Prospective Cohort Study. Matern. Child Health J. 2012, 16, 1374–1384. [Google Scholar] [CrossRef] [PubMed]
- von Elm, E.; Altman, D.G.; Egger, M.; Pocock, S.J.; Gøtzsche, P.C.; Vandenbroucke, J.P. The Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) statement: Guidelines for reporting observational studies. Ann. Intern. Med. 2007, 147, 573–577. [Google Scholar] [CrossRef] [Green Version]
- Blumfield, M.L.; Nowson, C.; Hure, A.J.; Smith, R.; Simpson, S.J.; Raubenheimer, D.; MacDonald-Wicks, L.; Collins, C.E. Lower Protein-to-Carbohydrate Ratio in Maternal Diet is Associated with Higher Childhood Systolic Blood Pressure up to Age Four Years. Nutrients 2015, 7, 3078–3093. [Google Scholar] [CrossRef] [Green Version]
- Cancer Council Victoria. Dietary Questionnaire for Epidemiological Studies Version 2 (DQESv2); Australia Cancer Council: Melbourne, VIC, Australia, 2014. [Google Scholar]
- Hodge, A.; Patterson, A.J.; Brown, W.J.; Ireland, P.; Giles, G. The Anti Cancer Council of Victoria FFQ: Relative validity of nutrient intakes compared with weighed food records in young to middle-aged women in a study of iron supplementation. Aust. N. Z. J. Public Health 2000, 24, 576–583. [Google Scholar] [CrossRef]
- Wechsler, D. Wechsler Preschool and Primary Scale of Intelligence, 3rd ed.; Harvcourt Assessment Inc.: San Antonio, TX, USA, 2002. [Google Scholar]
- Wechsler, D. WPPSI-III: Technical and Interpretative Manual; The Psychological Corporation: New York, NY, USA, 2002. [Google Scholar]
- Wechsler, D. WPPSI-III Australian: Administration and Scoring Manual; Harcourt Assessment: Sydney, NSW, Australia, 2004. [Google Scholar]
- Achenbach, T.; Rescorla, L. Manual for ASEBA School-age Forms and Profiles. In Research Center for Children, Youth, and Families; University of Vermont: Burlington, UK, 2001. [Google Scholar]
- Rescorla, L.A.; Achenbach, T.M.; Ivanova, M.Y.; Harder, V.S.; Otten, L.; Bilenberg, N.; Bjarnadottir, G.; Capron, C.; De Pauw, S.S.W.; Dias, P.; et al. International Comparisons of Behavioral and Emotional Problems in Preschool Children: Parents’ Reports From 24 Societies. J. Clin. Child Adolesc. Psychol. 2011, 40, 456–467. [Google Scholar] [CrossRef] [Green Version]
- Willett, W.C.; Howe, G.R.; Kushi, L.H. Adjustment for total energy intake in epidemiologic studies. Am. J. Clin. Nutr. 1997, 65, 1220S–1228S. [Google Scholar] [CrossRef]
- Van den Broeck, J.; Willie, D.; Younger, N. The World Health Organization child growth standards: Expected implications for clinical and epidemiological research. Eur. J. Pediatr. 2009, 168, 247–251. [Google Scholar] [CrossRef]
