
Impact of HLA-B27 and Disease Status on the Gut Microbiome of the Offspring of Ankylosing Spondylitis Patients

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview
2.2. Subjects
2.3. Processing of Fecal Samples
2.4. Sequencing and Analysis of 16S Ribosomal DNA from the Fecal Specimens
2.5. Genotyping and PRS Calculation
3. Results
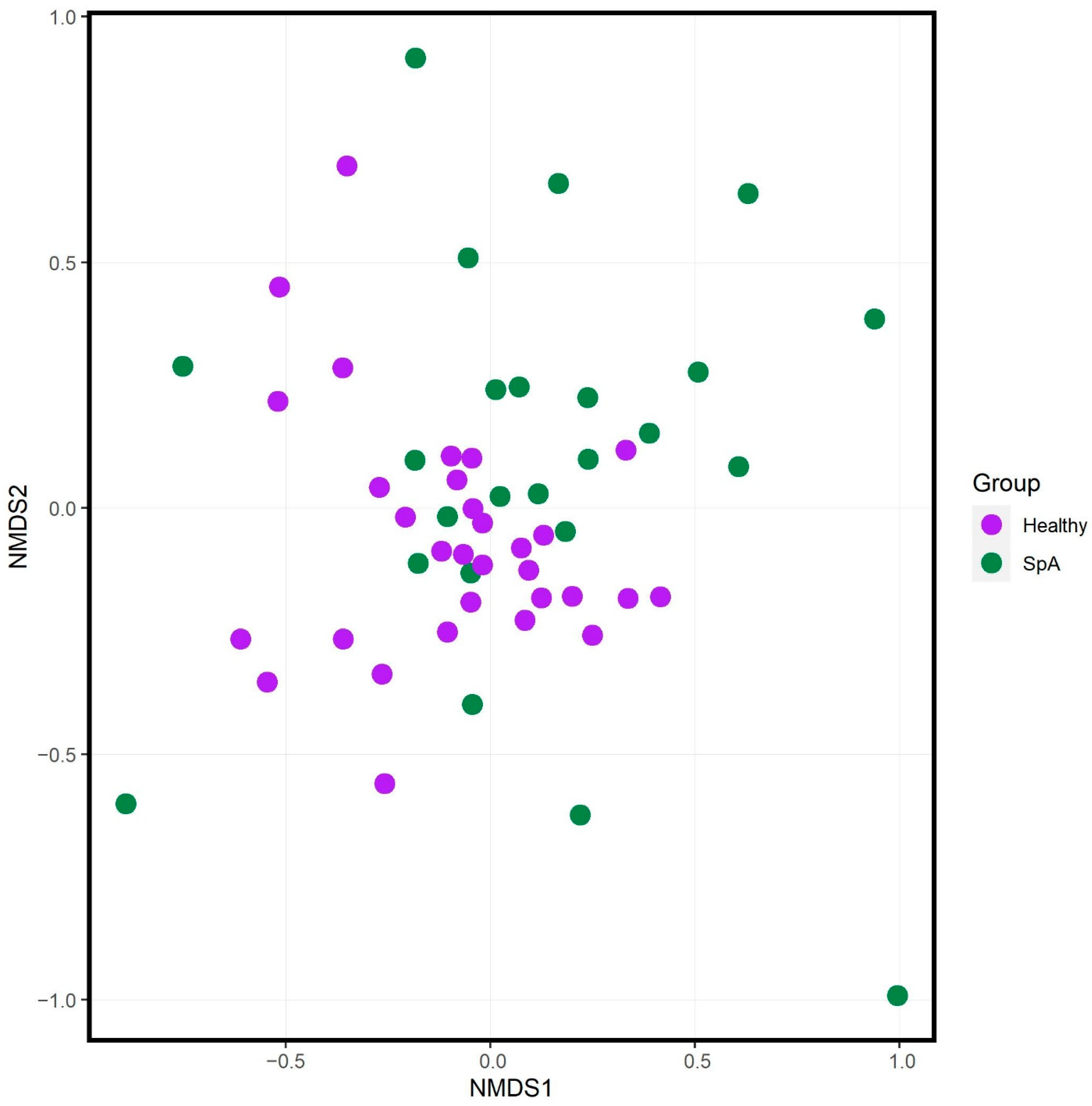
3.1. Offspring of Ankylosing Spondylitis Patients
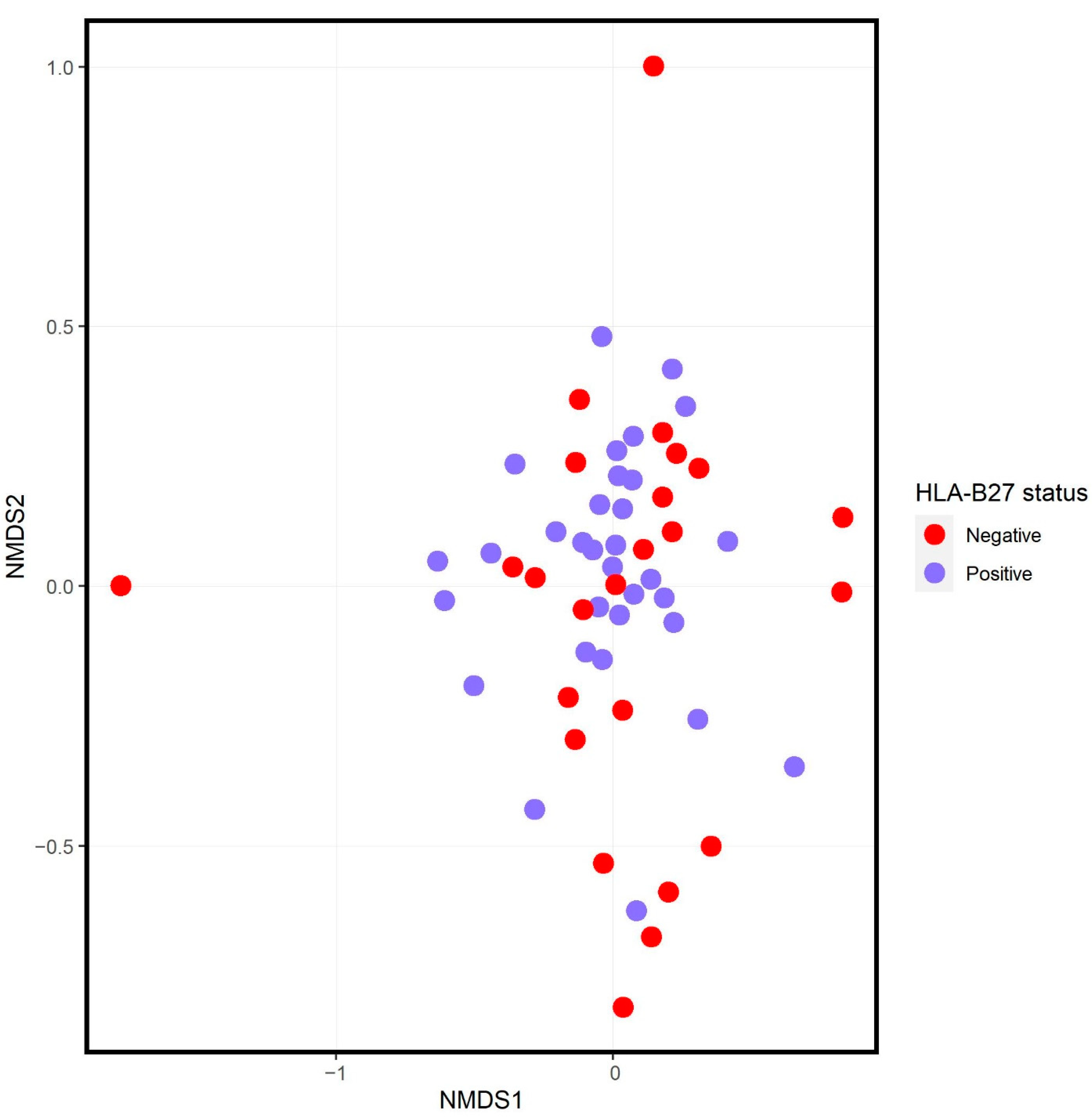
3.2. Comparison of HLA-B27+ SpA Subjects and Offspring
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reveille, J.D. Epidemiology of spondyloarthritis in North America. Am. J. Med. Sci. 2011, 341, 284–286. [Google Scholar] [CrossRef] [PubMed]
- Brewerton, D.A.; Hart, F.D.; Nicholls, A.; Caffrey, M.; James, D.C.; Sturrock, R.D. Ankylosing spondylitis and HL-A 27. Lancet 1973, 1, 904–907. [Google Scholar] [CrossRef]
- Srinivasalu, H.; Sikora, K.A.; Colbert, R.A. Recent Updates in Juvenile Spondyloarthritis. Rheum. Dis. Clin. N. Am. 2021, 47, 565–583. [Google Scholar] [CrossRef] [PubMed]
- Reveille, J.D.; Weisman, M.H. The epidemiology of back pain, axial spondyloarthritis and HLA-B27 in the United States. Am. J. Med. Sci. 2013, 345, 431–436. [Google Scholar] [CrossRef] [PubMed]
- van der Linden, S.M.; Valkenburg, H.A.; de Jongh, B.M.; Cats, A. The risk of developing ankylosing spondylitis in HLA-B27 positive individuals. A comparison of relatives of spondylitis patients with the general population. Arthritis Rheumatol. 1984, 27, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Australo-Anglo-American Spondyloarthritis Consortium (TASC); Reveille, J.D.; Sims, A.M.; Danoy, P.; Evans, D.M.; Leo, P.; Pointon, J.J.; Jin, R.; Zhou, X.; Bradbury, L.A.; et al. Genome-wide association study of ankylosing spondylitis identifies non-MHC susceptibility loci. Nat. Genet. 2010, 42, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Stoll, M.L.; Kumar, R.; Morrow, C.D.; Lefkowitz, E.J.; Cui, X.; Genin, A.; Cron, R.Q.; Elson, C.O. Altered microbiota associated with abnormal humoral immune responses to commensal organisms in enthesitis-related arthritis. Arthritis Res. Ther. 2014, 16, 486. [Google Scholar] [CrossRef]
