Cell Factories for Industrial Production Processes: Current Issues and Emerging Solutions



Department of Biotechnology and Biomedicine, Technical University of Denmark, Søltofts Plads 2800 Kgs. Lyngby, Denmark
*
Author to whom correspondence should be addressed.
Processes 2020, 8(7), 768; https://0-doi-org.brum.beds.ac.uk/10.3390/pr8070768
Submission received: 30 March 2020
/
Revised: 19 June 2020
/
Accepted: 22 June 2020
/
Published: 30 June 2020
(This article belongs to the Special Issue Bioreactor System: Design, Modeling and Continuous Production Process)
Abstract
:Despite all the progresses made by metabolic engineering, still only a few biotechnological processes are running at an industrial level. In order to boost the biotechnological sector, integration strategies as well as long-term views are needed. The aim of the present review is to identify the main drawbacks in biotechnological processes, and to propose possible solutions to overcome the issues in question. Novel cell factories and bioreactor design are discussed as possible solutions. In particular, the following microorganisms: Yarrowia lipolytica, Trichosporon oleaginosus, Ustilago cynodontis, Debaryomyces hansenii along with sequential bioreactor configurations are presented as possible cell factories and bioreactor design solutions, respectively.
1. Introduction
Biorefineries are defined as “… sustainable processing of biomass into a spectrum of marketable products (food, feed, materials, chemicals) and energy (fuels, power, heat)” [1]. They typically involve the key stages of feedstock pretreatment, fermentation product recovery and purification. Biorefineries can be broadly classified into one of the following groups according to the nature of the feedstock used to produce the desired bio-products: (1) sugar biorefineries, (2) vegetable oil biorefineries, (3) synthesis gas biorefineries and (4) lignocellulose biorefineries [2]. Sugar biorefineries, in which crude sugar provides the carbon source for the fermenting microorganism, are ubiquitous in industrial biotechnology (Table 1). Crude sugar can be readily metabolized, eliminating the need for complex and costly pretreatment steps. However, such sugars are typically derived from edible food crops which requires large areas of agricultural land and threatens food security. In vegetable oil biorefineries, bio-commodities are produced from oils. The products may be converted via transesterification into fatty acid esters, which can be used in diesel engines [3]. Unlike crude sugar and vegetable oil biorefineries, lignocellulosic biorefineries make use of non-edible waste biomass as feedstocks, protecting food security. However, lignocellulose must undergo a chemical or biological pretreatment step to release fermentable sugars before they can be metabolized by the microorganism and converted into an end-product of interest [2]. Synthesis gas, derived from the gasification of hydrocarbon-rich substances, is employed to produce bio-commodities in synthesis gas biorefineries. Although traditionally derived from coal, synthesis gas can be produced from the gasification of more renewable substances such as organic waste.
Recent advances in the field of metabolic engineering have enabled the construction of microbial cell factories for the production of wide-ranging valuable compounds [4]. The industrial biotechnology sector is rapidly growing, and a number of bio-commodities are in production today. A few relevant examples are reported in Table 1.
A number of companies have successfully employed microorganisms to convert plant-derived renewable feedstocks into market-relevant products at an industrial level as shown in Table 1. As the product of interest may be either native or heterologous, the potential applications of microbial cell factories are far reaching. Despite this, scale-up remains a major challenge in effective bioprocess development [1] and further research is critical to expediting the development of sustainable biorefinery platforms to capitalize on the benefits observed at the laboratory scale. Additional issues across the whole production chain must be overcome to ensure successful scale up and long term survival of more biorefineries. Key factors to consider include bioreactor configuration, feedstock and microbial host. Continuous fermentations offer inherent advantages such as relatively low operational costs, increased efficiency and reduced down time compared to the more widely used batch or fed-batch modes of operation [4]. Despite this, the successful implementation of continuous bioprocesses faces a number of challenges, which will be discussed in detail in this review. Microbial host and feedstock selection were also considered; such factors can have a major influence on the economic and technological feasibility of the bioprocess. The model host Saccharomyces cerevisiae has been widely employed for microbial cell factory development due to its well characterized genome, eukaryotic biosynthetic machinery and tolerance for industrial fermentation conditions. However, this organism is unable to metabolize second generation feedstocks such as lignocellulose and glycerol, and cultivation requires substantial volumes of freshwater. Alternative non-conventional yeasts, capable of overcoming some of these bottlenecks, are gaining increasing interest as potential microbial cell factories. Microorganisms present a natural biodiversity that may be exploited by finding strains that synthesize products of commercial value. In other words, microbes produce metabolites or enzymes that allow them to survive in a certain natural environment. Furthermore, those natural capabilities can be used in large-scale fermentation processes, for production of fine chemicals, antibiotics or drugs, among others. Thus, the aforementioned term “cell factory” describes those microorganisms that are used in the bioconversion of substrates into a product of industrial interest. A cell factory may be genetically manipulated to obtain some advantages during industrial bioprocesses. Moreover, physical parameters such pH or temperature, can be also altered during cultivation in order to obtain a more suitable environment for production. Desirable cultivation characteristics that an ideal cell factory must provide are i.e., lower production costs (including costs of downstream processing), wide range of substrate utilization, simplified product recovery and process operation, and increased profit margin of the process [14]. Promising examples of such fungi will be evaluated in this review. Finally, downstream processing methods to maximize product recovery and cell viability will be addressed.
2. Industrial Bioprocesses: Current Issues
In this section, the main obstacles susceptible to arise during the industrial application of a bioprocess are discussed, and subsequent solutions are suggested for each particular problem.
2.1. Feedstock
Several factors must be considered during feedstock selection, such as: (1) sugar and lignin composition, (2) seasonable dependency, (3) abundance and (4) transport costs. Lignocellulosic biomass is comprised of differing proportions of the carbohydrate polymers cellulose and hemicellulose and the aromatic polymer, lignin. Feedstocks with a low lignin content are desirable as relatively mild pretreatment conditions (low temperature and pressure) are required, minimizing both capital and operational costs. The abundance and seasonal dependency of the feedstock are also important parameters to take into account. For example, a feedstock may be highly abundant following an annual harvest, however, the biorefinery could suffer a feedstock shortage during the remainder of the year. Continuous bioprocesses are inherently more vulnerable to such supply shortages compared to more traditional batch-based processes, largely due to a lack of down-time. This can have a major impact on the economic viability of continuous biorefineries [4]. In addition, if a feedstock is highly abundant but scattered over a large geographical area, the costs and environmental impacts associated with its transportation will be increased dramatically.
A wide range of potential waste-derived feedstocks are available for the biorefinery industry as summarized in Table 2. Industrial waste products such as brewer’s spent grain, sugar beet pulp and sugarcane bagasse are one promising source. As the major by-product of the brewing industry, accounting for around 25% of the total product [15,16], brewer’s spent grain is both low in cost and unaffected by seasonal variations. Similarly, sugar beet pulp is the major by-product of sugar beet processing, with 1.5 Mt generated per 30 Mt of sugar beet [17,18]. The very low lignin content of just 2% (Table 2), renders sugar beet pulp a particularly desirable feedstock. The refining of sugarcane also yields vast quantities of agro-waste material continuously, with 270 kg of sugarcane bagasse produced per ton of sugarcane [19,20]. Agro-food waste products resulting from the industrial food processing sector are another source of organic wastes with potential as feedstocks. The processing of potatoes, for example, generates around 0.6 Mt y−1 of organic waste in Europe alone [16,17,18,19,20,21]. The growing fresh cut sector discards around 50% of the raw product produced during the peeling and cutting of fruit and vegetables to be sold in plastic bags [21,22]. Following the roasting of coffee beans, the silverskin, is discarded as the main by-product [16,17,18,19,20,21]. These industrial wastes are available all year round, however, relatively low quantities are produced annually, limiting their widespread application. The agricultural residues resulting from crop farming (such as: olive pomace/pits, corn stover, corncob) [17,18,19] are a more abundant source of lignocellulose with around 150 Mt y−1 (Table 2) produced. However, the high seasonal dependency of this organic waste could result in supply issues if selected as the sole feedstock. Finally, the organic fraction of municipal solid waste (OFMSW) comprises up to 50% of the two billion tons of municipal solid waste generated globally per year [23]. As such waste is generated worldwide in vast quantities, it is abundant and readily available with minimal transportation requirements. Despite this, the applications of this type of feedstock are limited, due to substantial variations in composition, which can hinder technological and economic feasibility [24].
