An Overview of Exosomes in Cancer Therapy: A Small Solution to a Big Problem



1
Department of Pharmaceutical Technology, Faculty of Pharmacy, University of Coimbra, Azinhaga de Santa Comba, Pólo III—Pólo das Ciências da Saúde, 3000-548 Coimbra, Portugal
2
Group of Pharmaceutical Technology, REQUIMTE/LAQV, Faculty of Pharmacy, University of Coimbra, Azinhaga de Santa Comba, Pólo III—Pólo das Ciências da Saúde, 3000-548 Coimbra, Portugal
*
Author to whom correspondence should be addressed.
Processes 2020, 8(12), 1561; https://0-doi-org.brum.beds.ac.uk/10.3390/pr8121561
Submission received: 23 October 2020
/
Revised: 23 November 2020
/
Accepted: 25 November 2020
/
Published: 27 November 2020
(This article belongs to the Special Issue Development and Characterization of Nanocarriers for Drugs)
Abstract
:Exosomes are defined as a type of extracellular vesicle released when multivesicular bodies of the endocytic pathway fuse with the plasma membrane. They are characterized by their role in extracellular communication, partly due to their composition, and present the ability to recognize and interact with cells from the immune system, enabling an immune response. Their targeting capability and nanosized dimensions make them great candidates for cancer therapy. As chemotherapy is associated with cytotoxicity and multiple drug resistance, the use of exosomes targeting capabilities, able to deliver anticancer drugs specifically to cancer cells, is a great approach to overcome these disadvantages. The objective is to assess treatment efficiency in reducing tumor cells, as well as overall safety and response by cancer carriers. So far, results show exosomes as a promising therapeutic strategy in the fight against cancer. This review summarizes the characteristics and composition of exosomes, as well as explaining in detail the involved parties in the origin of exosomes. Furthermore, some considerations about exosome application in immunotherapy are addressed. The main isolation and loading methods are described to give an insight into how exosomes can be obtained and manipulated. Finally, some therapeutic applications of exosomes in cancer therapy are described.
1. Introduction
Cancer remains one of the leading causes of death worldwide, despite significant therapeutic advancements and improved detection methods. The main characteristic of this disease is the uncontrolled cell proliferation. There are several types of cancer, related to different types of cells, with different behaviors and responses to treatment, and various contributing factors, such as diet, lifestyle, inherited genes, infectious microorganisms, as well as exposure to radiation and carcinogenic substances [1,2]. Usual cancer treatments involve chemotherapy, radiation therapy, and/or surgery. Chemotherapy is considered the most effective therapy. However, treatment fails in several situations where cancer cells show resistance to chemotherapeutic drugs. In fact, two of the biggest threats to chemotherapy are related with multiple drug resistance and with treatment toxicity [3]. In this regard, therapies based in nanoparticles (NPs) are being investigated, focused on efficient drug delivery methods, as a way to reduce adverse effects and improve the chances of a successful treatment [4].
In 1983, while observing the maturation of reticulocytes into erythrocytes, Johnstone et al. reported that fusion of multivesicular bodies (MVBs) with the plasma membrane (PM) led to the release of nanosized vesicles. Later, in 1987, these vesicles were named exosomes [5,6]. At first, it was believed that exosomes acted as cellular disposal systems, helping with the removal of unnecessary proteins [7]. However, nowadays it is known that exosomes play an important role in extracellular communication, capable of carrying proteins, nucleic acids, and lipids [8]. Taking advantage of their innate characteristics and functions, exosomes are being investigated as potential therapeutic agents. As discussed further on, exosomes are capable of triggering an immune response, prompting the development of vaccines in immunotherapy. In addition to the capacity for targeting and recognizing proteins on a cell membrane, they can also act as nanocarriers of drugs and nucleic acids for several pathologies, namely cancer.
This review aims to describe the composition of exosomes, as well as give insight into the underlying mechanism responsible for the origin of these vesicles. What could be simply described as a form of exocytosis, is in fact a much deeper process. With the help of several proteins working together as one complex, there are several steps involved in the development of exosomes. Although there is still much to learn about this mechanism, there is already proof of other pathways that lead to the biogenesis of exosomes [9,10,11].
Nowadays, there are many techniques used to obtain and purify exosomes. Some techniques are more popular than others due to their accessibility and simpler methodology (for example, centrifugation-based methods), while others, which present higher yields, are associated with elevated costs. Depending on the goal of each study, one technique may be preferable to another. For these reasons, several techniques were described, each with their own advantages and disadvantages.
Due to the capability of exosomes to carry molecules, several advances have been made in order to load exosomes with a desired drug or nucleic acid. It was, therefore, relevant to summarize some of the most commonly performed loading strategies. The different techniques were divided in pre- and post-loading methods, varying on whether the exosome loading occurs in a direct or indirect manner. Like isolation techniques, loading techniques also have advantages and disadvantages. These are mostly related to the drug loading capacity of the method, and to the stress induced on the vesicles.
Knowledge of these techniques provides some background into the posteriorly referred clinical applications. Several in vivo experiments were performed, using different isolation and loading methodologies. Results of different investigations will be discussed, evaluating the efficacy of exosome-based treatments using paclitaxel (PTX), small interfering RNA (siRNA), and doxorubicin (DOX) [12,13,14].
2. Exosomes
2.1. Structure and Functions
Cells from the most diverse organisms are known to release extracellular vesicles, which can be found in several biological fluids like blood, saliva, urine, lymph, and breast milk [15,16]. One of these vesicles is the exosome, a particle of endocytic origin with a size ranging from 30–100 nm [15,17,18]. The morphology of these particles has been described as being “saucer-like” [18] or “cup-shaped” [19], resembling a flattened sphere, depending on the cell of origin [18,19,20]. Exosomes are lined with a lipid bilayer that encloses cytosol from the secreting cells, and are composed by several lipids, proteins, and nucleic acids. Similar to their morphology, the composition of exosomes is also influenced by the parent cell that originated them [15,17,20].
Over the years, the protein composition of exosomes has been the subject of several studies. According to ExoCarta, an online database regarding exosomes, 9769 proteins have been identified in these systems [21], such as adhesion molecules, major histocompatibility complex (MHC) class I and II proteins, cytosolic chaperone proteins (for example, heat shock proteins (Hsp)) and metabolic enzymes [22,23]. One of the most commonly found protein family in exosomes are the tetraspanins (namely CD9, CD63, CD81, and CD82) [18,24]. The tetraspanins are a superfamily of proteins with four transmembrane domains and two extracellular segments [25,26]. These compartments are responsible for several processes that result in membrane fusion, organization of large molecular complexes and even protein trafficking and signaling [18,24]. Since the membrane of exosomes is enriched with tetraspanins, they are considered excellent biomarkers [24].
Another set of proteins present in the composition of exosomes are the Hsp. These systems act as molecular chaperones in response to stress. By helping to maintain an appropriate folding of proteins, Hsp prevent the formation of protein aggregates. These misfolded proteins could lead to the most diverse pathologies, such as neurodegenerative disorders like Parkinson or Alzheimer’s disease [27]. The correct manipulation of their biological activity could be the answer to develop new therapies for the mentioned diseases or for several types of cancer [28]. The expression of Hsp can be constitutive or induced due to stressful events (like exposing a cell to higher temperatures) [29]. In an attempt to induce the production of Hsp to be sorted into B-cell exosomes, Clayton et al. exposed the cell line to high temperatures (42 °C over 3 h). In this study, it was concluded that there was an increase both in B-cell exosomes secretion, as well as in the amount of produced Hsp. Further analysis showed that the Hsp are encapsulated in the lumen of the exosomes. This renders them unavailable to interact with the Hsp receptors present at the surface of target cells [27], suggesting that other mechanisms could be involved in order to allow the exosomal Hsp to interact with biological targets.
Exosomes from antigen-presenting cells (APC) carry MHC class II molecules [18], which could enable them to play a role in antigen presentation. Raposo et al. documented that peptide-MHC class II complexes present at the surface of exosomes were able to stimulate T cells [30]. It has also been shown that dendritic cells (DC) produce exosomes with MHC class I molecules and CD86, allowing them the potential to induce a response from CD8 + T-Cells [18,31].
Regarding their lipid composition, exosomes have resemblances with lipid rafts [15,32]—microdomains expressed at the PM with elevated content of sphingolipids, cholesterol, and phospholipids [33]. According to studies, in comparison with their parent cells, exosomes are usually enriched 2–3 times more in sphingomyelin, cholesterol, phosphatidylserine and glycosphingolipids, with values varying with each type of originating cell [32,34]. Besides their structural function in the PM and role in the formation of exosomes, lipids could also have an influence on the function of these particles in the body, making them important targets for future studies, as to better understand how exosomes work and how to easily manipulate them as therapeutic agents [32,34].
Besides their protein and lipid composition, exosomes also act as carriers of nucleic acids, namely messenger RNA (mRNA) and microRNA (miRNA) [35]. miRNAs are small, non-coding RNA molecules, that can regulate the expression of genes and complementary mRNAs, playing a role in cellular development, proliferation, and apoptosis [35,36,37]. The transfer of nucleic acids by exosomes from one cell to another can influence cells on an epigenetic level and lead to an exchange of features between cells, since receiving cells are getting RNA of proteins that wouldn’t be normally expressed in them [35,37]. Like other cell types, cancer cells can also release exosomes, and with them, their own nucleic acids. Melo et al., studied the effects of miRNAs transported by breast cancer cells-derived exosomes. In this study, it was suggested that these molecules have the capacity to induce the formation of tumours on cells that would otherwise be considered healthy [38]. As alarming as it may be, these conclusions shed a light on cancer diagnostic and treatment methods, with the possibility of using miRNAs transported by exosomes as biomarkers in melanoma, breast cancer or lung cancer [17,37]. Such mechanisms could be adapted to our advantage, using exosomes as vectors in genetic therapy, and loading specific sets of nucleic acids to use them as carriers between cells [39].
Due to their composition (see Figure 1), exosomes act as nanocarriers in the transfer of macromolecules around the organism. They play a fundamental role as mediators in cell to cell communication, whether cells are close to or far from each other [10,35]. These nanosystems are capable of eliciting biological responses like expression/suppression of proteins, induce immune responses, or modulate cancer progression [24,40].
2.2. Biogenesis
As previously mentioned, exosomes are considered extracellular vesicles with an endocytic origin [15]. When a cell membrane site is invaginated through the ubiquitination of surface receptors, it leads to the formation of early endosomes [16]. As they mature, there is a gradual increase in size and change in their content, mostly due to the accumulation of intraluminal vesicles (ILV). They eventually become late endosomes, also referred to as MVBs [17,41]. After their maturation, MVBs usually fuse with lysosomes, and their contents suffer lysosomal degradation. However, by a process that still is not quite understood, the membrane of MVBs can fuse with the PM of the cell, leading to the release of the accumulated ILVs, which are now called exosomes (see Figure 2) [17,24,41,42].
One of the most studied mechanisms for the formation and cargo sorting of ILVs into the MVBs involves the endosomal sorting complex required for transport (ESCRT) [10,17]. The ESCRT family is composed by almost twenty proteins, distributed by four multiprotein complexes (ESCRT-0, ESCRT-I, ESCRT-II and ESCRT-III), along with accessory protein Vps4. All of them are involved in the biogenesis of exosomes, transporting ubiquitinated proteins on the membrane of endosomes in the form of a cluster, so that they can later be included in a newly formed vesicle, inside the endosome [24]. The discovery of the ESCRT machinery began after the identification of several genes in Saccharomyces cerevisiae yeast, classified as “Vps” genes, whose proteins had the capacity to form complexes and mediate protein trafficking through endosomes [44]. Since then, the human analogues of these proteins have been discovered and categorized according to the ESCRT complex (see Table 1).
Sites of the cell membrane start forming early endosomes when proteins at their surface are ubiquitinated. Due to the action of Vps34, or class III Phosphatidylinositol-3-Kinase (PI3K), the lipid phosphatidylinositol (PI) present at the surface of the cell and the endosome is phosphorylated into phosphatidylinositol-3-phosphate (PI(3)P) [45]. The presence of PI(3)P acts as a recognition signal with specificity for the Hrs protein, recruiting the ESCRT-0 complex which binds to the ubiquitinated proteins at the surface of the endosome, initiating cargo sorting into the MVBs [41,44]. After binding with proteins, the ESCRT-0 begins to cluster and sequester them and, due to the Hrs subunit, binds itself with a Tsg101 unit from ESCRT-I, which proceeds to recruit the ESCRT-II complex via Vps28/Vps36 binding [44]. ESCRT-I and ESCRT-II work together to create buds and stabilize vesicle necks, allowing for ESCRT-III to promote the budding process and cleaving inward budding vesicles. These vesicles are released into the lumen of the endosome, originating ILVs [4,46]. Unlike the other three complexes, ESCRT-III exists in the cytosol as inactive monomers instead of an active complex, and depends on ESCRT-II to be activated (in yeasts, ESCRT-II protein Vps25 binds to Vps20, and afterwards Snf7, Vps24 and Vps2 bind to each other, sequentially) [42,45]. The exact cleaving mechanism is not fully understood, however it is known that Snf7/CHMP4 is responsible for recruiting the degradation of alpha-4 (Doa4) enzyme. This enzyme is responsible for the removal of ubiquitin from the cargo of MVBs, allowing them to be sorted and incorporated in vesicles [47,48]. Before the process is complete, the ESCRT-III complex is disassembled and recycled back to the cytosol after binding with the Vps4 ATPase enzyme, which requires adenosine triphosphate (ATP) (see Figure 3) [17,42,45].
