Potential Impact of Biodegradable Surfactants on Foam-Based Microalgal Cultures
by
 , , and
, , and
María Vázquez
1,* ,
,
José Carlos Castilla-Alcántara
2,
Inés Garbayo
1,
Carlos Vílchez
1 and
and
María Cuaresma
1 1
Algal Biotechnology Group, Ciderta and RENSMA (Center of Research in Natural Resources, Health and Environment), University of Huelva, 21007 Huelva, Spain
2
Instituto de Recursos Naturales y Agrobiología de Sevilla (IRNAS-CSIC), 41012 Seville, Spain
*
Author to whom correspondence should be addressed.
Processes 2020, 8(12), 1640; https://0-doi-org.brum.beds.ac.uk/10.3390/pr8121640
Submission received: 27 October 2020
/
Revised: 8 December 2020
/
Accepted: 10 December 2020
/
Published: 12 December 2020
(This article belongs to the Special Issue Extraction, Exploitation and Application of Algae Biomass)
Abstract
:Microalgae cultivation in liquid foams is a promising concept which requires the use of a surfactant as a foam stabilizing agent. The biodegradable character of a surfactant is a key aspect regarding its applicability in a liquid foam-bed photobioreactor (LF-PBR), since it might influence microalgal growth and the stability of the foam-based cultivation. In this work, the effects of the biodegradable surfactants bovine serum albumin (BSA), Saponin and Tween 20 on the whole microbial community of microalgal cultures (i.e., microalgal and bacterial populations) were studied. The three surfactants enhanced bacterial and microalgal growth in non-axenic microalgal cultures, but they differed in their efficiency to sustain bacterial growth. In this sense, Saponin was proven to enhance the growth of S. obliquus-associated bacteria in microalgae-free cultures, and to sustain it even when other nutrients were lacking, suggesting that Saponin can be used as an energy and nutrients source by these bacteria. The degradation and consumption of Saponin by S. obliquus-associated bacteria was also confirmed by the foaming capacity decrease in Saponin-added bacterial cultures. The biodegradable character of BSA, Saponin and Tween 20 reduces their suitability to be used in a LF-PBR since they would not be able to maintain stable foaming.
1. Introduction
Although microalgae offer a large variety of potential market applications [1,2,3], the number of microalgae-based production processes on an industrial scale is still limited by the high production and processing costs. Currently, large-scale microalgae production is performed in suspension-based cultivation systems, including open systems or raceways and tubular and flat plate photobioreactors [4,5]. The innovative concept of microalgae cultivation in liquid foams was developed as an attempt to reduce production and harvesting costs [6]. The high interfacial surface area of foam and the subsequent CO2/O2 transfer increase, along with the long gas residence time, allow the operation of a liquid foam-bed photobioreactor (LF-PBR) at high biomass concentrations, which contributes to the reduced energy requirements for microalgae harvesting. In this sense, the continuous operation of a LF-PBR at biomass densities higher than 20 g L−1 was already achieved, which is approximately a factor of 10 higher than those employed in conventional suspension-based cultivation systems [7]. In addition, the energy requirements could be lowered due to enhanced mass transfer, longer gas residence time and lower pressure drop in the LF-PBR [8].
The cultivation of microalgae in a LF-PBR requires the addition of a suitable surfactant to stabilize the liquid foams. It was previously determined that for an optimal microalgae cultivation process in a LF-PBR, the surfactant used should, among other characteristics, not be rapidly biodegradable [9]. The biodegradability of the surfactant was determined to be crucial due to two different reasons. First, the use of a biodegradable surfactant in a LF-PBR could result in a decrease in the foaming capacity of the surfactant-added culture. The biodegradation of the surfactant could have an impact on the stability of the surfactant concentration in the reactor, possibly resulting in the necessity of the continuous addition of surfactant in order to maintain an optimal surfactant concentration in the system. The biodegradation of a surfactant can be either partial or complete [10]. In the latter case, it is obvious that the foaming capacity of the surfactant-added culture would decrease as the surfactant is being degraded. As such, the biodegradability of a surfactant was considered to be a drawback for its application in a LF-PBR. However, whether surfactant biodegradation would influence the foaming capacity of a microbial culture has not been experimentally assessed yet. Secondly, microalgal cultures generally contain other associated microorganisms, including bacteria. Obtaining axenic microalgal cultures is an onerous task [11] and the scaling-up of microalgal cultures to large scales under axenic conditions would be extremely difficult and impractical. These bacterial populations that are naturally associated with microalgae are species-specific and their interactions with the microalgal cells can range from beneficial to detrimental to microalgal growth [12]. For instance, Wang and co-workers [13] found that adding bacterial culture medium to the dinoflagellate culture altered the balance between microalgal and bacterial populations in the culture, which in turn resulted in microalgal cell lysis. However, bacteria can also positively influence microalgal growth and even modify their biochemical composition. It is known that bacteria can enhance microalgal growth through the production of growth-promoting factors, such as indole-3-acetic acid, vitamins and siderophores, among others [12]. As an example, Azospirillum brasilense was found to promote the growth of Chlorella vulgaris and Chlorella sorokiniana when co-immobilized and grown in wastewater [14], but also an enhanced accumulation of starch and carbohydrates has been observed for these two Chlorella strains [15]. Considering the varied effects of the bacteria–microalga interactions summarized above, the presence of an additional carbon and energy source for the bacteria in the microalgal culture (i.e., a biodegradable surfactant) can have unpredictable effects over the culture, and further research in this sense is necessary.
Scenedesmus obliquus was shown to be a promising microalgal strain to be cultivated in a LF-PBR [16]. For this reason, S. obliquus-associated bacteria were previously used to assess the biodegradability of 10 surfactants as an attempt to find a suitable surfactant for its application in a LF-PBR [9]. Among the 10 surfactants evaluated, BSA, Saponin and Tween 20 were found to be easily biodegradable [9]. In that study, the biodegradability of the surfactants was investigated at concentrations of 1 and 10 CMC (i.e., critical micelle concentration) as an initial approximation, although the surfactant concentration required for the operation of a LF-PBR may depend on the particular design and size of the system. Therefore, in order to get a greater insight into the effect of a biodegradable surfactant on the microbial populations of a microalgal culture, a wider range of surfactant concentrations is studied in this work. Considering that S. obliquus has been observed to grow in symbiosis with bacteria [17], and that the co-cultivation of this microalga and selected bacterial strains could result in enhanced microalgal growth [18], one of the aims of this work is to further study the effects of these biodegradable surfactants on the growth of both the bacteria and the microalga. To the best of our knowledge, how the presence of biodegradable surfactants influences the dynamics of both a microalga and the coexistent bacterial populations has not been investigated yet.
Furthermore, in a previous study, BSA, Saponin and Tween 20 were shown to be the most easily biodegradable surfactants from a group of 10 compounds, and they were also the best tolerated by the microalgal species tested [9]. Considering that promoting bacterial growth may have a positive effect on microalgal growth, the use of a biodegradable surfactant in foam-based microalgal cultures might in turn enhance microalgal growth. However, very little is known regarding the performance of microalgal cultures with the addition of biodegradable surfactants. In this sense, this manuscript intends to provide insight into how using biodegradable surfactants could impact foam-based microalgae cultivation in terms of the following: (i) how an increase in the bacterial populations would affect microalgal growth and the performance of the microalgal culture as a whole and (ii) to what extent this can affect the foaming capacity of the cultures. In addition, the presented results might also be useful for other potential microbial applications, including the use of biodegradable surfactants.
Considering the background previously described, this work aims at studying the impact of three biodegradable surfactants (BSA, Saponin and Tween 20) on the microbial populations of Scenedesmus obliquus cultures, including both the microalga and the associated bacterial populations. Besides, a greater insight into the biodegradation of Saponin by S. obliquus-associated bacteria is provided by studying the effect of the biodegradation of Saponin on its foaming capacity using microalgae-free bacterial cultures. In addition, the effect of Saponin on the colony-forming characteristic of S. obliquus was also investigated.
2. Materials and Methods
2.1. Biological Material
Scenedesmus obliquus was kindly provided by Fitoplancton Marino S.L. (Cádiz, Spain). The microalga was cultivated in modified M-8 culture medium [19], in which urea was replaced by 3 g L−1 KNO3 in order to avoid the presence of any other source of organic carbon in the media apart from the surfactant. The microalga was maintained in linear phase cultures in shake flasks placed in a growth chamber at 25 °C. The cultures were continuously illuminated at 110 µmol photons m−2 s−1 with white fluorescent lamps and bubbled with air containing 2.5% (v/v) CO2 as unique carbon source.
The bacteria present in S. obliquus cultures were isolated by streaking aliquots of the microalgal culture broth on Trypton Glucose Agar (TGA) plates (Ref. 4021452, Biolife, Italy), and were incubated at 37 °C. Microalgae-free bacterial liquid cultures were prepared in 1 L flasks in Luria–Bertani (LB) medium. The cultures were inoculated with a mix of the morphologically different colonies previously grown in the TGA plates. The flasks were placed in an orbital shaker incubator at 37 °C and 115 rpm (MaxQ 4000, Thermo Scientific, Waltham, MA, USA). The cultures were maintained in these conditions for a maximum of a week before being used to inoculate cultures for the assays.
2.2. Surfactants
Lyophilized BSA powder (Sigma A7030, bovine albumin fraction V), Saponin (Sigma 84510, Premium quality, 8–20% sapogenins) and Tween 20 (Sigma P1379) were tested at specific concentrations expressed in critical micelle concentration (CMC) units. The CMC concentration is a specific value for each surfactant and it refers to the concentration at which the surfactant molecules start forming micelles in the liquid phase. The CMC values and other surfactant characteristics are enlisted in Table 1.