- Costello, M.E.; Ciccia, F.; Willner, D.; Warrington, N.; Robinson, P.C.; Gardiner, B.; Marshall, M.; Kenna, T.J.; Triolo, G.; Brown, M.A. Intestinal dysbiosis in ankylosing spondylitis. Arthritis Rheumatol. 2015, 67, 686–691. [Google Scholar] [CrossRef]
- Tito, R.Y.; Cypers, H.; Joossens, M.; Varkas, G.; Van Praet, L.; Glorieus, E.; Van den Bosch, F.; De Vos, M.; Raes, J.; Elewaut, D. Brief Report: Dialister as a Microbial Marker of Disease Activity in Spondyloarthritis. Arthritis Rheumatol. 2017, 69, 114–121. [Google Scholar] [CrossRef]
- Breban, M.; Tap, J.; Leboime, A.; Said-Nahal, R.; Langella, P.; Chiocchia, G.; Furet, J.P.; Sokol, H. Faecal microbiota study reveals specific dysbiosis in spondyloarthritis. Ann. Rheum. Dis. 2017, 76, 1614–1622. [Google Scholar] [CrossRef]
- Aggarwal, A.; Sarangi, A.N.; Gaur, P.; Shukla, A.; Aggarwal, R. Gut microbiome in children with enthesitis-related arthritis in a developing country and the effect of probiotic administration. Clin. Exp. Immunol. 2017, 187, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Sternes, P.R.; Wang, M.; Song, J.; Morrison, M.; Li, T.; Zhou, L.; Wu, X.; He, F.; Zhu, J.; et al. Shotgun metagenomics reveals an enrichment of potentially cross-reactive bacterial epitopes in ankylosing spondylitis patients, as well as the effects of TNFi therapy upon microbiome composition. Ann. Rheum. Dis. 2020, 79, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, J.T.; Davey, M.P. Time for a gut check: Evidence for the hypothesis that HLA-B27 predisposes to ankylosing spondylitis by altering the microbiome. Arthritis Rheum. 2011, 63, 3195–3198. [Google Scholar] [CrossRef] [PubMed]
- van der Linden, S.; Valkenburg, H.A.; Cats, A. Evaluation of diagnostic criteria for ankylosing spondylitis. A proposal for modification of the New York criteria. Arthritis Rheum. 1984, 27, 361–368. [Google Scholar] [CrossRef]