Even though the ESCRT pathway is considered a key mechanism in the biogenesis of MVBs, there is evidence of ESCRT-independent mechanisms. Stuffers et al. depleted cells of Hrs, Tsg101, Vps22 and Vps24, subunits of each ESCRT complex, in order to evaluate the production of MVBs and the secretion of exosomes in the absence of ESCRT proteins. After transfecting siRNA against the mentioned proteins using Hep-2 and HeLa cells, the integrity of the ESCRT complex was lost, yet MBVs were still being formed, suggesting the existence of mechanisms independent of ESCRT [11].
One of the supposed mechanisms involved in MVBs formation revolves around tetraspanins, namely, CD63 [25], which, unlike other proteins of the tetraspanins family, is abundantly found intracellularly in ILVs of late endosomes [49]. The melanosomal protein Pmel17 (essential in the maturation of melanosomes) is transported into ILVs without the need for ubiquitination and, therefore, does not need to be sorted by the ESCRT complex [50]. After depleting CD63 in MNT-1 cells with CD63-specific siRNA, there was a reduction of ILVs on endosomes, with consequent impairment of melanosome maturation [51], suggesting an important role for CD63 in ESCRT-independent mechanisms.
Besides proteins, lipids also seem to play a role when it comes to generate ILVs without the ESCRT machinery (see Figure 4). Ceramide has been the subject of several studies, as to better understand how it operates. Studying the endosomal trafficking of the proteolipid protein (PLP), it was observed that the ESCRT complex was not involved in PLP sorting after using siRNA to knock down Hrs, Tsg101 or Vps4 coding genes. In the absence of ESCRT, ILVs containing PLP were still being formed, with a high concentration of sphingolipids in the vesicles [52]. Sphingomyelinases remove the phosphocholine group of sphingomyelin leading to the formation of ceramide. In order to analyse the role of ceramide in PLP vesicle sorting and exosome biogenesis, Oli-neu cells (mouse oligodendroglial cells) were treated with sphingomyelinase inhibitor GW4869. Other inhibitors, spiroepoxide and glutathione, were also used. The same effect could be observed in the three tests, with a marked decrease of exosome secretion from the cells, concluding that ceramide is also involved in ESCRT-independent ILV formation [52]. A similar study was performed in human embryonic kidney (HEK) 293 cells to prove that miRNA secretion depends on ceramide. First, RNases were added to the medium in order to assess if extracellular miRNA is contained in exosomes. After the cells were incubated with the RNases, miRNA could still be detected. Exosomes play an important role in protecting carried molecules from external threats. Afterwards, two tests were performed, one with GW4869 and another with siRNA to knock down the expression of sphingomyelinase. It was concluded that depletion of ceramide lead to a decrease of both exosomes and miRNA in the extracellular medium. Although the exact mechanism of how miRNAs are sorted into exosomes is still unknown, an ESCRT-independent mechanism involving ceramide could be responsible for this phenomenon [9].
Several studies are trying to discover more about how MVBs fuse with the PM and release exosomes to the extracellular microenvironment. Two groups of proteins seem to be a part of this mechanism, small GTPases of the RAB family and sets of soluble N-ethylmaleimide-sensitive fusion attachment protein receptors (SNAREs) [15]. The use of small-hairpin RNA (shRNA) to knock down Rab proteins (Rab2b, Rab9a, Rab5a, Rab27a, and Rab27b) led to an inhibition of exosome secretion in HeLa cells [53]. A focused analysis on proteins Rab27a and Rab27b led to the discovery that these two proteins are involved in MVBs distribution to the cell periphery, after using total internal reflection fluorescence microscopy to observe MVBs. The silencing of Rab27 proteins caused a reduction in the fusion events between MVBs and the PM, but also led to an increase in MVB size, suggesting that, in the absence of these proteins, MVBs could fuse with each other or form complexes with other vesicles, preventing them from docking with the PM [53].
Over the years, several studies have proven the involvement of SNAREs in vesicle fusion with the PM, for example, in lysosomal exocytosis [54]. However, the SNARE complexes that intervene in that process could be different from the SNAREs involved in fusion of the MVB with the PM [31]. Fader et al. observed that the vesicle-associated membrane protein 7 (VAMP7), a member of the SNARE family, was necessary for the fusion of MVBs with the PM, leading to exosome secretion. Inhibition of these protein in K562 cells (myeloid leukaemia cell line) led to a decrease in exosome secretion [55]. In another study, Proux-Gillardeaux et al. concluded that the inhibition of VAMP7 on Madin-Darby canine kidney cells impaired lysosome secretion but had no influence on exosome release [56]. Although contradictory, these results do not discard the role of SNAREs in the biogenesis of exosomes, but instead indicate that different cell types could require other proteins from the SNARE family. Further studies should be conducted to better understand how these molecules work [8,15].
2.3. Exosome-Like Endogenous Nanosystems
Prolonged cancer therapies cause systemic toxicity and healthy cell damage, limiting the efficacy of cytostatic drugs [57]. New drug delivery systems based on NPs could be a way to reduce side effects and improve the treatment of patients. These nanosystems can target tumours while protecting the drug from degradation and enhancing endocytosis and drug uptake in the cells [57,58]. As a result of the flawed angiogenesis and poor lymphatic drainage characteristic of tumour cells, nanocarriers accumulate around the blood vessels, by a phenomenon termed the enhanced permeability and retention effect [58]. Also, some nanosystems (such as exosomes) can actively target cells by recognition of surface proteins or other biomolecules. Unfortunately, synthetic nanosystems have the disadvantage of being recognized and eliminated by the reticuloendothelial system [59]. On the other hand, NPs like exosomes have an endogenous nature, making them the perfect candidates for drug delivery given their high stability, low immunogenicity, and biocompatible properties [14].
Exosomes can be administered through several routes. The most frequent one is intravenously, followed by intranasal administration, with reports of oral and subcutaneous administration of extracellular vesicles [60,61,62]. Owing to their composition, the administration of exosomes derived from cells similar to the targeted tissues leads to a higher targeting specificity [62]. The composition of exosomes gives them an innate ability to target other cells, and their structure allows the encapsulation of molecules that can be delivered to such cells. However, cells release exosomes in relatively low amounts, and current purification methods can be inefficient, leading to low yields [57]. This issue has led to the development of several studies with the objective of directing exosomal targeting and to formulate nanocarriers with the same characteristics of exosomes, enhancing their therapeutic effect.
As previously referred, exosomes can target cells based on their molecular composition. Such targeting ligands can, however, be artificially modified [63]. One method commonly used to engineer desired targeting ligands on exosomes revolves around plasmid transfection. Alvarez-Erviti et al. transfected plasmids encoding Lamp2b constructs into DCs. One of these constructs was comprised of the rabies viral glycoprotein (RVG) peptide, that binds to acetylcholine receptors. After exosome purification, quantitative PCR assay confirmed the expression of RVG-Lamp2b complexes on the surface of the dendritic cell-derived exosomes. Using fluorescence microscopy, it could be observed that the exosomes targeted neuronal cells (microglia, oligodendrocytes, and neurons). The method used for exosomal targeting was, therefore, successful [64].
Since the amount of exosomes released by cells is low, Jang et al. studied a technique to obtain high amounts of exosome-like nanovesicles. Monocytes and macrophages were loaded with DOX, a chemotherapeutic drug. In an extrusion process, the cells were passed through filters with different pore sizes (from 10µm to 1µm). The obtained products were nanovesicles with the same protein composition of the PM of the cells they derived, similar to the composition of exosomes released by the same cells. The protein content (analysed by western blotting) of exosomes and the obtained nanovesicles revealed the presence of CD63, Tsg101, Moesin and Beta-actin in both particles (see Figure 5) [57].
In the same study, mice were transplanted with CT26 cells (a colorectal carcinoma cell line that causes tumours when introduced in mice). After tumour growth was observed, exosome-like nanovesicles loaded with DOX were administered. Tumour reduction could be observed and there was no reported decrease in body weight and white cells count. In addition to these promising results in terms of therapeutic efficacy, the yield of the exosome-like particles was about 100-fold higher than the quantity of naturally released exosomes [57]. Hence, the method used to obtain exosome mimics is a great alternative to study the effects of these particles as drug delivery systems.
2.4. Immunotherapeutic Potential of Exosomes
As approached before, due to their composition, exosomes could elicit some sort of immune response. Tumour-derived exosomes (TEXs) carry a various array of proteins—such as MCH class I and II, CD9, CD63—including tumour specific antigens. Due to their composition, TEXs can initiate a tumour-targeted immune response. DCs recognize and process antigens from TEXs, presenting them to helper T lymphocytes, causing their activation. Furthermore, TEXs can stimulate natural killer (NK) cells. Incubation of Hsp70-expressing TEXs and CD94+ NK cells stimulates the activation of NKs and release of granzyme B [65].
In contrast, TEXs can also have an immunosuppressive effect. For example, the presence of TGF-β1 and galectin-1 produce a suppressive effect in CD4+ and CD8+ T lymphocytes [65]. In addition, TEXs are known to be involved in tumour progression, aiding the process of angiogenesis and metastasis, and inhibiting cancer cells apoptosis [66]. One of the challenges against cancer therapies is the presence of PD-L1 on the surface of tumour cells. When PD-L1 binds to its receptor on T cells, it leads to the suppression of T lymphocyte activity. TEXs have been found to express PD-L1, which can be used as a biomarker. High levels of TEXs containing PD-L1 generally indicate a negative prognosis in patients treated with PD-L1 inhibitors [67].
Since its first application, chimeric antigen receptor T (CAR-T) cell immunotherapy became a promising oncologic therapeutic. Using a viral vector, T cells can be recombined with a specific antigen receptor. Through this method, T lymphocytes are targeted to a certain tumour-associated antigen with a higher precision, leading to a more effective cytotoxic response [68]. Unfortunately, there are still many limitations associated with this method. Since CAR-T cells are active, they can expand uncontrollably. In addition, ten days after CAR-T cell infusion, two thirds of the patients usually experience an adverse effect called cytokine release syndrome (CRS). CRS is caused by an uncontrollable release of cytokines by the modified T cells [68]. A way to surpass this effect would be with CAR-T cell derived exosomes. These exosomes, which are released by CAR-T cells, retain the therapeutic capacity of their parent cells. Administrating CAR-T cell derived exosomes would enable control of the in vivo expansion that CAR-T cells go through. In this sense, CRS could be avoided [69].
So far, CAR-T cell therapy has only proved itself efficient in haematological conditions (such as lymphomas). Owing to their nanometric size, CAR-T cell derived exosomes can easily cross the blood-brain barrier or tumour cell membranes. This particularity could suggest the application of CAR-T therapeutic towards other pathologies [68,69].
3. Isolation Techniques
3.1. Traditional Methods
Due to their composition and functions, exosomes have gained attention as potential tools in the diagnosis and treatment of oncologic diseases. Although there is no standardization when it comes to exosome isolation, several methods have been developed to facilitate their extraction [70,71].
The following topics give insight into some techniques applied for exosomal extraction, as well as the advantages and disadvantages behind each method. In order to obtain exosomes of higher purity, some of these techniques are performed sequentially [72]. To confirm the obtained particles after each procedure, performing a Western blot and/or a flow cytometry analysis is common practice [73,74].
3.1.1. Differential Centrifugation/Ultracentrifugation
In centrifugation, particles in a sample are subjected to a centrifugal force. This causes sequential sedimentation of the particles according to their size and density, with heavier/bigger particles depositing first [71,73]. In differential centrifugation, three successive centrifugations are performed with increasing centrifugal force and duration, aiming to remove cells, cellular debris and macromolecules from the rest of the sample [71]. The three centrifugal forces and durations are 300× g for 10 min, 2000× g for 10 min and 10,000× g for 30 min, respectively [74]. Between each sequence, the supernatant is aspired [73]. After the differential centrifugation is performed, an ultracentrifugation step is followed, applying forces of 100,000× g for 70 min, causing the exosomes to form a pellet. This step can be repeated by carefully removing the supernatant and re-suspending the exosome pellet in phosphate buffered saline (PBS) and subjecting it to another centrifugation [71,73,74].
The method of differential centrifugation/ultracentrifugation is the most widely used when it comes to exosome isolation [71]. It is considered as being easy to perform and not requiring much technical expertise nor sample pre-treatment. In terms of disadvantages, the extreme forces applied in ultracentrifugation could damage the exosomes, making them unviable for further testing. Some nanovesicles tend to be trapped by bigger particles during centrifugation and lost during supernatant aspiration, resulting in low yields [70].