2.3. Cultivation Conditions to Assess the Biodegradability of BSA, Saponin and Tween 20 in Scenedesmus obliquus Cultures
The effects of the surfactants BSA, Saponin and Tween 20 on the bacterial and microalgal populations were assessed in cultures of S. obliquus. The three surfactants were assessed at concentrations of 1, 5, 10, 25 and 50 CMC. Duplicate cultures were prepared in 100 mL flasks with 75 mL of modified M-8 culture medium, in which the original nitrogen source (i.e., urea) was replaced by KNO3. For media preparation, the surfactants were dissolved in autoclaved M-8 culture medium by magnetic stirring in order to obtain stock solutions at 50 CMC. The culture medium and stock solutions were vacuum filtered through cellulose nitrate filters (0.2 µm pore size, 11407-47-ACN, Sartorius, Göttingen, Germany). The different culture media were prepared by mixing the corresponding volumes of the filtered 50 CMC stock solutions and filtered surfactant-free medium. The stock solutions were kept homogenous by being magnetically stirred during the media preparation process.
The flasks were inoculated from a S. obliquus culture maintained in the linear phase of growth. The inoculum volume was adjusted in order to get an initial O.D.750 of 0.2. The inoculum was not previously centrifuged in order to avoid bacteria loss. The cultures were placed on an orbital shaker (MAXI HD, OVAN, Badalona, Spain) set at 155 rpm. They were incubated at room temperature and continuously illuminated at 110 µmol photons m−2 s−1 by fluorescent lamps from above.
2.4. Growth Measurements in Scenedesmus obliquus Cultures
Samples of 1 mL of culture were taken and centrifuged at 3000× g for 5 min (MiniSpin centrifuge, Eppendorf, Hamburg, Germany). Bacterial concentration was measured as the optical density at 600 nm (O.D.600) of the supernatant. The resulting microalgal pellets were resuspended in 1 mL of fresh culture media and the microalgae concentration was measured as the optical density at 750 nm (O.D.750) of these suspensions in order to reduce possible interferences due to bacteria presence. Bacterial and microalgal growth were expressed as the increment in optical density (O.D.) during 2.8 days of cultivation, measured at 600 and 750 nm, respectively (ΔO.D.600 and ΔO.D.750). The parameter O.D.600/O.D.750 was calculated and used as an estimation of the bacteria to microalgae ratio.
In addition, samples of 10 mL of culture were taken at the beginning and at the end of the experiment in order to measure the dry weight of the cultures in duplicate and to calculate the volumetric microalgal biomass productivities achieved by each culture along the experiment. Microalgae samples were filtered through pre-washed, pre-dried and pre-weighed filters (glass microfiber filters with a pore size of 0.7 µm) (Filter Lab MFV5, Filtros Anoia S.L., Barcelona, Spain). Filters were then dried at 80 °C over at least 16 h and cooled down in a desiccator for at least 2 h. The filter weight was determined on a 0.01 mg precision balance (Sartorius CP225D, Sartorius AG, Göttingen, Germany). The dry weight concentration, expressed as g L−1, was calculated by differential weight. Microalgal biomass productivities were calculated as the increase in volumetric biomass dry weight that occurred for a certain period of time and it was expressed in grams of biomass produced per liter of culture per day (g L−1 d−1).
2.5. Gram Staining
For a general bacterial classification, S. obliquus-associated bacteria were Gram-stained [22]. For the Gram staining, the bacterial suspension sample was fixed by adding a few drops of methanol and attached to a glass microscope slide by gentle heating. A few drops of crystal violet stain (Crystal violet oxalate, Scharlau, Sentmenat, Spain) were added and the sample was allowed to stand for one minute. Then, the sample was gently washed with distilled water and a few drops of Lugol solution (Lugol’s solution, Scharlau, Sentmenat, Spain) were added. The sample was gently washed with distilled water after 1 min. A few drops of acetone were added to the sample in order to decolorize the Gram-negative bacteria. After 20 s, the sample was washed with distilled water and a few drops of Safranin solution (Safranin O, Scharlau, Sentmenat, Spain) were added to the sample to dye the Gram-negative bacteria. After washing the sample with distilled water, the dyed sample was observed under the optical microscope using the oil immersion objective lens.
2.6. Inverted Optical Microscope
Surfactant-added microalgal cultures were occasionally observed under an inverted optical microscope (Olympus cooperation IX71S8F-3, SN: 8F15047, Camera: XC30, Software: cellsens standard 1.14) in order to distinguish morphological changes in the microalgal cells and large increases in bacterial populations. Surfactant-free cultures and cultures with Saponin at 100 CMC were incubated in batch in an orbital shaker incubation set at 25 °C, containing a 2.5% CO2-enriched air atmosphere and providing an illumination of 120 µmol photons m−2 s−1. The observations under the inverted microscope were conducted after 5 days of cultivation under the aforementioned conditions.
2.7. Assessment of the Effects of Surfactant Biodegradation on Foamability
The effects of Saponin biodegradation on the foaming capacity (also referred to as foamability) of this surfactant were tested using S. obliquus-associated bacteria. Prior to the experiment, both the surfactant and the bacterial culture medium concentration were optimized. For the determination of the optimal Saponin concentration, the foamability of Saponin solutions was measured for Saponin solutions at 0.5, 1.0, 1.5 and 2.0 CMC prepared in 100 mL of Luria–Bertani (LB) medium. Foamability was measured as the up-flow speed of the foam produced by bubbling the solution with small bubbles at a superficial velocity of 3.68 mm s−1. For this, 100 mL of each solution was placed in a 1 L glass graduated cylinder and the foam up-flow speed was measured after 30 s of foaming. The minimum Saponin concentration at which the maximum foamability was achieved was chosen for the experiment. On the other hand, the optimal concentration of LB medium was determined. For that, cultures of S. obliquus-associated bacteria were prepared in 1 L flasks in LB medium diluted 2-, 5-, 10- and 20-fold. The volume of inoculum was adapted to get an initial optical density at 600 nm (O.D.600) of approximately 0.1. The cultures were prepared in duplicate and incubated in and orbital shaker incubator (MaxQ 4000, Thermo Scientific, Waltham, MA, USA) at 37 °C and 115 rpm. Bacterial growth was measured as O.D.600 and the optimal LB medium concentration was determined as the lowest concentration that resulted in bacterial growth cease during the experimental time (i.e., 72 h).
According to the aforementioned tests, the experiment to evaluate the effects of biodegradability on the surfactant foamability was performed using Saponin at 2 CMC and twenty-time-diluted LB culture medium (LB/20). The conditions assessed included Saponin-free culture medium, Saponin-added culture medium, Saponin-free bacterial cultures and Saponin-added bacterial cultures. The cultures were prepared in duplicate in 1 L flasks and incubated in an orbital shaker incubator (MaxQ 4000, Thermo Scientific, Waltham, MA, USA) at 37 °C and 115 rpm for 11 days. Samples of 105 mL were taken daily. Bacterial growth was measured as O.D.600 and the foamability of the cultures was assessed as described above.
3. Results and Discussion
3.1. Surfactants Biodegradation in Microalgal Cultures
The biodegradation of the surfactants BSA, Saponin and Tween 20 was investigated in microalgal cultures in order to get a greater insight into the microalgae–bacteria interactions. In this study, biodegradation was investigated in S. obliquus cultures because, at the time of the experimentation, this microalga was shown to be the most tolerant to a group of 10 surfactants evaluated [9]; therefore, its potential for microalgae cultivation in foam was considered the highest among the microalgae previously tested in related studies [9,16]. Besides, BSA, Saponin and Tween 20 showed to be the most easily biodegraded by Scenedesmus obliquus-associated bacteria out of 10 evaluated surfactants [9].