Alternative non-lignocellulosic compounds with potential as low cost biorefinery feedstocks include synthesis gas and glycerol. As synthesis gas can be generated from the gasification of OFMSW, it could provide a widely available and low-cost feedstock. However, the range of microorganisms able to metabolize synthesis gas is limited [10] and the high temperatures and pressures required can result in high capital costs. Glycerol is a major by-product of biodiesel production, with around 100 kg of crude glycerol generated per ton of biodiesel [25]. In 2017, Rodrigues and co-workers estimated global biodiesel production will exceed 110,000 mL in the coming years [26]. Although glycerol does not contain troublesome lignin, is available all year round and is produced in specific geographical areas, only certain microorganisms are able to use it as a sole carbon source. Many potential waste derived feedstocks are available, however, there is unlikely to be a single sustainable solution and the appropriate choice is likely to be heavily influenced by the nature and geographical location of the individual bioprocess.
2.2. Inhibitory Compounds from the Feedstock
Lignocellulosic materials must be pretreated to breakdown the external lignin layer and complex polysaccharides in order to release monomeric fermentable sugars. Several methods for lignocellulosic biomass pretreatment have been reported in the literature [27,28]. In those involving harsher conditions such as high temperature and pressure, a number of unwanted side-products are produced [29]. These include phenolic compounds such as vanillin, ferulic acid and p-coumaric acid; furan derivatives, such as furfural and hydroxymethylfurfural (HMF), and small organic acids including levulinic acid, formic acid and acetic acid. Such compounds can penetrate the cell membrane and inhibit cellular metabolism. The extent of the effect of each compound on cell viability and productivity is dependent on its concentration and the microorganism in question [29]. Similarly, non-lignocellulosic feedstocks can also contain inhibitory compounds. Crude glycerol, for example, is known to contain salts, alcohols and fatty acids, which can interfere with the fermentation process and should therefore be removed [26]. During non-continuous processes, the removal of such inhibitory compounds is relatively straightforward with an additional step in the production chain. In continuous processes, however, integrating a removal step prior to feeding the continuous reactor is more complicated and can increase down-time, thus increasing operational costs [30]. Several innovative pretreatment methods have been reported, which yield low inhibitor concentrations [16] or involve detoxification steps to effectively remove inhibitors [31]; despite this, as such approaches make use of specific reagents and require further steps, respectively, associated costs remain high.
2.3. Changes in the pH Value
Major discrepancies in pH occur across the various stages of the biorefinery process (biomass pretreatment, enzymatic hydrolysis and fermentation). Steam explosion and acid hydrolysis, two of the most widely used methods for biomass pretreatment, are carried out at pH 2–3 [31,32]. In alkaline pretreatment methods, on the other hand, the pH ranges between 8 and 10 [33]. As most enzymes are denatured at such extreme values, the pH must be adjusted prior to the enzyme hydrolysis step. Although the optimal pH for this step is strongly influenced by the enzyme(s) used, it typically ranges between 4.8 and 6 [34]. In the subsequent fermentation stage, the pH must be adjusted again to ensure optimal performance of the microorganism. For example, S. cerevisiae grows best between pH 4 and 5 [35], Clostridium acetobutylicum at around pH 6 [22] and Lactobacillus spp. at pH 7 [36]. As the adjustment of pH requires excessive water and/or chemical consumption it is both expensive and time-consuming, hindering overall process productivity. This is particularly problematic in continuous processes where unexpected pH deviations can have a domino effect on all subsequent steps if undetected. This can ultimately lead to loss of the entire production, which often runs for over 20 days, resulting in substantial losses in both time and profit [4].
2.4. Water Demand
Biorefinery operation typically consumes vast quantities of freshwater, which is further exacerbated by the use of irrigated crop-based feedstocks [37]. Although, not considered at laboratory scale, water availability becomes increasingly crucial during industrial scale up, particularly in more arid climates. Water is required for all stages of the bioprocess from feedstock pretreatment to fermentation to downstream processing. It is also employed in the implementation of temperature and pH control [38]. Large quantities of water are necessary for most of the feedstock pretreatment methods discussed in Section 2.2. For example, in acid or alkaline pretreatment methods the catalyst must be mixed with water. In steam explosion and autohydrolysis methods, the biomass is immersed in water and undergoes organic solvent pretreatment before being washed with water. The subsequent enzymatic hydrolysis, media preparation and fermentation steps also require considerable volumes of freshwater. This water demand is even greater for continuous processes where plant cooling demand is increased. Cooling water may be recirculated in order to reduce both water and energy usage, however, more sustainable solutions are needed for the remaining steps of the bioprocess [39]. One possible solution is the use of a halophilic microorganism for the fermentation stage, this would allow use of saline water, the most abundant source on the planet, in place of freshwater. Saline water may not be suitable for the prior biomass pretreatment and enzymatic hydrolysis steps, which are sensitive to salinity deviations [40]. Despite this, its use in the fermentation step would reduce the overall freshwater demand of the process and hence production costs [41].
2.5. Metabolic Balance
Traditional multiple unit operation approaches (pretreatment–fermentation–recovery) are both time and resource intensive. Consolidated bioprocessing (CBP) strategies are an innovative potential solution with growing research interest [42,43]. Here the substrate hydrolysis and fermentation steps are combined in a single unit operation. This is achieved through the overexpression of genes encoding enzymes for both complex substrate metabolism and desirable product biosynthesis. This approach has significant metabolic constraints, however, as a strain engineered to degrade the feedstock and produce bio-chemicals cannot be optimized to efficiently perform both simultaneously. Typically, either low feedstock degradation rates or poor bio-product yields are reported [42,43]. One potential solution is the construction of a co-culture CBP system in which each microorganism is engineered and optimized to perform a single purpose efficiently [44].
2.6. Product Inhibition
Another challenge often faced during the scale up of biotechnological processes is product toxicity. Accumulation of the bio-product of interest can impair the cellular viability and productivity of the host if it exceeds a critical concentration in the fermentation broth [45]. The result of this is low substrate conversion and product yields. Ethanol, butanol and 1,3 propanediol are some examples of classical fermentation products affected by this issue. In a traditional batch ethanol fermentation, the fermenting yeast is able to tolerate ethanol concentrations up to around 10% (v/v), concentrations above this toxic level result in cell death [46]. During ABE (Acetone-Butanol-Ethanol) fermentation, Clostridia species ferment sugars into acetone, butanol and ethanol. Traditional batch fermentation is completely inhibited when the butanol concentration reaches around 13 g/L [47]. Similarly, 1,3 propanediol fermentations are inhibited when the product concentration exceeds around 80 g/L, this is associated with an increase in membrane fluidity and ultimately leads to cell death [48]. Several studies have focused on the metabolic engineering of the microbial strains in order to increase their tolerance towards specific products [49,50]. However, as reported in Section 2.5, if a microorganism is engineered to serve several purposes, low performances are typically observed. There is a critical demand for the development of alternative solutions to the classical metabolic engineering of a single strain to overcome such issues.
2.7. Continuous Cultivation of Microorganisms
Continuous fermentations are especially suited for the production of biomass for applications such as primary metabolite synthesis or dairy starting cultures. With reduced operational costs, increased efficiency and reduced down-time compared to batch processes, continuous fermentations are desirable for industrial biotechnological applications. However, higher initial capital expenditure is necessary. In addition, contamination risk is increased in continuous fermentations due to the long cultivation times, often exceeding 20–50 days, particularly as a constant supply of sterile fresh medium is needed. Genetic instability of cultivation is another challenge associated with continuous operation as cell mutations can accumulate over time. Such mutations may hinder productivity in longer operations [4].