3.1.2. Density Gradient Centrifugation
A variation of ultracentrifugation is the method of density gradient centrifugation. In this technique, the obtained supernatant from a differential centrifugation is placed in a density gradient medium [73]. A sucrose gradient medium is commonly used, being built into an ultracentrifuge tube. Instead of sucrose, an iodixanol gradient can also be used, with reported improvements when it comes to the separation of nanovesicles from viral particles [75]. The density of the medium increases linearly from top to bottom, and after applying a centrifugal force, the particles in the sample separate themselves based on their densities [73]. The sample is placed on the top of the density gradient and ultra-centrifuged at 100,000–200,000× g for long periods of time, ranging from one to five hours, with some studies even going as far as 16 h [72,74]. As mentioned, the particles in the sample sediment along the medium until they reach a density equal to their own, the isopycnic position [73]. For exosomes, that density is between 1.10 g/mL to 1.19 g/mL [71,74]. The particles of interest can then be extracted by simply collecting the fractions of the suspension.
Unlike the traditional method for ultracentrifugation, this approach prevents exosomes mixing with residual proteins or previously separated particles. Having higher separation efficiency, this technique allows one to obtain exosomes with a higher purity [71,73]. Regardless of the efficiency of the method, it is more complex in terms of technique, involving the preparation of the density gradient medium, with more costs associated. Additionally, the process itself requires more time to be performed than traditional ultracentrifugation [71].
3.1.3. Immunoaffinity Isolation Techniques
The surface of exosomes is covered by proteins and other macromolecules. These molecules can be targeted by corresponding ligands, similar to antigen-antibody interactions [73]. A method for isolation of exosomes has emerged based on the immunoaffinity between their proteins. Immunoaffinity isolation of exosomes uses magnetic beads coated with monoclonal antibodies. These antibodies specifically target proteins present at the membrane of exosomes, such as CD9, CD63 and CD81 [71,76]. The magnetic beads are added to a sample and attach themselves to the exosomes. After a magnetic force is applied, the magnetic beads are retained and the rest of the sample is discarded, while the exosomes attached to the beads remain [71]. This method allows for the targeting of exosomes derived from specific cells with unique markers. Mathivanan et al. used this technique to isolate exosomes derived from LIM1215 cells, a colorectal carcinoma cell line. In order to capture the exosomes, the group used beads coated with A33 antibodies. A33 is a protein commonly expressed in colon epithelial cells. It was safe to assume that exosomes released from LIM1215 cells would express the A33 protein and be targeted by the A33-antibody coated beads [77].
After binding to the exosomes, removing the magnetic beads is a difficult task, which limits the use of the exosomes for further studies [71]. In addition, the reagents necessary for this technique are costly, impairing accessibility to the method. In order to avoid interference, cells and cell debris must be absent from the sample. This suggests the need to perform other isolation techniques first [73,77].
3.1.4. Size Exclusion Chromatography (SEC)
The basis of SEC is to sort particles in a sample according to their size. The particles move across a column that contains the stationary phase, a porous gel [74]. Based on their sizes, the particles will move across the column at different rates. Since larger particles cannot penetrate through the pores of the stationary phase, these elute first by the mobile phase [73]. Smaller particles are retained, meaning that they are eluted at a slower rate. Afterwards, the eluted fraction containing exosomes is collected. Prior to SEC, it is common to perform a low-speed centrifugation to remove larger components from the sample, like cells, cell debris and macromolecules. The sample is also filtered in order to concentrate extracellular vesicles [74].
3.1.5. Hydrophilic Polymer Precipitation
Polyethylene glycol (PEG) is a polymer commonly used to precipitate macromolecules, proteins, nucleic acids, viruses, and other particles [78]. Due to their hydrophilic nature, PEGs complex with water molecules, forcing less soluble components to precipitate [73]. The common procedure starts by incubating the sample with a precipitating solution at 4 °C, followed by low-speed centrifugation (1500× g). The pelleted exosomes can then be re-suspended in PBS [78].
This technique is simple, fast, easy to execute, and does not deform the exosomes. Several precipitating kits specialized for exosome precipitation are commercially available, but are associated with high costs [71]. Cells and cell debris need to be removed from the sample to avoid interference. Like the exosomes, proteins, nucleic acids, and other particles present in the sample can precipitate after incubation with PEG [78]. Therefore, performing additional techniques might be necessary in order to minimize the presence of impurities [73,78].
3.2. Novel Methods
Although widely used, traditional methods still present various limitations: long running times, high costs, and damage to the integrity of the nanovesicles. In recent years, new techniques have been developed to isolate and identify exosomes, attempting to surpass some disadvantages of traditional methods [71].
3.2.1. Stirred Ultrafiltration
The principle of stirred ultrafiltration is similar to traditional membrane filtration. Particles in a sample are separated depending on their size. Based on the pore size of the ultrafiltration membrane, exosomes and small molecules pass through the filter along with the solvent [71]. The sample is subjected to constant stirring to prevent the membrane from becoming clogged by bigger particles that were retained. It also requires an external source of pressure, usually nitrogen, to push the sample through the filtration membrane. Afterwards, the membrane is usually rinsed with PBS [70].
This method allows the purification of large volumes of sample and to obtain large amounts of exosomes in a short time span. In comparison to the forces applied in ultracentrifugation, there is a lower risk of damaging the integrity of exosomes when using this technique [71]. However, the membrane only discriminates particles based on their size. Small molecules and other particles can pass through the filter, diminishing the purity of the isolate [73].
3.2.2. Nanoplasmon-Enchanced Scattering (nPES)
The purpose of nPES is not directed towards exosome isolation in the same way as the previously mentioned methods. Instead, the main goal of this technique is to detect and quantify exosomes present in a sample in a rapid, sensitive, and specific manner [71]. The methodology resembles an enzyme-linked immunosorbent assay (ELISA). A sensor chip is coated with antibodies against typical exosomal markers (for example, CD81) [79]. When a sample is added to a well, exosomes expressing the marker are retained and accumulated in the wells. Afterwards, gold NPs, also coated with exosomal markers (antibodies for CD9 or CD63), are added to the medium. According to their size and shape, the gold NPs scatter light at different wavelengths. The sensor chip is then placed under a dark field microscope to analyse the light scattering [79]. Based on the scatter area, the number of exosomes can be quantified [71].
Liang et al. observed that exosomes secreted by pancreatic cancer cells express the ephrin type-A receptor 2 (EphA2). In this study, they used antibodies for EphA2 and used nPES to identify exosomes bearing this protein. This method successfully detected EphA2-expressing exosomes. In addition, it was also possible to quantify exosomes of patients with pancreatic cancer and to compare results with post-treatment samples. Besides diagnosis, the nPES technique could give insight into the success rate of oncologic treatments by monitoring exosomes expressing oncologic markers [79].
4. Exosome Loading Strategies
Some exosomes naturally function as cargo deliverers from one cell to another. Taking advantage of this ability, these particles can be artificially loaded with drugs and nucleic acids. The several existing techniques can be divided in pre-loading and post-loading methods. In pre-loading methods, the exosomes are indirectly loaded with the drug, unlike post-loading methods.
4.1. Pre-Loading Methods
Incubation with Donor Cells
4.2. Post-Loading Methods
4.2.1. Incubation with Exosomes
Extracted and purified exosomes are incubated with a drug, similarly to the previous method. Due to the concentration gradient, the drug molecules diffuse into the exosomes [63].
Both incubation techniques, considered passive methods for drug loading into nanosystems, are simple to execute. However, due to the hydrophobic interactions between certain drugs and the lipid bilayer of the nanovesicles, the loading efficiency is relatively low [63].
4.2.2. Electroporation
In electroporation, exosomes are placed in a conductive solution and subjected to an electrical field. The applied current causes the phospholipid bilayer of the exosomes to rearrange, forming small pores in the membrane [63]. The presence of these pores leads to the diffusion of drugs and/or nucleotides into the interior of the exosomes. After the loading process is completed, the integrity of the PM is restored [80].
Electroporation is the selected method to encapsulate nucleotides. siRNA possesses a negative charge and cannot diffuse through the hydrophilic shell of the PM [81]. Nevertheless, if an optimized buffer is not used, electroporation could cause RNA aggregation and disrupt the stability of the exosomes, reducing the loading efficiency [80]. Johnsen et al. suggested the use of a trehalose containing buffer to maintain the integrity of the exosomes [82].
4.2.3. Sonication
This technique uses ultrasonic frequencies to agitate particles in a suspension. The exosomes are mixed with the drug and a probe sonicator induces deformation of the PM of the nanovesicles [80]. While the membrane is deformed, drugs diffuse inside the exosomes. The integrity of the membrane is restored about an hour after the sonication process, as previously reported [12].
Kim et al. loaded exosomes with PTX using different methods: incubation with free exosomes, electroporation, and sonication. The obtained exosome-PTX nanosystems were purified using SEC and analysed by high performance liquid chromatography (HPLC) to evaluate the loading efficiency of each method. The study concluded that sonication led to higher amounts of PTX loading into the exosomes, followed by electroporation and drug incubation with free exosomes [12].
4.2.4. Freeze/Thaw Cycles
In this method, exosomes are incubated with the drug. Afterwards, these are rapidly frozen at −80 °C and thawed at room temperature [80]. In each cycle, the PM is disrupted, allowing drug molecules to diffuse inside the exosomes. The cycles are repeated several times to achieve equal drug concentrations inside and outside the nanovesicles [83].
In terms of loading capacity, this method is lower than other techniques. such as electroporation or sonication [63].
4.2.5. Saponin Assisted Incubation
In this methodology, exosomes are incubated with the drug and saponins. Saponins are surfactant molecules that form complexes with cholesterol, present in the PM of exosomes, generating pores [63]. This increase in permeability allows for the easier loading of hydrophilic molecules when compared with a simple incubation [84].
The exact mechanism of how saponins interact with cholesterol is not yet understood. However, it is known that they interact in the same way as with cholesterol on the PM of red blood cells. This fact could be quite harmful when used in in vivo studies, due to the haemolytic activity of saponins. To perform this technique, the saponin concentrations should be minimal, and the exosomes must be thoroughly washed after being incubated with saponins [85].
5. Exosomes as Nanosystems of Nucleic Acids and Drugs
5.1. Paclitaxel
PTX is one of the most used anticancer drugs. It is naturally found in the bark of Taxus brevifolia, although new extraction methods have been developed to obtain this drug [86]. PTX promotes the assembly of tubulin into microtubules, stabilizing the latter and preventing their dissociation. Since the microtubules do not dissociate, cell cycle progression is blocked, and the growth of cancerous cells is hampered (see Figure 6) [87].
PTX has been used in the treatment of several types of cancer, namely colorectal, ovarian, breast or lung cancer [86,87]. The efficiency of PTX and other chemotherapeutic drugs is limited by the emergence of multiple drug resistance (MDR) [89]. The overexpression of ATP-binding cassette (ABC) transporters, specifically the drug efflux P-glycoprotein (Pgp) transporter, is one of the mechanisms mediating MDR in cancer cells [12]. To overcome MDR, Kim et al. evaluated oncological treatment efficiency of PTX-loaded exosomes.
The exosomes used in this study were extracted from a murine macrophage cell line, using a precipitating polymer (ExoQuick-TC™ kit). As previously mentioned, three loading methods were tested in order to evaluate the loading capacity of each technique. The exosomes loaded with PTX were purified using SEC and analysed by HPLC. The exosome-PTX complex obtained by sonication was used for further studies since it showed better values of loaded PTX [12].
To evaluate the antineoplastic effect of exosome-PTX complex, a Lewis Lung Carcinoma mouse model was used. Using a lentiviral vector, 3LL-M27 cells (a carcinoma cell line overexpressing Pgp) were transfected in order to encode fluorescent proteins. As such, tumoral growth can be assessed via fluorescence imaging techniques [12].
The mice were injected intravenously with the modified 3LL-M27 cells. The carcinoma cells were allowed to establish for 48 h. To begin the treatment, the mice were split through different groups, where saline, free PTX and with the exosome-PTX system were administered, respectively. Tumour progression was accompanied by monitoring and quantifying the chemiluminescent signal emitted by the modified carcinoma cells. In vivo images show the progression of the metastasis in the different treatment groups (Figure 7a). 22 days after the administration of the 3LL-M27 cells, the mice were sacrificed. Lung sections were observed via confocal microscopy (Figure 7b). Mice administered with saline present the biggest tumour growth. Both free PTX and exosome-PTX complex inhibit tumour progression. When comparing PTX based treatments, the exosome-PTX complex proved to be more effective at stopping metastasis progression when compared with free PTX [12].
5.2. siRNA
siRNA belongs to a class of small, double stranded, non-coding RNAs, composed of 20–30 nucleotides. Through a mechanism of RNA interference, these molecules can target complementary mRNA. This way, it causes mRNA degradation and subsequent gene silencing [90].
Due to their mechanism and the possibility of exosome loading, exosome-siRNA complexes are emerging as a therapeutic agent in oncologic conditions. Due to an uncontrolled tumour growth and metastasis, head and neck cancer (HNC) still has a poor prognosis [13]. One of the reasons that make HNC so malignant is the epithelial-mesenchymal transition (EMT). EMT is a transforming process that some epithelial cells go through, leading to the formation of mesenchymal cells. EMT is usually associated with tumour growth and cancer progression. Cancer cells become more invasive and form metastases easier [91]. The transient receptor potential polycystic 2 (TRPP2), an ion channel, is one of the regulating mechanisms of EMT in HCN. Targeting TRPP2 could be a way to inhibit tumour progression in HNC, which was the goal of a developed study by Wang et al. [13].