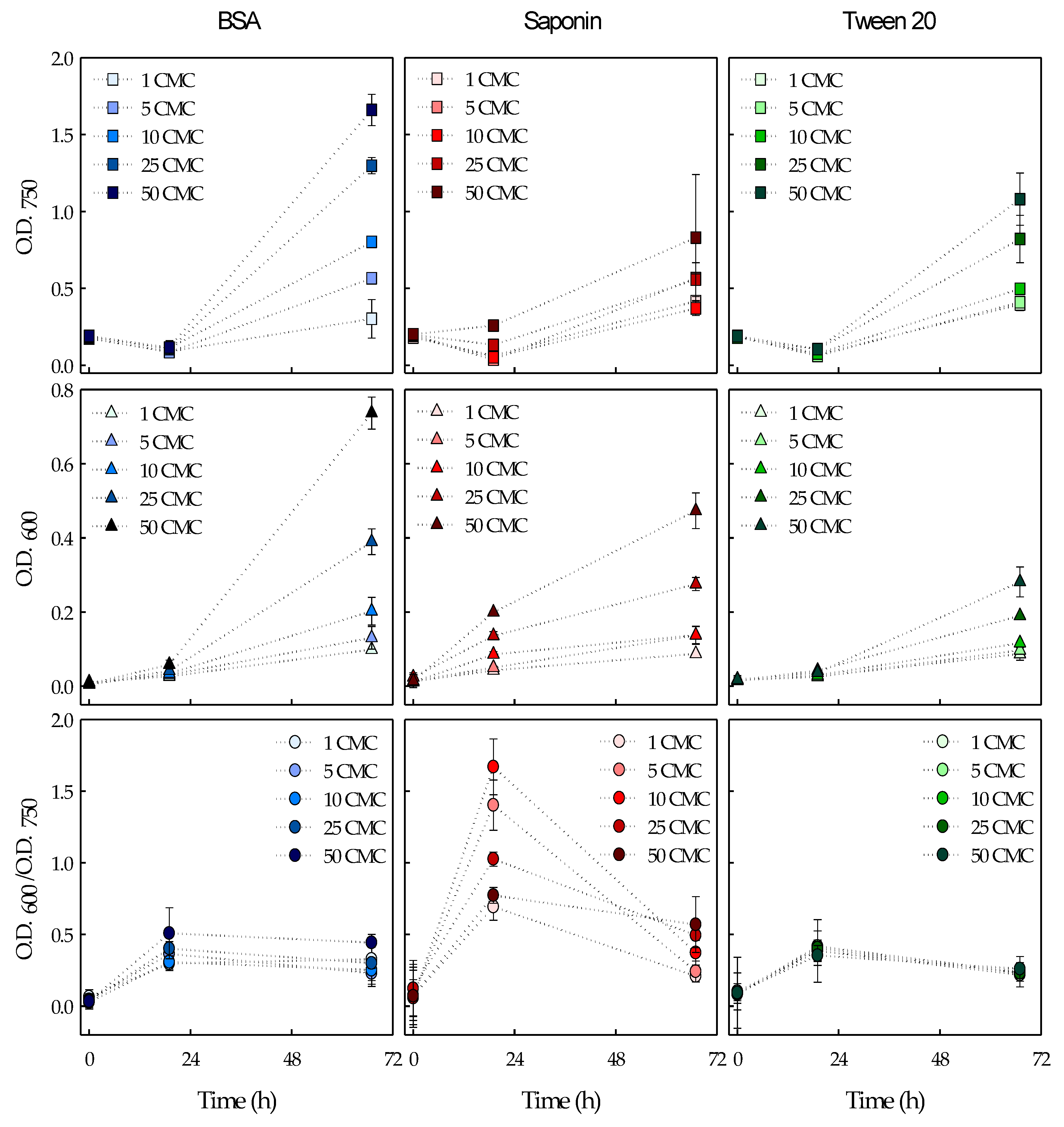
BSA, Saponin and Tween 20 were added to S. obliquus cultures at different concentrations: 1, 5, 10, 25 and 50 CMC. The lack of toxicity of these surfactants to S. obliquus was already proven in the range of concentrations used in this work [9]. Bacterial and microalgal growth in the presence of the biodegradable surfactants was measured in terms of optical density at 600 and 750 nm, respectively, since this is a rapid and commonly-used technique for the estimation of bacterial and microalgal concentrations in liquid cultures [23,24]. The evolution of bacterial and microalgal concentrations and the ratio between both populations in the microalgal cultures incubated with the different surfactants are shown in Figure 1.
As can be seen in Figure 1, the three surfactants promoted both bacterial and microalgal growth in the range of concentrations tested. Bacterial and microalgal growth increased with the increasing surfactant concentrations in all cases, especially at above the 10 CMC concentration. In addition, the photosynthetic efficiency of all the cultures, measured as the maximum photosynthetic efficiency of Photosystem II (Fv/Fm), remained unaffected during the experiment (values remained between 0.67 and 0.75 along the experimental time), which restates the non-toxic nature of these surfactants to S. obliquus.
Besides, both the bacteria and the microalga showed a lag phase of at least 20 h when cultivated at the different concentrations of the three surfactants, with the exception of the bacteria in Saponin cultures, which showed growth from the beginning of the experiment (Figure 1). This resulted in a marked increase in the bacteria-to-microalgae ratio during the first day of cultivation in the presence of Saponin. Microorganisms usually show a latency time to acclimate themselves to the new substrate [25,26], which is the surfactants in this case. Bacteria started growing earlier in Saponin-enriched cultures than in cultures with BSA and Tween 20. Differences in bacterial growth at different surfactant concentrations were already noticeable after 24 h of cultivation in cultures with Saponin, unlike in those with BSA and Tween 20. This possibly indicates that the associated changes in the bacterial populations (i.e., growth of the bacterial populations responsible for the surfactant degradation) or the changes in the bacterial metabolism to degrade the surfactants (e.g., gene expression and/or enzyme induction) might have been more complex for the uptake and utilization of BSA and Tween 20 than for Saponin. Moreover, the bacteria-to-microalgae ratio (i.e., O.D.600/O.D.750) increased in the presence of the three surfactants. The experimental cultivation conditions may have contributed to the general increase in this ratio, which was initially 0.071 ± 0.027. Unlike the experimental cultures, the inoculum was obtained from a culture that was bubbled with CO2-enriched air, which favors microalgal growth over bacterial growth. Nevertheless, the surfactants influenced the bacteria-to-microalgae ratio differently. For BSA-added cultures, slightly higher ratios were found with increased BSA concentrations. In these cultures, the ratio increased in the first 24 h of cultivation and remained constant until the end of the experiment. As aforementioned, the addition of Saponin resulted in steeply increased bacteria-to-microalgae ratios in the first day of cultivation. This increase was especially steep at intermediate Saponin concentrations (i.e., 5-25 CMC). Finally, the addition of Tween 20 resulted in slightly higher bacteria to microalgae ratios, but this increase was similar over the range of concentrations tested.
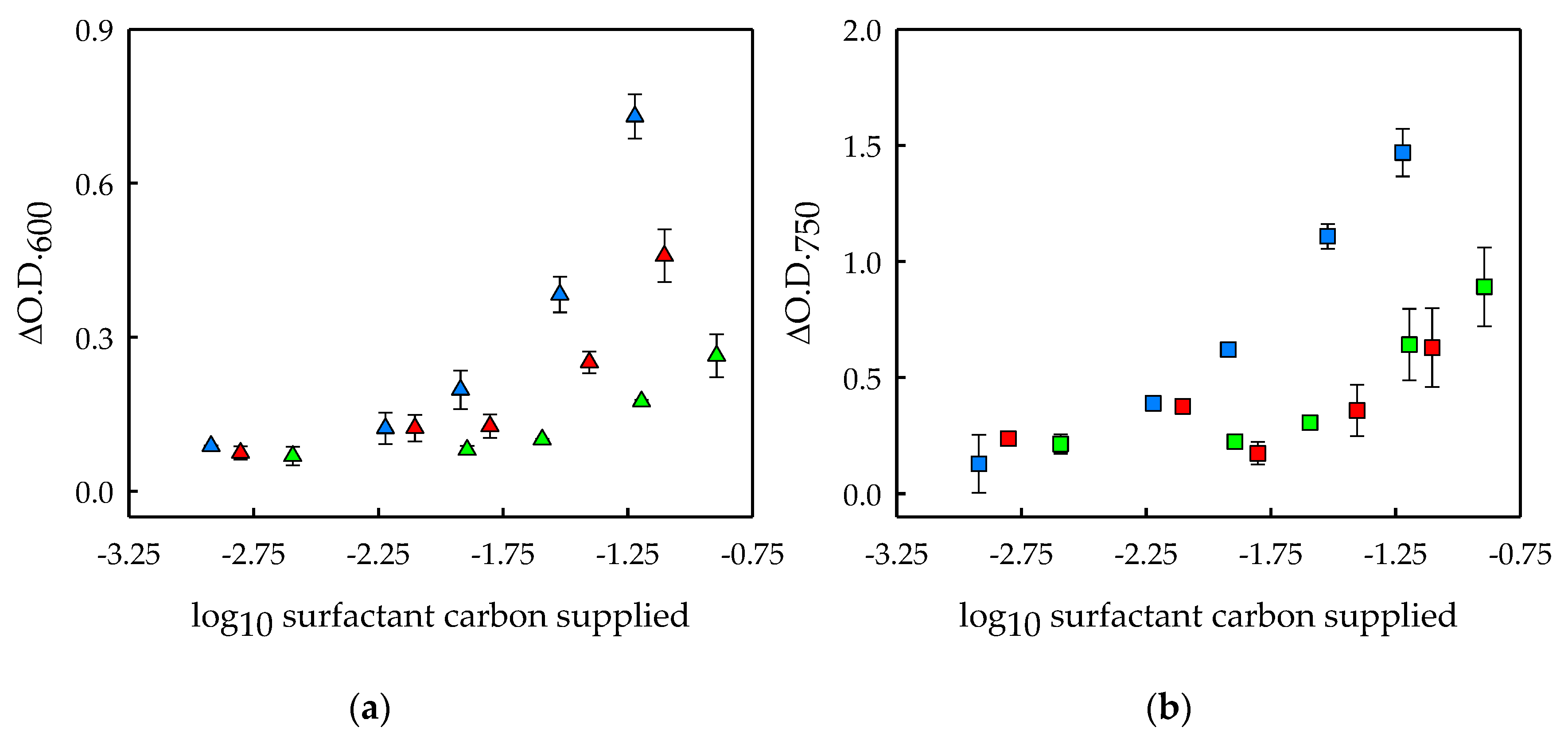
In order to get a better insight into the effect of each biodegradable surfactant on the bacterial and microalgal populations, the increment in optical density at 600 and 750 nm and the ratio between these two values were calculated at the end of the experiment, and are shown in Figure 2. Since the surfactants have varied CMC values and chemical compositions, the carbon supplied by each surfactant differed when added at the same concentration in CMC units (Table 1). Subsequently, the X-axis of Figure 2 is expressed as the decimal logarithm of the milligrams of carbon added to each culture, calculated from the carbon content of the surfactants.
As previously discussed, bacterial growth was stimulated by the three biodegradable surfactants evaluated at concentrations up to 50 CMC (Figure 1). However, same carbon dosages supplied by the different surfactants resulted in different bacterial growth—BSA enhanced bacterial growth to the largest extent while Tween 20 resulted in the lowest bacterial growth (Figure 2a). This can be clearly exemplified by the bacterial growth observed in cultures with the highest concentrations (25 and 50 CMC) (Figure 2a). These differences might be explained by the energy the bacteria obtain from the degradation of these molecules with respect to the energy required for this process. Moreover, the differences in the bacterial growth sustained by the different surfactants might be attributed to the degradation of these compounds to a different extent. BSA molecules have to be hydrolyzed by extracellular enzymes to small peptides that can be transported across the bacterial cell wall [27,28,29,30]. This possibly implies that BSA molecules are degraded to a large extent, which would explain the large increase in bacterial growth. Similarly, Saponin has been reported to be completely biodegraded [26]. This is known as ultimate biodegradation or mineralization [31], and it might explain the intermediate bacterial growth observed in Saponin-added cultures. Lastly, Tween 20 produced the lowest bacterial growth. This may be due to a partial biodegradation of the molecules, which is in agreement with previous observations [32,33].
