
Human Milk Oligosaccharides: A Comprehensive Review towards Metabolomics

 ,
,  ,
,
Abstract
:1. Introduction
2. Human Milk Oligosaccharides: Structure and Composition
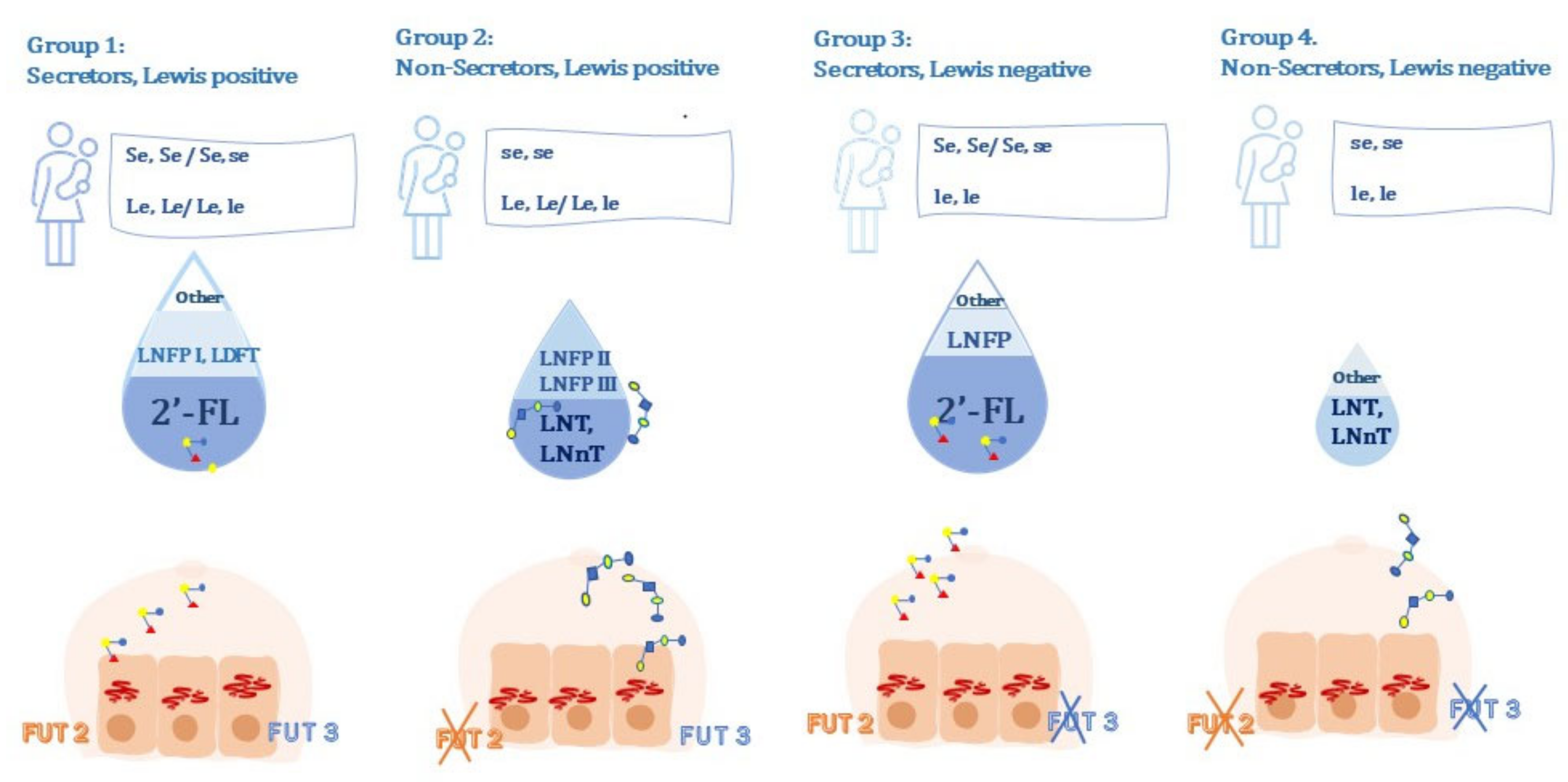
- Group 1: secretor, Lewis-positive, (Se, Le) (FUT2 active, FUT3 active);
- Group 2: non-secretor, Lewis-positive, (se, Le) (FUT2 inactive, FUT3 active);
- Group 3: secretor, Lewis-negative, (Se, le) (FUT2 active, FUT3 inactive);
- Group 4: non-secretor, Lewis-negative, (se, le) (FUT2 inactive, FUT3 inactive) [37].
3. Human Milk Oligosaccharides: Biological Functions behind Physiology
3.1. Prebiotic Effect
3.2. Maturation of the Intestinal Barrier
3.3. Brain Development
3.4. Development of the Immune System
3.5. Protection against Allergies
3.6. Protection from Infections
3.7. Protection against Necrotizing Enterocolitis
4. Metabolomics and Human Milk Oligosaccarides
4.1. Methods: Inclusion and Exclusion Criteria
4.2. Metabolomics and Human Milk Oligosaccharides: Review of the Literature’s Results
5. Human Milk Oligosaccharides Interindividual Variability: Is It Always about the Genes?
6. Formula Milk and HMOs Supplementation: Where Next?
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
List of Abbreviations
| HMOs | Human Milk Oligosaccharide |
| SCFAs | Short-Chain Fatty Acids |
| Gal | β-d-galactose |
| Glc | β-d-glucose |
| GlcNAc | β-d-N-acetyglucosamine |
| Fuc | α-l-fucose |
| Sia | sialic acid α-d-N-acetylneuraminic acid |
| LNnT | Lacto-N-neotetraose |
| LNT | Lacto-N-tetraose |
| LNT-II | Lacto-N-triose II |
| 2′-FL | 2′-fucosyllactose |
| 3-FL | 3-fucosyllactose |
| 3′-SL | 3′-sialyllactose |
| 6′-SL | 6-’sialyllactose |
| LNFP-I | Lacto-N-fucopentaose I |
| LNFP-II | Lacto-N-fucopentaose II |
| LNFP-III | Lacto-N-fucopentaose III |
| Th2 | T helper 2 lymphocyte |
| IL-4 | Interleukin-4 |
| IFN-γ | Interferon gamma |
| IgE | Immunoglobulin E |
| Th | T helper |
| DSLNT | disialyllacto-N-tetraose |
| LDFT | Lactodifucotetraose |
| LNDFH-I | Lacto-N-difucoesaose I |
| LNDFH-II | Lacto-N-difucohesaose II |
| DFLNH | difucosyllacto-N-hexaose |
| LNH | Lacto-N-hexaose |
| LNnFP-V | Lacto-N-neofucopentaose V |
| LNF-I | Lacto-N-fucopentaose |
| 3′-GL | 3′-galactosyllactose |
| 6′-GL | 6′-galactosyllactose |
| MAPK | Mitogen-Activated Protein Kinase |
| TRL | Toll Like Receptor |
| Sieglecs | sialic acid binding Immunoglobulin-like lectins |
| APC | Antigen Presenting Cells |
| TNF-α | Tumor Necrosis Factor alpha |
| NEC | necrotizing enterocolitis |
| HIV | human immunodeficiency virus |
| LC | lectin receptors |
References
- Victora, C.G.; Bahl, R.; Barros, A.; A França, G.V.; Horton, S.; Krasevec, J.; Murch, S.; Sankar, M.J.; Walker, N.; Rollins, N.C.; et al. Breastfeeding in the 21st century: Epidemiology, mechanisms, and lifelong effect. Lancet 2016, 387, 475–490. [Google Scholar] [CrossRef] [Green Version]
- Ballard, O.; Morrow, A.L. Human Milk Composition: Nutrients and bioactive factors. Pediatr. Clin. N. Am. 2012, 60, 49–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lessen, R.; Kavanagh, K. Position of the Academy of Nutrition and Dietetics: Promoting and Supporting Breastfeeding. J. Acad. Nutr. Diet. 2015, 115, 444–449. [Google Scholar] [CrossRef]
- Italianer, M.; Naninck, E.; Roelants, J.; Van Der Horst, G.; Reiss, I.; Goudoever, J.; Joosten, K.; Chaves, I.; Vermeulen, M. Circadian Variation in Human Milk Composition, a Systematic Review. Nutrients 2020, 12, 2328. [Google Scholar] [CrossRef]
- Martin, C.R.; Ling, P.-R.; Blackburn, G.L. Review of Infant Feeding: Key Features of Breast Milk and Infant Formula. Nutrients 2016, 8, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmeira, P.; Carneiro-Sampaio, M. Immunology of breast milk. Rev. Assoc. Med. Bras. 2016, 62, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Polonovski, M.; Montreuil, J. Etude chromatographique des polyosides de lait de femme [Chromatographic study of the polyosides of human milk]. C. R. Hebd. Seances Acad. Sci. 1954, 238, 2263–2264. [Google Scholar]
- Craft, K.M.; Townsend, S.D. Mother Knows Best: Deciphering the Antibacterial Properties of Human Milk Oligosaccharides. Acc. Chem. Res. 2019, 52, 760–768. [Google Scholar] [CrossRef] [Green Version]
- Duijts, L.; Jaddoe, V.W.V.; Hofman, A.; Moll, H.A. Prolonged and Exclusive Breastfeeding Reduces the Risk of Infectious Diseases in Infancy. Pediatrics 2010, 126, e18–e25. [Google Scholar] [CrossRef] [Green Version]
- Mudd, A.T.; Fleming, S.A.; Labhart, B.; Chichlowski, M.; Berg, B.M.; Donovan, S.M.; Dilger, R.N. Dietary Sialyllactose Influences Sialic Acid Concentrations in the Prefrontal Cortex and Magnetic Resonance Imaging Measures in Corpus Callosum of Young Pigs. Nutrients 2017, 9, 1297. [Google Scholar] [CrossRef] [Green Version]
- Doherty, A.M.; Lodge, C.J.; Dharmage, S.; Dai, X.; Bode, L.; Lowe, A.J. Human Milk Oligosaccharides and Associations With Immune-Mediated Disease and Infection in Childhood: A Systematic Review. Front. Pediatr. 2018, 6, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, L. Human Milk Oligosaccharides: Structure and Functions. Nestle Nutr. Inst. Workshop Ser. 2020, 94, 1–9. [Google Scholar] [CrossRef]
- Sprenger, G.A.; Baumgärtner, F.; Albermann, C. Production of human milk oligosaccharides by enzymatic and whole-cell microbial biotransformations. J. Biotechnol. 2017, 258, 79–91. [Google Scholar] [CrossRef]
- Thurl, S.; Munzert, M.; Henker, J.; Boehm, G.; Müller-Werner, B.; Jelinek, J.; Stahl, B. Variation of human milk oligosaccharides in relation to milk groups and lactational periods. Br. J. Nutr. 2010, 104, 1261–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, L.; Kuhn, L.; Kim, H.-Y.; Hsiao, L.; Nissan, C.; Sinkala, M.; Kankasa, C.; Mwiya, M.; Thea, D.; Aldrovandi, G.M. Human milk oligosaccharide concentration and risk of postnatal transmission of HIV through breastfeeding. Am. J. Clin. Nutr. 2012, 96, 831–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karczewski, K.J.; Snyder, M.P. Integrative omics for health and disease. Nat. Rev. Genet. 2018, 19, 299–310. [Google Scholar] [CrossRef]