Wang et al. investigated if TRPP2 siRNA would have any effect in TRPP2 gene knockdown and further influence HNC growth and metastasis. To achieve this, a TRPP2 siRNA-exosome complex was prepared and used. Exosomes were obtained from HEK 293 cells using PEG as the precipitating agent (a centrifugation was previously performed to remove cells and cell debris). Exosomes were incubated with the TRPP2 siRNA, with the latter being loaded and encapsulated. siRNA is susceptible to the action of many enzymes, such as nucleases, so it’s fundamental that exosomes provide some sort of protection to the cargo they carry. To assess this, an agarose gel electrophoresis was performed to evaluate the stability of free TRPP2 siRNA and the TRPP2 siRNA-exosome complex against RNA nucleases (see Figure 8). Free TRPP2 siRNA was degraded after 5 min, whereas exosome-encapsulated TRPP2 siRNA maintained stability, proving that exosomes shield siRNA from enzyme degradation [13].
FaDu cells, a human squamous carcinoma cell line, were used to evaluate TRPP2 siRNA-exosome complex efficiency. The cells were incubated with free TRPP2 siRNA (used as the control) and with the exosomes. Western blot analysis was performed to confirm if TRPP2 suppression occurred. The results indicated that there was a significant reduction of TRPP2 expression in FaDu cells treated with the TRPP2 siRNA-exosomes (see Figure 9). Since TRPP2 is associated with EMT, the authors of the study also confirmed the influence that the treatment had in this process. Common biomarkers of EMT are E-cadherin (at low levels), vimentin and N-cadherin (both with increased levels). For these biomarkers, western blot analysis was also performed, all of them with promising results (Figure 9). There was an increase in the expression of E-cadherin and a reduction of both vimentin and N-cadherin, showing the potential of TRPP2 siRNA-exosomes as a treatment option for HNC [13].
5.3. Doxorubicin-Loaded Nanoparticles
DOX belongs to anthracyclines, a class of drugs routinely used in chemotherapy. Its use is recommended in the treatment of several cancers (ovarian, breast, lung, Hodgkin’s lymphoma). The way DOX fights cancer cells has been described by two mechanisms. One occurs in the nucleus of the cell, where DOX intercalates itself with DNA, impairing the activity of topoisomerase-II and blocking nucleic acid transcription. The other way involves oxidation of DOX into semiquinone, with posterior transformation into DOX again. This process generates reactive oxygen species that cause oxidative stress and cell membrane damage, eventually causing cell death [92]. Besides MDR caused by ABC transporters (like the previously mentioned Pgp), the use of DOX is limited due to the cardiotoxic effect it causes.
As previously mentioned, the use of nano-based technologies in drug delivery seems like an almost perfect solution. However, exogenous particles have the disadvantage of being recognized and eliminated by the immune system [63]. To overcome this, Yong et al. developed biomimetic NPs, converging the efficiency of NPs with the endogenous benefits of exosomes [14].
Porous silicon NPs were loaded with DOX. The choice of these NPs was based on their biocompatibility and drug loading capacity. Afterwards, the DOX-loaded NPs were incubated with H22 cells, a mouse hepatocellular carcinoma cell line. Exosomes were obtained by centrifugation followed by differential centrifugation. Cells were able to incorporate DOX-NPs and release DOX-NP-Exosomes. The same procedure was applied to other cell lines, originating DOX-NP-Exosomes from different origins [14].
Another cause of MDR is the existence of cancer stem cells (CSC). This cell population has high expressions of ABC transporters and an elevated self-renewal rate. These properties render most treatments ineffective. In this sense, H22 CSCs were treated with DOX-NP-Exosomes to evaluate the cytotoxic effectiveness of the nano-complex. The procedure was done with different groups, containing free DOX, DOX-NPs and DOX-NP-Exosomes, respectively. The results are represented in Figure 10. In comparison with the other methods, there is an accentuated reduction of H22 CSCs in cells treated with the DOX-NP-Exosome complex [14].
To assess in vivo results, a model was used with H22 tumour-bearing mice. The mice were administered with different DOX formulations (free DOX, DOX-NPs and DOX-NP-Exosomes), with a drug concentration of 0.5 mg/kg. A fourth group was administered with free DOX at a higher concentration, namely 4 mg/kg. Free DOX and DOX-NPs (at a dosage of 0.5 mg/kg) showed weak tumour growth inhibiting capabilities. The most effective treatment was with DOX-NP-Exosomes, which proved to be even more effective than high dose of free DOX (4 mg/kg). This formulation achieved the biggest reduction in tumour mass (see Figure 11) and also increased mice survival time. Yong et al. developed an exosomal formulation, containing NPs loaded with a therapeutic agent. The presented drug delivery system proved itself to be biocompatible, not triggering an immune response. In terms of MDR, this system exhibits a higher tumour accumulation in comparison with other formulations (like free DOX) that suffer efflux by the action of ABC transporters [14].
6. Conclusions
MDR by cancer cells is one of the greatest threats to a positive outcome in oncological treatment. Although many resistance mechanisms have been acknowledged, most of them still remain without a successful solution. In this sense, exosome-based treatments shed a light on this area. As seen in the mentioned in vivo studies, exosome-containing formulations were more efficient in diminishing tumour cells when compared with conventional treatments (administration of free drug). In many studies, adverse reactions that are commonly associated with anticancer agents were much less frequent in mice groups treated with exosomes. However, these studies were only performed in animals, and the results obtained could not be applicable in humans.
In previous years, clinical trials have been performed to evaluate the effectiveness of vaccines containing exosomes in immunotherapy. In 2005, a Phase I Clinical Trial was performed using autologous exosomes derived from DCs, loaded with melanoma antigen gene (MAGE) proteins. This study aimed to test the safety and effectiveness of these exosomes on 13 patients with non-small cell lung cancer. It was possible to observe an increase of activity by NK cells, as well as an immune response against MAGE (determined by delayed type hypersensitivity test). Overall, there was a prolonged stabilization of the disease and only mild adverse reactions were reported (mostly, topical reactions related with the injection) [93]. Two years later, in 2007, a Phase I Clinical Trial was conducted covering 40 patients suffering from colorectal cancer. The exosomes used in this study were obtained from ascitic fluid. One group was administered with the exosomes alone, while another group was administered with a combination of exosomes and granulocyte-macrophage colony-stimulating factor (GM-CSF). The results showed that the groups inoculated with the GM-CSF adjuvant had a more efficient induction of a tumour-antigen specific response by cytotoxic T lymphocytes. Similar to the previously mentioned clinical trial, the reported adverse effects were related with injection site reactions, and some patients claimed to feel fatigued [94]. In addition to the previously mentioned trials, other studies took place to assess exosome safety and efficiency in cancer therapies. In recent years, more clinic trials have been registered and are still recruiting candidates (Table 2). A Phase I clinical trial is currently active, with seven patients enrolled. The aim is to deliver curcumin conjugated with plant-derived exosomes to normal colon tissue and colon tumours. The effects of curcumin on both normal cells and cancerous colon cells are to be evaluated by measuring biomarkers and serum cytokine levels. In another Phase I study, grape-derived exosomes are being used to prevent oral mucositis caused by chemoradiation treatments of HNC. More recently, a Phase I active trial is identifying the maximum tolerated dose and dose-limiting toxicities of exosomes loaded with siRNA against KRAS G12D for the treatment of metastatic pancreatic cancer [95].
Also, other than to treat oncologic diseases, there are trials focused on the treatment of other illnesses, like those presenting a chronic character (for example, chronic kidney disease or type 1 diabetes) [96].
There is still much to be learned about the biogenesis of exosomes. It is known that the ESCRT is involved, as well as several ESCRT-independent methods. However, there are many in-between steps that are still unexplained and require further studies. To fully understand the processes that are behind the formation of these nanovesicles would be a great advantage. Furthermore, most laboratorial techniques for exosome isolation suffer from low yields, so a better knowledge of the nature of exosomes could help develop new techniques aiming to improve exosome production and collection.
Until now, there have been no approved products containing exosomes. Hopefully, that paradigm will change in the following years. Many studies concerning exosomes have proved their role in stimulating the immune system and their ability to load therapeutic molecules. Recent reports have tried to develop new formulations based on the characteristics of exosomes. With positive results in several investigations, it is undeniable that exosomes stand as a promising multiparametric nano-approach for cancer therapy.
Author Contributions
P.R. wrote the paper; C.M. analyzed and complete the paper; F.V. and A.F. revised the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This work received financial support from National Funds (FCT/MEC, Fundação para a Ciência e Tecnologia/Ministério da Educação e Ciência) through project UID/QUI/50006/2013, co-financed by European Union (FEDER under the Partnership Agreement PT2020). It was also supported by the grant FCT PTDC/BTM-MAT/30255/2017 (POCI-01- 0145-FEDER-030255) from the Portuguese Foundation for Science and Technology (FCT) and the European Community Fund (FEDER) through the COMPETE2020 program.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Anand, P.; Kunnumakara, A.B.; Sundaram, C.; Harikumar, K.B.; Tharakan, S.T.; Lai, O.S.; Sung, B.; Aggarwal, B.B. Cancer is a preventable disease that requires major lifestyle changes. Pharm. Res. 2008, 25, 2097–2116. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Horn, G.; Moulton, K.; Oza, A.; Byler, S.; Kokolus, S.; Longacre, M. Cancer development, progression, and therapy: An epigenetic overview. Int. J. Mol. Sci. 2013, 14, 21087–21113. [Google Scholar] [CrossRef] [PubMed]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The different mechanisms of cancer drug resistance: A brief review. Adv. Pharm. Bull. 2017, 7, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 1–18. [Google Scholar] [CrossRef]
- Johnstone, R.M.; Adam, M.; Hammond, J.R.; Orr, L.; Turbide, C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 1987, 262, 9412–9420. [Google Scholar]
- Pan, B.T.; Johnstone, R.M. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: Selective externalization of the receptor. Cell 1983, 33, 967–978. [Google Scholar] [CrossRef]
- Vlassov, A.V.; Magdaleno, S.; Setterquist, R.; Conrad, R. Exosomes: Current knowledge of their composition, biological functions, and diagnostic and therapeutic potentials. Biochim. Biophys. Acta Gen. Subj. 2012, 1820, 940–948. [Google Scholar] [CrossRef]
- Kowal, J.; Tkach, M.; Théry, C. Biogenesis and secretion of exosomes. Curr. Opin. Cell Biol. 2014, 29, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory Mechanisms and Intercellular Transfer of MicroRNAs in Living Cells. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar] [CrossRef] [Green Version]
- Li, S.P.; Lin, Z.-X.; Jiang, X.-Y.; Yu, X.-Y. Exosomal cargo-loading and synthetic exosome-mimics as potential therapeutic tools. Acta Pharmacol. Sin. 2018, 39, 542–551. [Google Scholar] [CrossRef] [Green Version]
- Stuffers, S.; Wegner, C.S.; Stenmark, H.; Brech, A. Multivesicular endosome biogenesis in the absence of ESCRTs. Traffic 2009, 10, 925–937. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Haney, M.J.; Zhao, Y.; Mahajan, V.; Deygen, I.; Klyachko, N.L.; Inskoe, E.; Piroyan, A.; Sokolsky, M.; Okolie, O.; et al. Development of exosome-encapsulated paclitaxel to overcome MDR in cancer cells. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 655–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Chen, L.; Huang, Y.; Li, K.; Jinye, A.; Fan, T.; Zhao, R.; Xia, X.; Shen, B.; Du, J.; et al. Exosome-delivered TRPP2 siRNA inhibits the epithelial-mesenchymal transition of FaDu cells. Oncol. Lett. 2019, 17, 1953–1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, T.; Zhang, X.; Bie, N.; Zhang, H.; Zhang, X.; Li, F.; Hakeem, A.; Hu, J.; Gan, L.; Santos, H.A.; et al. Tumor exosome-based nanoparticles are efficient drug carriers for chemotherapy. Nat. Commun. 2019, 10, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, Secretion, and Intercellular Interactions of Exosomes and Other Extracellular Vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Tamkovich, S.N.; Tutanov, O.S.; Laktionov, P.P. Exosomes: Generation, structure, transport, biological activity, and diagnostic application. Biochem. Suppl. Ser. A Membr. Cell Biol. 2016, 10, 163–173. [Google Scholar] [CrossRef]
- Frydrychowicz, M.; Kolecka-Bednarczyk, A.; Madejczyk, M.; Yasar, S.; Dworacki, G. Exosomes—Structure, biogenesis and biological role in non-small-cell lung cancer. Scand. J. Immunol. 2015, 81, 2–10. [Google Scholar] [CrossRef]
- Théry, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef]
- Xiao, C.; Song, F.; Zheng, Y.L.; Lv, J.; Wang, Q.F.; Xu, N. Exosomes in Head and Neck Squamous Cell Carcinoma. Front. Oncol. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Ha, D.; Yang, N.; Nadithe, V. Exosomes as therapeutic drug carriers and delivery vehicles across biological membranes: Current perspectives and future challenges. Acta Pharm. Sin. B 2016, 6, 287–296. [Google Scholar] [CrossRef] [Green Version]
- ExoCarta—Exosome Protein, RNA and Lipid Database. Available online: http://www.exocarta.org/ (accessed on 28 April 2020).