3. Industrial Bioprocesses: Possible Solutions
The aforementioned challenges should be tackled through a combination of strategically designed microbial cell factories and innovative bioreactor processes. In this section, potential mitigations for each of the discussed bottlenecks are considered.
3.1. Cell Factories
Microbial cell factories are employed for the bioconversion of substrates into valuable products. Cell factories may be used to overproduce a native metabolite, a heterologous product, or a combination of native and heterologous proteins [51]. Microbes such as baker’s yeast and lactic acid bacteria [52] have been exploited by humans for millennia for the production of fermented foods and beverages. The native secondary metabolites of certain cell factories also have important pharmaceutical applications such as the antibiotic penicillin produced by Penicillium chrysogenum. As a result of advances in genome sequencing and molecular biology tools, the production of non-native products became possible in the late 20th century. In 1991 Baily defined metabolic engineering as “… the improvement of cellular activities by manipulation of enzymatic, transport, and regulatory functions of the cell with the use of recombinant DNA technology” [53]. Using such tools, cells became programmable and cell factory design evolved into an engineering discipline of its own. Scientists became capable of introducing heterologous genes into well-characterized workhorses like S. cerevisiae and Aspergillus niger. A popular analogy compares cells to airplanes where each individual part has a function and where parts can be combined to contribute to an overall function of the system [50]. Such techniques were initially limited to the heterologous expression of one or two genes for the production of a small molecule or protein of interest. A major industrial example of this, is the establishment of recombinant S. cerevisiae and Escherichia coli strains for the production of insulin [54]. However, as tools for high-throughput sequencing, gene assembly and cloning are becoming increasingly affordable, genome scale engineering is now feasible, allowing the construction of entire heterologous pathways. Examples of cell factories with more complex engineered metabolic pathways, which have been successfully scaled up to industrial scale include those for the antimalarial drug artemisinin and the sweetener stevia [55] using S. cerevisiae cell factories.
3.1.1. Glycerol as Feedstock: Yarrowia lipolytica
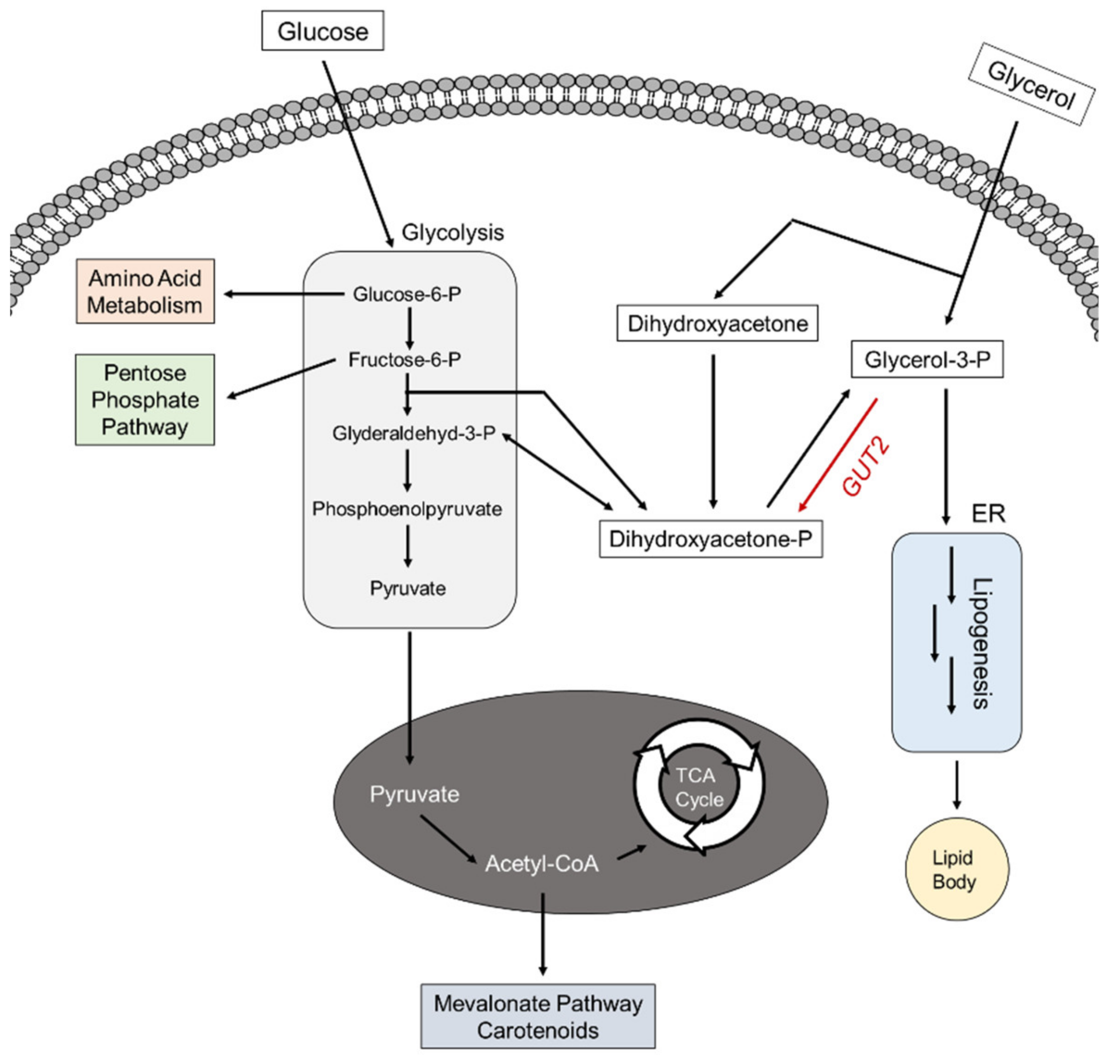
The oleaginous non-conventional yeast, Yarrowia lipolytica, was first discovered in 1970 by Wickerham et al. [56]. As a strict aerobe, Y. lipolytica is unable to metabolize sugars in the absence of oxygen [57]. Y. lipolytica is a promising candidate for the production of liquid biofuels as it is capable of accumulating high concentrations of lipids (>50% of dried biomass). In addition, unlike most species, which grow preferentially on glucose, this species prefers the much cheaper substrate, glycerol [58]. In 2013, Workman et al. discovered that the specific growth rate of Y. lipolytica was 25% higher at 0.3 h-1 when grown in media supplemented with glycerol compared to glucose, irrespective of the initial glycerol concentration [58]. In subsequent co-substrate cultivations, glycerol was consumed preferentially with glucose catabolism only initiating upon glycerol depletion [58]. This is largely due to the greater number of genes encoding proteins for glycerol uptake than hexose transport in its genome. In addition, glycerol may enter the central carbon metabolism via two different pathways through the use of either dihydroxyacetone or glycerol-3-phosphate as an intermediate, as summarized in Figure 1.
Although both routes are active, the primary pathway is that involving the glycerol-3-phosphate intermediate [58]. A correlation between glycerol metabolism and the species ability to accumulate lipids in lipid bodies has been elucidated [59]. When the gene encoding glycerol-3-phosphate dehydrogenase (GUT2), an important enzyme in the primary glycerol metabolic pathway, was knocked out, only the dihydroxyacetone-mediated pathway remained active. This resulted in a shift in carbon flux toward lipid synthesis and lipid accumulation increased threefold [59]. The use of crude glycerol also effects lipid production and accumulation, especially when stearin is used as co-substrate [41]. As a result of recent advancements in synthetic biology and metabolic engineering tools, heterologous expression of genes is now relatively straightforward. Such genes may be either expressed from plasmids or chromosomally integrated. CRISPR-Cas9-mediated genome editing methods, which offer greatly enhanced integration efficiency through the induction of targeted double strand breaks in the genome, have also been adapted for applications in Yarrowia with reasonable efficiency [60]. Omega-3 fatty acids have been found to have several health benefits [61]. Dupont recently commercialized the production of two omega-3 fatty acid products using recombinant Y. lipolytica. Yarrowia biomass is also a rich source of lipids and protein and has been approved for use as animal feed by the European Federation of Feedstuff Producers. A bioprocess is currently being developed for the sustainable production of Yarrowia biomass from flaxseed production waste [62]. Furthermore, Y. lipolytica is a promising cell factory for the biosynthesis of carotenoids such as beta-carotene, astaxanthin and zeaxanthin, compounds with applications as food colorants or antioxidants [60].