- Meltzer, H.M.; Brantsaeter, A.L.; Ydersbond, T.A.; Alexander, J.; Haugen, M.; The MoBa Dietary Support Group. Methodological challenges when monitoring the diet of pregnant women in a large study: Experiences from the Norwegian Mother and Child Cohort Study (MoBa). Matern. Child Nutr. 2008, 4, 14–27. [Google Scholar] [CrossRef]
- Trumbo, P.; Schlicker, S.; Yates, A.A.; Poos, M. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein and Amino Acids. J. Am. Diet. Assoc. 2002, 102, 1621–1630. [Google Scholar] [CrossRef]
- Sharma, S.S.; Greenwood, D.C.; Simpson, N.A.B.; Cade, J. Is dietary macronutrient composition during pregnancy associated with offspring birth weight? An observational study. Br. J. Nutr. 2018, 119, 330–339. [Google Scholar] [CrossRef] [Green Version]
- Hrolfsdottir, L.; Schalkwijk, C.G.; Birgisdottir, B.E.; Gunnarsdottir, I.; Maslova, E.; Granström, C.; Strøm, M.; Olsen, S.; Halldorsson, T.I. Maternal diet, gestational weight gain, and inflammatory markers during pregnancy. Obesity 2016, 24, 2133–2139. [Google Scholar] [CrossRef]
- Savard, C.; Lemieux, S.; Weisnagel, S.J.; Fontaine-Bisson, B.; Gagnon, C.; Robitaille, J.; Morisset, A.-S. Trimester-Specific Dietary Intakes in a Sample of French-Canadian Pregnant Women in Comparison with National Nutritional Guidelines. Nutrients 2018, 10, 768. [Google Scholar] [CrossRef] [Green Version]
- National Health and Medical Research Council (NHMRC). Eat for Health: Australian Dietary Guidelines. 2013. Available online: https://www.eatforhealth.gov.au/sites/default/files/content/n55_australian_dietary_guidelines.pdf (accessed on 19 May 2021).
- Rizzo, T.; Metzger, B.E.; Burns, W.J.; Burns, K. Correlations between Antepartum Maternal Metabolism and Intelligence of Offspring. N. Engl. J. Med. 1991, 325, 911–916. [Google Scholar] [CrossRef]
- Xu, T.; Faleschini, S.; Rifas-Shiman, S.L.; Monthé-Drèze, C.; Oken, E.; Hivert, M.; Tiemeier, H. Maternal glucose tolerance in pregnancy and child cognitive and behavioural problems in early and mid-childhood. Paediatr. Périnat. Epidemiol. 2021, 35, 109–119. [Google Scholar] [CrossRef]
- Fraser, A.; Nelson, S.; Macdonald-Wallis, C.; Lawlor, D.A. Associations of Existing Diabetes, Gestational Diabetes, and Glycosuria with Offspring IQ and Educational Attainment: The Avon Longitudinal Study of Parents and Children. Exp. Diabetes Res. 2012, 2012, 963735. [Google Scholar] [CrossRef] [Green Version]
- Lovegrove, A.; Edwards, C.H.; De Noni, I.; Patel, H.; El, S.N.; Grassby, T.; Zielke, C.; Ulmius, M.; Nilsson, L.; Butterworth, P.J.; et al. Role of polysaccharides in food, digestion, and health. Crit. Rev. Food Sci. Nutr. 2017, 57, 237–253. [Google Scholar] [CrossRef] [Green Version]
- Australian Bureau of Statistics (ABS). Australian Health Survey: Nutrition First Results-Foods and Nutrients, 2011–2012; ABS: Canberra, Australia, 2014. [Google Scholar]