- Conde-Vancells, J.; Rodriguez-Suarez, E.; Embade, N.; Gil, D.; Matthiesen, R.; Valle, M.; Elortza, F.; Lu, S.C.; Mato, J.M.; Falcón-Pérez, J.M. Characterization and comprehensive proteome profiling of exosomes secreted by hepatocytes. J. Proteome Res. 2008, 7, 5157–5166. [Google Scholar] [CrossRef] [Green Version]
- Février, B.; Raposo, G. Exosomes: Endosomal-derived vesicles shipping extracellular messages. Curr. Opin. Cell Biol. 2004, 16, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Andreu, Z.; Yáñez-mó, M. Tetraspanins in extracellular vesicle formation and function. Front. Immunol. 2014, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Charrin, S.; le Naour, F.; Silvie, O.; Milhiet, P.-E.; Boucheix, C.; Rubinstein, E. Lateral organization of membrane proteins: Tetraspanins spin their web. Biochem. J. 2009, 420, 133–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemler, M.E. Tetraspanin Proteins Mediate Cellular Penetration, Invasion, and Fusion Events and Define a Novel Type of Membrane Microdomain. Annu. Rev. Cell Dev. Biol. 2003, 19, 397–422. [Google Scholar] [CrossRef] [PubMed]
- Clayton, A.; Turkes, A.; Navabi, H.; Mason, M.D.; Tabi, Z. Induction of heat shock proteins in B-cell exosomes. J. Cell Sci. 2005, 118, 3631–3638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponomarenko, M.; Stepanenko, I.; Kolchanov, N. Heat Shock Proteins. In Brenner’s Encyclopedia of Genetics, 2nd ed.; Maloy, S., Hughes, K., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 402–405. [Google Scholar]
- Reddy, V.S.; Madala, S.K.; Trinath, J.; Reddy, G.B. Extracellular small heat shock proteins: Exosomal biogenesis and function. Cell Stress Chaperones 2018, 23, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Nijman, H.W.; Stoorvogel, W.; Liejendekker, R.; Harding, C.V.; Melief, C.J.; Geuze, H.J. B Lymphocytes Secrete Antigen-Presenting Vesicles. J. Exp. Med. 1996, 183, 1161–1172. [Google Scholar] [CrossRef]
- Bobrie, A.; Colombo, M.; Raposo, G.; Théry, C. Exosome Secretion: Molecular Mechanisms and Roles in Immune Responses. Traffic 2011, 12, 1659–1668. [Google Scholar] [CrossRef]
- Skotland, T.; Hessvik, N.P.; Sandvig, K.; Llorente, A. Exosomal lipid composition and the role of ether lipids and phosphoinositides in exosome biology. J. Lipid Res. 2019, 60, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Hemler, M.E. Tetraspanin functions and associated microdomains. Nat. Rev. Mol. Cell Biol. 2005, 6, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Subra, C.; Laulagnier, K.; Perret, B.; Record, M. Exosome lipidomics unravels lipid sorting at the level of multivesicular bodies. Biochimie 2007, 89, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Xu, Q. Functions and applications of exosomes. Acta Pol. Pharm.—Drug Res. 2014, 71, 537–543. [Google Scholar]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef]
- Gusachenko, O.N.; Zenkova, M.A.; Vlassov, V.V. Nucleic acids in exosomes: Disease markers and intercellular communication molecules. Biochemistry 2013, 78, 1–7. [Google Scholar] [CrossRef]
- Melo, S.A.; Sugimoto, H.; O’Connell, J.T.; Kato, N.; Villanueva, A.; Vidal, A.; Qiu, L.; Vitkin, E.; Perelman, L.T.; Melo, C.A.; et al. Cancer Exosomes Perform Cell-Independent MicroRNA Biogenesis and Promote Tumorigenesis. Cancer Cell 2014, 26, 707–721. [Google Scholar] [CrossRef] [Green Version]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Tvall, J.O.L.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Kalluri, R.; Lebleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Stoorvogel, W.; Kleijmeer, M.J.; Geuze, H.J.; Raposo, G. The biogenesis and functions of exosomes. Traffic 2002, 3, 321–330. [Google Scholar] [CrossRef]
- Hanson, P.I.; Cashikar, A. Multivesicular Body Morphogenesis. Annu. Rev. Cell Dev. Biol. 2012, 28, 337–362. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Henne, W.M.; Buchkovich, N.J.; Emr, S.D. The ESCRT Pathway. Dev. Cell 2011, 21, 77–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurley, J.H. The ESCRT complexes. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 463–487. [Google Scholar] [CrossRef] [PubMed]
- Wollert, T.; Hurley, J.H. Molecular mechanism of multivesicular body biogenesis by ESCRT complexes. Nature 2010, 464, 864–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babst, M. A Protein’s Final ESCRT. Traffic 2005, 6, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Katzmann, D.J.; Odorizzi, G.; Emr, S.D. Receptor downregulation and multivesicular-body sorting. Nat. Rev. Mol. Cell Biol. 2002, 3, 893–905. [Google Scholar] [CrossRef] [PubMed]
- Pols, M.S.; Klumperman, J. Trafficking and function of the tetraspanin CD63. Exp. Cell Res. 2009, 315, 1584–1592. [Google Scholar] [CrossRef]
- Theos, A.C.; Truschel, S.T.; Tenza, D.; Hurbain, I.; Harper, D.C.; Berson, J.F.; Thomas, P.C.; Raposo, G.; Marks, M.S. A lumenal domain-dependent pathway for sorting to intralumenal vesicles of multivesicular endosomes involved in Organelle Morphogenesis. Dev. Cell 2006, 10, 343–354. [Google Scholar] [CrossRef] [Green Version]