3.1.2. Tolerance and Conversion of Lignocellulosic Inhibitors: Trichosporon oleaginosus
Trichosporon oleaginosus (previously known as Cryptococcus curvatus, Candida curvata, Cutaneotrichosporon oleaginosus and Apriotrichum curvatum) is an oleaginous yeast within the basidomycete phylum [63,64], which was first isolated from the floors of cheese plants and floor drains at Iowa State University [65]. It is capable of naturally accumulating high lipid contents of over 20% of its biomass and under certain circumstances this may be increased further to as high as 60% [66]. T. oleaginosus is able to metabolize a broad range of carbon and nitrogen sources and grows optimally at a temperature of 28–30 °C and pH between 5.4 and 5.8 [66,67]. Although glucose is the preferred substrate of this yeast, it is able to metabolize a range of complex monosaccharide mixtures. T. oleaginosus can metabolize a wide variety of recalcitrant feedstocks [65] and tolerate several lignocellulosic pretreatment side-products, such as acetic acid, furfural and ammonia [68,69,70]. In addition, volatile fatty acids may be used as a substrate by the strain up to certain concentrations, above which they become toxic and can impair cell growth. Of the potential organic acid substrates, acetic acid is likely to be the most economically viable as higher lipid contents can be achieved compared to butyric or propionic acids [71]. A further advantage of T. oleaginosus is its ability to both tolerate and metabolize lignin-derived aromatic compounds, whilst remaining oleaginous [72].
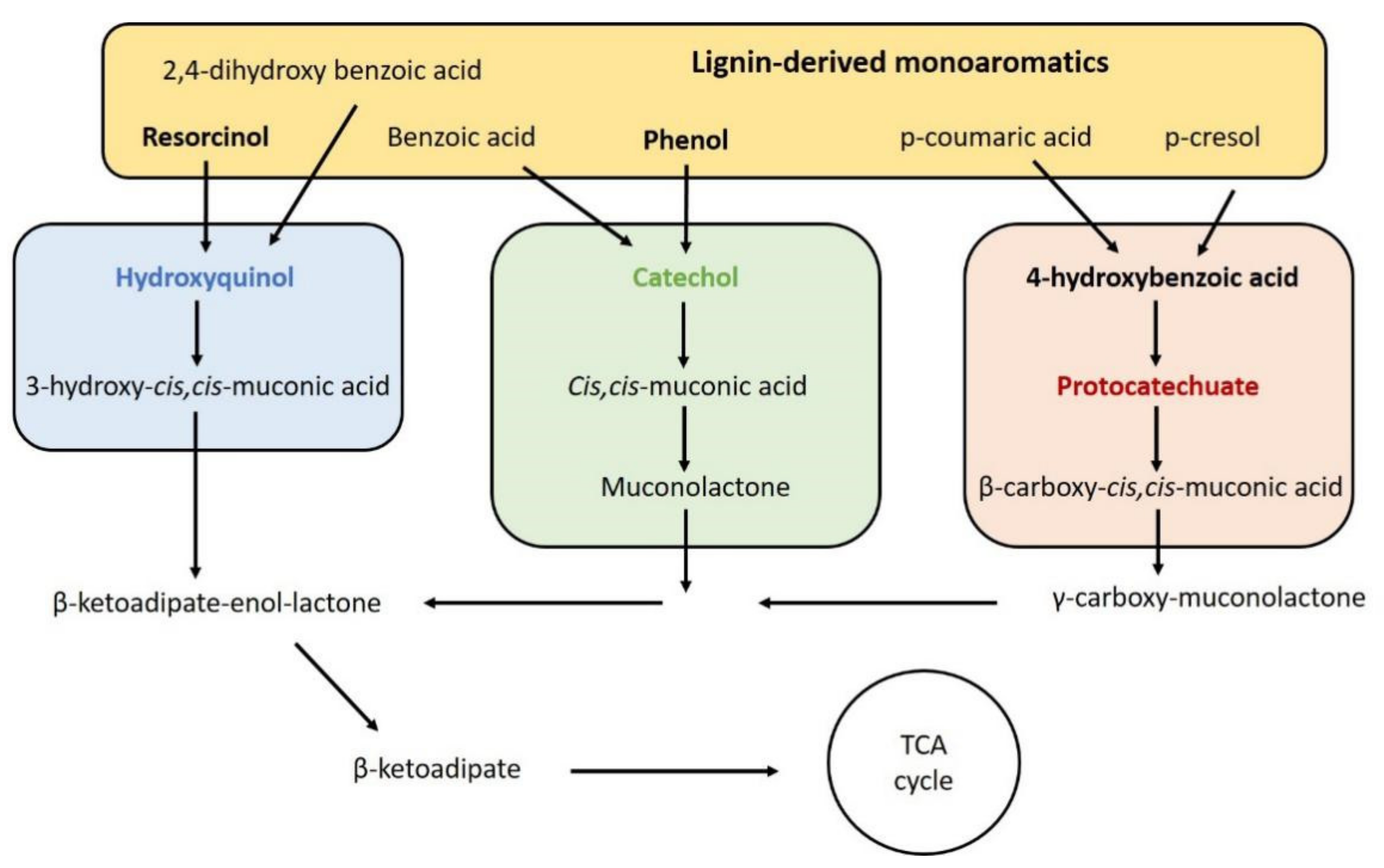
The microbial conversion of plant-derived phenolic compounds to aromatic compounds is known as “biological funneling”. The three main funneling pathways for aromatic metabolism are the hydroxyquinol, catechol, and protocatechuate pathways (Figure 2). Allison et al. selected resorcinol, phenol and 4-hydroxybenzoic acid (pHBA) as representative compounds for each of the funneling pathways, based on their relatively low minimum inhibitory concentration (MIC) [72]. When T. oleaginosus was cultivated on each of these three model lignin-derived aromatics, the specific growth rates achieved using resorcinol and pHBA were very similar at 0.097 h−1 and 0.092 h−1, respectively, whilst the growth rate observed using phenol was lower at 0.058 h−1. As expected, the growth rate on all of the phenolic compounds was lower than that on the preferential glucose substrate (≈0.157 h−1) [72,73]. Despite this, the three compounds were completely consumed after just several hours of cultivation. Interestingly, in high nitrogen media lipid accumulation was enhanced for the resorcinol and pHBA cultures at 7.21 and 7.55 %w/w, whilst it decreased in equivalent phenol cultures (5.26 %w/w), compared to the glucose control (6.72 %w/w) [74].
3.1.3. Tolerance to a Broad Range of pH Values: Ustilago cynodontis
Ustilago cynodontis is a true smut fungus from the Ustilaginaceae family, with the ability to produce a broad range of chemicals with potential applications in the food, pharmaceutical and chemical industries. Secondary metabolites of U. cynodontis of particular biotechnological interest include polyols, organic acids and extracellular glycolipids [75,76], as well as tricarboxylic acid (TCA) cycle intermediates. Overproduction of glycolipids in U. cynodontis cultures has been demonstrated by Morita et al. in 2008 [77]. In this work, a final mannosyerythritol lipid (MEL) titer of 1.4 g/L was achieved using soybean oil as a substrate. The MELs, which can be readily produced from vegetable oils by the fungus, are attractive biosurfactant candidates due to their biodegradability, mild production conditions and multifunctionality [78,79]. In addition, U. cynodontis can metabolize a variety of carbohydrate polymers and monomers derived from the degradation of renewable non-food biomass [75,80,81].