- Petty, R.E.; Southwood, T.R.; Manners, P.; Baum, J.; Glass, D.N.; Goldenberg, J.; He, X.; Maldonado-Cocco, J.; Orozco-Alcala, J.; Prieur, A.M.; et al. International League of Associations for Rheumatology classification of juvenile idiopathic arthritis: Second revision, Edmonton, 2001. J. Rheumatol. 2004, 31, 390–392. [Google Scholar]
- Rudwaleit, M.; van der Heijde, D.; Landewe, R.; Listing, J.; Akkoc, N.; Brandt, J.; Braun, J.; Chou, C.T.; Collantes-Estevez, E.; Dougados, M.; et al. The development of Assessment of SpondyloArthritis international Society classification criteria for axial spondyloarthritis (part II): Validation and final selection. Ann. Rheum. Dis. 2009, 68, 777–783. [Google Scholar] [CrossRef]
- Stoll, M.L.; Weiss, P.F.; Weiss, J.E.; Nigrovic, P.A.; Edelheit, B.S.; Bridges, S.L., Jr.; Danila, M.I.; Spencer, C.H.; Punaro, M.G.; Schikler, K.; et al. Age and fecal microbial strain-specific differences in patients with spondyloarthritis. Arthritis Res. Ther. 2018, 20, 14. [Google Scholar] [CrossRef]
- Cary, S.G.; Blair, E.B. New Transport Medium for Shipment of Clinical Specimens. I. Fecal Specimens. J. Bacteriol. 1964, 88, 96–98. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Feranchuk, S.; Belkova, N.; Potapova, U.; Kuzmin, D.; Belikov, S. Evaluating the use of diversity indices to distinguish between microbial communities with different traits. Res. Microbiol. 2018, 169, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Bray, J.R.; Curtis, J.T. An ordination of upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Kelly, B.J.; Gross, R.; Bittinger, K.; Sherrill-Mix, S.; Lewis, J.D.; Collman, R.G.; Bushman, F.D.; Li, H. Power and sample-size estimation for microbiome studies using pairwise distances and Permanova. Bioinformatics 2015, 31, 2461–2468. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Svetnik, V.; Liaw, A.; Tong, C.; Culberson, J.C.; Sheridan, R.P.; Feuston, B.P. Random forest: A classification and regression tool for compound classification and QSAR modeling. J. Chem. Inf. Comput. Sci. 2003, 43, 1947–1958. [Google Scholar] [CrossRef]