- Casado, B.; Affolter, M.; Kussmann, M. OMICS-rooted studies of milk proteins, oligosaccharides and lipids. J. Proteom. 2009, 73, 196–208. [Google Scholar] [CrossRef]
- Koletzko, B. Interindividual variation of human milk metabolome. Am. J. Clin. Nutr. 2019, 110, 1–3. [Google Scholar] [CrossRef]
- Jenness, R. The composition of human milk. Semin Perinatol. 1979, 3, 225–239. [Google Scholar]
- Urashima, T.; Asakuma, S.; Leo, F.; Fukuda, K.; Messer, M.; Oftedal, O.T. The Predominance of Type I Oligosaccharides Is a Feature Specific to Human Breast Milk. Adv. Nutr. 2012, 3, 473S–482S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahl, B.; Thurl, S.; Zeng, J.; Karas, M.; Hillenkamp, F.; Steup, M.; Sawatzki, G. Oligosaccharides from Human Milk as Revealed by Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry. Anal. Biochem. 1994, 223, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Coppa, G.V.; Gabrielli, O.; Pierani, P.; Catassi, C.; Carlucci, A.; Giorgi, P.L. Changes in carbohydrate composition in human milk over 4 months of lactation. Pediatrics 1993, 91, 637–641. [Google Scholar]
- Townsend, R.R.; Hardy, M.R.; Lee, Y.C. Separation of oligosaccharides using high-performance anion-exchange chromatography with pulsed amperometric detection. Methods Enzymol. 1989, 179, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Urashima, T.; Hirabayashi, J.; Sato, S.; Kobata, A. Human Milk Oligosaccharides as Essential Tools for Basic and Application Studies on Galectins. Trends Glycosci. Glycotechnol. 2018, 30, SE51–SE65. [Google Scholar] [CrossRef] [Green Version]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef] [Green Version]
- Ray, C.; Kerketta, J.A.; Rao, S.; Patel, S.; Dutt, S.; Arora, K.; Pournami, F.; Bhushan, P. Human Milk Oligosaccharides: The Journey Ahead. Int. J. Pediatr. 2019, 2019, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soyyılmaz, B.; Mikš, M.H.; Röhrig, C.H.; Matwiejuk, M.; Meszaros-Matwiejuk, A.; Vigsnæs, L.K. The Mean of Milk: A Review of Human Milk Oligosaccharide Concentrations throughout Lactation. Nutrients 2021, 13, 2737. [Google Scholar] [CrossRef]
- Vandenplas, Y.; Berger, B.; Carnielli, V.P.; Ksiazyk, J.; Lagström, H.; Luna, M.S.; Migacheva, N.; Mosselmans, J.-M.; Picaud, J.-C.; Possner, M.; et al. Human Milk Oligosaccharides: 2′-Fucosyllactose (2′-FL) and Lacto-N-Neotetraose (LNnT) in Infant Formula. Nutrients 2018, 10, 1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thurl, S.; Henker, J.; Siegel, M.; Tovar, K.; Sawatzki, G. Detection of four human milk groups with respect to Lewis blood group dependent oligosaccharides. Glycoconj. J. 1997, 14, 795–799. [Google Scholar] [CrossRef]
- Kunz, C.; Rudloff, S.; Baier, W.; Klein, N.; Strobel, S. OLIGOSACCHARIDES INHUMANMILK: Structural, Functional, and Metabolic Aspects. Annu. Rev. Nutr. 2000, 20, 699–722. [Google Scholar] [CrossRef]
- Kunz, C.; Rudloff, S. Compositional Analysis and Metabolism of Human Milk Oligosaccharides in Infants. Nestle Nutr. Inst. Workshop Ser. 2017, 88, 137–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprenger, N.; Lee, L.Y.; De Castro, C.A.; Steenhout, P.; Thakkar, S.K. Longitudinal change of selected human milk oligosaccharides and association to infants’ growth, an observatory, single center, longitudinal cohort study. PLoS ONE 2017, 12, e0171814. [Google Scholar] [CrossRef] [Green Version]
- Blank, D.; Dotz, V.; Geyer, R.; Kunz, C. Human Milk Oligosaccharides and Lewis Blood Group: Individual High-Throughput Sample Profiling to Enhance Conclusions From Functional Studies. Adv. Nutr. 2012, 3, 440S–449S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akkerman, R.; Faas, M.M.; De Vos, P. Non-digestible carbohydrates in infant formula as substitution for human milk oligosaccharide functions: Effects on microbiota and gut maturation. Crit. Rev. Food Sci. Nutr. 2018, 59, 1486–1497. [Google Scholar] [CrossRef]
- Xu, Z.; Vo, L.; Macher, B.A. Structure-Function Analysis of Human 1,3-Fucosyltransferase. Amino acids involved in acceptor substrate specificity. J. Biol. Chem. 1996, 271, 8818–8823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erney, R.M.; Malone, W.T.; Skelding, M.B.; Marcon, A.A.; Kleman–Leyer, K.M.; O’Ryan, M.L.; Ruiz–Palacios, G.; Hilty, M.D.; Pickering, L.K.; Prieto, P. Variability of Human Milk Neutral Oligosaccharides in a Diverse Population. J. Pediatr. Gastroenterol. Nutr. 2000, 30, 181–192. [Google Scholar] [CrossRef]
- Van Leeuwen, S.S. Challenges and Pitfalls in Human Milk Oligosaccharide Analysis. Nutrients 2019, 11, 2684. [Google Scholar] [CrossRef] [Green Version]
- Nordgren, J.; Sharma, S.; Bucardo, F.; Nasir, W.; Günaydın, G.; Ouermi, D.; Nitiema, L.W.; Becker-Dreps, S.; Simpore, J.; Hammarström, L.; et al. Both Lewis and Secretor Status Mediate Susceptibility to Rotavirus Infections in a Rotavirus Genotype–Dependent Manner. Clin. Infect. Dis. 2014, 59, 1567–1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.; Koleva, P.; du Toit, E.; Geddes, D.T.; Munblit, D.; Prescott, S.L.; Eggesbø, M.; Johnson, C.C.; Wegienka, G.; Shimojo, N.; et al. The Milk Metabolome of Non-secretor and Lewis Negative Mothers. Front. Nutr. 2021, 7, 576966. [Google Scholar] [CrossRef]
- Samuel, T.M.; Binia, A.; De Castro, C.A.; Thakkar, S.K.; Billeaud, C.; Agosti, M.; Al-Jashi, I.; Costeira, M.J.; Marchini, G.; Martinez-Costa, C.; et al. Impact of maternal characteristics on human milk oligosaccharide composition over the first 4 months of lactation in a cohort of healthy European mothers. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Newburg, D.S.; Ruiz-Palacios, G.M.; Morrow, A.L. Human milk glycans protect infants against enteric pathogens. Annu. Rev. Nutr. 2005, 25, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Kunz, C.; Meyer, C.; Collado, M.C.; Geiger, L.; García-Mantrana, I.; Bertua-Ríos, B.; Martinez-Costa, C.; Borsch, C.; Rudloff, S. Influence of Gestational Age, Secretor, and Lewis Blood Group Status on the Oligosaccharide Content of Human Milk. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 789–798. [Google Scholar] [CrossRef]