U. cynodontis is also a promising microbial alternative for the production of itaconic acid, a valuable intermediate in the production of a number of resins, plastics, adhesives, as well as bio-based polymers and novel fuels [81,82]. Itaconate is an unsaturated dicarboxylic acid with two pKa values at 3.8 and 5.5 [83]. As U. cynodontis is highly tolerant to low pH it is well suited to itaconate production. Of the several Ustilaginaceae family members in which itaconic acid has been evaluated to date, U. cynodontis (strain 2217) is the highest producer (≈3.3 g/L) [84]. Very recently, an engineered strain of U. cynodontis was used to improve itaconate production up to 0.61 g/g, in pH-controlled fed-batch reactions (pH 3.6), using glucose as a carbon source [85]. This specific U. cynodontis strain displays a stable yeast-like morphology resulting from deletion of the fuz7 gene, which encodes a MAPK protein involved in the regulation of tube formation and filamentous growth [85]. Differences in growth and itaconate production were observed depending on the pH of the culture medium. For example, at pH 1.9 both growth and itaconate production were inhibited, while at pH 3.6 optimal growth and maximum titers were observed [86]. Through the overexpression of genes involved in itaconate transport and pathway regulation (MTTA and RIA1), itaconate production was enhanced further [85].
3.1.4. Use of Non-Pure Water: Debaryomyces hansenii
Debaryomyces hansenii is a halophilic, non-conventional yeast, found naturally in several environments including seawater, soil, air and the ice of Antarctic and Arctic glaciers [87,88,89]. The presence of sodium in the growth medium was found to provide protection against external stresses, such as oxidative stress and low pH [90,91,92], leading to its classification as a halotolerant yeast. Additionally, Gustafsson and Norkrans in 1976 [93] and Adler et al. in 1985 [94], described the role of glycerol production and accumulation as a compatible solute in response to high extracellular osmolarity. More recent work on the characterization of D. hansenii in controlled bioreactors, revealed that high salt concentrations are necessary for optimal cell performance in this yeast, especially sodium salts [41]. This confirmed that D. hansenii is halophilic rather than simply halotolerant as previously hypothesized. At moderate to high salinity levels of around 0.5–1 M, D. hansenii exhibits an increased maximum specific growth rate (µmax). Very high concentrations of around 2 M, however, trigger a shift in metabolic strategy, where µmax is decreased and the biomass yield on substrate is increased. Hence, the survival strategy is switched from growing at the fastest possible rate to maximizing overall yield: slowing down the growth rate whilst still proliferating [41]. D. hansenii is of great interest within the food industry due to its “salt loving” character, where among other applications, it is used in the ripening process of sausages through the secretion of exopeptidases, development of flavor characteristics and production of cheeses [95,96,97,98,99].
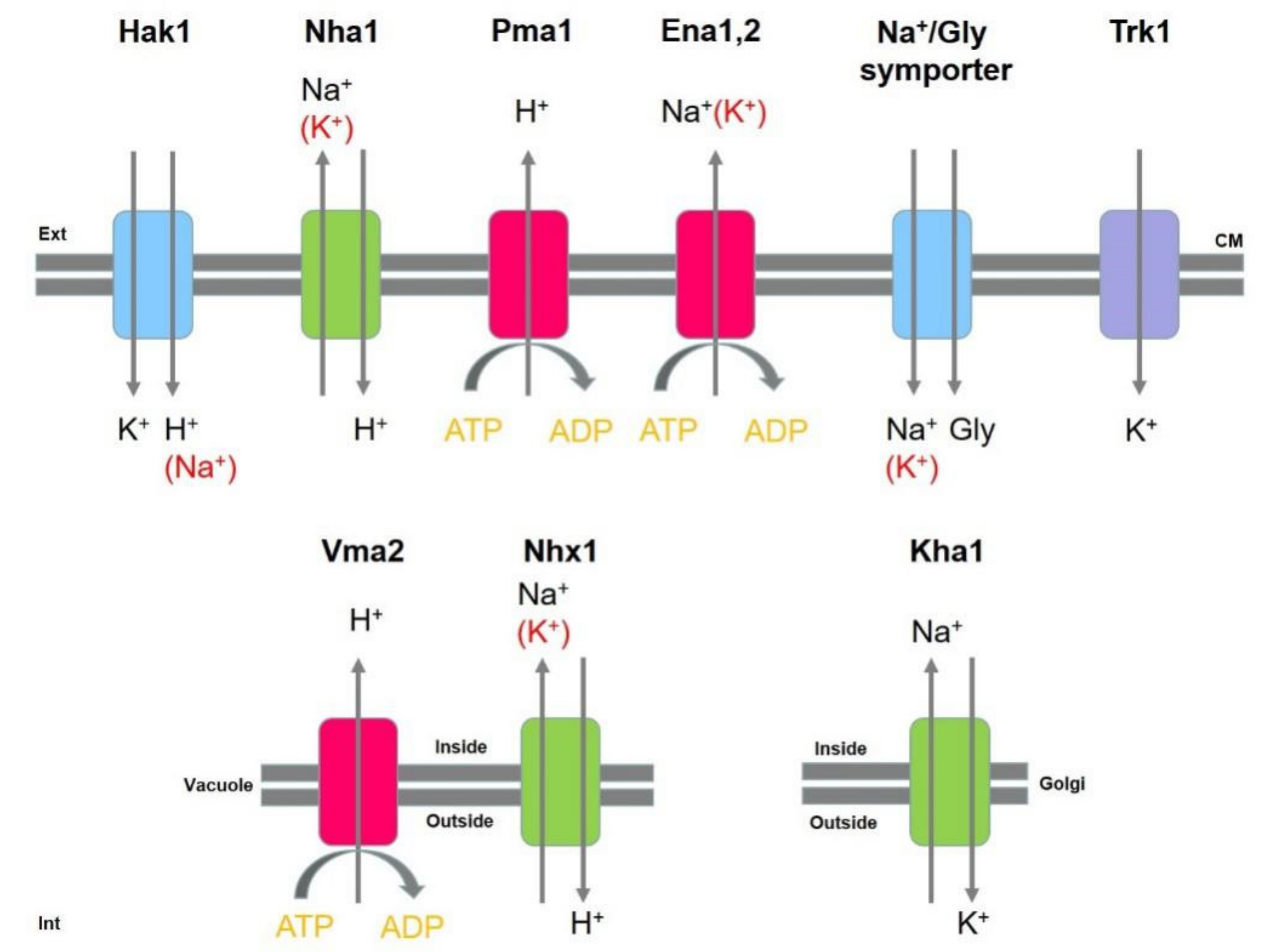
Although halotolerance is not yet completely understood in D. hansenii, the role of several membrane transporters and pumps in osmotic stress tolerance has been demonstrated. Figure 3 shows a schematic overview of the main transport systems and pumps involved in the maintenance of the intracellular osmolarity in D. hansenii. By transporting Na+ and K+ into the cell, H+-ATPases minimize excess H+ accumulation in the cytosol. Examples of H+-ATPases include, DhPma1 and DhVma2, which are located in the cell membrane and vacuole, respectively [100]. Two additional transport systems are used to extrude Na+ out of the cell: DhNha1, an active Na+/H+-antiporter [101], and DhEna1/DhEna2, which actively secretes Na+ by consuming ATP [102]. DhENA1 is expressed at high Na+ concentrations, whilst DhENA2 is only transcribed under high pH conditions. In addition, DhNhx1 and DhKha1 are H+/K+-antiporters described to decrease the cytosolic Na+ concentration by transferring Na+ into vacuoles and the Golgi apparatus respectively [103,104]. Due to their importance in cell performance, a further two K+ transporters have been identified in D. hansenii, DhHak1 and DhTrk1. DhHak1 is a K+/H+-symporter, which may also function as a K+/Na+-symporter in the presence of Na+. DhHAK1 (High Affinity K+ transport), on the other hand, is expressed only when the ion concentration is low [105]. As the K+ concentration increases, DhHAK1 is repressed and K+ uptake is then performed by DhTrk1 [106]. Finally, a Na+/glycerol-symporter has been also studied [107], using this symporter, D. hansenii is able to accumulate glycerol and use it as a compatible solute, as aforementioned. In the absence of Na+, K+ may also function as a co-transporter with glycerol.