- Li, Z.; Wu, X.; Leo, P.J.; De Guzman, E.; Akkoc, N.; Breban, M.; Macfarlane, G.J.; Mahmoudi, M.; Marzo-Ortega, H.; Anderson, L.K.; et al. Polygenic Risk Scores have high diagnostic capacity in ankylosing spondylitis. Ann. Rheum. Dis. 2021, 80, 1168–1174. [Google Scholar] [CrossRef]
- Jia, X.; Han, B.; Onengut-Gumuscu, S.; Chen, W.M.; Concannon, P.J.; Rich, S.S.; Raychaudhuri, S.; de Bakker, P.I. Imputing amino acid polymorphisms in human leukocyte antigens. PLoS ONE 2013, 8, e64683. [Google Scholar] [CrossRef]
- McCarthy, S.; Das, S.; Kretzschmar, W.; Delaneau, O.; Wood, A.R.; Teumer, A.; Kang, H.M.; Fuchsberger, C.; Danecek, P.; Sharp, K.; et al. A reference panel of 64,976 haplotypes for genotype imputation. Nat. Genet. 2016, 48, 1279–1283. [Google Scholar] [CrossRef]
- Klingberg, E.; Magnusson, M.K.; Strid, H.; Deminger, A.; Stahl, A.; Sundin, J.; Simren, M.; Carlsten, H.; Ohman, L.; Forsblad-d’Elia, H. A distinct gut microbiota composition in patients with ankylosing spondylitis is associated with increased levels of fecal calprotectin. Arthritis Res. Ther. 2019, 21, 248. [Google Scholar] [CrossRef] [PubMed]
- Asquith, M.; Sternes, P.R.; Costello, M.E.; Karstens, L.; Diamond, S.; Martin, T.M.; Li, Z.; Marshall, M.S.; Spector, T.D.; le Cao, K.A.; et al. HLA Alleles Associated with Risk of Ankylosing Spondylitis and Rheumatoid Arthritis Influence the Gut Microbiome. Arthritis Rheumatol. 2019, 71, 1642–1650. [Google Scholar] [CrossRef] [PubMed]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermudez-Humaran, L.G.; Gratadoux, J.J.; Blugeon, S.; Bridonneau, C.; Furet, J.P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef] [PubMed]
- Stebbings, S.; Munro, K.; Simon, M.A.; Tannock, G.; Highton, J.; Harmsen, H.; Welling, G.; Seksik, P.; Dore, J.; Grame, G.; et al. Comparison of the faecal microflora of patients with ankylosing spondylitis and controls using molecular methods of analysis. Rheumatology 2002, 41, 1395–1401. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhao, H.; Xiao, X.Y.; Chen, B.D.; Guo, R.J.; Wang, Q.; Chen, H.; Zhao, L.D.; Zhang, C.C.; Jiao, Y.H.; et al. Metagenomic profiling of the pro-inflammatory gut microbiota in ankylosing spondylitis. J. Autoimmun. 