- Van Leeuwen, S.S.; Stoutjesdijk, E.; Kate, G.A.T.; Schaafsma, A.; Dijck-Brouwer, J.; Muskiet, F.A.J.; Dijkhuizen, L. Regional variations in human milk oligosaccharides in Vietnam suggest FucTx activity besides FucT2 and FucT3. Sci. Rep. 2018, 8, 16790. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.B.; Odamaki, T.; Xiao, J.-Z. Insights into the reason of Human-Residential Bifidobacteria (HRB) being the natural inhabitants of the human gut and their potential health-promoting benefits. FEMS Microbiol. Rev. 2020, 44, 369–385. [Google Scholar] [CrossRef]
- Hundshammer, C.; Minge, O. In Love with Shaping You—Influential Factors on the Breast Milk Content of Human Milk Oligosaccharides and Their Decisive Roles for Neonatal Development. Nutrients 2020, 12, 3568. [Google Scholar] [CrossRef] [PubMed]
- De Leoz, M.L.A.; Kalanetra, K.M.; Bokulich, N.A.; Strum, J.S.; Underwood, M.A.; German, J.B.; Mills, D.A.; Lebrilla, C.B. Human Milk Glycomics and Gut Microbial Genomics in Infant Feces Show a Correlation between Human Milk Oligosaccharides and Gut Microbiota: A Proof-of-Concept Study. J. Proteome Res. 2014, 14, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borewicz, K.; Gu, F.; Saccenti, E.; Arts, I.C.; Penders, J.; Thijs, C.; van Leeuwen, S.; Lindner, C.; Nauta, A.; Van Leusen, E.; et al. Correlating Infant Fecal Microbiota Composition and Human Milk Oligosaccharide Consumption by Microbiota of 1-Month-Old Breastfed Infants. Mol. Nutr. Food Res. 2019, 63, e1801214. [Google Scholar] [CrossRef]
- LoCascio, R.G.; Desai, P.; Sela, D.A.; Weimer, B.; Mills, D.A. Broad Conservation of Milk Utilization Genes in Bifidobacterium longum subsp. infantis as Revealed by Comparative Genomic Hybridization. Appl. Environ. Microbiol. 2010, 76, 7373–7381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, M.A.E.; O’Neill, I.J.; Kujawska, M.; Javvadi, S.G.; Wijeyesekera, A.; Flegg, Z.; Chalklen, L.; Hall, L.J. Breast milk-derived human milk oligosaccharides promote Bifidobacterium interactions within a single ecosystem. ISME J. 2019, 14, 635–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thongaram, T.; Hoeflinger, J.L.; Chow, J.; Miller, M.J. Human milk oligosaccharide consumption by probiotic and human-associated bifidobacteria and lactobacilli. J. Dairy Sci. 2017, 100, 7825–7833. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, J.G.; Chain, F.; Martín, R.; Humaran, L.G.B.; Courau, S.; Langella, P. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb. Cell Factories 2017, 16, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondue, P.; Milani, C.; Arnould, E.; Ventura, M.; Daube, G.; Lapointe, G.; Delcenserie, V. Bifidobacterium mongoliense genome seems particularly adapted to milk oligosaccharide digestion leading to production of antivirulent metabolites. BMC Microbiol. 2020, 20, 1–12. [Google Scholar] [CrossRef]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; Reyes-Gavilán, C.G.D.L.; Salazar, N. Intestinal Short Chain Fatty Acids and their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [Green Version]
- Besten, G.D.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarr, A.J.; Galley, J.D.; Fisher, S.E.; Chichlowski, M.; Berg, B.M.; Bailey, M.T. The prebiotics 3′Sialyllactose and 6′Sialyllactose diminish stressor-induced anxiety-like behavior and colonic microbiota alterations: Evidence for effects on the gut–brain axis. Brain Behav. Immun. 2015, 50, 166–177. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; O’Riordan, M.X. Regulation of Bacterial Pathogenesis by Intestinal Short-Chain Fatty Acids. Adv. Appl. Microbiol. 2013, 85, 93–118. [Google Scholar] [CrossRef] [Green Version]
- Azagra-Boronat, I.; Massot-Cladera, M.; Mayneris-Perxachs, J.; Knipping, K.; Land, B.V.; Tims, S.; Stahl, B.; Garssen, J.; Franch, À.; Castell, M.; et al. Immunomodulatory and Prebiotic Effects of 2′-Fucosyllactose in Suckling Rats. Front. Immunol. 2019, 10, 1773. [Google Scholar] [CrossRef] [Green Version]
- Gerbe, F.; Jay, P. Intestinal tuft cells: Epithelial sentinels linking luminal cues to the immune system. Mucosal Immunol. 2016, 9, 1353–1359. [Google Scholar] [CrossRef] [PubMed]
- Cilieborg, M.S.; Sangild, P.T.; Jensen, M.L.; Østergaard, M.V.; Christensen, L.; Rasmussen, S.; Mørbak, A.L.; Jorgensen, C.B.; Bering, S.B. α1,2-Fucosyllactose Does Not Improve Intestinal Function or Prevent Escherichia coli F18 Diarrhea in Newborn Pigs. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 310–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mezoff, E.A.; Hawkins, J.A.; Ollberding, N.J.; Karns, R.; Morrow, A.L.; Helmrath, M.A. The human milk oligosaccharide 2′-fucosyllactose augments the adaptive response to extensive intestinal. Am. J. Physiol. Liver Physiol. 2016, 310, G427–G438. [Google Scholar] [CrossRef] [Green Version]
- Kuntz, S.; Rudloff, S.; Kunz, C. Oligosaccharides from human milk influence growth-related characteristics of intestinally transformed and non-transformed intestinal cells. Br. J. Nutr. 2008, 99, 462–471. [Google Scholar] [CrossRef] [Green Version]
- Hester, S.N.; Donovan, S.M. Individual and Combined Effects of Nucleotides and Human Milk Oligosaccharides on Proliferation, Apoptosis and Necrosis in a Human Fetal Intestinal Cell Line. Food Nutr Sci. 2012, 3, 1567–1576. [Google Scholar] [CrossRef] [Green Version]