The improved performance of D. hansenii under very harsh conditions such as high salinity, osmotic pressure, media acidification or nutrient scarcity, makes this yeast a suitable candidate for cultivation in high salinity media such as seawater. This is particularly desirable in arid climates where freshwater supplies are limited. Contamination risks are also reduced as such levels of salinity are toxic for most microorganisms, which could reduce sterilization requirements. The use of seawater for D. hansenii cultivation therefore offers the potential of reduced operational costs and increased production yields. Additional complex feedstock sources could also be explored for this yeast as a cell factory for biomass revalorization. An example is the use of algal biomass hydrolysates, which are rich in salt and inhibit growth of less tolerant organisms. Although D. hansenii would be highly suited as cell factory in processes involving algal biomass hydrolysates or seawater, as salt may be corrosive to traditional plant infrastructure, more work is needed to determine compatibility. Alternative materials may be required for unit operations continuously exposed to high salinity to facilitate cultivation of halophilic microorganisms in industrial biorefineries. The greatest challenge associated with the development of D. hansenii, however, is the lack of efficient molecular genome engineering tools despite its genome being completely sequenced in 2004 [108]. Further research is therefore critical to alleviating this bottleneck to allow the far-reaching biotechnological benefits of D. hansenii to be realized.
3.2. Bioreactor Configurations
During fermentation, the bioreactor is typically operated in either batch, fed-batch or continuous mode. Batch configuration offers the major advantage of inherent simplicity; however, increased downtime requirements typically limit potential yields. In contrast, in fed-batch and continuous configurations downtime is reduced significantly, at a cost of increased process complexity and control requirements [109]. The appropriate selection is highly dependent on the individual process, as bioreactor configuration can strongly influence overall process productivity, the metabolism and growth morphology of the particular microorganism should be taken into careful consideration. The nature of the bioreactor configuration is particularly important for secondary metabolite production; such molecules are typically decoupled from growth and therefore not produced when all required nutrients are in excess. Upon encountering unfavorable conditions, e.g., the depletion of a nutrient, the biosynthesis of secondary metabolites will be initiated by the microorganism. If a metabolite is produced under the condition of carbon source limitation, batch configuration would limit production to the very end of the fermentation. For this application fed-batch is likely to be more appropriate as this “stress” condition can be mimicked throughout the cultivation by slowly feeding the carbon source to limit accumulation in the medium. Using this strategy, the product of interest is synthesized throughout the fermentation rather than in the final stages only, maximizing overall productivity [110]. Substantial progress in the field of metabolic engineering for cell factory construction has been achieved recently, however, the development of novel reactor configurations to provide innovative solutions for particular fermentation processes is still necessary to ensure more biotechnological processes can be implemented at an industrial level.
3.2.1. Sequential Bioreactors
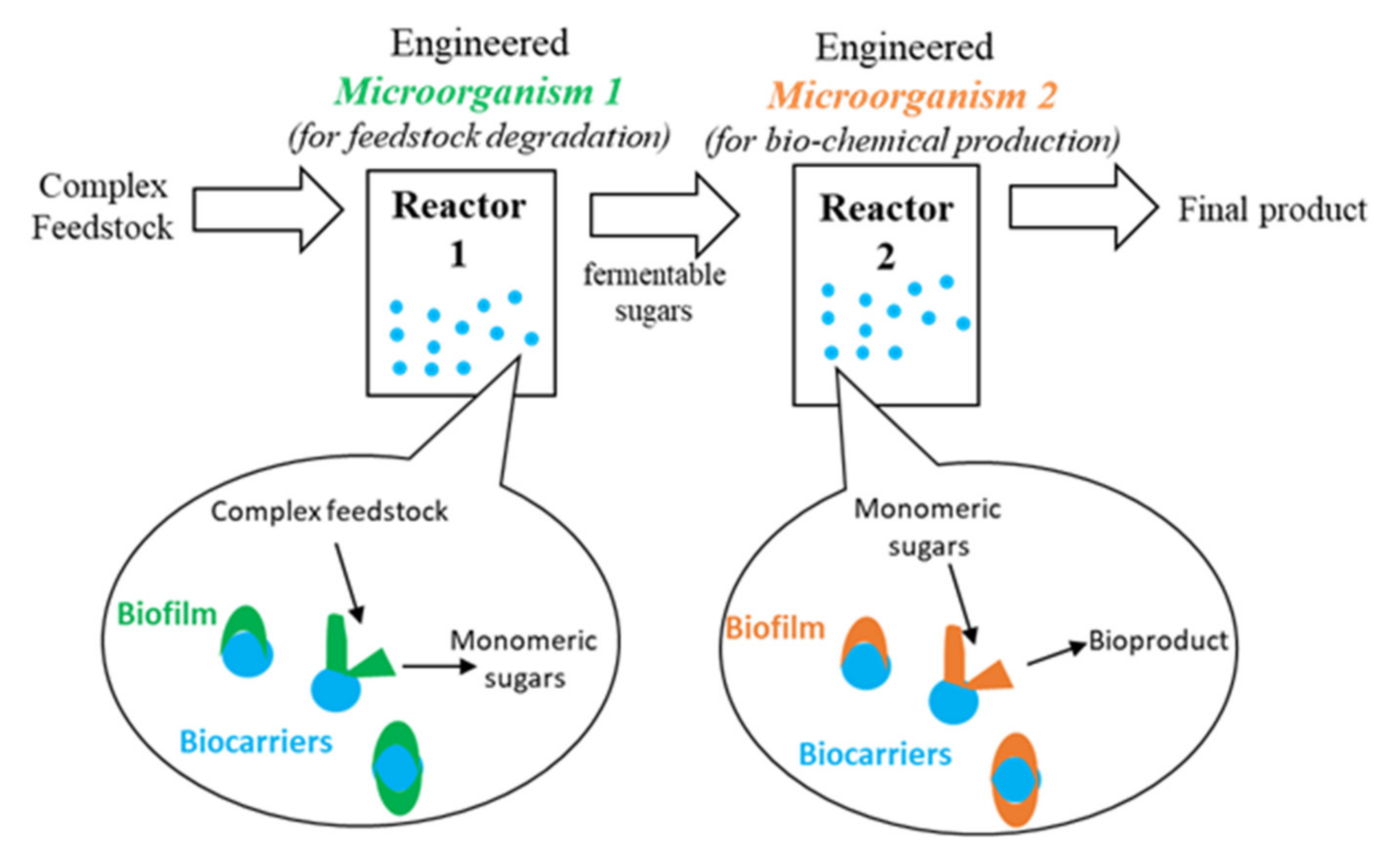
To ensure the successful establishment of more biorefineries, the aforementioned bottlenecks of poor metabolic flux balance and product inhibition must be alleviated. As discussed in Section 2.5, a CBP system is necessary to reduce reliance on expensive chemicals or enzymes for feedstock degradation. To reduce the metabolic burden associated with the expression of the enzymes required for feedstock degradation and product formation, two different microorganisms can be engineered, each optimized to perform one task. In order to avoid growth competition this could be achieved through the use of a sequential bioreactor configuration as depicted in Figure 4.