- Dundar, A.N.; Gocmen, D. Effects of autoclaving temperature and storing time on resistant starch formation and its functional and physicochemical properties. Carbohydr. Polym. 2013, 97, 764–771. [Google Scholar] [CrossRef] [Green Version]
- Muir, J.G.; O’Dea, K. Measurement of resistant starch: Factors affecting the amount of starch escaping digestion in vitro. Am. J. Clin. Nutr. 1992, 56, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Higgins, J.A.; Brown, M.A.; Storlien, L.H. Consumption of resistant starch decreases postprandial lipogenesis in white adipose tissue of the rat. Nutr. J. 2006, 5, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solinas, G.; Borén, J.; Dulloo, A.G. De novo lipogenesis in metabolic homeostasis: More friend than foe? Mol. Metab. 2015, 4, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Unger, R.H.; Clark, G.O.; Scherer, P.E.; Orci, L. Lipid homeostasis, lipotoxicity and the metabolic syndrome. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2010, 1801, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Opazo-Ríos, L.; Mas, S.; Marín-Royo, G.; Mezzano, S.; Gómez-Guerrero, C.; Moreno, J.A.; Egido, J. Lipotoxicity and Diabetic Nephropathy: Novel Mechanistic Insights and Therapeutic Opportunities. Int. J. Mol. Sci. 2020, 21, 2632. [Google Scholar] [CrossRef] [Green Version]
- Birt, D.F.; Boylston, T.; Hendrich, S.; Jane, J.-L.; Hollis, J.; Li, L.; McClelland, J.; Moore, S.; Phillips, G.J.; Rowling, M.; et al. Resistant Starch: Promise for Improving Human Health. Adv. Nutr. 2013, 4, 587–601. [Google Scholar] [CrossRef] [Green Version]
- Ashwar, B.A.; Gani, A.; Shah, A.; Wani, I.A.; Masoodi, F.A. Preparation, health benefits and applications of resistant starch-a review. Starch Stärke 2016, 68, 287–301. [Google Scholar] [CrossRef]
- Psichas, A.; Sleeth, M.L.; Murphy, K.G.; Brooks, L.; A Bewick, G.; Hanyaloglu, A.C.; A Ghatei, M.; Bloom, S.R.; Frost, G. The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents. Int. J. Obes. 2015, 39, 424–429. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, E.; Leonel, A.; Teixeira, L.; Silva, A.; Silva, J.; Pelaez, J.; Capettini, L.; Lemos, V.; Santos, R.; Alvarez-Leite, J. Butyrate impairs atherogenesis by reducing plaque inflammation and vulnerability and decreasing NFκB activation. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 606–613. [Google Scholar] [CrossRef]