- van Niel, G.; Charrin, S.; Simoes, S.; Romao, M.; Rochin, L.; Saftig, P.; Marks, M.S.; Rubinstein, E.; Raposo, G. The Tetraspanin CD63 Regulates ESCRT-Independent and -Dependent Endosomal Sorting during Melanogenesis. Dev. Cell 2011, 21, 708–721. [Google Scholar] [CrossRef] [Green Version]
- Trajkovic, K. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 2008, 319, 1244–1247. [Google Scholar] [CrossRef]
- Ostrowski, M.; Carmo, N.B.; Krumeich, S.; Fanget, I.; Raposo, G.; Savina, A.; Moita, C.F.; Schauer, K.; Hume, A.N.; Freitas, R.P.; et al. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat. Cell Biol. 2010, 12, 19–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, S.K.; Huynh, C.; Proux-Gillardeaux, V.; Galli, T.; Andrews, N.W. Identification of SNAREs Involved in Synaptotagmin VII-regulated Lysosomal Exocytosis. J. Biol. Chem. 2004, 279, 20471–20479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fader, C.M.; Sánchez, D.G.; Mestre, M.B.; Colombo, M.I. TI-VAMP/VAMP7 and VAMP3/cellubrevin: Two v-SNARE proteins involved in specific steps of the autophagy/multivesicular body pathways. Biochim. Biophys. Acta Mol. Cell Res. 2009, 1793, 1901–1916. [Google Scholar] [CrossRef] [Green Version]
- Proux-Gillardeaux, V.; Raposo, G.; Irinopoulou, T.; Galli, T. Expression of the Longin domain of TI-VAMP impairs lysosomal secretion and epithelial cell migration. Biol. Cell 2007, 99, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.C.; Kim, O.Y.; Yoon, C.M.; Choi, D.-S.; Roh, T.-Y.; Park, J.; Nilsson, J.; Lötvall, J.; Kim, Y.-K.; Gho, Y.S. Bioinspired exosome-mimetic nanovesicles for targeted delivery of chemotherapeutics to malignant tumors. ACS Nano 2013, 7, 7698–7710. [Google Scholar] [CrossRef]
- Peer, D.; Karp, J.M.; Hong, S.; Farokhzad, O.C.; Margalit, R.; Langer, R. Nanocarriers as an emerging platform for cancer therapy. Nat. Nanotechnol. 2007, 2, 751–760. [Google Scholar] [CrossRef]
- Batrakova, E.V.; Kim, M.S. Using exosomes, naturally-equipped nanocarriers, for drug delivery. J. Control. Release 2015, 219, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Trelis, M.; Galiano, A.; Bolado, A.; Toledo, R.; Marcilla, A.; Bernal, D. Subcutaneous injection of exosomes reduces symptom severity and mortality induced by Echinostoma caproni infection in BALB/c mice. Int. J. Parasitol. 2016, 46, 799–808. [Google Scholar] [CrossRef]
- Betker, J.L.; Angle, B.M.; Graner, M.W.; Anchordoquy, T.J. The potential of exosomes from cow milk for oral delivery. J. Pharm. Sci. 2019, 108, 1496–1505. [Google Scholar] [CrossRef]
- Jong, B.; Barros, E.R.; Hoenderop, J.G.J.; Rigalli, J.P. Recent Advances in Extracellular Vesicles as Drug Delivery Systems and Their Potential in Precision Medicine. Pharmaceutics 2020, 12, 1006. [Google Scholar] [CrossRef]
- Luan, X.; Sansanaphongpricha, K.; Myers, I.; Chen, H.; Yuan, H.; Sun, D. Engineering exosomes as refined biological nanoplatforms for drug delivery. Acta Pharmacol. Sin. 2017, 38, 754–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; A Wood, M.J. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol. 2011, 29, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Naseri, M.; Bozorgmehr, M.; Zöller, M.; Pirmardan, E.R.; Madjd, Z. Tumor-derived exosomes: The next generation of promising cell-free vaccines in cancer immunotherapy. OncoImmunology 2020, 9, 1779991. [Google Scholar] [CrossRef] [PubMed]
- Gomez, C.T.; Goreham, R.V.; Serra, J.J.B.; Nann, T.; Kussmann, M. ‘Exosomics’—A review of biophysics, biology and biochemistry of exosomes with a focus on human breast milk. Front. Genet. 2018, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Mathew, M.; Zade, M.; Mezghani, N.; Patel, R.; Wang, Y.; Momen-Heravi, F. Extracellular Vesicles as Biomarkers in Cancer Immunotherapy. Cancers 2020, 12, 2825. [Google Scholar] [CrossRef]
- Tang, X.J.; Sun, X.-Y.; Huang, K.-M.; Zhang, L.; Yang, Z.-S.; Zou, D.-D.; Wang, B.; Warnock, G.L.; Dai, L.-J.; Luo, J. Therapeutic potential of CAR-T cell-derived exosomes: A cell-free modality for targeted cancer therapy. Oncotarget 2015, 6, 44179–44190. [Google Scholar] [CrossRef] [Green Version]
- Dutta, A. Exosomes-based cell-free cancer therapy: A novel strategy for targeted therapy. Immunol. Med. 2020, 41, 1–8. [Google Scholar] [CrossRef]
- Lobb, R.J.; Becker, M.; Wen, S.W.; Wong, C.S.F.; Wiegmans, A.P.; Leimgruber, A.; Möller, A. Optimized exosome isolation protocol for cell culture supernatant and human plasma. J. Extracell. Vesicles 2015, 4, 27031. [Google Scholar] [CrossRef]
- Yu, L.L.; Zhu, J.; Liu, J.-X.; Jiang, F.; Ni, W.-K.; Qu, L.-S.; Ni, R.-Z.; Lu, C.-H.; Xiao, M. A Comparison of Traditional and Novel Methods for the Separation of Exosomes from Human Samples. BioMed Res. Int. 2018, 2018, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kamerkar, S.; Lebleu, V.S.; Sugimoto, H.; Yang, S.; Ruivo, C.F.; Melo, S.A.; Lee, J.J.; Kalluri, R. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature 2017, 546, 498–503. [Google Scholar] [CrossRef]
- Li, P.; Kaslan, M.; Lee, S.H.; Yao, J.; Gao, Z. Progress in exosome isolation techniques. Theranostics 2017, 7, 789–804. [Google Scholar] [CrossRef] [PubMed]