On the other hand, microalgal growth was measured as the increment of optical density at 750 nm after 2.8 days of cultivation (Figure 2b). As discussed above (Figure 1), a higher surfactant concentration resulted in increased microalgal growth for the three surfactants evaluated. With respect to the carbon content of the surfactants (Figure 2a), the highest microalgal growth stimulation was found for BSA-added cultures, while Tween 20 and Saponin promoted microalgal growth to a similar lower extent. The microalgal growth was approximately 7-, 4- and 3-fold higher at 50 CMC of BSA, Saponin and Tween, respectively, than for their corresponding cultures at 1 CMC. The microalgal growth enhancement at increased surfactant concentrations was noticeable with the naked eye.
The microalgal growth stimulation could be explained by the surfactants’ assimilation by the microalgal cells, or by an enhanced bacterial growth and the subsequent accumulation of released microalgal growth-promoting compounds in the broth. The ability of S. obliquus to solely degrade these surfactants remains unknown. However, it was previously shown that BSA, which stimulated S. obliquus growth to the highest extent, was not consumed by C. sorokiniana cells [6], and therefore that hypothesis was disregarded for that specific case. However, the breakdown of the BSA protein by the bacteria might result in peptides or free aminoacids that could possibly be utilized by the microalga. In this sense, the assimilation of peptones (i.e., partially hydrolyzed proteins) and free aminoacids by microalgae has been reported [34,35,36]. Likewise, the sugar moiety of Saponin, which is composed of galacturonic acid, glucuronic acid and galactose, might have also been mixotrophically metabolized by S. obliquus. In this sense, it is known that some Scenedesmus species can grow on other sugars apart from glucose, including galacturonic acid [37] and galactose [38]. On the other hand, Tween 20 might have been preferentially or solely degraded by bacteria. In this sense, Tween 80 was reported to be more intensively utilized by bacteria than by some microalgae [39]. Nevertheless, some Scenedesmus strains are able to utilize Tween compounds as a carbon source, while other microalgal strains are not [37]. Therefore, it is hard to predict if the S. obliquus strain used in this study had the ability to utilize Tween 20.
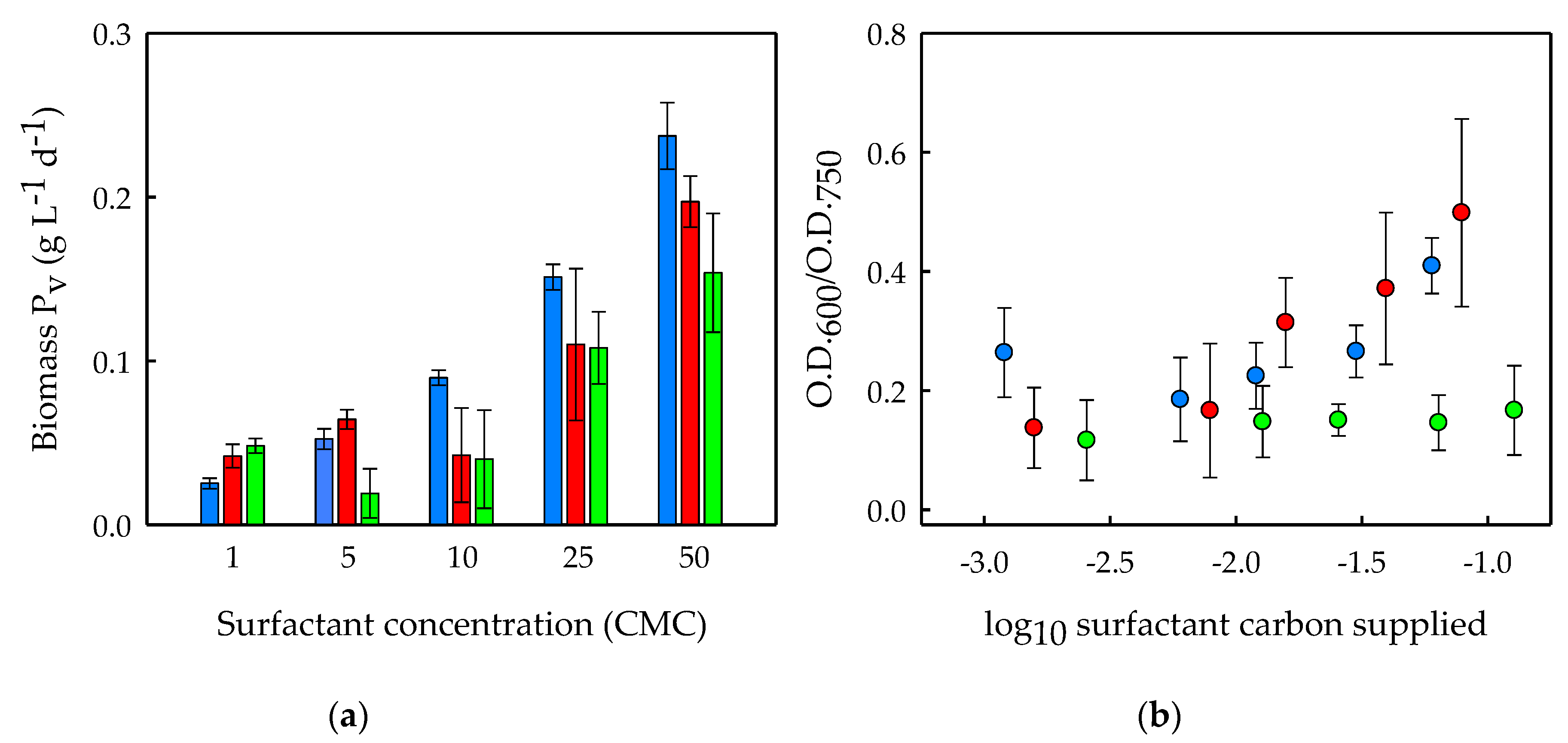
The biomass productivities of S. obliquus achieved during the 2.8 days of cultivation in the presence of the surfactants were also calculated and are shown in Figure 3a. As can be seen, increasing BSA, Saponin and Tween 20 concentrations resulted in increased microalgal biomass productivities. The biomass productivities at 50 CMC were approximately 9.5-, 4.5- and 3-fold higher than those at 1 CMC of BSA, Saponin and Tween 20, respectively. As mentioned above, this might be due to the possible metabolization of the surfactants by the microalga. However, the increased microalgal biomass productivities might have also been a result of the growth-promoting effects of the bacteria on S. obliquus. Enhanced microalgal growth can be expected in the cultures where bacteria grow to a higher extent, since microalgal growth can be enhanced by a variety of compounds produced by bacteria, such as CO2, vitamins, siderophores or growth-promoting factors [12]. In this sense, symbiotic relationships have been already described between S. obliquus and bacteria [17], and a maximum of 2.5-fold biomass growth enhancement has been reported for S. obliquus through its co-cultivation with bacteria [18], which could be in agreement with our findings.
Furthermore, the biodegradable surfactants were shown to affect the bacteria-to-microalgae ratio differently. The bacteria-to-microalgae ratios of the cultures at the end of the experiments are shown in Figure 3b. As can be seen, increasing BSA and Saponin concentrations resulted in similar increases in the bacteria-to-microalgae ratios. However, no effect on this ratio was found in cultures with Tween 20 at any of the concentrations tested, indicating that this surfactant enhanced bacterial and microalgal growth to a similar extent when added at concentrations up to 50 CMC. The fact that the bacteria to microalgae ratio is not affected by the concentration of Tween 20 in the medium might indicate a co-dependent growth of the two microbial communities (algal and bacterial populations) in Tween 20-added cultures, while the addition of the other surfactants (i.e., BSA and Saponin) seems to favor bacterial growth over microalgal growth.