- Yu, Y.; Patch, C.; Weston-Green, K.; Zhou, Y.; Zheng, K.; Huang, X.-F. Dietary Galacto-Oligosaccharides and Resistant Starch Protect Against Altered CB1 and 5-HT1A and 2A Receptor Densities in Rat Brain: Implications for Preventing Cognitive and Appetite Dysfunction During a High-Fat Diet. Mol. Nutr. Food Res. 2018, 62, e1800422. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Keenan, M.J.; Fernandez-Kim, S.O.; Pistell, P.J.; Ingram, D.K.; Li, B.; Raggio, A.M.; Shen, L.; Zhang, H.; McCutcheon, K.L.; et al. Dietary resistant starch improves selected brain and behavioral functions in adult and aged rodents. Mol. Nutr. Food Res. 2013, 57, 2071–2074. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Keenan, M.J.; Martin, R.J.; Tulley, R.T.; Raggio, A.M.; McCutcheon, K.L.; Zhou, J. Dietary Resistant Starch Increases Hypothalamic POMC Expression in Rats. Obesity 2009, 17, 40–45. [Google Scholar] [CrossRef] [Green Version]
- Gong, P.; Li, J.; Wang, J.; Lei, X.; Chen, D.; Zhang, K.; Zhang, W.; Zhen, A.; Gao, X.; Zhang, F. Variations in 5-HT2A influence spatial cognitive abilities and working memory. Can. J. Neurol. Sci. 2011, 38, 303–308. [Google Scholar] [CrossRef] [Green Version]
- Wingen, M.; Kuypers, K.; Ramaekers, J.G. Selective verbal and spatial memory impairment after 5-HT1A and 5-HT2A receptor blockade in healthy volunteers pre-treated with an SSRI. J. Psychopharmacol. 2007, 21, 477–485. [Google Scholar] [CrossRef]
- Yu, Y.; Wu, Y.; Patch, C.; Wu, Z.; Szabo, A.; Li, D.; Huang, X.-F. DHA prevents altered 5-HT1A, 5-HT2A, CB1 and GABAA receptor binding densities in the brain of male rats fed a high-saturated-fat diet. J. Nutr. Biochem. 2013, 24, 1349–1358. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Yu, Y.; Wu, Y.; Patch, C.; Szabo, A.; Huang, X.-F. Reduction of histamine H1 receptor binding induced by high-fat diet can be prevented by DHA and dietary fiber in specific brain areas of male rats. Brain Res. Bull. 2013, 97, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Hure, A.; Young, A.; Smith, R.; Collins, C. Diet and pregnancy status in Australian women. Public Health Nutr. 2009, 12, 853–861. [Google Scholar] [CrossRef] [Green Version]
- Ashman, A.M.; Collins, C.E.; Hure, A.J.; Jensen, M.; Oldmeadow, C. Maternal diet during early childhood, but not pregnancy, predicts diet quality and fruit and vegetable acceptance in offspring. Matern. Child Nutr. 2016, 12, 579–590. [Google Scholar] [CrossRef]
- Isaacs, E.; Oates, J.; ILSI Europe, a.i.s.b.l. Nutrition and cognition: Assessing cognitive abilities in children and young people. Eur. J. Nutr. 2008, 47, 4–24. [Google Scholar] [CrossRef]
- Deary, I.J.; Johnson, W.; Houlihan, L.M. Genetic foundations of human intelligence. Human Genet. 2009, 126, 215–232. [Google Scholar] [CrossRef] [Green Version]
- Ford, N.D.; Stein, A.D. Risk factors affecting child cognitive development: A summary of nutrition, environment, and maternal–child interaction indicators for sub-Saharan Africa. J. Dev. Orig. Health Dis. 2016, 7, 197–217. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Characteristics | ||