This concept is often employed in biogas plants; hydrolysis and acidogenesis take place first in a smaller tank, followed by methanogenesis in a larger tank [111]. This method could be adapted for the production of bio-commodities from lignocellulosic biomass. The system would be comprised of two vessels, reactor 1 and reactor 2, as depicted in Figure 4. In the first reactor, an engineered organism efficiently breaks down the raw lignocellulosic feedstock into fermentable sugars. The resulting product, consisting largely of monomeric sugars, is subsequently fed into reactor 2 where it is converted into the product of interest by a second engineered microorganism. Biofilm reactors can be used for this application to enhance performance, here the microorganisms attach to the surface of small beads as they grow and are retained in their respective reactors [112,113]. In the case of continuous bioreactor configurations, determination of the optimal dilution rate (D) and reactor volumes must consider both microorganisms, a compromise may therefore be necessary to maximize overall performance. A major issue associated with the proposed configuration is substrate consumption in the first reactor. The purpose of the microorganism in the first reactor 1 is to hydrolyze complex substrate to release simple sugars. However, care must be taken to minimize consumption of the resulting sugars and maximize availability for the second microorganism. Two possible strategies could be used to overcome this issue:
- (1)
- Use of a feedstock containing a carbon source more preferable to the microorganism than the hydrolysis product. For example, a feedstock containing or supplemented with crude glycerol could be used for certain microorganisms with a preference for glycerol. Provided an appropriate dilution rate is used, the resulting glucose will not be metabolized in reactor 1 and hence will be transferred to reactor 2 for use in product formation.
- (2)
- Through careful characterization of substrate consumption and cellulose hydrolysis kinetics in the first reactor, the dilution rate can be optimized such that recovery of the sugar product is maximized while ensuring minimal carbon source availability for substrate degradation by the first microorganism.
Through coupling this innovative bioreactor configuration with strategic metabolic engineering approaches, an effective CBP process can be developed. Metabolic flux imbalances can be effectively minimized by distributing the pathways across two microorganisms. In addition, the use of a two-stage bioreactor process eliminates the substrate competition challenge associated with traditional co-cultures, ultimately improving overall process productivity.
The proposed configuration could be adopted to treat different feedstocks by using different degrading microorganisms. Depending on both the feedstock and the microorganism used in the first reactor, more or less inhibiting molecules will be encountered. However, due to the nature of the pretreatment performed in bioreactor 1 (which is merely biological), no further purification and/or sterilization steps will be required from there onwards.
3.2.2. Integrated Downstream Process Units
Most valuable biosynthetic products are unfortunately toxic to the host microorganism at the high concentrations required to ensure economic feasibility at an industrial scale [114]. Metabolic engineering techniques coupled with adaptive laboratory evolution (ALE) can increase tolerance to these products. However, in most cases toxicity thresholds for such molecules remain too low to ensure sufficient titers for successful industrial production. In situ recovery methods, in which the fermentation and recovery steps are integrated can provide a potential solution. Several in situ recovery methods have been reported in the literature [114], each of them favored by different fermentation configurations.
For batch reactors, gas stripping and pervaporation are among the most suitable in situ recovery methods. In a gas stripping process, the toxic product preferentially diffuses into a solvent gas stream which passes through the fermentation broth. This technique effectively reduces product accumulation without harming the culture and does not require expensive equipment or extensive plant modifications. However, it strictly depends on affinity of the product for the gaseous solvent, which is typically steam, air, an inert gas or hydrocarbon gas [115]. In a recent study, a gas recycle with a flowrate between 0.3 and 0.6 vvm was used, this effectively kept butanol concentrations below inhibitory levels throughout the batch fermentation [116]. Pervaporation methods involve attaching an additional process unit to the bioreactor. Separation is achieved via partial vaporization through either a non-porous or porous membrane. The driving force for the separation is the difference in the partial pressure of the components on either side of the membrane. An appropriate membrane should be selected such that affinity for the product of interest is maximized whilst minimizing costs [117]. Using of a novel silicalite-1/polydimethylsiloxane/polyvinylidene fluoride hybrid membrane, an ethanol productivity of 1.6 g/L*h was achieved in an integrated ethanol fermentation–pervaporation process [118]. Adsorption is a more promising method for fed-batch processes; such techniques make use of a porous solid adsorbent or a resin, to which the compound binds to the surface [119]. Recently, Raganati et al., obtained encouraging results using an Amberlite XAD-7 resin for the adsorption of butanol produced during an ABE fermentation [120]. Numerous studies involving the optimization of these techniques are available in the literature [118,119,120]. However, examples of effective integrated fermentation and downstream processing methods for the recovery of heterologous products of metabolically engineered microorganisms at an industrial scale remain sparse.
4. Conclusions
In this review, key issues associated with biorefinery platforms were discussed in detail. Those most significant included a lack of a reliable substrate supply, high water consumption rates and the metabolic burden associated with the heterologous pathways. Novel cell factories such as Yarrowia lipolytica, Trichosporon oleaginosus, Ustilago cynodontis and Debaryomyces hansenii with promising natural characteristics of preferential glycerol catabolism, the ability to metabolize lignocellulosic inhibitors, broad pH tolerance and affinity for non-pure water sources, respectively, were reviewed. An innovative bioreactor configuration using a sequential fermentation system was introduced; by distributing the key pathways across two microorganisms, the metabolic burden can be minimized and productivity enhanced. The solutions proposed could be adopted in a synergistic way in order to boost further biotechnology processes to an industrial level.
Author Contributions
Conceptualization, A.P. and J.L.M.; Investigation, C.N., I.H.J. and A.P.; Writing-Original Draft Preparation, C.N., I.H.J., J.L.M. and A.P.; Writing-Review and Editing, C.N., I.H.J., J.L.M. and A.P.; Supervision, J.L.M. and A.P.; Funding acquisition, J.L.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Novo Nordisk Fonden within the framework of the Fermentation Based Biomanufacturing Initiative. Grant Number NNF17SA0031362.
Acknowledgments
The authors thank Laura Walls and Christopher Workman for their kind revision of the English language.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Cherubini, F.; Jungmeier, G.; Wellisch, M.; Willke, T.; Skiadas, I.V.; Van Ree, R.; De Jong, E. Toward a common classification approach for biorefinery systems. Biofuels Bioprod. Biorefining 2009, 3, 534–546. [Google Scholar] [CrossRef]
- Hingsamer, M.; Jungmeier, G. Biorefineries. In The Role of Bioenergy in the Emerging Bioeconomy; Lago, C., Caldes, N., Lecon, Y., Eds.; Elsevier Inc.: Madrid, Spain, 2019; pp. 179–222. [Google Scholar]
- Caullet, C.; Le Notre, J. Vegetable oil Biorefineries. In Industrial Biorefinery & White Biotechnology; Pandey, A., Hofer, R., Larroche, C., Eds.; Elsevier Inc.: London, UK, 2015; pp. 247–270. [Google Scholar]
- Waites, M.J.; Morgan, N.L.; Rockey, J.S.; Higton, G. Industrial Microbiology: An Introduction; Blackwell Science Ltd.: Oxford, UK, 2001. [Google Scholar]
- Novo Nordisk. Available online: https://www.novonordisk.com/ (accessed on 14 April 2020).
- Kjeldsen, T.; Balschmidt, P.; Diers, I.; Hach, M.; Kaarsholm, N.C.; Ludvigsen, S. Expression of insulin in yeast: The importance of molecular adaptation for secretion and conversion. Biotechnol. Genet. Eng. Rev. 2001, 18, 89–121. [Google Scholar] [CrossRef] [PubMed]
- Biosynthia. Available online: https://www.biosyntia.com/ (accessed on 14 April 2020).
- Novozymes. Available online: https://www.novozymes.com/en (accessed on 14 April 2020).
- LanzaTech. Available online: https://www.lanzatech.com/ (accessed on 14 April 2020).
- Marcellin, E.; Behrendorff, J.B.Y.H.; Nagaraju, S.; DeTissera, S.; Segovia, S.; Palfreyman, R.W.; Daniell, J.; Licona-Cassani, C.; Quek, L.-E.; Speight, R.; et al. Low carbon fuels and commodity chemicals from waste gases – systematic approach to understand energy metabolism in a model acetogen. Green Chem. 2016, 18, 3020–3028. [Google Scholar] [CrossRef] [Green Version]
- DSM. Available online: https://www.dsm.com/corporate/home.html (accessed on 15 April 2020).