- Holscher, H.D.; Davis, S.R.; Tappenden, K. Human Milk Oligosaccharides Influence Maturation of Human Intestinal Caco-2Bbe and HT-29 Cell Lines. J. Nutr. 2014, 144, 586–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holscher, H.; Bode, L.; Tappenden, K. Human Milk Oligosaccharides Influence Intestinal Epithelial Cell Maturation In Vitro. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.; Elderman, M.; Cheng, L.; De Haan, B.J.; Nauta, A.; De Vos, P. Modulation of Intestinal Epithelial Glycocalyx Development by Human Milk Oligosaccharides and Non-Digestible Carbohydrates. Mol. Nutr. Food Res. 2019, 63, e1900303. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Kong, C.; Walvoort, M.T.C.; Faas, M.M.; De Vos, P. Human Milk Oligosaccharides Differently Modulate Goblet Cells Under Homeostatic, Proinflammatory Conditions and ER Stress. Mol. Nutr. Food Res. 2019, 64, e1900976. [Google Scholar] [CrossRef] [Green Version]
- Chleilat, F.; Klancic, T.; Ma, K.; Schick, A.; Nettleton, J.E.; Reimer, R.A. Human Milk Oligosaccharide Supplementation Affects Intestinal Barrier Function and Microbial Composition in the Gastrointestinal Tract of Young Sprague Dawley Rats. Nutrients 2020, 12, 1532. [Google Scholar] [CrossRef]
- Berger, P.K.; Plows, J.F.; Jones, R.B.; Alderete, T.; Yonemitsu, C.; Poulsen, M.; Ryoo, J.H.; Peterson, B.S.; Bode, L.; Goran, M.I. Human milk oligosaccharide 2’-fucosyllactose links feedings at 1 month to cognitive development at 24 months in infants of normal and overweight mothers. PLoS ONE 2020, 15, e0228323. [Google Scholar] [CrossRef]
- Fleming, S.; Hauser, J.; Yan, J.; Donovan, S.; Wang, M.; Dilger, R. A Mediation Analysis to Identify Links between Gut Bacteria and Memory in Context of Human Milk Oligosaccharides. Microorganisms 2021, 9, 846. [Google Scholar] [CrossRef]
- Vázquez, E.; Barranco, A.; Ramírez, M.; Gruart, A.; Delgado-García, J.M.; Martínez-Lara, E.; Blanco, S.; Martín, M.J.; Castanys, E.; Buck, R.; et al. Effects of a human milk oligosaccharide, 2′-fucosyllactose, on hippocampal long-term potentiation and learning capabilities in rodents. J. Nutr. Biochem. 2015, 26, 455–465. [Google Scholar] [CrossRef]
- Vazquez, E.; Barranco, A.; Ramirez, M.; Gruart, A.; Delgado-Garcia, J.M.; Jimenez, M.L.; Buck, R.; Rueda, R. Dietary 2’-Fucosyllactose Enhances Operant Conditioning and Long-Term Potentiation via Gut-Brain Communication through the Vagus Nerve in Rodents. PLoS ONE 2016, 11, e0166070. [Google Scholar] [CrossRef]
- Fleming, S.; Mudd, A.T.; Hauser, J.; Yan, J.; Metairon, S.; Steiner, P.; Donovan, S.M.; Dilger, R.N. Human and Bovine Milk Oligosaccharides Elicit Improved Recognition Memory Concurrent With Alterations in Regional Brain Volumes and Hippocampal mRNA Expression. Front. Neurosci. 2020, 14, 770. [Google Scholar] [CrossRef] [PubMed]
- Wang, B. Molecular Mechanism Underlying Sialic Acid as an Essential Nutrient for Brain Development and Cognition. Adv. Nutr. 2012, 3, 465S–472S. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Yu, B.; Karim, M.; Hu, H.; Sun, Y.; McGreevy, P.; Petocz, P.; Held, S.; Brand-Miller, J. Dietary sialic acid supplementation improves learning and memory in piglets. Am. J. Clin. Nutr. 2007, 85, 561–569. [Google Scholar] [CrossRef] [Green Version]
- Oliveros, E.; Vázquez, E.; Barranco, A.; Ramírez, M.; Gruart, A.; Delgado-García, J.M.; Buck, R.; Rueda, R.; Martín, M.J. Sialic Acid and Sialylated Oligosaccharide Supplementation during Lactation Improves Learning and Memory in Rats. Nutrients 2018, 10, 1519. [Google Scholar] [CrossRef] [Green Version]
- Obelitz-Ryom, K.; Bering, S.B.; Overgaard, S.H.; Eskildsen, S.F.; Ringgaard, S.; Olesen, J.L.; Skovgaard, K.; Pankratova, S.; Wang, B.; Brunse, A.; et al. Bovine Milk Oligosaccharides with Sialyllactose Improves Cognition in Preterm Pigs. Nutrients 2019, 11, 1335. [Google Scholar] [CrossRef] [Green Version]
- Triantis, V.; Bode, L.; van Neerven, R. Immunological Effects of Human Milk Oligosaccharides. Front. Pediatr. 2018, 6, 190. [Google Scholar] [CrossRef] [PubMed]
- Engfer, M.B.; Stahl, B.; Finke, B.; Sawatzki, G.; Daniel, H. Human milk oligosaccharides are resistant to enzymatic hydrolysis in the upper gastrointestinal tract. Am. J. Clin. Nutr. 2000, 71, 1589–1596. [Google Scholar] [CrossRef] [Green Version]
- Kulinich, A.; Liu, L. Human milk oligosaccharides: The role in the fine-tuning of innate immune responses. Carbohydr. Res. 2016, 432, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.C.; Wang, M.; Donovan, S.M. The role of early life nutrition in the establishment of gastrointestinal microbial composition and function. Gut Microbes 2017, 8, 143–171. [Google Scholar] [CrossRef]
- Rapoport, E.M.; Kurmyshkina, O.V.; Bovin, N.V. Mammalian galectins: Structure, carbohydrate specificity, and functions. Biochemistry 2008, 73, 393–405. [Google Scholar] [CrossRef]
- Koliwer-Brandl, H.; Siegert, N.; Umnus, K.; Kelm, A.; Tolkach, A.; Kulozik, U.; Kuballa, J.; Cartellieri, S.; Kelm, S. Lectin inhibition assays for the analysis of bioactive milk sialoglycoconjugates. Int. Dairy J. 2011, 21, 413–420. [Google Scholar] [CrossRef]
- Nio-Kobayashi, J. Tissue- and cell-specific localization of galectins, β-galactose-binding animal lectins, and their potential functions in health and disease. Anat. Sci. Int. 2016, 92, 25–36. [Google Scholar] [CrossRef]