|---|---|---|
| Pregnant Women | Median (IQR) 1 | Range Difference |
| Maternal Age (y) | 29 (7) | 22.4 |
| Education | n | % |
| No formal qualification | 1 | 1.6 |
| Year 10 or equivalent | 10 | 16 |
| Year 12 or equivalent | 11 | 17 |
| Trade/apprenticeship | 2 | 3.1 |
| Certificate/diploma | 14 | 22 |
| University degree | 23 | 36 |
| Higher university degree | 3 | 4.7 |
| Missing | 0 | 0 |
| Household Weekly Income | n | % |
| No income | 0 | 0 |
| $AUD 1 1–299 | 4 | 6 |
| $AUD 1 300–699 | 13 | 20 |
| $AUD 1 700–999 | 13 | 20 |
| $AUD 1 1000 or more | 30 | 47 |
| Unsure | 4 | 6 |
| Missing | 0 | 0 |
| Marital Status | n | % |
| Never married | 20 | 32 |
| Married | 40 | 63 |
| Separated/divorced | 3 | 4.8 |
| Widowed | 0 | 0 |
| Missing | 1 | 1.6 |
| Maternal Smoking | n | % |
| Yes | 7 | 11 |
| No | 57 | 89 |
| Missing | 0 | 0 |
| Maternal Depression | n | % |
| Yes | 17 | 27 |
| No | 46 | 72 |
| Missing | 1 | 1.6 |
| Maternal Anxiety | n | % |
| Yes | 9 | 14 |
| No | 54 | 84 |
| Missing | 1 | 1.6 |
| Previous Live Births(>37 Weeks Gestation) | n | % |
| None | 34 | 53 |
| 1–2 | 26 | 41 |
| 3–4 | 4 | 6.2 |
| >5 | 0 | 0 |
| Missing | 0 | 0 |
| Nutrients | Daily NRVs | Daily Intake | % of Energy |
|---|---|---|---|
| Energy (KJ) | 7095.7 (5860.3, 8610.73) | n/a | |
| Protein (g) | 5–20 † | 78.5 (66.0, 101.7) | 19.4 (17.8, 21.1) |
| Total fat (g) | 20–35 † | 71.2 (58.1, 85.9) | 37.3 (34.2, 39.7) |
| SFA (g) | ≤10 † | 29.4 (23.2, 36.9) | 15.9 (13.1, 17.7) |
| PUFA (g) | 10.9 (8.1, 13.0) | 5.2 (4.4, 6.5) | |
| MUFA (g) | 24.5 (19.9, 29.4) | 12.7 (11.8, 13.9) | |
| Total carb. (g) | 45–65 † | 181.8 (153.7, 234.3) | 42.1 (39.5, 44.8) |
| Sugars (g) | ≤25% † | 89.9 (72.3, 110.6) | 19.9 (17.3, 22.0) |
| Starch (g) | 96.9 (79.6, 118.2) | 21.3 (20.0, 23.6) | |
| Fibre (g) | 25 (AI) § | 19.4 (15.1, 23.7) | 4.3 (3.7, 4.9) |
| P:C ratio (g) | 2.0 (1.0, 3.0) | n/a | |
| Energy-adjusted values (n = 64) | |||
| Energy (KJ) | 7317.0 (5984.2, 8706.3) | n/a | |
| Protein (g) | 5–20 † | 81.1 (69.2, 103.6) | 19.2 (17.7, 21.0) |
| Total fat (g) | 20–35 † | 72.3 (60.4, 87.4) | 37.3 (34.7, 40.0) |
| SFA (g) | ≤10 † | 30.3 (25.0, 38.8) | 16.0 (13.4, 17.8) |
| PUFA (g) | 11.2 (8.8, 13.1) | 5.2 (4.4, 6.6) | |
| MUFA (g) | 24.9 (20.9, 31.4) | 12.6 (11.8, 13.9) | |
| Total carb. (g) | 45–65 † | 186.1 (156.8, 237.6) | 42.0 (39.5, 44.7) |
| Total Sugars (g) | ≤25% † | 92.6 (74.3, 113.2) | 19.9 (16.6, 22.0) |
| Starch (g) | 99.2 (80.4, 120.9) | 21.3 (19.8, 23.7) | |
| Fibre (g) | 25 (AI) § | 20.9 (15.7, 24.3) | 4.2 (3.6, 4.9) |
| P:C ratio (g) | 2.0 (1.0, 3.0) | n/a |
| Variables 1 | Beta- Coefficient | 95% Confidence Interval | p-Value 2 | R-Value |
|---|---|---|---|---|