3.2. Morphological Changes in S. obliquus Cultures at High Saponin Concentrations
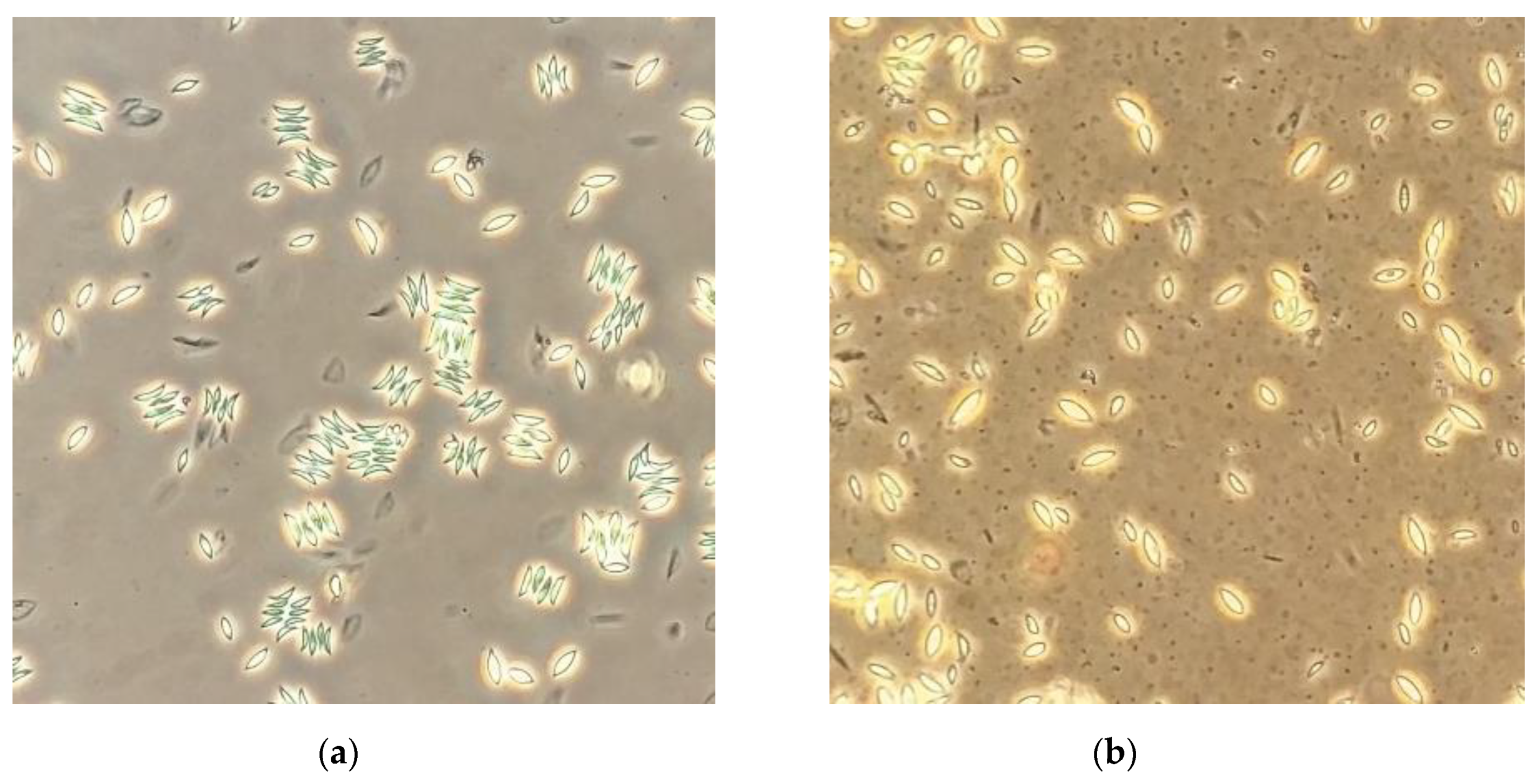
Morphological changes in the colony-forming strain S. obliquus were occasionally noticed when the microalga was cultivated at high concentrations of Saponin. Thus, samples of S. obliquus cultures taken at a high concentration (100 CMC) of the surfactant Saponin were observed under an inverted microscope in order to gather qualitative information on this event (Figure 4). In cultures at high Saponin concentrations (Figure 4b), the microalgal colonies disintegrated and the cells became rounder than those forming the colonies in the surfactant-free cultures (Figure 4a). It has been reported that surfactants can produce morphological changes in microalgal cells [40]. Since Saponin promoted bacterial growth in the microalgal cultures, the changes observed in the microalgal colonies might have also been due to the high bacterial concentration in the microalgal cultures. As an example in this sense, the high competitive pressure exerted by large populations of other microorganisms such as Microcystis aeruginosa has been shown to inhibit the formation of colonies of S. obliquus [41].
3.3. Effects of Surfactant Biodegradation on Foaming Capacity
The bacteria present in Scenedesmus obliquus cultures were previously proved to be responsible for the biodegradation of the surfactants tested in this study [9]. In order to partly characterize these bacteria, they were isolated from an S. obliquus culture by streaking aliquots of the culture broth on Tryptone Glucose Agar (TGA) plates. The Gram-staining of the bacteria isolated from S. obliquus cultures showed that this consortium was composed of more than one strain, including Gram-negative and Gram-positive bacteria. Moreover, the differences in the morphological features of the colonies grown on the agar plates allowed us to identify the presence of at least four bacterial species. As such, more than one bacterial strain might be responsible for the biodegradation of the surfactants tested. In this sense, other studies on the bacterial communities of S. obliquus cultures have found bacteria from at least ten different orders [42], and twenty-five different species were found in S. obliquus cultures by Wang and co-workers [18], among which five were able to grow using the nutrients secreted by the alga. As an attempt to understand the implications of using a biodegradable surfactant for microalgae cultivation in surfactant-stabilized foams, the effects of surfactant degradation by S. obliquus-associated bacteria on the surfactant foaming capacity were further studied. For that, Saponin was used as an example of biodegradable surfactant.
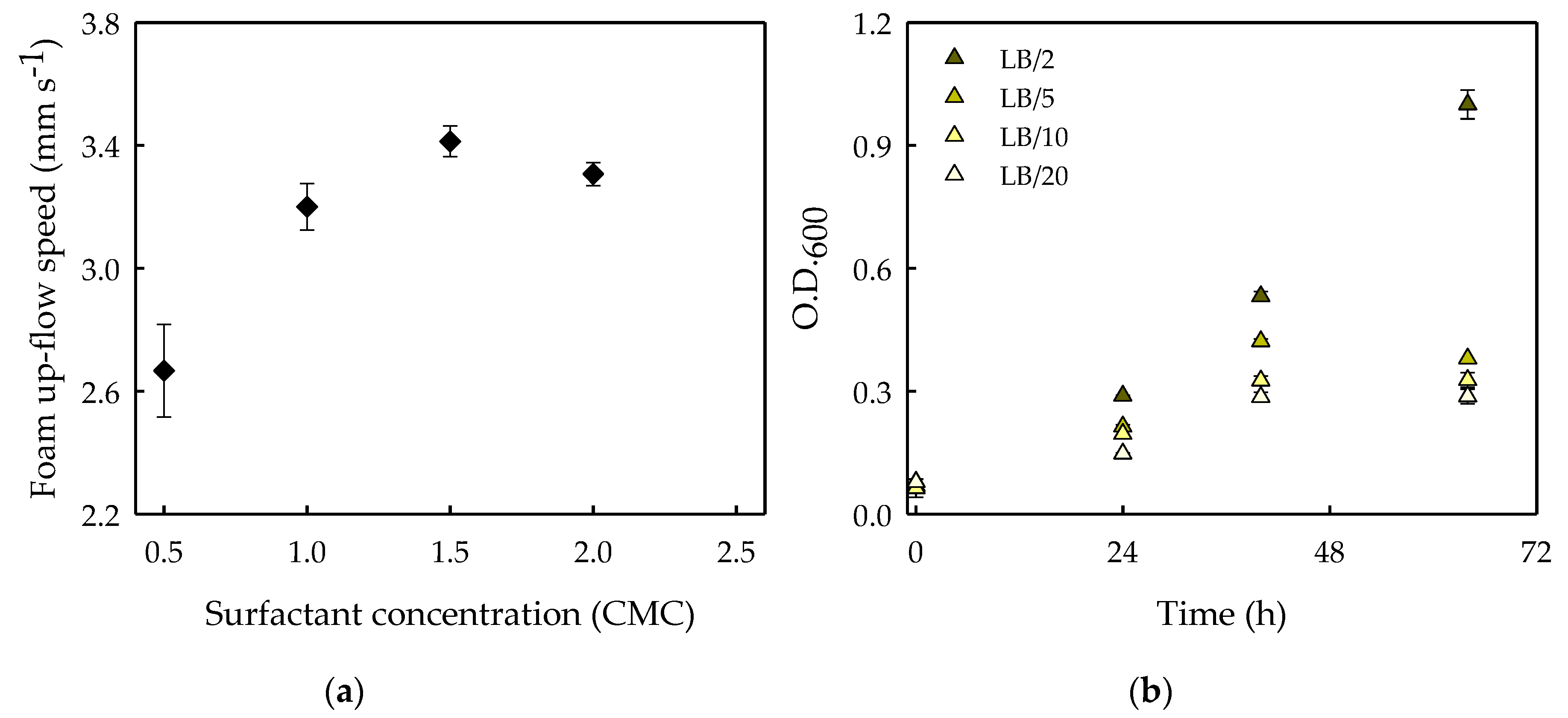
In order to make the biodegradation effects on foamability noticeable, the minimal surfactant concentration that allows us to reach the highest foam production speed in our system was experimentally determined. In this sense, the foamability of Saponin solutions at different concentrations was measured as the foam production speed achieved after 30 s of foaming (Figure 5a). According to the results, Saponin was employed at 2 CMC, since that was the minimum surfactant concentration at which the maximum foam production speed (3.4 ± 0.05 mm s−1) was achieved in Luria–Bertani (LB) medium in our system (Figure 5a). The biodegradability experiment was performed in LB medium because it is a widely used culture medium for the growth of bacteria [43].
The optimal concentration of the bacterial culture medium (LB) was also determined. For that, LB medium diluted 2-, 5-, 10- and 20-fold was inoculated with S. obliquus-associated bacteria, and the analysis of optical density at 600 nm allowed us to follow the bacterial growth in the different solutions (Figure 5b). The optimal medium concentration for the biodegradation experiment should ensure that the contained nutrients in the LB-diluted media would be depleted before the end of the experiment (in less than 72 h of cultivation) in order to distinguish when bacterial growth was supported by the use of the surfactant as a nutrients and energy source. As such, inoculated cultures with the addition of Saponin should be the only ones that might continue growing after the nutrients were depleted from the culture medium.
As can be seen in Figure 5b, only the cultures in the 2-fold diluted medium continued growing after 48 h of cultivation as a result of the still-available nutrients in the medium. Besides, it was decided to use 20-fold diluted LB medium to perform the biodegradation experiment, since this was the lowest concentrated medium which resulted in bacterial growth cease during the experimental time.