- Cerliani, J.P.; Stowell, S.R.; Mascanfroni, I.D.; Arthur, C.M.; Cummings, R.D.; Rabinovich, G.A. Expanding the Universe of Cytokines and Pattern Recognition Receptors: Galectins and Glycans in Innate Immunity. J. Clin. Immunol. 2010, 31, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Doha, K.S.U.; Kitova, E.N.; Kitov, P.I.; St-Pierre, Y.; Klassen, J.S. Human Milk Oligosaccharide Specificities of Human Galectins. Comparison of Electrospray Ionization Mass Spectrometry and Glycan Microarray Screening Results. Anal. Chem. 2017, 89, 4914–4921. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Liu, S.; Kling, D.; Leone, S.; Lawlor, N.; Huang, Y.; Feinberg, S.B.; Hill, D.; Newburg, D.S. The human milk oligosaccharide 2′-fucosyllactose modulates CD14 expression in human enterocytes, thereby attenuating LPS-induced inflammation. Gut 2014, 65, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Zuurveld, M.; Van Witzenburg, N.P.; Garssen, J.; Folkerts, G.; Stahl, B.; Land, B.V.; Willemsen, L.E.M. Immunomodulation by Human Milk Oligosaccharides: The Potential Role in Prevention of Allergic Diseases. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Bode, L.; Kunz, C.; Muhly-Reinholz, M.; Mayer, K.; Seeger, W.; Rudloff, S. Inhibition of monocyte, lymphocyte, and neutrophil adhesion to endothelial cells by human milk oligosaccharides. Thromb. Haemost. 2004, 92, 1402–1410. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Lawlor, N.; Newburg, D.S. Human Milk Components Modulate Toll-Like Receptor–Mediated Inflammation. Adv. Nutr. 2016, 7, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Kiewiet, M.B.G.; Groeneveld, A.; Nauta, A.; de Vos, P. Human Milk Oligosaccharides and Its Acid Hydrolysate LNT2 Show Immunomodulatory Effects via TLRs in a Dose and Structure-Dependent Way. J. Funct. Food 2019, 59, 184. [Google Scholar] [CrossRef]
- Thomas, P.G.; Carter, M.R.; Atochina, O.; Da’Dara, A.A.; Piskorska, D.; McGuire, E.; Harn, D.A. Maturation of Dendritic Cell 2 Phenotype by a Helminth Glycan Uses a Toll-Like Receptor 4-Dependent Mechanism. J. Immunol. 2003, 171, 5837–5841. [Google Scholar] [CrossRef] [Green Version]
- Goehring, K.C.; Marriage, B.J.; Oliver, J.S.; Wilder, J.A.; Barrett, E.G.; Buck, R.H. Similar to Those Who Are Breastfed, Infants Fed a Formula Containing 2′-Fucosyllactose Have Lower Inflammatory Cytokines in a Randomized Controlled Trial. J. Nutr. 2016, 146, 2559–2566. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Ouwehand, A.C.; Isolauri, E.; Hashimoto, H.; Benno, Y.; Salminen, S. Comparison of mucosal adhesion and species identification of bifidobacteria isolated from healthy and allergic infants. FEMS Immunol. Med Microbiol. 2001, 30, 43–47. [Google Scholar] [CrossRef]
- Walsh, C.; Lane, J.A.; van Sinderen, D.; Hickey, R.M. Human milk oligosaccharides: Shaping the infant gut microbiota and supporting health. J. Funct. Foods 2020, 72, 104074. [Google Scholar] [CrossRef] [PubMed]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Akay, H.K.; Tokman, H.B.; Hatipoglu, N.; Hatipoglu, H.; Siraneci, R.; Demirci, M.; Borsa, B.A.; Yuksel, P.; Karakullukcu, A.; Kangaba, A.A.; et al. The relationship between bifidobacteria and allergic asthma and/or allergic dermatitis: A prospective study of 0–3 years-old children in Turkey. Anaerobe 2014, 28, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Cukrowska, B.; Bierła, J.B.; Zakrzewska, M.; Klukowski, M.; Maciorkowska, E. The Relationship between the Infant Gut Microbiota and Allergy. The Role of Bifidobacterium breve and Prebiotic Oligosaccharides in the Activation of Anti-Allergic Mechanisms in Early Life. Nutrients 2020, 12, 946. [Google Scholar] [CrossRef] [Green Version]
- -Courtade, L.C.; Han, S.; Lee, S.-H.; Mian, F.M.; Buck, R.; Forsythe, P. Attenuation of food allergy symptoms following treatment with human milk oligosaccharides in a mouse model. Allergy 2015, 70, 1091–1102. [Google Scholar] [CrossRef]
- Verheijden, K.A.; Braber, S.; Leusink-Muis, T.; Thijssen, S.; Boon, L.; Kraneveld, A.D.; Garssen, J.; Folkerts, G.; Willemsen, L.E. Regulatory T Cell Depletion Abolishes the Protective Effect of Dietary Galacto-Oligosaccharides on Eosinophilic Airway Inflammation in House Dust Mite–Induced Asthma in Mice. J. Nutr. 2015, 146, 831–837. [Google Scholar] [CrossRef] [Green Version]
- Zehra, S.; Khambati, I.; Vierhout, M.; Mian, M.F.; Buck, R.; Forsythe, P. Human Milk Oligosaccharides Attenuate Antigen-Antibody Complex Induced Chemokine Release from Human Intestinal Epithelial Cell Lines. J. Food Sci. 2018, 83, 499–508. [Google Scholar] [CrossRef]
- Bachrach, V.R.G.; Schwarz, E.; Bachrach, L.R. Breastfeeding and the Risk of Hospitalization for Respiratory Disease in Infancy. Arch. Pediatr. Adolesc. Med. 2003, 157, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Lis-Kuberka, J.; Orczyk-Pawiłowicz, M. Sialylated Oligosaccharides and Glycoconjugates of Human Milk. The Impact on Infant and Newborn Protection, Development and Well-Being. Nutrients 2019, 11, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, H.; Zhu, L.; Faden, H.S. The milk-based diet of infancy and the gut microbiome. Gastroenterol. Rep. 2019, 7, 246–249. [Google Scholar] [CrossRef]
- Laucirica, D.R.; Triantis, V.; Schoemaker, R.; Estes, M.K.; Ramani, S. Milk Oligosaccharides Inhibit Human Rotavirus Infectivity in MA104 Cells. J. Nutr. 2017, 147, 1709–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comstock, S.S.; Li, M.; Wang, M.; Monaco, M.H.; Kuhlenschmidt, T.B.; Kuhlenschmidt, M.S.; Donovan, S.M. Dietary Human Milk Oligosaccharides but Not Prebiotic Oligosaccharides Increase Circulating Natural Killer Cell and Mesenteric Lymph Node Memory T Cell Populations in Noninfected and Rotavirus-Infected Neonatal Piglets. J. Nutr. 2017, 147, 1041–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, J.; Piskarev, V.E.; Xia, M.; Huang, P.; Jiang, X.; Likhosherstov, L.M.; Novikova, O.S.; Newburg, D.S.; Ratner, D.M. Identifying human milk glycans that inhibit norovirus binding using surface plasmon resonance. Glycobiology 2013, 23, 1491–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrow, A.L.; Ruiz-Palacios, G.M.; Jiang, X.; Newburg, D.S. Human-Milk Glycans That Inhibit Pathogen Binding Protect Breast-feeding Infants against Infectious Diarrhea. J. Nutr. 2005, 135, 1304–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newburg, D.S.; Ruiz-Palacios, G.M.; Altaye, M.; Chaturvedi, P.; Meinzen-Derr, J.; Guerrero, M.D.L.; Morrow, A.L. Innate protection conferred by fucosylated oligosaccharides of human milk against diarrhea in breastfed infants. Glycobiology 2003, 14, 253–263. [Google Scholar] [CrossRef]