| Full Scale IQ | ||||
| Energy | −1.27 | −11.45 to 8.91 | 0.80 | 0.15 |
| Protein (% E) | 5.29 | −19.53 to 30.10 | 0.68 | 0.16 |
| Total fat (% E) | 14.01 | −9.19 to 37.20 | 0.23 | 0.18 |
| PUFA (% E) | 1.70 | −6.96 to 10.37 | 0.70 | 0.15 |
| CHO (% E) | −20.55 | −46.12 to 5.01 | 0.11 | 0.19 |
| P:C ratio | −0.73 | −7.20 to 5.75 | 0.82 | 0.15 |
| Protein (g) | 0.33 | −9.79 to 9.13 | 0.94 | 0.15 |
| PUFA (g) | 0.53 | −6.61 to 7.67 | 0.88 | 0.05 |
| Total sugars (g) | −0.51 | −8.50 to 7.49 | 0.90 | 0.15 |
| Starch (g) | −6.81 | −16.02 to 2.40 | 0.14 | 0.19 |
| Verbal IQ | ||||
| Energy | 3.79 | −9.73 to 17.31 | 0.58 | 0.06 |
| Protein (% E) | −1.51 | −34.61 to 31.59 | 0.93 | 0.05 |
| Total fat (% E) | 19.83 | −10.80 to 50.45 | 0.20 | 0.08 |
| PUFA (% E) | 0.20 | −11.21 to 11.61 | 0.97 | 0.05 |
| CHO (% E) | −19.41 | −53.73 to 14.91 | 0.26 | 0.07 |
| P:C ratio | −3.56 | −12.12 to 5.00 | 0.41 | 0.06 |
| Protein (g) | 3.05 | −9.51 to 15.62 | 0.63 | 0.05 |
| PUFA (g) | 1.98 | −7.45 to 11.41 | 0.68 | 0.05 |
| Total sugars (g) | 2.46 | −8.16 to 13.09 | 0.64 | 0.05 |
| Starch (g) | −2.23 | −14.75 to 10.27 | 0.72 | 0.05 |
| Performance IQ | ||||
| Energy | −5.75 | −17.14 to 5.64 | 0.32 | 0.20 |
| Protein (% E) | 14.14 | −13.68 to 41.96 | 0.31 | 0.20 |
| Total fat (% E) | 11.70 | −14.69 to 38.11 | 0.38 | 0.20 |
| PUFA (% E) | −0.39 | −10.19 to 9.42 | 0.94 | 0.19 |
| CHO (% E) | −24.67 | −53.48 to 4.14 | 0.09 | 0.23 |
| P:C ratio | 1.85 | −5.46 to 9.15 | 0.61 | 0.19 |
| Protein (g) | −2.91 | −13.56 to 7.74 | 0.59 | 0.19 |
| PUFA (g) | −3.09 | −11.12 to 4.93 | 0.44 | 0.20 |
| Total sugars (g) | −3.78 | −12.71 to 5.16 | 0.40 | 0.20 |
| Starch (g) | −11.02 | −21.19 to −0.84 | 0.03 | 0.26 |
| Processing Speed Composite | ||||
| Energy | 0.92 | −9.83 to 11.67 | 0.86 | 0.18 |
| Protein (% E) | 2.73 | −20.56 to 26.03 | 0.81 | 0.18 |
| Total fat (% E) | 6.14 | −16.24 to 28.52 | 0.58 | 0.19 |
| PUFA (% E) | −0.13 | −8.09 to 7.84 | 0.98 | 0.18 |
| CHO (% E) | −12.23 | −36.63 to 12.17 | 0.32 | 0.20 |
| P:C ratio | 3.17 | −2.72 to 9.05 | 0.28 | 0.20 |
| PUFA (g) | 0.31 | −6.90 to 7.52 | 0.93 | 0.18 |
| Protein (g) | 1.14 | −8.22 to 10.49 | 0.81 | 0.18 |
| Total sugars (g) | 0.80 | −6.86 to 8.47 | 0.83 | 0.18 |
| Starch (g) | −4.69 | −15.57 to 6.19 | 0.39 | 0.20 |
| General Language Composite | ||||
| Energy | 2.86 | −12.76 to 18.49 | 0.72 | 0.06 |
| Protein (% E) | 4.05 | −34.13 to 42.24 | 0.83 | 0.05 |
| Total fat (% E) | 22.65 | −12.72 to 58.01 | 0.20 | 0.09 |
| PUFA (% E) | 4.01 | −9.11 to 17.14 | 0.54 | 0.06 |
| CHO (% E) | −24.03 | −63.56 to 15.51 | 0.23 | 0.08 |
| P:C ratio | −1.88 | −11.82 to 8.05 | 0.71 | 0.06 |
| Protein (g) | 3.05 | −11.46 to 17.56 | 0.68 | 0.06 |
| PUFA (g) | 4.14 | −6.71 to 14.98 | 0.45 | 0.07 |