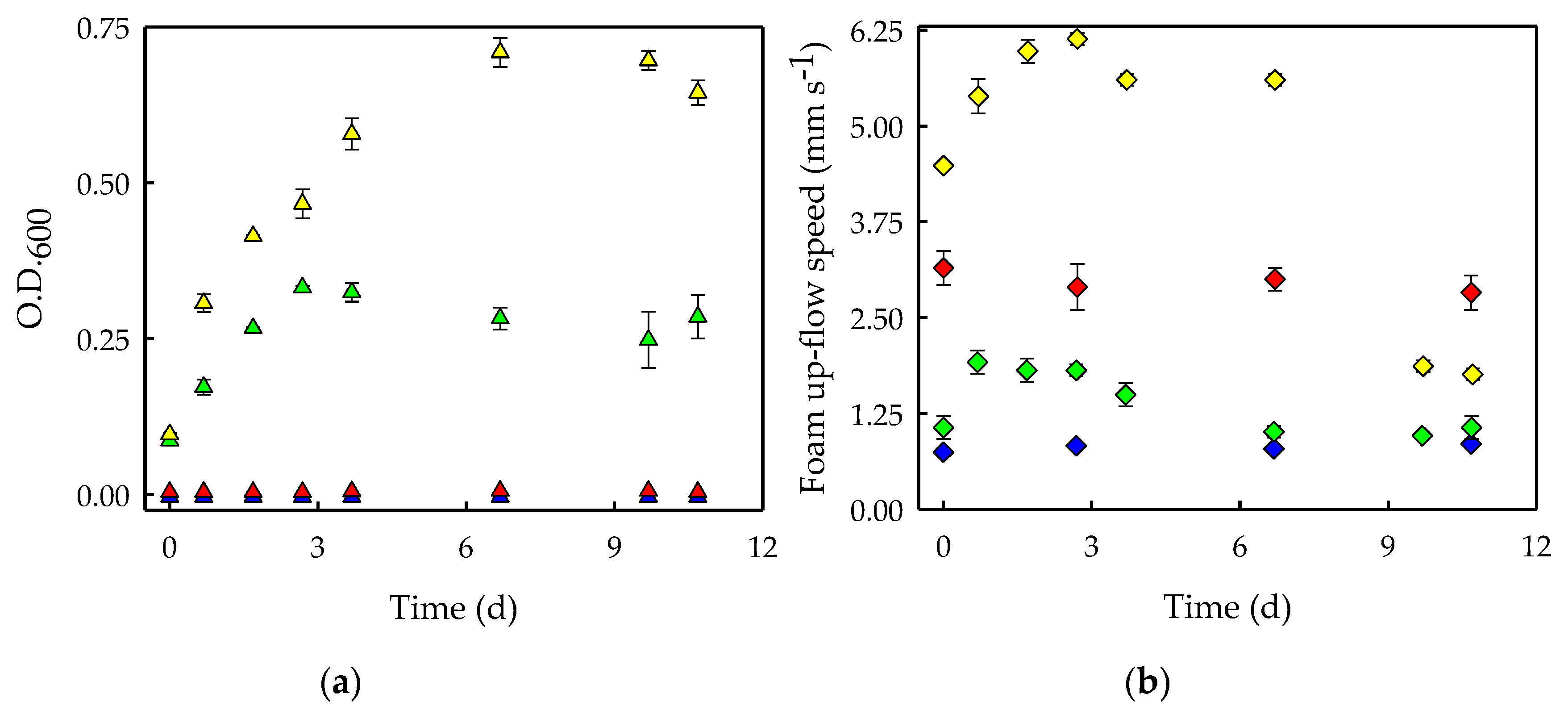
Once the optimal surfactant concentration (i.e., 2 CMC) and culture medium (i.e., LB/20) were identified, the biodegradability experiment was performed. Bacterial growth (Figure 6a) and foamability (Figure 6b) were analyzed in Saponin-free culture media, in a Saponin-added culture media, in Saponin-free bacterial cultures, and in Saponin-added bacterial cultures. Bacterial growth was measured as optical density at 600 nm (Figure 6a). As expected, the optical density remained constant for non-inoculated solutions during the experimental time, indicating that contamination did not occur in Saponin-free (blue triangles) or in Saponin-added (red triangles) culture media. The Saponin-free bacterial cultures (green triangles) grew during the 2–3 first days, after which they stopped growing due to nutrients depletion. However, the bacterial cultures showed a faster growth when Saponin was present (yellow triangles) and bacteria were able to continue growing for 7 days after nutrients from the LB/20 medium were depleted. This indicates that beyond the biodegradation of Saponin, this surfactant supports the growth of S. obliquus-associated bacteria. In this sense, it has been stated that microorganisms can either utilize surfactants as substrates to obtain energy and nutrients, or co-metabolize surfactants by microbial metabolic reactions [25]. Moreover, the fact that the cultures with Saponin grew faster than those without the surfactant suggests that either S. obliquus-associated bacteria consumed Saponin and nutrients from LB/20 medium simultaneously, or that Saponin is a more efficient substrate to promote bacterial growth compared to other nutrients present in the culture media.
On the other hand, the foamability of the cultures was measured in terms of the foam up-flow speed achieved in the first 30 s of foaming (Figure 6b). The foamability of Saponin-free culture media (blue diamonds) remained constant, as did that of the Saponin-added culture media (red diamonds). This indicates that the foamability of Saponin was not affected by the cultivation conditions (i.e., mechanical shaking and incubation at 37 °C). On the other hand, the foamability of Saponin-free bacterial cultures (green diamonds) increased with bacterial growth during the first 24 h of cultivation, indicating that the presence of bacterial cells promoted the foaming. However, the foamability of these cultures remained constant for the rest of the experiment. Finally, the foamability of the Saponin-added bacterial cultures (yellow diamonds) increased during the first 3 days, which can be explained by the large increase in bacterial cells during that period (Figure 6a, yellow triangles). However, it was observed that foamability did not proportionally increase with bacterial growth beyond a certain bacterial concentration (green triangles). This may explain the foaming capacity of Saponin-added bacterial cultures (yellow diamonds), which remained stable between days 3 and 9 despite the bacterial growth observed. Lastly, the further bacterial growth in Saponin-added cultures supported by the degradation and consumption of this surfactant was confirmed by the drop in the foaming capacity of the cultures with this surfactant, which occurred simultaneously with the cease of growth (Figure 6a, yellow triangles). It can be suggested that at that time Saponin would have been completely biodegraded and there would be no remaining nutrients to support bacterial growth further. This finding is in agreement with the previously reported complete biodegradation of this biosurfactant [26].
Biodegradable surfactants, and in particular BSA, Saponin and Tween 20, were previously considered not suitable to be employed in foam-based microalgae cultivation in a LF-PBR. This statement was based on the ability of S. obliquus-associated bacteria to degrade them [9]. However, the present results show that the addition of these surfactants in microalgal cultures promotes not only the growth of the bacteria, but also that of the microalga within the first days of cultivation. It remains still uncertain whether this microalgal growth stimulation is based on the ability of S. obliquus to degrade these compounds and/or is a consequence of the enhanced bacterial growth and potential beneficial interactions with the coexistent bacteria. Nevertheless, the present work indicates that beyond this initial microalgal biomass enhancement, the presence of biodegradable surfactants in non-axenic microalgal cultures would have further implications, such as the loss of the foaming capacity of the cultures and morphological changes in the microalgal cell colonies. This work ultimately indicates that biodegradable surfactants would not be adequate to perform stable microalgae production in a LF-PBR.
4. Conclusions
The biodegradable surfactants BSA, Saponin and Tween 20 enhance the growth of both the microalga and the bacteria naturally present in non-axenic mono-algal cultures of S. obliquus. Microalgal biomass productivity was improved 9.5-, 4.5- and 3-fold in cultures supplemented with BSA, Saponin and Tween 20 at 50 CMC compared to those at 1 CMC, respectively. This can be explained by either the assimilation of these surfactants by S. obliquus and/or by possible beneficial interactions between the microalga and the bacterial consortia in the cultures, such as the production of microalgal growth-promoting factors by the bacteria. It was observed that increasing BSA and Saponin concentrations in the cultures leads to similarly increased bacteria-to-algae ratios, while Tween 20 promotes bacterial and microalgal growth equally at all the surfactant concentrations tested. Our results suggest that using a biodegradable surfactant in a LF-PBR would result in an imbalance in the microbial populations that is specific to the particular biodegradable surfactant employed. Besides this, high concentrations of Saponin, and/or the consequent high bacterial concentration, led to microalgal colony disintegration and morphological changes in the cells. This could influence the performance of foam-based microalgal cultures, for instance, in terms of the extent to which the cells/colonies are dragged up towards the foam phase in a LF-PBR. Moreover, it was proven that Saponin can be used as a nutrient and energy source by microalgae-free S. obliquus-associated bacteria, which in turn results in a decreased foaming capacity of the cultures. Overall, despite the fact that the supplementation of these biodegradable surfactants to microalgal cultures results in microalgal growth enhancement, their application in a LF-PBR for microalgae production in liquid foams would not be suitable due to the consequent foamability losses, as was proven for Saponin in this work.