- Szatanek, R.; Baran, J.; Siedlar, M.; Baj-Krzyworzeka, M. Isolation of extracellular vesicles: Determining the correct approach (Review). Int. J. Mol. Med. 2015, 36, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantin, R.; Diou, J.; Bélanger, D.; Tremblay, A.M.; Gilbert, C. Discrimination between exosomes and HIV-1: Purification of both vesicles from cell-free supernatants. J. Immunol. Methods 2008, 338, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Tauro, B.J.; Greening, D.W.; Mathias, R.A.; Ji, H.; Mathivanan, S.; Scott, A.M.; Simpson, R.J. Comparison of ultracentrifugation, density gradient separation, and immunoaffinity capture methods for isolating human colon cancer cell line LIM1863-derived exosomes. Methods 2012, 56, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Mathivanan, S.; Lim, J.W.E.; Tauro, B.J.; Ji, H.; Moritz, R.L.; Simpson, R.J. Proteomics analysis of A33 immunoaffinity-purified exosomes released from the human colon tumor cell line LIM1215 reveals a tissue-specific protein signature. Mol. Cell. Proteom. 2010, 9, 197–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konoshenko, M.Y.; Lekchnov, E.A.; Vlassov, A.V.; Laktionov, P.P. Isolation of Extracellular Vesicles: General Methodologies and Latest Trends. BioMed Res. Int. 2018, 2018, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Liang, K.; Liu, F.; Fan, J.; Sun, D.; Liu, C.; Lyon, C.J.; Bernard, D.W.; Li, Y.; Yokoi, K.; Katz, M.H.; et al. Nanoplasmonic quantification of tumour-derived extracellular vesicles in plasma microsamples for diagnosis and treatment monitoring. Nat. Biomed. Eng. 2017, 1, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antimisiaris, S.G.; Mourtas, S.; Marazioti, A. Exosomes and exosome-inspired vesicles for targeted drug delivery. Pharmaceutics 2018, 10, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahlgren, J.; Karlson, T.D.L.; Brisslert, M.; Sani, F.V.; Telemo, E.; Sunnerhagen, P.; Valadi, H. Plasma exosomes can deliver exogenous short interfering RNA to monocytes and lymphocytes. Nucleic Acids Res. 2012, 40, e130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnsen, K.B.; Gudbergsson, J.M.; Skov, M.N.; Christiansen, G.; Gurevich, L.; Moos, T.; Duroux, M. Evaluation of electroporation-induced adverse effects on adipose-derived stem cell exosomes. Cytotechnology 2016, 68, 2125–2138. [Google Scholar] [CrossRef] [Green Version]
- Costa, A.P.; Xu, X.; Burgess, D.J. Freeze-anneal-thaw cycling of unilamellar liposomes: Effect on encapsulation efficiency. Pharm. Res. 2014, 31, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Fuhrmann, G.; Serio, A.; Mazo, M.; Nair, R.; Stevens, M.M. Active loading into extracellular vesicles significantly improves the cellular uptake and photodynamic effect of porphyrins. J. Control. Release 2015, 205, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Podolak, I.; Galanty, A.; Sobolewska, D. Saponins as cytotoxic agents: A review. Phytochem. Rev. 2010, 9, 425–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Chen, L. Progress in research on paclitaxel and tumor immunotherapy. Cell. Mol. Biol. Lett. 2019, 24, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Weaver, B.A. How Taxol/paclitaxel kills cancer cells. Mol. Biol. Cell 2014, 25, 2677–2681. [Google Scholar] [CrossRef] [PubMed]
- Fong, A.; Durkin, A.; Lee, H. The potential of combining tubulin-targeting anticancer therapeutics and immune therapy. Int. J. Mol. Sci. 2019, 20, 586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Song, L.; Liu, S.; Zhao, S.; Jiang, Q.; Ding, B. A Tailored DNA Nanoplatform for Synergistic RNAi-/Chemotherapy of Multidrug-Resistant Tumors. Angew. Chem. Int. Ed. 2018, 57, 15486–15490. [Google Scholar] [CrossRef]
- Dana, H.; Chalbatani, G.M.; Mahmoodzadeh, H.; Karimloo, R.; Rezaiean, O.; Moradzadeh, A.; Mehmandoost, N.; Moazzen, F.; Mazraeh, A.; Marmari, V.; et al. Molecular Mechanisms and Biological Functions of siRNA. Int. J. Biomed. Sci. 2017, 13, 48–57. [Google Scholar] [PubMed]
- Dave, B.; Mittal, V.; Tan, N.M.; Chang, J.C. Epithelial-mesenchymal transition, cancer stem cells and treatment resistance. Breast Cancer Res. 2011, 14, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorn, C.F.; Oshiro, C.; Marsh, S.; Hernandez-Boussard, T.; McLeod, H.; Klein, T.E.; Altman, R.B. Doxorubicin pathways: Pharmacodynamics and adverse effects. Pharm. Genom. 2011, 21, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Morse, M.A.; Garst, J.; Osada, T.; Khan, S.; Hobeika, A.C.; Clay, T.M.; Valente, N.; Shreeniwas, R.; Sutton, M.A.; Delcayre, A.; et al. A phase I study of dexosome immunotherapy in patients with advanced non-small cell lung cancer. J. Transl. Med. 2005, 3, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, S.; Wei, D.; Wu, Z.; Zhou, X.; Wei, X.; Huang, H.; Li, G. Phase I clinical trial of autologous ascites-derived exosomes combined with GM-CSF for colorectal cancer. Mol. Ther. 2008, 16, 782–790. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov—Find Studies. Available online: https://clinicaltrials.gov/ct2/home (accessed on 3 October 2020).
- Chen, Y.S.; Harn, H.-J.; Lin, E.-Y.; Chiou, T.-W. Exosomes in clinical trial and their production in compliance with good manufacturing practice. Tzu Chi Med J. 2020, 32, 113–120. [Google Scholar] [CrossRef]
Figure 1.
Structure and overall composition of an exosome [15].