2020, 107, 102360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Han, R.; Zhang, X.; Fang, G.; Chen, J.; Li, J.; Xu, S.; Qian, L.; Chen, W.; Pan, F. Fecal microbiota in patients with ankylosing spondylitis: Correlation with dietary factors and disease activity. Clin. Chim. Acta Int. J. Clin. Chem. 2019, 497, 189–196. [Google Scholar] [CrossRef]
- Tejesvi, M.V.; Arvonen, M.; Kangas, S.M.; Keskitalo, P.L.; Pirttila, A.M.; Karttunen, T.J.; Vahasalo, P. Faecal microbiome in new-onset juvenile idiopathic arthritis. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2016, 35, 363–370. [Google Scholar] [CrossRef]
- Muller, P.H.; De Meij, T.G.; Westedt, M.; De Groot, E.F.; Allaart, C.F.; Brinkman, D.M.; Schonberg-Meinema, D.; van der Berg, M.; Van Suijlekom-Smit, L.W.; Van Rossum, M.A.; et al. Disturbance of Microbial Core Species in New-Onset Juvenile Idiopathic Arthritis. J. Pediatr. Infect. Dis. 2017, 12, 131–135. [Google Scholar]
- Wen, C.; Zheng, Z.; Shao, T.; Liu, L.; Xie, Z.; Le Chatelier, E.; He, Z.; Zhong, W.; Fan, Y.; Zhang, L.; et al. Quantitative metagenomics reveals unique gut microbiome biomarkers in ankylosing spondylitis. Genome Biol. 2017, 18, 142. [Google Scholar] [CrossRef]
- Chen, Z.; Qi, J.; Wei, Q.; Zheng, X.; Wu, X.; Li, X.; Liao, Z.; Lin, Z.; Gu, J. Variations in gut microbial profiles in ankylosing spondylitis: Disease phenotype-related dysbiosis. Ann. Transl. Med. 2019, 7, 571. [Google Scholar] [CrossRef]
- Vatanen, T.; Kostic, A.D.; d’Hennezel, E.; Siljander, H.; Franzosa, E.A.; Yassour, M.; Kolde, R.; Vlamakis, H.; Arthur, T.D.; Hamalainen, A.M.; et al. Variation in Microbiome LPS Immunogenicity Contributes to Autoimmunity in Humans. Cell 2016, 165, 842–853. [Google Scholar] [CrossRef] [PubMed]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Di Paola, M.; Ramazzotti, M.; Albanese, D.; Pieraccini, G.; Banci, E.; Miglietta, F.; Cavalieri, D.; Lionetti, P. Diet, Environments, and Gut Microbiota. A Preliminary Investigation in Children Living in Rural and Urban Burkina Faso and Italy. Front. Microbiol. 2017, 8, 1979. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | Value |