Author Contributions
Conceptualization and visualization, C.V., I.G., M.C. and M.V.; methodology and validation, J.C.C.-A. and M.V.; investigation and writing—original draft preparation, J.C.C.-A. and M.V.; formal analysis and data curation, M.C. and M.V.; supervision, project administration, writing—review and editing and funding acquisition, C.V., I.G. and M.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was performed in the framework of EU MIRACLES project which received funding from the European Union’s Seventh Framework Programme for research, technological development and demonstration under grant agreement no 613588. The authors also acknowledge the funding received from Rensma, University of Huelva, for publishing open access.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Camacho, F.; Macedo, A.; Malcata, F. Potential industrial applications and commercialization of microalgae in the functional food and feed industries: A short review. Mar. Drugs 2019, 17, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forján, E.; Navarro, F.; Cuaresma, M.; Vaquero, I.; Ruíz-Domínguez, M.C.; Gojkovic, Ž.; Vázquez, M.; Márquez, M.; Mogedas, B.; Bermejo, E.; et al. Microalgae: Fast-growth sustainable green factories. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1705–1755. [Google Scholar] [CrossRef]
- Ruiz, J.; Olivieri, G.; De Vree, J.; Bosma, R.; Willems, P.; Reith, J.H.; Eppink, M.H.M.; Kleinegris, D.M.M.; Wijffels, R.H.; Barbosa, M.J. Towards industrial products from microalgae. Energy Environ. Sci. 2016, 9, 3036–3043. [Google Scholar] [CrossRef] [Green Version]
- Acién Fernández, F.G.; Fernández Sevilla, J.M.; Molina Grima, E. Photobioreactors for the production of microalgae. Rev. Environ. Sci. Bio/Technology 2013, 12, 131–151. [Google Scholar] [CrossRef]
- Norsker, N.H.; Barbosa, M.J.; Vermuë, M.H.; Wijffels, R.H. Microalgal production—A close look at the economics. Biotechnol. Adv. 2011, 29, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Janoska, A.; Lamers, P.P.; Hamhuis, A.; van Eimeren, Y.; Wijffels, R.H.; Janssen, M. A liquid foam-bed photobioreactor for microalgae production. Chem. Eng. J. 2017, 313, 1206–1214. [Google Scholar] [CrossRef] [Green Version]
- Janoska, A.; Barten, R.; de Nooy, S.; van Rijssel, P.; Wijffels, R.H.; Janssen, M. Improved liquid foam-bed photobioreactor design for microalgae cultivation. Algal Res. 2018, 33, 55–70. [Google Scholar] [CrossRef]
- Janoska, A.; Andriopoulos, V.; Wijffels, R.H.; Janssen, M. Potential of a liquid foam-bed photobioreactor for microalgae cultivation. Algal Res. 2018, 36, 193–208. [Google Scholar] [CrossRef]
- Janoska, A.; Vázquez, M.; Janssen, M.; Wijffels, R.H.; Cuaresma, M.; Vílchez, C. Surfactant selection for a liquid foam-bed photobioreactor. Biotechnol. Prog. 2018, 34, 711–720. [Google Scholar] [CrossRef]
- Ying, G.-G. Distribution, Behavior, Fate, and Effects of Surfactants and Their Degradation Products in the Environment. Environ. Int. 2004, 77–109. [Google Scholar] [CrossRef]
- Ramanan, R.; Kim, B.H.; Cho, D.H.; Oh, H.M.; Kim, H.S. Algae-bacteria interactions: Evolution, ecology and emerging applications. Biotechnol. Adv. 2016, 34, 14–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuentes, J.L.; Garbayo, I.; Cuaresma, M.; Montero, Z.; González-Del-Valle, M.; Vílchez, C. Impact of microalgae-bacteria interactions on the production of algal biomass and associated compounds. Mar. Drugs 2016, 14, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, Z.; Su, J.; Tian, Y.; Ning, X.; Hong, H.; Zheng, T. Lysis of a red-tide causing alga, Alexandrium tamarense, caused by bacteria from its phycosphere. Biol. Control 2010, 52, 123–130. [Google Scholar] [CrossRef]
- De-Bashan, L.E.; Hernandez, J.P.; Morey, T.; Bashan, Y. Microalgae growth-promoting bacteria as “helpers” for microalgae: A novel approach for removing ammonium and phosphorus from municipal wastewater. Water Res. 2004, 38, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Choix, F.J.; de-Bashan, L.E.; Bashan, Y. Enhanced accumulation of starch and total carbohydrates in alginate-immobilized Chlorella spp. induced by Azospirillum brasilense: I. Autotrophic conditions. Enzym. Microb. Technol. 2012, 51, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, M.; Fuentes, J.L.; Hincapié, A.; Garbayo, I.; Vílchez, C.; Cuaresma, M. Selection of microalgae with potential for cultivation in surfactant-stabilized foam. Algal Res. 2018, 31, 216–224. [Google Scholar] [CrossRef]
- Ferreira, A.; Ribeiro, B.; Marques, P.A.S.S.; Ferreira, A.F.; Dias, A.P.; Pinheiro, H.M.; Reis, A.; Gouveia, L. Scenedesmus obliquus mediated brewery wastewater remediation and CO2 biofixation for green energy purposes. J. Clean. Prod. 2017, 165, 1316–1327. [Google Scholar] [CrossRef]
- Wang, R.; Xue, S.; Zhang, D.; Zhang, Q.; Wen, S.; Kong, D.; Yan, C.; Cong, W. Construction and characteristics of artificial consortia of Scenedesmus obliquus-bacteria for S. obliquus growth and lipid production. Algal Res. 2015, 12, 436–445. [Google Scholar] [CrossRef]
- Cuaresma, M.; Janssen, M.; Vílchez, C.; Wijffels, R.H. Productivity of Chlorella sorokiniana in a short light-path (SLP) panel photobioreactor under high irradiance. Biotechnol. Bioeng. 2009, 104, 352–359. [Google Scholar] [CrossRef]
- Clarkson, J.R.; Cui, Z.F.; Darton, R.C. Protein denaturation in foam: II. Surface activity and conformational change. J. Colloid Interface Sci. 1999, 215, 333–338. [Google Scholar] [CrossRef]
- Helenius, A.; McCaslin, D.R.; Fries, E.; Tanford, C. Properties of detergents. Methods Enzymol. 1979, 56, 734–749. [Google Scholar] [CrossRef] [PubMed]
- Gram, C. Ueber die isolirte Farbung der Schizomyceten in Schnitt-und Trockenpraparaten. Fortschr. Med. 1884, 2, 185–189. [Google Scholar]
- Griffiths, M.J.; Garcin, C.; van Hille, R.P.; Harrison, S.T.L. Interference by pigment in the estimation of microalgal biomass concentration by optical density. J. Microbiol. Methods 2011, 85, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.A.; Curtis, B.S.; Curtis, W.R. Improving accuracy of cell and chromophore concentration measurements using optical density. BMC Biophys. 2013, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Jurado, E.; Fernndez-Serrano, M.; Ros, F.; Lechug, M. Aerobic Biodegradation of Surfactants. Biodegrad.-Life Sci. 2013. [Google Scholar] [CrossRef] [Green Version]
- Mølgaard, P.; Chihaka, A.; Lemmich, E.; Furu, P.; Windberg, C.; Ingerslev, F.; Halling-Sørensen, B. Biodegradability of the molluscicidal saponins of Phytolacca dodecandra. Regul. Toxicol. Pharmacol. 2000, 32, 248–255. [Google Scholar] [CrossRef]
- Cadoret, A.; Conrad, A.; Block, J.C. Availability of low and high molecular weight substrates to extracellular enzymes in whole and dispersed activated sludges. Enzym. Microb. Technol. 2002, 31, 179–186. [Google Scholar] [CrossRef]
- Confer, D.R.; Logan, B.E. Molecular weight distribution of hydrolysis products during the biodegradation of model macromolecules in suspended and biofilm cultures. II. Dextran and dextrin. Water Res. 1997, 31, 2137–2145. [Google Scholar] [CrossRef]
- Mosquera-Corral, A.; Montràs, A.; Heijnen, J.J.; Van Loosdrecht, M.C.M. Degradation of polymers in a biofilm airlift suspension reactor. Water Res. 2003, 37, 485–492. [Google Scholar] [CrossRef]
- Wang, C.; Lan, C.Q. Effects of shear stress on microalgae—A review. Biotechnol. Adv. 2018, 36, 986–1002. [Google Scholar] [CrossRef]
- Raymond, J.W.; Rogers, T.N.; Shonnard, D.R.; Kline, A.A. A review of structure-based biodegradation estimation methods. J. Hazard. Mater. 2001, 84, 189–215. [Google Scholar] [CrossRef]
- Yeh, D.H.; Pennell, K.D.; Pavlostathis, S.G. Toxicity and biodegradability screening of nonionic surfactants using sediment-derived methanogenic consortia. Water Sci. Technol. 1998, 38, 55–62. [Google Scholar] [CrossRef]
- Chang, Y.C.; Huang, S.C.; Chen, K.F. Evaluation of the effects of nanoscale zero-valent iron (nZVI) dispersants on intrinsic biodegradation of trichloroethylene (TCE). Water Sci. Technol. 2014, 69, 2357–2363. [Google Scholar] [CrossRef] [PubMed]
- Flynn, K.; Butler, I. Nitrogen sources for the growth of marine microalgae: Role of dissolved free amino acids. Mar. Ecol. Prog. Ser. 1986, 34, 281–304. [Google Scholar] [CrossRef]
- Isleten-Hosoglu, M.; Gultepe, I.; Elibol, M. Optimization of carbon and nitrogen sources for biomass and lipid production by Chlorella saccharophila under heterotrophic conditions and development of Nile red fluorescence based method for quantification of its neutral lipid content. Biochem. Eng. J. 2012, 61, 11–19. [Google Scholar] [CrossRef]
- Xiong, W.; Li, X.; Xiang, J.; Wu, Q. High-density fermentation of microalga Chlorella protothecoides in bioreactor for microbio-diesel production. Appl. Microbiol. Biotechnol. 2008, 78, 29–36. [Google Scholar] [CrossRef]
- Tian-Yuan, Z.; Yin-Hu, W.; Lin-Lan, Z.; Xiao-Xiong, W.; Hong-Ying, H. Screening heterotrophic microalgal strains by using the Biolog method for biofuel production from organic wastewater. Algal Res. 2014, 6, 175–179. [Google Scholar] [CrossRef]
- Pancha, I.; Chokshi, K.; Mishra, S. Enhanced biofuel production potential with nutritional stress amelioration through optimization of carbon source and light intensity in Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2015, 179, 565–572. [Google Scholar] [CrossRef]
- Tuchman, N.C.; Schollett, M.A.; Rier, S.T.; Geddes, P. Differential heterotrophic utilization of organic compounds by diatoms and bacteria under light and dark conditions. Hydrobiologia 2006, 561, 167–177. [Google Scholar] [CrossRef]
- Azizullah, A.; Richter, P.; Häder, D.P. Toxicity assessment of a common laundry detergent using the freshwater flagellate Euglena gracilis. Chemosphere 2011, 84, 1392–1400. [Google Scholar] [CrossRef]
- Zhu, S.; Wang, Y.; Huang, W.; Xu, J.; Wang, Z.; Xu, J.; Yuan, Z. Enhanced Accumulation of Carbohydrate and Starch in Chlorella zofingiensis Induced by Nitrogen Starvation. Appl. Biochem. Biotechnol. 2014, 174, 2435–2445. [Google Scholar] [CrossRef] [PubMed]
- Greses, S.; Christian, J.; Aguado, D.; Ferrer, J.; Seco, A. Microbial community characterization during anaerobic digestion of Scenedesmus spp. under mesophilic and thermophilic conditions. Algal Res. 2017, 27, 121–130. [Google Scholar] [CrossRef]
- Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Scenedesmus obliquus (upper figures) and bacterial (middle figures) growth expressed as optical density at 750 nm (O.D.750) and 600 nm (O.D.600), respectively. Bacteria to microalgae ratio (lower figures) expressed as the ratio between the optical density at 600 and 750 nm (O.D.600/O.D.750). Cultures with BSA (left), Saponin (middle) and Tween 20 (right) at concentrations of 1, 5, 10, 25 and 50 CMC. The average values of duplicate cultures are shown, together with their corresponding standard deviations.