Figure 1.
Structure and overall composition of an exosome [15].


Figure 3.
Overall mechanism of the ESCRT during MVB sorting [48].
Figure 3.
Overall mechanism of the ESCRT during MVB sorting [48].

Figure 4.
Several machineries involved in the biogenesis of exosomes [8].
Figure 4.
Several machineries involved in the biogenesis of exosomes [8].

Figure 5.
Western blot results comparison between the nanovesicles (NV) and exosomes (EXO) [57].
Figure 5.
Western blot results comparison between the nanovesicles (NV) and exosomes (EXO) [57].

Figure 6.
Representation of how PTX promotes microtubule stabilization [88].
Figure 6.
Representation of how PTX promotes microtubule stabilization [88].

Figure 7.
(a) In vivo imaging of chemiluminescent signal monitoring in each treatment group [12]. (b) Confocal microscopy of lung sections of the sacrificed mice. It is possible to observe no detection of fluorescence in the Exosome-PTX treated cells, when compared to the non-treated control group (saline) [12].
Figure 7.
(a) In vivo imaging of chemiluminescent signal monitoring in each treatment group [12]. (b) Confocal microscopy of lung sections of the sacrificed mice. It is possible to observe no detection of fluorescence in the Exosome-PTX treated cells, when compared to the non-treated control group (saline) [12].

Figure 8.
Results of agarose gel electrophoresis. Each sample was incubated with RNA nucleases, free siRNA was mostly degraded after 5 min [13].
Figure 8.
Results of agarose gel electrophoresis. Each sample was incubated with RNA nucleases, free siRNA was mostly degraded after 5 min [13].

Figure 9.
Results of western blot analysis for TRPP2 expression and EMT biomarkers. Free siRNA was used as control [13].
Figure 9.
Results of western blot analysis for TRPP2 expression and EMT biomarkers. Free siRNA was used as control [13].

Figure 10.
H22 CSCs colony size after DOX administration, delivered as free drug, in NPs and NPs contained in exosomes. The experiments are performed in triplicate (mean±SD) with statistically significant difference (***) between groups [14].
Figure 10.
H22 CSCs colony size after DOX administration, delivered as free drug, in NPs and NPs contained in exosomes. The experiments are performed in triplicate (mean±SD) with statistically significant difference (***) between groups [14].

Figure 11.
Tumour masses of mice after intravenous administration of different formulations of DOX [14].
Figure 11.
Tumour masses of mice after intravenous administration of different formulations of DOX [14].


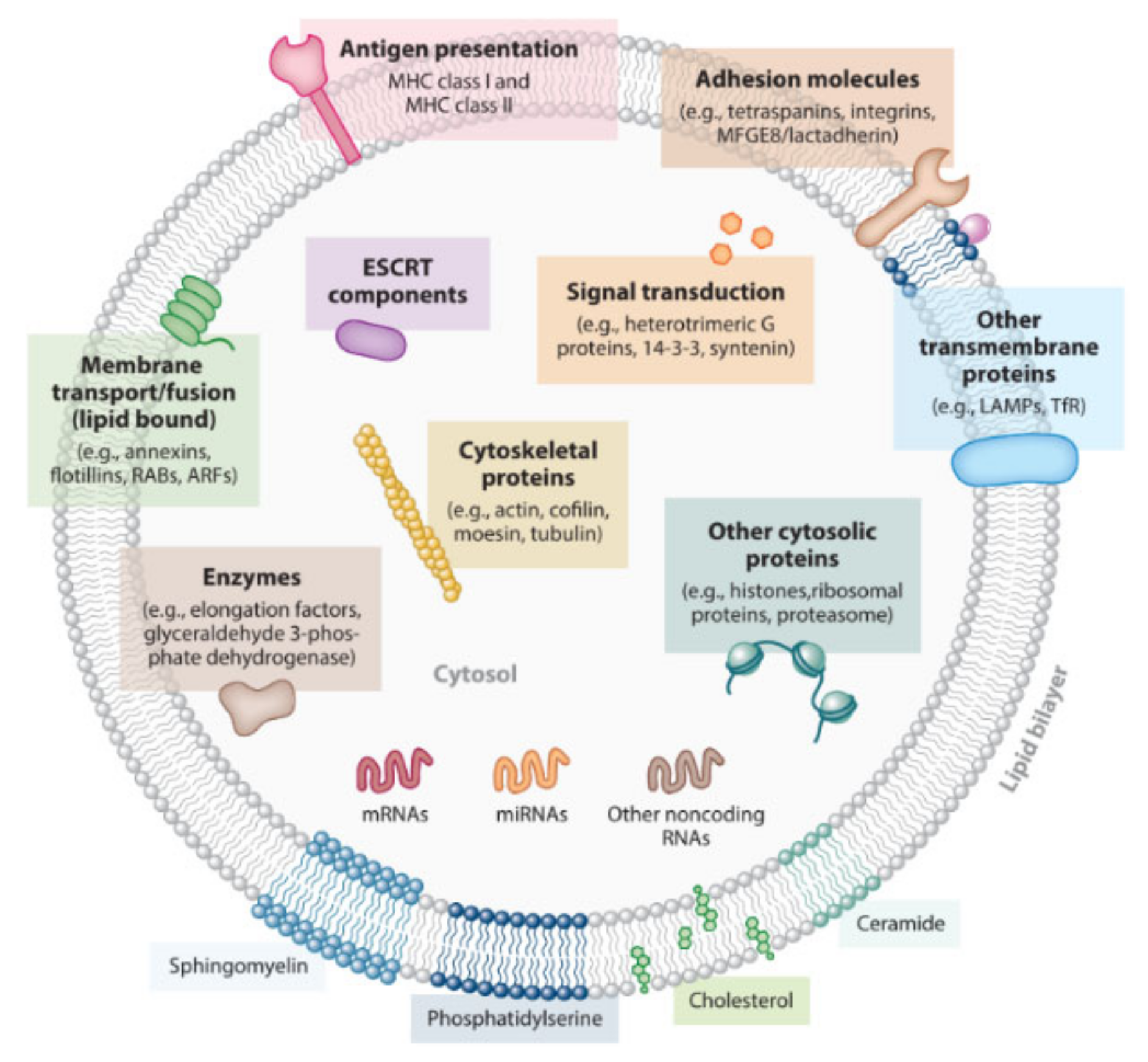{kind=link}
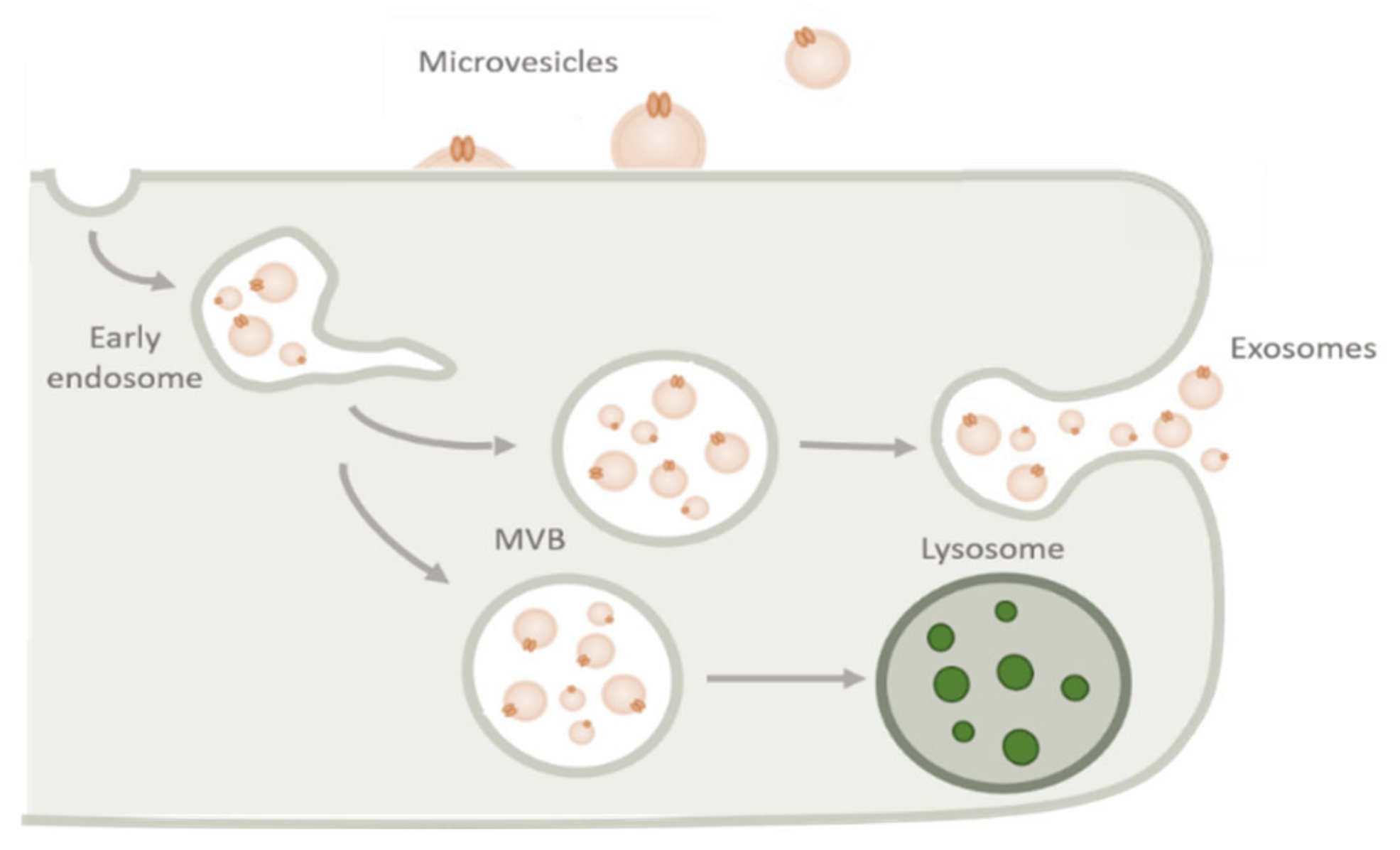{kind=link}
{kind=link}
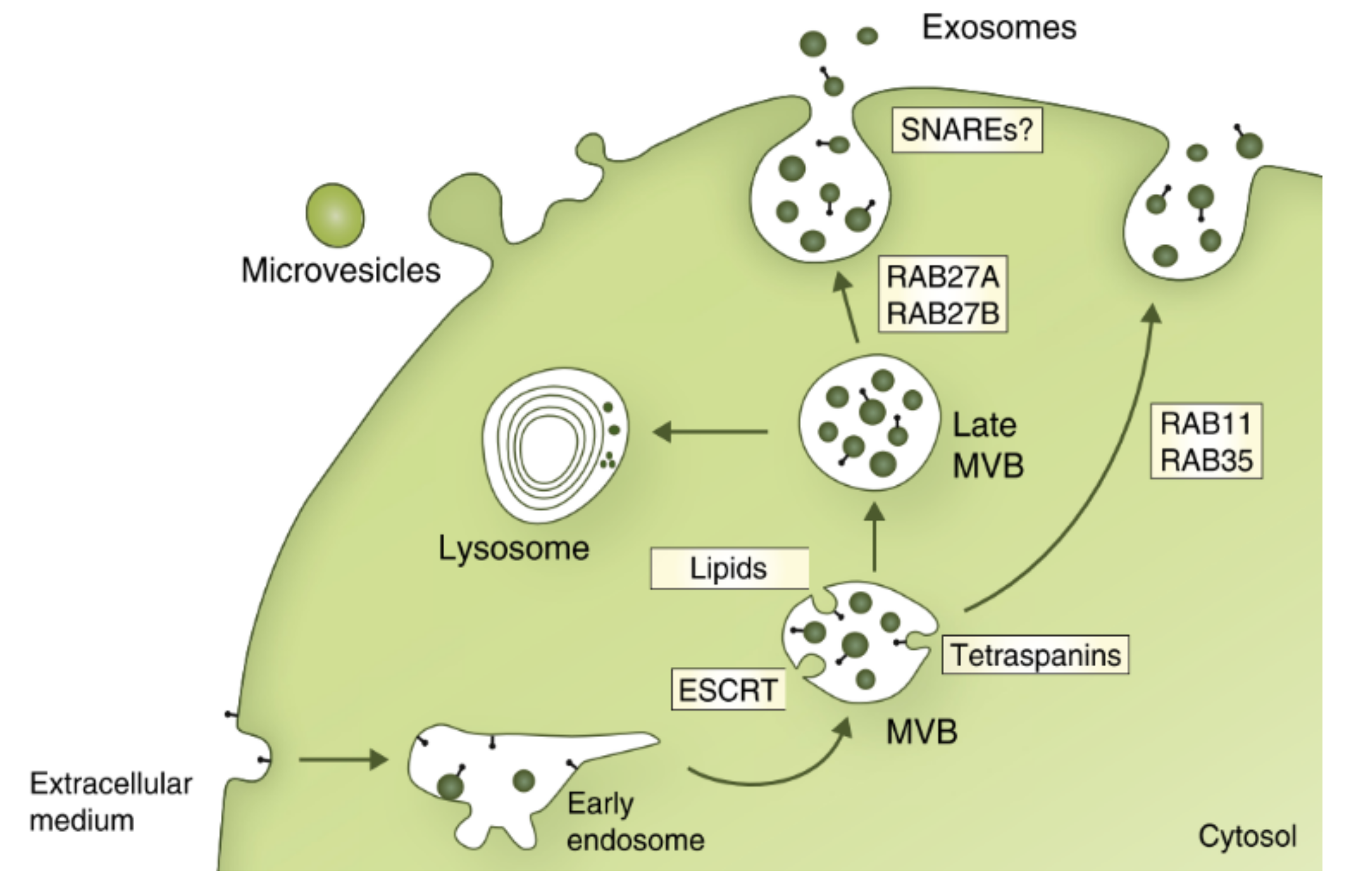{kind=link}
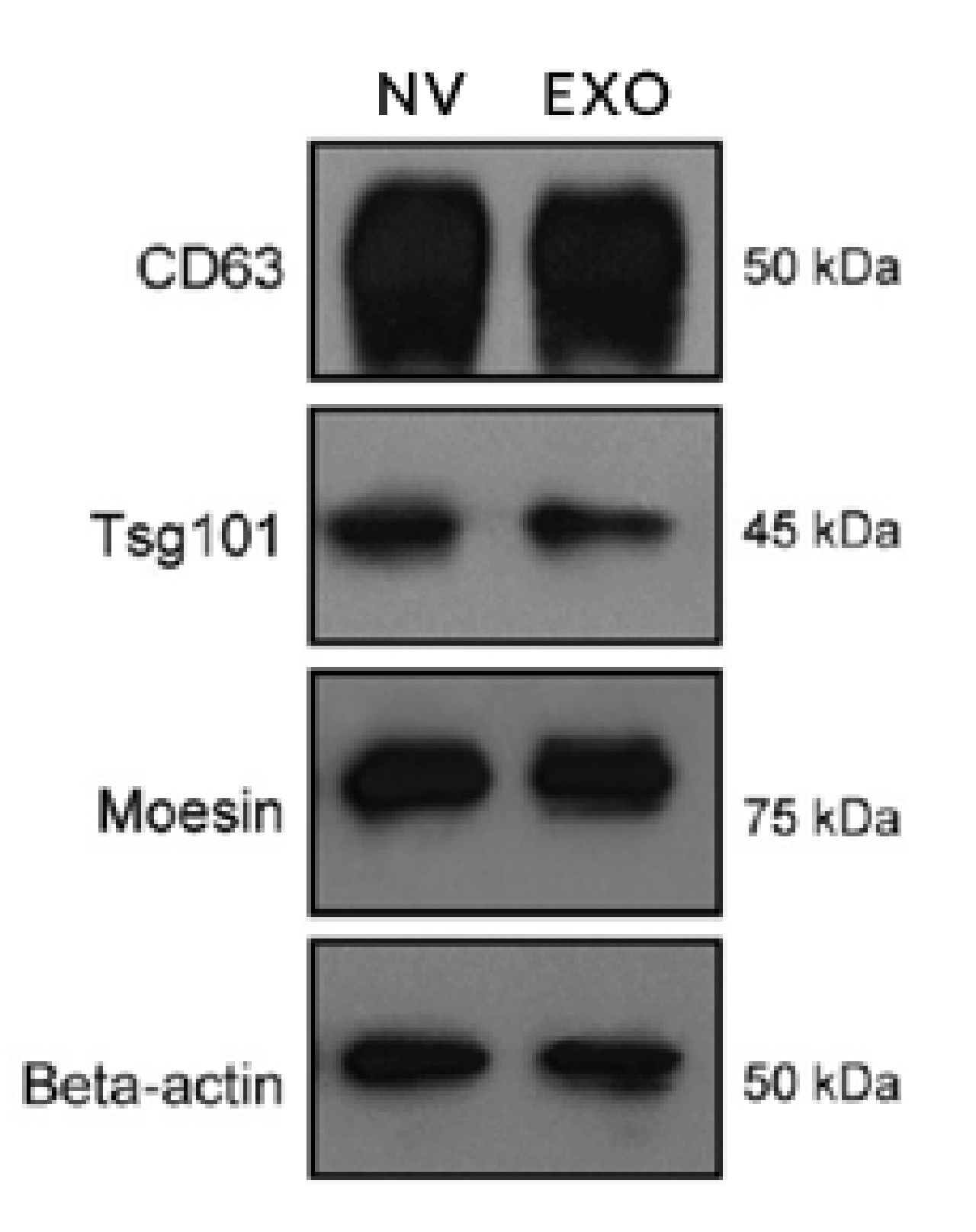{kind=link}
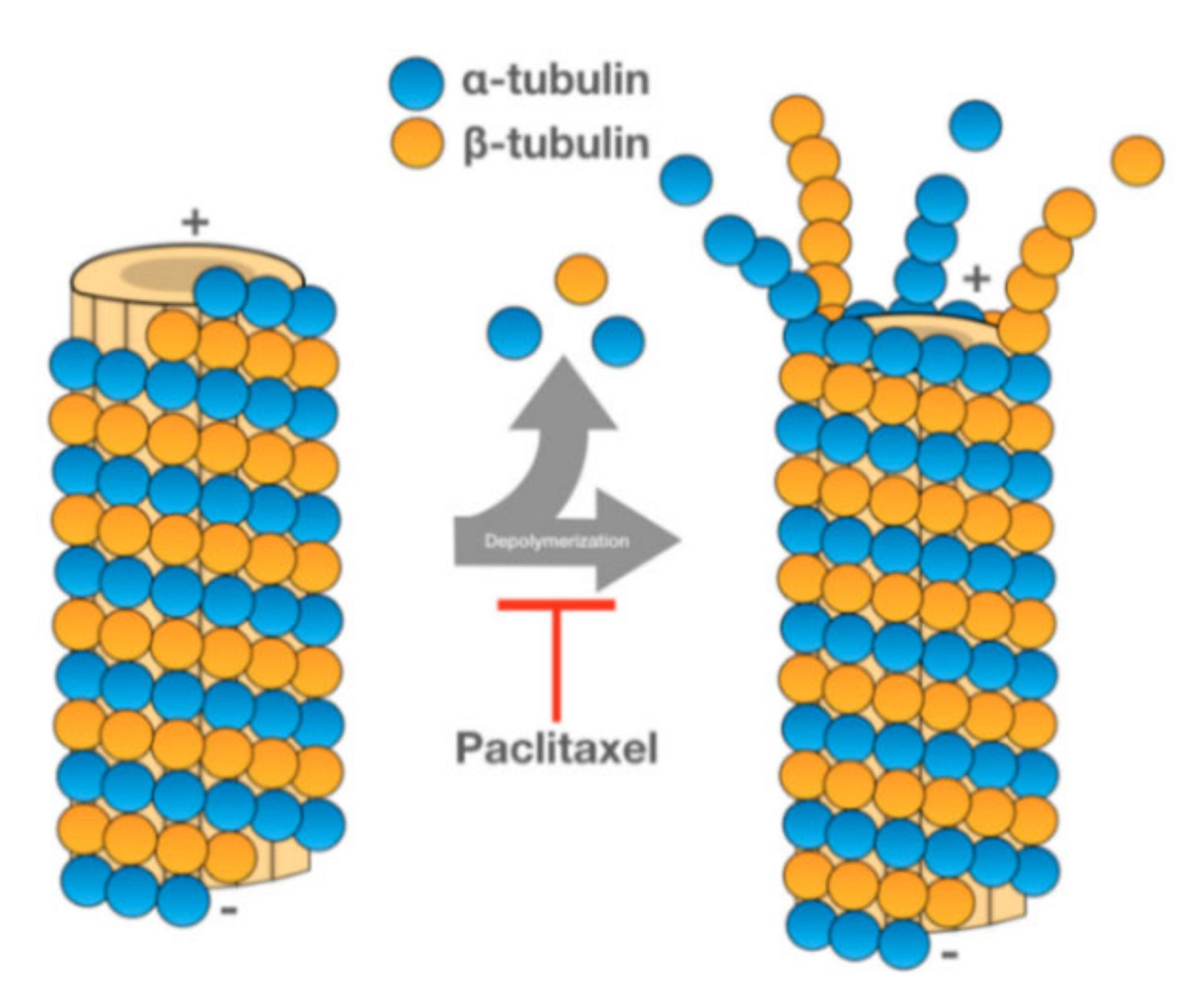{kind=link}
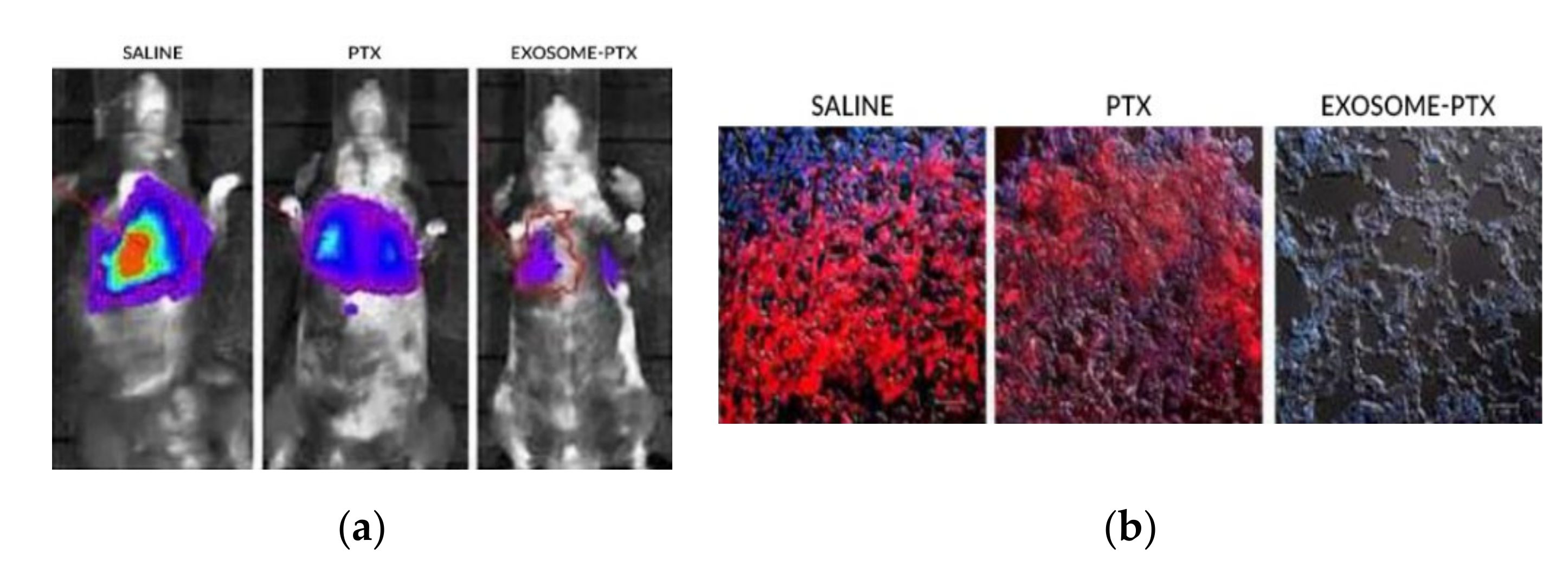{kind=link}
{kind=link}
{kind=link}
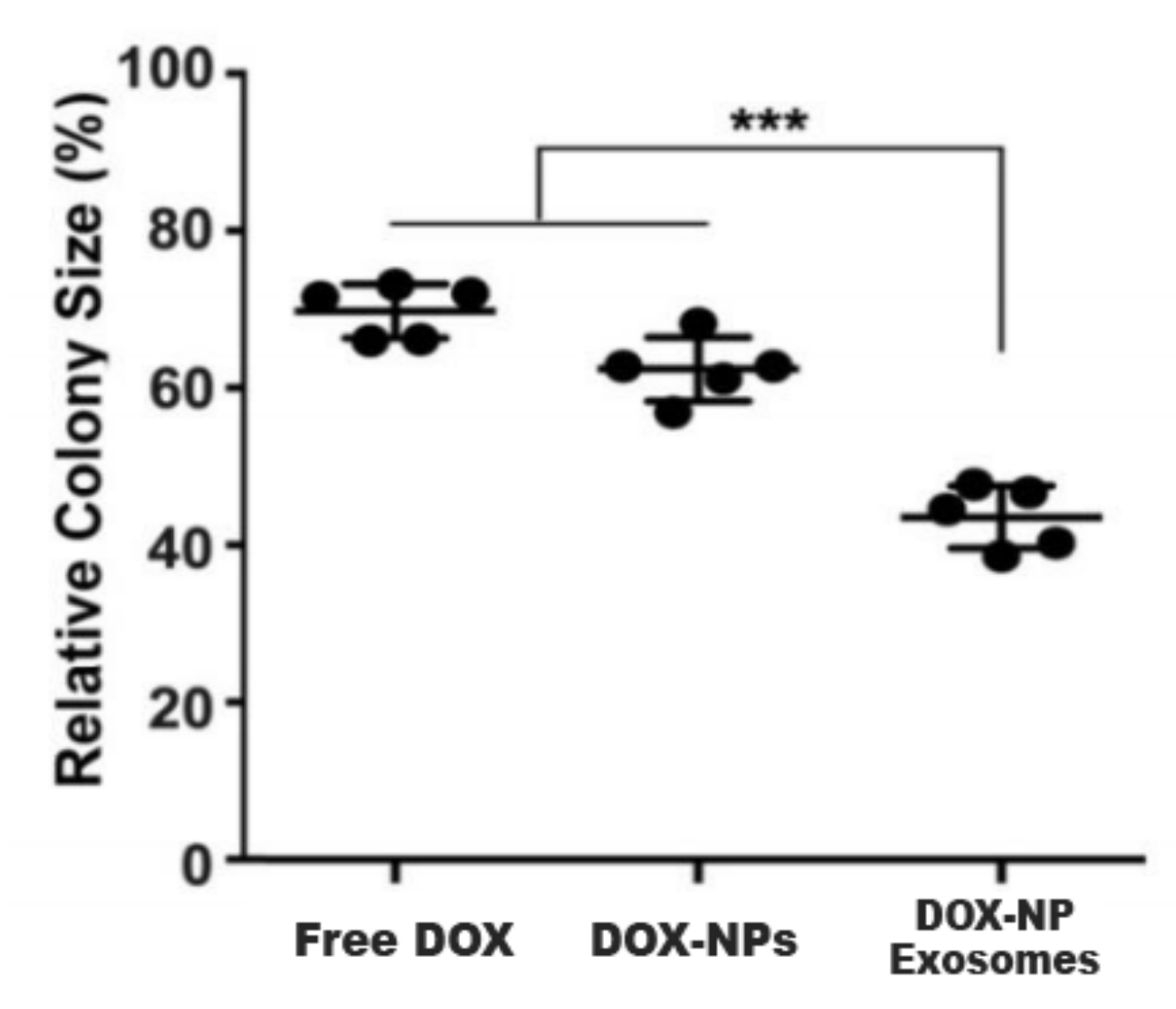{kind=link}
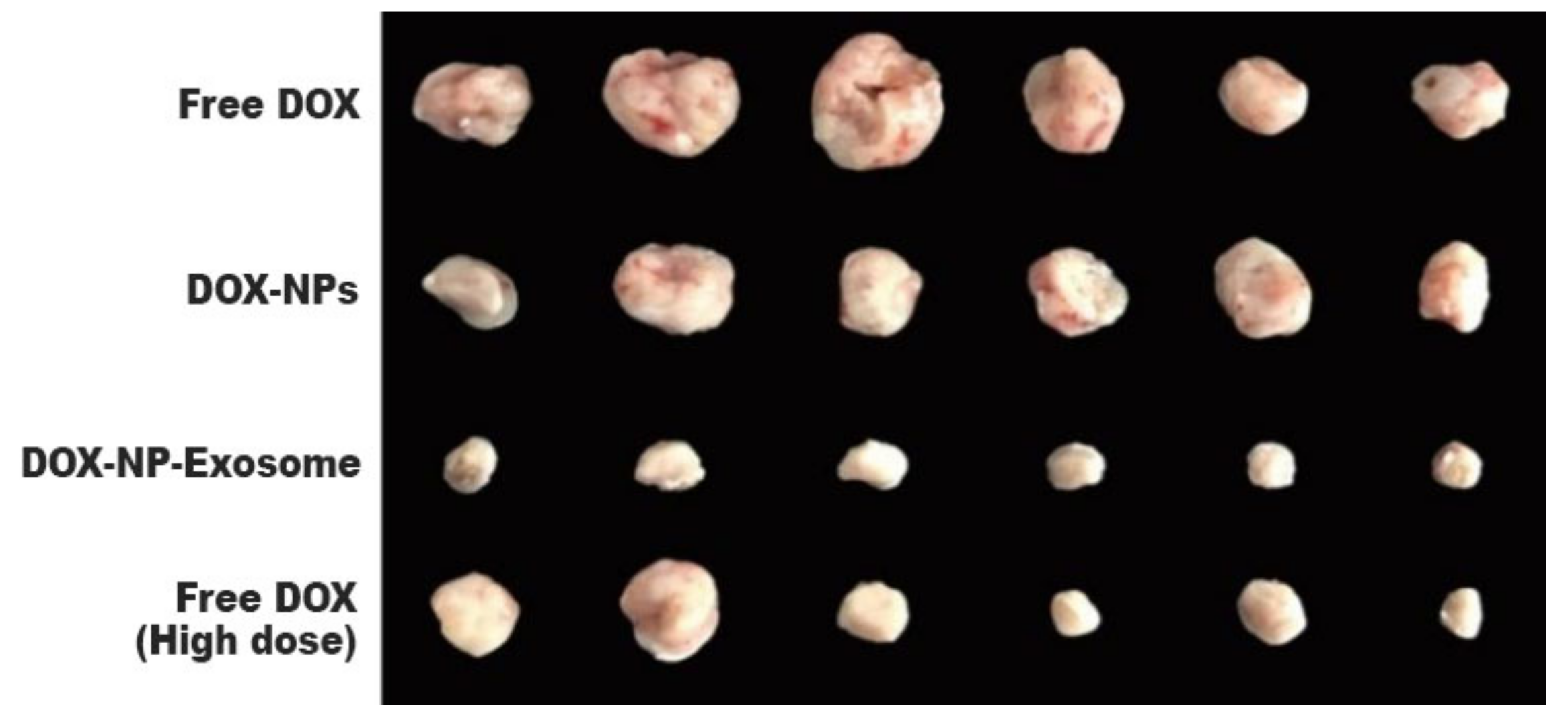{kind=link}
Table 1.
The ESCRT proteins in Saccharomyces cerevisiae yeast and corresponding human analogue [42,44].
| Complexes | Yeast | Human |
|---|---|---|
| ESCRT-0 | Vps27 | Hrs |
| Hse1 | STAM1,2 | |
| ESCRT-I | Vps23 | Tsg101 |
| Vps28 | Vps28 | |
| Vps37 | Vps37 A, B, C, D | |
| Mvb12 | Mvb12 A, B | |
| ESCRT-II | Vps36 | EAP45 |
| Vps22 | EAP30 | |
| Vps25 | EAP20 | |
| ESCRT-III | Vps20 | CHMP6 |
| Snf7 | CHMP4 A, B, C | |
| Vps24 | CHMP3 | |
| Vps2 | CHMP2 A, B | |
| Vps4 Complex | Vps4 | Vps4 A, B |
Table 2.
On-going clinical trials with exosome-based therapies for oncological treatment [91].
Table 2.
On-going clinical trials with exosome-based therapies for oncological treatment [91].
| Year | Disease | Phase | Exosome Source | Formulation | Status |
|---|---|---|---|---|---|
| 2011 | Colon cancer | Phase I | Plant derived | Exosomes loaded with curcumin | Active |
| 2012 | Head and neck cancer | Phase I | Plant derived | Powder to prevent oral mucositis derived from chemoradiation | Active |
| 2013 | Malignant ascites | Phase II | Tumour cell derived | Exosomes loaded with Chemotherapeutic drugs | Unknown |
| 2016 | Malignant pleural effusion | Phase II | Tumour cell derived | Exosomes loaded with cisplatin | Recruiting |
| 2020 | Stage IV pancreatic cancer | Phase I | Mesenchymal Stromal Cells | Exosomes loaded with KRAS G12D siRNA | Not yet recruiting |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rodrigues, P.; Melim, C.; Veiga, F.; Figueiras, A. An Overview of Exosomes in Cancer Therapy: A Small Solution to a Big Problem. Processes 2020, 8, 1561. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8121561
AMA Style
Rodrigues P, Melim C, Veiga F, Figueiras A. An Overview of Exosomes in Cancer Therapy: A Small Solution to a Big Problem. Processes. 2020; 8(12):1561. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8121561
Chicago/Turabian StyleRodrigues, Paulo, Catarina Melim, Francisco Veiga, and Ana Figueiras. 2020. "An Overview of Exosomes in Cancer Therapy: A Small Solution to a Big Problem" Processes 8, no. 12: 1561. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8121561
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.