|---|---|
| n | 31 |
| Demographics | |
| Male Sex | 23 (74.2%) |
| Age (years) | 45.7 ± 7.1 |
| Race | |
| White | 20 (64.5%) |
| Asian | 8 (25.8%) |
| American Indian | 1 (3.2%) |
| Multiracial | 2 (6.5%) |
| Hispanic ethnicity | 3 (9.7%) |
| Treatment | |
| NSAIDs | 18 (58.1%) |
| Methotrexate | 1 (3.2%) |
| Etanercept | 5 (16.1%) |
| TNFi mAb | 11 (35.5%) |
| Secukinumab | 1 (3.2%) |
| Body mass index (mg/kg2 ± SD) | 25.5 ± 4.0 |
| BASDAI | 2.5 ± 2.3 |
| BASFI | 1.7 ± 2.4 |
| ASDAS-CRP | 1.8 ± 1.0 |
| Feature | HLA-B27 Negative | HLA-B27 Positive |
|---|---|---|
| n | 24 | 32 |
| Demographics | ||
| Male Sex | 12 (50%) | 12 (38%) |
| Age (years) | 10.5 ± 4.0 | 10.7 ± 3.7 |
| Race | ||
| White | 11 (46%) | 11 (34%) |
| Asian | 4 (17%) | 4 (12%) |
| American Indian | 0 | 2 (6.2) |
| Multiracial | 9 (38%) | 15 (47%) |
| Hispanic ethnicity | 6 [25] | 5 (16%) |
| NSAID usage | 2 (8.3%) | 1 (3.1%) |
| Body mass index (mg/kg2 ± SD) | 17.5 ± 3.2 | 18.0 ± 3.8 |
| Organism | BaseMean | LFC | LFCSE | padj |
|---|---|---|---|---|
| Blautia | 2539 | −1.91 | 0.40 | 6.13 × 10−5 |
| Coprococcus | 1146 | −1.35 | 0.38 | 0.005 |
| Blautia obeum | 1142 | −2.17 | 0.55 | 0.001 |
| Blautia | 743 | −1.31 | 0.42 | 0.016 |
| Faecalibacterium prausnitzii | 611 | 0.72 | 0.26 | 0.034 |
| Feature | Offspring | SpA |
|---|---|---|
| n | 31 | 24 |
| Demographics | ||
| Male Sex | 12 (38.7%) | 15 (62%) |
| Age (years) | 10.6 ± 3.7 | 13.5 ± 2.9 |
| Race | ||
| White | 11 (36%) | 17 (71%) |
| Asian | 4 (13%) | 1 (4.2%) |
| Black | 0 | 4 (17%) |
| American Indian | 2 (6.5%) | 0 |
| Multiracial | 14 (45%) | 1 (4.2%) |
| Unknown | 0 | 1 (4.2%) |
| Hispanic ethnicity | 5 (16.1%) | 0 |
| NSAID usage | 0 | 7 (29.2%) |
| Sacroiliitis | 0 | 17 (71%) |
| Enthesitis | 0 | 11 (46%) |
| Acute anterior uveitis | 0 | 0 |
| Body mass index (mg/kg2 ± SD) | 17.8 ± 3.7 | 18.1 ± 3.8 |
| Organism | BaseMean | LFC | LFCSE | padj |
|---|---|---|---|---|
| Bacteroides | 3233 | 1.27 | 0.47 | 0.047 |
| Blautia | 1803 | 1.47 | 0.35 | <0.001 |