- Puccio, G.; Alliet, P.; Cajozzo, C.; Janssens, E.; Corsello, G.; Sprenger, N.; Wernimont, S.; Egli, D.; Gosoniu, L.; Steenhout, P. Effects of Infant Formula With Human Milk Oligosaccharides on Growth and Morbidity: A Randomized Multicenter Trial. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 624–631. [Google Scholar] [CrossRef] [Green Version]
- Kwon, S.-J.; Na, D.H.; Kwak, J.H.; Douaisi, M.; Zhang, F.; Park, E.J.; Park, J.-H.; Youn, H.; Song, C.-S.; Kane, R.S.; et al. Nanostructured glycan architecture is important in the inhibition of influenza A virus infection. Nat. Nanotechnol. 2016, 12, 48–54. [Google Scholar] [CrossRef]
- Lucas, A.; Cole, T. Breast milk and neonatal necrotising enterocolitis. Lancet 1990, 336, 1519–1523. [Google Scholar] [CrossRef]
- Bode, L. Human Milk Oligosaccharides in the Prevention of Necrotizing Enterocolitis: A Journey From in vitro and in vivo Models to Mother-Infant Cohort Studies. Front. Pediatr. 2018, 6, 385. [Google Scholar] [CrossRef]
- Tanner, S.; Berryhill, T.F.; Ellenburg, J.L.; Jilling, T.; Cleveland, D.S.; Lorenz, R.G.; Martin, C.A. Pathogenesis of Necrotizing Enterocolitis: Modeling the innate immune response. Am. J. Pathol. 2014, 185, 4–16. [Google Scholar] [CrossRef] [Green Version]
- Gabrielli, O.; Zampini, L.; Galeazzi, T.; Padella, L.; Santoro, L.; Peila, C.; Giuliani, F.; Bertino, E.; Fabris, C.; Coppa, G.V. Preterm Milk Oligosaccharides During the First Month of Lactation. Pediatrics 2011, 128, e1520–e1531. [Google Scholar] [CrossRef]
- Jantscher-Krenn, E.; Zherebtsov, M.; Nissan, C.; Goth, K.; Guner, Y.S.; Naidu, N.; Choudhury, B.; Grishin, A.V.; Ford, H.R.; Bode, L. The human milk oligosaccharide disialyllacto-N-tetraose prevents necrotising enterocolitis in neonatal rats. Gut 2011, 61, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.Y.; Li, B.; Koike, Y.; Määttänen, P.; Miyake, H.; Cadete, M.; Johnson-Henry, K.C.; Botts, S.; Lee, C.; Abrahamsson, T.R.; et al. Human Milk Oligosaccharides Increase Mucin Expression in Experimental Necrotizing Enterocolitis. Mol. Nutr. Food Res. 2018, 63, e1800658. [Google Scholar] [CrossRef]
- Autran, C.A.; Kellman, B.P.; Kim, J.H.; Asztalos, E.; Blood, A.; Spence, E.C.H.; Patel, A.; Hou, J.; Lewis, N.; Bode, L. Human milk oligosaccharide composition predicts risk of necrotising enterocolitis in preterm infants. Gut 2017, 67, 1064–1070. [Google Scholar] [CrossRef] [PubMed]
- Masi, A.C.; Embleton, N.D.; Lamb, C.A.; Young, G.; Granger, C.L.; Najera, J.; Smith, D.P.; Hoffman, K.L.; Petrosino, J.F.; Bode, L.; et al. Human milk oligosaccharide DSLNT and gut microbiome in preterm infants predicts necrotising enterocolitis. Gut 2020. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, M.; Guo, H.; Yan, J.; Liu, F.; Chen, J.; Li, Y.; Ren, F. Human Milk Oligosaccharides Protect against Necrotizing Enterocolitis by Inhibiting Intestinal Damage via Increasing the Proliferation of Crypt Cells. Mol. Nutr. Food Res. 2019, 63, e1900262. [Google Scholar] [CrossRef]
- Zhang, W.; He-Yang, J.; Tu, W.; Zhou, X. Sialylated human milk oligosaccharides prevent intestinal inflammation by inhibiting toll like receptor 4/NLRP3 inflammasome pathway in necrotizing enterocolitis rats. Nutr. Metab. 2021, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, C.P.; Wipf, P.; Yamaguchi, Y.; Fulton, W.B.; Kovler, M.; Niño, D.F.; Zhou, Q.; Banfield, E.; Werts, A.D.; Ladd, M.R.; et al. The human milk oligosaccharides 2’-fucosyllactose and 6’-sialyllactose protect against the development of necrotizing enterocolitis by inhibiting toll-like receptor 4 signaling. Pediatr. Res. 2020, 89, 91–101. [Google Scholar] [CrossRef]
- Marincola, F.C.; Noto, A.; Caboni, P.; Reali, A.; Barberini, L.; Lussu, M.; Murgia, F.; Santoru, M.L.; Atzori, L.; Fanos, V. A metabolomic study of preterm human and formula milk by high resolution NMR and GC/MS analysis: Preliminary results. J. Matern. Neonatal Med. 2012, 25, 62–67. [Google Scholar] [CrossRef]
- Praticò, G.; Capuani, G.; Tomassini, A.; Baldassarre, M.E.; Delfini, M.; Miccheli, A. Exploring human breast milk composition by NMR-based metabolomics. Nat. Prod. Res. 2013, 28, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Totten, S.M.; Wu, L.D.; Parker, E.A.; Davis, J.C.C.; Hua, S.; Stroble, C.; Ruhaak, L.R.; Smilowitz, J.T.; German, J.B.; Lebrilla, C.B. Rapid-throughput glycomics applied to human milk oligosaccharide profiling for large human studies. Anal. Bioanal. Chem. 2014, 406, 7925–7935. [Google Scholar] [CrossRef] [PubMed]
- Totten, S.M.; Zivkovic, A.M.; Wu, S.; Ngyuen, U.; Freeman, S.L.; Ruhaak, L.R.; Darboe, M.K.; German, J.B.; Prentice, A.M.; Lebrilla, C.B. Comprehensive Profiles of Human Milk Oligosaccharides Yield Highly Sensitive and Specific Markers for Determining Secretor Status in Lactating Mothers. J. Proteome Res. 2012, 11, 6124–6133. [Google Scholar] [CrossRef]
- Smilowitz, J.T.; O’Sullivan, A.; Barile, D.; German, J.B.; Lönnerdal, B.; Slupsky, C.M. The Human Milk Metabolome Reveals Diverse Oligosaccharide Profiles. J. Nutr. 2013, 143, 1709–1718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Leeuwen, S.; Schoemaker, R.; Gerwig, G.J.; Kan, E.J.M.V.L.-V.; Dijkhuizen, L.; Kamerling, J.P. Rapid milk group classification by 1H NMR analysis of Le and H epitopes in human milk oligosaccharide donor samples. Glycobiology 2014, 24, 728–739. [Google Scholar] [CrossRef] [Green Version]
- Alderete, T.L.; Autran, C.; Brekke, B.E.; Knight, R.; Bode, L.; Goran, M.I.; Fields, D.A. Associations between human milk oligosaccharides and infant body composition in the first 6 mo of life. Am. J. Clin. Nutr. 2015, 102, 1381–1388. [Google Scholar] [CrossRef]