Figure 1.
Scenedesmus obliquus (upper figures) and bacterial (middle figures) growth expressed as optical density at 750 nm (O.D.750) and 600 nm (O.D.600), respectively. Bacteria to microalgae ratio (lower figures) expressed as the ratio between the optical density at 600 and 750 nm (O.D.600/O.D.750). Cultures with BSA (left), Saponin (middle) and Tween 20 (right) at concentrations of 1, 5, 10, 25 and 50 CMC. The average values of duplicate cultures are shown, together with their corresponding standard deviations.

Figure 2.
Microbial growth in cultures with the addition of BSA (blue symbols), Saponin (red symbols) or Tween 20 (green symbols). (a) Bacterial growth expressed as the increment in optical density at 600 nm after 2.8 days of cultivation (ΔO.D.600) in the supernatant of Scenedesmus obliquus cultures enriched with BSA, Saponin and Tween 20 at concentrations of 1, 5, 10, 25 and 50 CMC; (b) Scenedesmus obliquus growth expressed as the increment in optical density at 750 nm after 2.8 days of cultivation (ΔO.D.750) in cultures enriched with BSA, Saponin and Tween 20 at concentrations of 1, 5, 10, 25 and 50 CMC. Surfactant concentration is expressed as the carbon dosage resulting from surfactant addition (logarithmic values, in milligrams). The average values of duplicate cultures are shown, together with their corresponding standard deviations.
Figure 2.
Microbial growth in cultures with the addition of BSA (blue symbols), Saponin (red symbols) or Tween 20 (green symbols). (a) Bacterial growth expressed as the increment in optical density at 600 nm after 2.8 days of cultivation (ΔO.D.600) in the supernatant of Scenedesmus obliquus cultures enriched with BSA, Saponin and Tween 20 at concentrations of 1, 5, 10, 25 and 50 CMC; (b) Scenedesmus obliquus growth expressed as the increment in optical density at 750 nm after 2.8 days of cultivation (ΔO.D.750) in cultures enriched with BSA, Saponin and Tween 20 at concentrations of 1, 5, 10, 25 and 50 CMC. Surfactant concentration is expressed as the carbon dosage resulting from surfactant addition (logarithmic values, in milligrams). The average values of duplicate cultures are shown, together with their corresponding standard deviations.

Figure 3.
(a) Volumetric biomass productivity of Scenedesmus obliquus (Biomass Pv), expressed as grams of biomass produced per liter of culture broth and per day, and (b) bacteria-to-microalgae ratio, expressed as the ratio between the optical density at 600 nm and that at 750 nm after 2.8 days of cultivation (O.D.600/O.D.750) in cultures enriched with BSA (blue bars and symbols), Saponin (red bars and symbols) and Tween 20 (green bars and symbols) at concentrations of 1, 5, 10, 25 and 50 CMC. The average values of duplicate cultures are shown, together with their corresponding standard deviations. Surfactant concentration is expressed in (a) CMC units and (b) as the carbon dosage resulting from surfactant addition (logarithmic values, in milligrams).
Figure 3.
(a) Volumetric biomass productivity of Scenedesmus obliquus (Biomass Pv), expressed as grams of biomass produced per liter of culture broth and per day, and (b) bacteria-to-microalgae ratio, expressed as the ratio between the optical density at 600 nm and that at 750 nm after 2.8 days of cultivation (O.D.600/O.D.750) in cultures enriched with BSA (blue bars and symbols), Saponin (red bars and symbols) and Tween 20 (green bars and symbols) at concentrations of 1, 5, 10, 25 and 50 CMC. The average values of duplicate cultures are shown, together with their corresponding standard deviations. Surfactant concentration is expressed in (a) CMC units and (b) as the carbon dosage resulting from surfactant addition (logarithmic values, in milligrams).

Figure 4.
Scenedesmus obliquus observed under an inverted optical microscope after five days of cultivation: (a) with no surfactant added and (b) with Saponin at 100 CMC.
Figure 4.
Scenedesmus obliquus observed under an inverted optical microscope after five days of cultivation: (a) with no surfactant added and (b) with Saponin at 100 CMC.

Figure 5.
(a) Foamability of the surfactant Saponin in 100 mL of LB culture medium expressed in terms of the foam up-flow speed achieved after 30 s of foaming (mm s−1) as a function of the surfactant concentration (CMC). The average values of three experimental replicates are shown, together with their corresponding standard deviations. (b) Bacterial growth expressed as optical density at 600 nm (O.D.600) in LB culture medium 2-, 5-, 10- and 20-fold diluted. The average values of duplicate cultures are shown, together with their corresponding standard deviations.
Figure 5.
(a) Foamability of the surfactant Saponin in 100 mL of LB culture medium expressed in terms of the foam up-flow speed achieved after 30 s of foaming (mm s−1) as a function of the surfactant concentration (CMC). The average values of three experimental replicates are shown, together with their corresponding standard deviations. (b) Bacterial growth expressed as optical density at 600 nm (O.D.600) in LB culture medium 2-, 5-, 10- and 20-fold diluted. The average values of duplicate cultures are shown, together with their corresponding standard deviations.

Figure 6.
(a) Bacterial growth expressed as optical density at 600 nm (O.D.600) during the biodegradability experiments with Saponin. (b) Foamability expressed as foam up-flow speed (mm s−1). Culture conditions: Saponin-free culture media (blue symbols), Saponin-added culture media (red symbols), Saponin-free bacterial cultures (green symbols) and Saponin-added bacterial cultures (yellow symbols). Saponin was used at a concentration of 2 CMC. The average values of duplicate cultures are shown, together with their corresponding standard deviations.
Figure 6.
(a) Bacterial growth expressed as optical density at 600 nm (O.D.600) during the biodegradability experiments with Saponin. (b) Foamability expressed as foam up-flow speed (mm s−1). Culture conditions: Saponin-free culture media (blue symbols), Saponin-added culture media (red symbols), Saponin-free bacterial cultures (green symbols) and Saponin-added bacterial cultures (yellow symbols). Saponin was used at a concentration of 2 CMC. The average values of duplicate cultures are shown, together with their corresponding standard deviations.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characteristics of the surfactants employed. P.I. stands for the online product information sheet.
Table 1.
Characteristics of the surfactants employed. P.I. stands for the online product information sheet.
| Surfactant | Type | CMC (g L−1) | Molecular Weight (g mol−1) | Carbon Content (% w/w) | Chemical Composition |
|---|---|---|---|---|---|
| BSA | Non-catalytic protein, anionic character | 0.03 [20] | 2754.11 | 53.64 | Bovine serum albumin derived from cows |
| Saponin | Non-ionic | 0.05 (P.I.) | 486.69 | 42.73 | Glycoside + Triterpene |
| Tween 20 | Non-ionic | 0.06 [21] | 1228.00 | 56.73 | Polyoxyethylene (20) sorbitanmonolaurate |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vázquez, M.; Castilla-Alcántara, J.C.; Garbayo, I.; Vílchez, C.; Cuaresma, M. Potential Impact of Biodegradable Surfactants on Foam-Based Microalgal Cultures. Processes 2020, 8, 1640. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8121640
AMA Style
Vázquez M, Castilla-Alcántara JC, Garbayo I, Vílchez C, Cuaresma M. Potential Impact of Biodegradable Surfactants on Foam-Based Microalgal Cultures. Processes. 2020; 8(12):1640. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8121640
Chicago/Turabian StyleVázquez, María, José Carlos Castilla-Alcántara, Inés Garbayo, Carlos Vílchez, and María Cuaresma. 2020. "Potential Impact of Biodegradable Surfactants on Foam-Based Microalgal Cultures" Processes 8, no. 12: 1640. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8121640
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.