| Bacteroides ovatus | 1132 | 1.12 | 0.40 | 0.040 |
| Faecalibacterium prausnitzii | 618 | −1.19 | 0.38 | 0.018 |
| Escherichia coli | 277 | 2.59 | 0.78 | 0.011 |
| Parabacteroides | 274 | 2.92 | 1.03 | 0.040 |
| Bifidobacterium | 128 | −2.12 | 0.59 | 0.004 |
| Coprococcus | 109 | −1.95 | 0.51 | 0.002 |
| Ruminococcus | 89 | −26.0 | 2.47 | <0.001 |
| Ruminococcus torques | 78 | 2.96 | 0.62 | <0.001 |
| Unspecified Lachnospiraceae | 67 | −2.12 | 0.76 | 0.047 |
| Coprococcus | 53 | −2.22 | 0.49 | <0.001 |
| Bacteroides | 50 | 3.01 | 0.97 | 0.018 |
| Unspecified Clostridiales | 46 | −10.8 | 2.44 | <0.001 |
| Unspecified Christensenellaceae | 43 | −5.96 | 2.01 | 0.043 |
| Bacteroides eggerthii | 27 | 4.49 | 0.89 | <0.001 |
| Ruminococcus | 20 | −24.2 | 2.94 | <0.001 |
| Bacteroides plebeius | 14 | −9.59 | 2.94 | 0.013 |
| Eubacterium biforme | 11 | −24.1 | 2.94 | <0.001 |
| Parabacteroides | 10 | 2.55 | 0.94 | 0.047 |
| Coprococcus eutactus | 6.6 | −5.87 | 1.81 | 0.013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoll, M.L.; DeQuattro, K.; Li, Z.; Sawhney, H.; Weiss, P.F.; Nigrovic, P.A.; Wright, T.B.; Schikler, K.; Edelheit, B.; Morrow, C.D.; et al. Impact of HLA-B27 and Disease Status on the Gut Microbiome of the Offspring of Ankylosing Spondylitis Patients. Children 2022, 9, 569. https://0-doi-org.brum.beds.ac.uk/10.3390/children9040569
Stoll ML, DeQuattro K, Li Z, Sawhney H, Weiss PF, Nigrovic PA, Wright TB, Schikler K, Edelheit B, Morrow CD, et al. Impact of HLA-B27 and Disease Status on the Gut Microbiome of the Offspring of Ankylosing Spondylitis Patients. Children. 2022; 9(4):569. https://0-doi-org.brum.beds.ac.uk/10.3390/children9040569
Chicago/Turabian StyleStoll, Matthew L., Kimberly DeQuattro, Zhixiu Li, Henna Sawhney, Pamela F. Weiss, Peter A. Nigrovic, Tracey B. Wright, Kenneth Schikler, Barbara Edelheit, Casey D. Morrow, and et al. 2022. "Impact of HLA-B27 and Disease Status on the Gut Microbiome of the Offspring of Ankylosing Spondylitis Patients" Children 9, no. 4: 569. https://0-doi-org.brum.beds.ac.uk/10.3390/children9040569