- Spevacek, A.R.; Smilowitz, J.T.; Chin, E.L.; Underwood, M.A.; German, J.B.; Slupsky, C.M. Infant Maturity at Birth Reveals Minor Differences in the Maternal Milk Metabolome in the First Month of Lactation. J. Nutr. 2015, 145, 1698–1708. [Google Scholar] [CrossRef] [Green Version]
- Sundekilde, U.K.; Downey, E.; O’Mahony, J.A.; O’Shea, C.-A.; Ryan, C.A.; Kelly, A.L.; Bertram, H.C. The Effect of Gestational and Lactational Age on the Human Milk Metabolome. Nutrients 2016, 8, 304. [Google Scholar] [CrossRef] [Green Version]
- Charbonneau, M.R.; O’Donnell, D.; Blanton, L.V.; Totten, S.M.; Davis, J.C.; Barratt, M.J.; Cheng, J.; Guruge, J.; Talcott, M.; Bain, J.R.; et al. Sialylated Milk Oligosaccharides Promote Microbiota-Dependent Growth in Models of Infant Undernutrition. Cell 2016, 164, 859–871. [Google Scholar] [CrossRef] [Green Version]
- McGuire, M.K.; Meehan, C.L.; McGuire, M.A.; Williams, J.E.; Foster, J.; Sellen, D.W.; Kamau-Mbuthia, E.W.; Kamundia, E.W.; Mbugua, S.; Moore, S.E.; et al. What’s normal? Oligosaccharide concentrations and profiles in milk produced by healthy women vary geographically. Am. J. Clin. Nutr. 2017, 105, 1086–1100. [Google Scholar] [CrossRef]
- Dessì, A.; Briana, D.; Corbu, S.; Gavrili, S.; Marincola, F.C.; Georgantzi, S.; Pintus, R.; Fanos, V.; Malamitsi-Puchner, A. Metabolomics of Breast Milk: The Importance of Phenotypes. Metabolites 2018, 8, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Gallego, C.; Morales, J.M.; Monleón, D.; Du Toit, E.; Kumar, H.; Linderborg, K.M.; Zhang, Y.; Yang, B.; Isolauri, E.; Salminen, S.; et al. Human Breast Milk NMR Metabolomic Profile across Specific Geographical Locations and Its Association with the Milk Microbiota. Nutrients 2018, 10, 1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saben, J.; Sims, C.; Abraham, A.; Bode, L.; Andres, A. Human Milk Oligosaccharide Concentrations and Infant Intakes Are Associated with Maternal Overweight and Obesity and Predict Infant Growth. Nutrients 2021, 13, 446. [Google Scholar] [CrossRef] [PubMed]
- Biddulph, C.; Holmes, M.; Kuballa, A.; Davies, P.; Koorts, P.; Carter, R.; Maher, J. Human Milk Oligosaccharide Profiles and Associations with Maternal Nutritional Factors: A Scoping Review. Nutrients 2021, 13, 965. [Google Scholar] [CrossRef]
- Sims, C.R.; Lipsmeyer, M.E.; Turner, D.E.; Andres, A. Human milk composition differs by maternal BMI in the first 9 months postpartum. Am. J. Clin. Nutr. 2020, 112, 548–557. [Google Scholar] [CrossRef]
- Azad, M.B.; Robertson, B.; Atakora, F.; Becker, A.B.; Subbarao, P.; Moraes, T.J.; Mandhane, P.J.; Turvey, S.E.; Lefebvre, D.L.; Sears, M.R.; et al. Human Milk Oligosaccharide Concentrations Are Associated with Multiple Fixed and Modifiable Maternal Characteristics, Environmental Factors, and Feeding Practices. J. Nutr. 2018, 148, 1733–1742. [Google Scholar] [CrossRef]
- Marriage, B.J.; Buck, R.H.; Goehring, K.C.; Oliver, J.S.; Williams, J.A. Infants Fed a Lower Calorie Formula With 2’FL Show Growth and 2’FL Uptake Like Breast-Fed Infants. J. Pediatr. Gastroenterol. Nutr. 2015, 61, 649–658. [Google Scholar] [CrossRef] [Green Version]
- Berger, B.; Porta, N.; Foata, F.; Grathwohl, D.; Delley, M.; Moine, D.; Charpagne, A.; Siegwald, L.; Descombes, P.; Alliet, P.; et al. Linking Human Milk Oligosaccharides, Infant Fecal Community Types, and Later Risk To Require Antibiotics. Mbio 2020, 11, e03196-19. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Authors, Year | Milk Sample | Technique | Results | Clinical Significance |
|---|---|---|---|---|
| Newburg et al. [108], 2004 | 93 healthy Mexican breast-feeding mothers (67 Lea−b+, 24 Lea−b−, 2 Lea+b−) | HPLC | ↑ LNF-I and 2′-FL in milk from Lea−b− mothers, ↑ LNF-II, LNF-III, 3-FL Lea−b+ mothers ↓ 2-linked fucosyloligosaccharides in milk consumed by infants with diharrea due to E. coli heat-stable enterotoxin (milk of non secretors mothers) | Different Lewis phenotypes correlate to different patterns of HMOs Non secretor genotype, results in lower amounts of 2′-FL and higher risk of diarrhea due to E. coli heat-stable enterotoxin |
| Thurl et al. [14], 2010 | 30 healthy Caucasian breast-feeding mothers (22 Lea−b+, 3 Lea−b−, 5 Lea+b−) | HPAEC | ↑ LNFP-II, LNDFH-II, 3-FL in in milk from Lea−b+ mothers No α1,4-fucosyloligosaccharides in milk from Lea−b− mothers No α1,2-fucosyl-oligosaccharides in milk from Lea+b− (Non-secretors) mothers Highest levels of LNT and LNH reached at 1 week postpartum; 50% decrease by 3 months postpartum | Different Lewis phenotypes correlates to different patterns of HMOs HMO concentrations vary during lactation, reaching their highest concentration during the first week of life |
| Gabrielli et al. [114], 2011 | 63 breast-feeding mothers who delivered preterm (25–30 w) | HPAEC | Higher total HMO concentration in milk from Group 1; reduction in total HMOs during the first month in all groups; 2′-FL, DFLNH, and LNFP main HMOs in Group 1; DFLNH, LNFP-II, and 3-FL main HMOs in Group 2; 2′-FL and LNFP-I main HMOs in Group 3; MFLNH-II and LNFP-III main HMOs in Group 4 | Different genotypes correlate to different patterns of HMOs (Group 1–4) HMO concentrations vary during lactation, with concentration decreasing more slowly in Group 1 |
| Totten et al. [125], 2012 | 60 Gambian breastfeeding mothers (27 secretors Lea−b+, 6 non secretors Lea+b-, 27 secretors Lea−b-, 10 non-secretors Lea−b-) | HPLC−CHIP/TOF MS | Prevalence of fucosylated HMOs in secretors; Lower amounts of fucosylated HMOs in non-secretor individuals | Secretor status influences HMO expression more than the Lewis status |
| Bode L. et al. [15], 2012 | 81 BM HIV positive that transmitted the virus vs. 86 BM HIV that did not transmitted the virus vs. 36 BM HIV negative | HPLC | ↑ non-3′-SL protection against HIV transmission | Good study population, support the hypothesis of the protective effect of some HMOs against HIV transmission |