| Total sugars (g) | 3.04 | −9.21 to 15.30 | 0.62 | 0.06 |
| Starch (g) | −6.11 | −20.47 to 8.24 | 0.40 | 0.07 |
| Variables 1 | Beta- Coefficient | 95% Confidence Interval | p-Value 2 | R-Value |
|---|---|---|---|---|
| Total Problems Score | ||||
| Energy | 15.02 | −15.61 to 45.66 | 0.33 | 0.25 |
| Protein (% E) | 18.11 | −56.01 to 92.22 | 0.63 | 0.23 |
| PUFA (% E) | −19.98 | −47.31 to 7.34 | 0.15 | 0.27 |
| Total fat (% E) | −14.87 | −90.95 to 61.21 | 0.70 | 0.13 |
| CHO (% E) | 26.5887 | −57.25 to 110.42 | 0.53 | 0.24 |
| P:C ratio | 12.88 | −6.93 to 32.69 | 0.20 | 0.26 |
| Protein (g) | 15.55 | −12.71 to 43.81 | 0.27 | 0.25 |
| PUFA (g) | −4.60 | −26.18 to 16.98 | 0.67 | 0.23 |
| Total sugars (g) | 17.51 | −6.50 to 41.51 | 0.15 | 0.27 |
| Starch (g) | 12.0573 | −18.14 to 42.2 | 0.43 | 0.24 |
| Internalizing Broad Band Score | ||||
| Energy | −1.47 | −32.06 to 29.13 | 0.92 | 0.27 |
| Protein (% E) | 4.11 | −69.30 to 77.52 | 0.91 | 0.27 |
| Total fat (% E) | −10.67 | −85.89 to 64.54 | 0.78 | 0.27 |
| PUFA (% E) | −21.99 | −48.82 to 4.84 | 0.11 | 0.32 |
| CHO (% E) | 23.54 | −59.35 to 106.44 | 0.57 | 0.28 |
| P:C ratio | 11.54 | −8.10 to 31.18 | 0.24 | 0.30 |
| Protein (g) | −0.64 | −28.95 to 27.66 | 0.96 | 0.27 |
| PUFA (g) | −13.84 | −34.79 to 7.11 | 0.19 | 0.30 |
| Total sugars (g) | 4.56 | −19.69 to 28.81 | 0.71 | 0.27 |
| Starch (g) | −0.26 | −30.30 to 29.79 | 0.99 | 0.27 |
| Externalizing Broad Band Score | ||||
| Energy | 23.00 | −7.57 to 53.58 | 0.14 | 0.22 |
| Protein (% E) | 18.93 | −56.11 to 93.97 | 0.61 | 0.19 |
| Total fat (% E) | 1.68 | −75.49 to 78.86 | 0.97 | 0.18 |
| PUFA (% E) | −17.55 | −45.39 to 10.29 | 0.21 | 0.21 |
| CHO (% E) | −0.52 | −88.14 to 87.11 | 0.99 | 0.11 |
| P:C ratio | 11.48 | −8.67 to 31.63 | 0.26 | 0.21 |
| Protein (g) | 22.50 | −5.70 to 50.70 | 0.12 | 0.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taylor, R.M.; Blumfield, M.L.; Ashton, L.M.; Hure, A.J.; Smith, R.; Buckley, N.; Drysdale, K.; Collins, C.E. Macronutrient Intake in Pregnancy and Child Cognitive and Behavioural Outcomes. Children 2021, 8, 425. https://0-doi-org.brum.beds.ac.uk/10.3390/children8050425
Taylor RM, Blumfield ML, Ashton LM, Hure AJ, Smith R, Buckley N, Drysdale K, Collins CE. Macronutrient Intake in Pregnancy and Child Cognitive and Behavioural Outcomes. Children. 2021; 8(5):425. https://0-doi-org.brum.beds.ac.uk/10.3390/children8050425
Chicago/Turabian StyleTaylor, Rachael M., Michelle L. Blumfield, Lee M. Ashton, Alexis J. Hure, Roger Smith, Nick Buckley, Karen Drysdale, and Clare E. Collins. 2021. "Macronutrient Intake in Pregnancy and Child Cognitive and Behavioural Outcomes" Children 8, no. 5: 425. https://0-doi-org.brum.beds.ac.uk/10.3390/children8050425