| Smilowitz et al. [126], 2013 | 52 healthy women (34–38 GW) | 1H-NMR | ↑ 2′-FL, LDFT and Lacto-N-fucopentose in BM of mothers with secretor phenotype | Differentiation between the two phenotypes according to the type of HMOs detected |
| Praticò et al. [123], 2014 | 20 healthy women | 1H-NMR | Se+/Le+: fucosylated HMOs Se-/Le+: absence of alpha-1,2-fucosylated HMOs Se-/Le-: absence of LNFP-II, LNDFH-I and LNDFH-II | Metabolomics approach linked the phenotype (3 different classes) with HMO content of the milk Different HMOs could impact the infant gut microbiota differently |
| Van Leeuwen et al. [127], 2014 | 32 breast-feeding mothers | 1H-NMR | Absence of alpha-1-2-fucosylated HMOs in Non-secretors Abundance of α1-2 fucosylated HMOs (2′-FL) in Secretors | Different Se/Le genotypes correlate to different patterns of HMOs (Group 1–4) |
| Alderete et al. [128], 2015 | 31 mother–infant pairs | HPLC |
LNFP-II and DSLNT correlate with greater fat mass LNFP-I was inversely associated with infant weight and fat mass | Explores the correlation among HMO composition and infant growth and obesity during the first 6 months of life |
| Spevacek et al. [129], 2015 | 15 BM preterm vs. 13 BM preterm | 1H-NMR | ↑ 3-FL during the 28 days of lactation in both milk ↑ LNFP and LNnT in preterm C at the beginning of lactation | HMO production is finely regulated by the mammary gland Dynamic composition in both milk in the first month of lactation Preterm milk varies the most |
| Sundekilde et al. [130], 2016 | 15 BM preterm vs. 30 at term | 1H-NMR | ↑ Lacto-N-tetraose, LNDFH-I, 3′-SL, and 6′-SL in preterm | First study comparing preterm vs. at term milk in a lactation period (up to 14 weeks) Metabolomics linked to clinics These HMOs may be the cause of low incidence of NEC in breastfed preterms |
| Charbonneau et al. [131], 2016 | 88 Malawian breast-feeding mothers (69 secretors, 19 non-secretors) and their newborns (59 with severe stunting, 29 healthy) 215 Malawian breast-feeding mothers (155 secretors, 60 non-secretors) | LC-TOF MS | Among secretors, no significant differences in HMO profiles; Among non-secretors, fucosylated and sialylated HMOs were more abundant in milk from mothers who delivered healthy infants compared to those who delivered stunted children | Lower fucosylated and sialylated HMOs in milk of mothers with severely stunted infants Breast milk from non-secretor Malawian mothers might be less supportive for their children’s growth |
| McGuire et al. [132], 2017 | 410 BM healthy women from different geographical location | HPLC | ↑ 3-FL in Swedish mothers ↑ DSLNT in Gambian mothers | HMOs vary in different locations around the world |
| Dessì et al. [133], 2018 | 53 BM of healthy mothers | 1H-NMR | 2′-FL. LDFT. LNFP-I secretor phenotype 3-FL and LNFD III non -secretor phenotype | HMO content differentiated in secretor and non-secretor phenotype |
| Gomez-Gallego et al. [134], 2018 | 79 BM of healthy mothers from Finland, Spain, South Africa, China (vaginal birth vs. c-section) | 1H-NMR | ↑ LNFP-I and 2′-FL in Finland and Spain ↑ 3-FL and LNFP-III in South Africa and China | The effect of c-section on BM is different depending on the location Different HMO production among the geographical location may results in different roles for human milk in relation to infant health and development. |
| Samuel et al. [40], 2019 | 331 European mothers from Spain, France, Italy, Norway, Portugal, Romania and Sweden 290 of 331 also tested for ppBMI correlation (229 normal, 61 overweight) | LC-FLD H-NMR | Highest concentration of 2′-FL, LNFP-I and DFLNH in Group 3 and 1 Highest concentration of LNFP-II and LNnFP-V in Group 2 Highest concentration of LNFP-III and LNT in Group 4 | Different Se/Le genotypes correlate to different patterns of HMOs (Group 1–4) Lower concentrations of LNT, LNnT and LNFP-V and higher concentration of 3′-SL, 6′-GL in overweight women |
| Wang et al. [39], 2021 | 10 breast-feeding mothers | 1H-NMR | 2′-FL and LNFP-I in milk from Secretors, absent in non-secretors. LNFP-II, 3-FL, LDFT, and LNFP-III in milk of Lewis positive, absent in Lewis negative. Significant differences exist in the non-HMO milk metabolome of non-secretor, Lewis negative mothers compared to other phenotypes. | Different Se/Le genotypes correlate to different patterns of HMOs (Group 1–4) Mammary gland metabolome is globally influenced by the inactivity of FUT2 and FUT3: Se–Le– phenotype shows a different non-HMO metabolome compared to other phenotypes |
| Saben JL et al. [135], 2021 | 194 mother–child pairs (Mothers classified by BMI: 68 NW, 51 OW, 75 OB) 155 infants tested for daily HM intake | HPLC | Among non-secretors, significantly lower concentration of 3′-SL in OW compared to NW. Among secretors, lower concentration of LNT, LNnT in OW compared to NW; higher LNnT and 3-FL in OB women compared to NW and OW. | Composition in terms of HMOs might be influenced by maternal weight, especially in non-secretor women. Infants born to OB women consumed fewer LNH, DFLNH and other HMOs compare to those born to OW and NW mothers, while they presented the highest consumption of LNnT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corona, L.; Lussu, A.; Bosco, A.; Pintus, R.; Cesare Marincola, F.; Fanos, V.; Dessì, A. Human Milk Oligosaccharides: A Comprehensive Review towards Metabolomics. Children 2021, 8, 804. https://0-doi-org.brum.beds.ac.uk/10.3390/children8090804
Corona L, Lussu A, Bosco A, Pintus R, Cesare Marincola F, Fanos V, Dessì A. Human Milk Oligosaccharides: A Comprehensive Review towards Metabolomics. Children. 2021; 8(9):804. https://0-doi-org.brum.beds.ac.uk/10.3390/children8090804
Chicago/Turabian StyleCorona, Laura, Anna Lussu, Alice Bosco, Roberta Pintus, Flaminia Cesare Marincola, Vassilios Fanos, and Angelica Dessì. 2021. "Human Milk Oligosaccharides: A Comprehensive Review towards Metabolomics" Children 8, no. 9: 804. https://0-doi-org.brum.beds.ac.uk/10.3390/children8090804