- Cargill. Available online: https://www.cargill.com/home (accessed on 15 April 2020).
- Lonza. Available online: https://www.lonza.com/ (accessed on 15 April 2020).
- Workman, M.; Andersen, M.R.; Thykaer, J. Integrated Approaches for Assessment of Cellular Performance in Industrially Relevant Filamentous Fungi. Ind. Biotechnol. 2013, 9, 337–344. [Google Scholar] [CrossRef]
- Klímek, P.; Wimmer, R.; Mishra, P.K.; Kúdela, J. Utilizing brewer’s-spent-grain in wood-based particleboard manufacturing. J. Clean. Prod. 2017, 141, 812–817. [Google Scholar] [CrossRef]
- Procentese, A.; Raganati, F.; Olivieri, G.; Russo, M.E.; Rehmann, L.; Marzocchella, A. Deep Eutectic Solvents pretreatment of agro-industrial food waste. Biotechnol. Biofuels 2018, 11, 37. [Google Scholar] [CrossRef]
- Procentese, A.; Raganati, F.; Olivieri, G.; Russo, M.E.; De La Feld, M.; Marzocchella, A. Renewable feedstocks for biobutanol production by fermentation. New Biotechnol. 2017, 39, 135–140. [Google Scholar] [CrossRef]
- Berlowska, J.; Cieciura-Wloch, W.; Kalinowska, H.; Kręgiel, D.; Borowski, S.; Pawlikowska, E.; Binczarski, M.; Witonska, I.A. Enzymatic Conversion of Sugar Beet Pulp: A Comparison of Simultaneous Saccharification and Fermentation and Separate Hydrolysis and Fermentation for Lactic Acid Production. Food Technol. Biotechnol. 2018, 56, 188–196. [Google Scholar] [CrossRef]
- Prasad, S.; Singh, A.; Joshi, H. Ethanol as an alternative fuel from agricultural, industrial and urban residues. Resour. Conserv. Recycl. 2007, 50, 1–39. [Google Scholar] [CrossRef]
- Ameram, N.; Muhammad, S.; Yusof, N.A.A.N.; Ishak, S.; Ali, A.; Shoparwe, N.F.; Ter, T.P. Chemical composition in sugarcane bagasse: Delignification with sodium hydroxide. Malays. J. Fundam. Appl. Sci. 2019, 15, 232–236. [Google Scholar] [CrossRef]
- Procentese, A.; Raganati, F.; Olivieri, G.; Russo, M.E.; De La Feld, M.; Marzocchella, A. Agro Food Wastes and Innovative Pretreatments to Meet Biofuel Demand in Europe. Chem. Eng. Technol. 2019, 42, 954–961. [Google Scholar] [CrossRef]
- Procentese, A.; Raganati, F.; Olivieri, G.; Russo, M.E.; Marzocchella, A. Pre-treatment and enzymatic hydrolysis of lettuce residues as feedstock for bio-butanol production. Biomass- Bioenergy 2017, 96, 172–179. [Google Scholar] [CrossRef]
- RTaherzadeh, M.; Bolton, K.; Wong, J.; Pandey, A. Sustainable Resource Recovery and Zero Waste Approaches; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar] [CrossRef]
- Rajendran, K.; Lin, R.; Wall, D.M.; Murphy, J.D. Influential Aspects in Waste Management Practices. Sustainable Resource Recovery and Zero Waste Approaches 2019, 65–78. [Google Scholar] [CrossRef]
- McCoy, M. GLYCERIN SURPLUS. Chem. Eng. News Arch. 2006, 84, 7. [Google Scholar] [CrossRef]
- Rodrigues, A.; Bordado, J.; Dos Santos, R.G. Upgrading the Glycerol from Biodiesel Production as a Source of Energy Carriers and Chemicals—A Technological Review for Three Chemical Pathways. Energies 2017, 10, 1817. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Barrett, D.M.; Delwiche, M.J.; Stroeve, P. Methods for Pretreatment of Lignocellulosic Biomass for Efficient Hydrolysis and Biofuel Production. Ind. Eng. Chem. Res. 2009, 48, 3713–3729. [Google Scholar] [CrossRef]
- Zhang, X.; Yuan, Q.; Cheng, G. Deconstruction of corncob by steam explosion pretreatment: Correlations between sugar conversion and recalcitrant structures. Carbohydr. Polym. 2017, 156, 351–356. [Google Scholar] [CrossRef]
- Kim, D. Physico-Chemical Conversion of Lignocellulose: Inhibitor Effects and Detoxification Strategies: A Mini Review. Molecul 2018, 23, 309. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Franden, M.A.; Yang, Q.; Chou, Y.-C.; Zhang, M.; Pienkos, P.T. Identification of Inhibitors in Lignocellulosic Slurries and Determination of Their Effect on Hydrocarbon-Producing Microorganisms. Front. Bioeng. Biotechnol. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Kucharska, K.; Rybarczyk, P.; Hołowacz, I.; Łukajtis, R.; Glinka, M.; Kamiński, M. Pretreatment of Lignocellulosic Materials as Substrates for Fermentation Processes. Mol. 2018, 23, 2937. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Bao, J. Lignocellulose Pretreatment Using Acid as Catalyst. In Handbook of Biorefinery Research and Technology; Park, J., Ed.; Springer: Dordrecht, The Netherlands, 2018. [Google Scholar]
- Chin, J.Y.; Ab Rahim, S.K.E.; Abdullah, N.S. Studies on Alkaline Pretreatment of Sugarcane Bagasse and Rice Straw Hydrolysis for the Recovery of Reducing Sugar. Int. J. Curr. Res. Sci. Eng. Technol. 2018, 1, 26–31. [Google Scholar] [CrossRef]
- Novozymes. Bioenergy Application sheet Novozyme Cellic CTec3 HS. Available online: https://www.novozymes.com/en (accessed on 14 April 2020).
- Azhar, S.H.M.; Abdulla, R.; Jambo, S.A.; Marbawi, H.; Gansau, J.A.; Faik, A.A.M.; Rodrigues, K.F. Yeasts in sustainable bioethanol production: A review. Biochem. Biophys. Rep. 2017, 10, 52–61. [Google Scholar] [CrossRef]
- Hetényi, K.; Nemeth, A.; Sevella, B. Role of pH-regulation in lactic acid fermentation: Second steps in a process improvement. Chem. Eng. Process. Process. Intensif. 2011, 50, 293–299. [Google Scholar] [CrossRef]
- Emmenegger, M.F.; Pfister, S.; Koehler, A.; De Giovanetti, L.; Arena, A.P.; Zah, R. Taking into account water use impacts in the LCA of biofuels: An Argentinean case study. Int. J. Life Cycle Assess. 2011, 16, 869–877. [Google Scholar] [CrossRef]
- Chiu, Y.-W.; Suh, S.; Pfister, S.; Hellweg, S.; Koehler, A. Measuring ecological impact of water consumption by bioethanol using life cycle impact assessment. Int. J. Life Cycle Assess. 2011, 17, 16–24. [Google Scholar] [CrossRef]
- Hossain, G.; Liu, L.; Du, G. Industrial Bioprocesses and the Biorefinery Concept. Current Developments in Biotechnology and Bioengineering 2017, 3–27. [Google Scholar] [CrossRef]
- Yang, M.; Wang, J.; Nan, Y.; Zhang, J.; Li, L.; Liu, G.; Vepsäläinen, J.; Kuittinen, S.; Pappinen, A. Effect of salts formed by neutralization for the enzymatic hydrolysis of cellulose and acetone–butanol–ethanol fermentation. RSC Adv. 2019, 9, 33755–33760. [Google Scholar] [CrossRef] [Green Version]
- Navarrete, C.; Frost, A.T.; Ramos-Moreno, L.; Martinez, J.L. A Physiological Characterization in Controlled Bioreactors Reveals a Novel Survival Strategy for Debaryomyces hansenii at High Salinity and Confirms its Halophilic Behavior. Biorxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Conway, P.M.; Cervenka, N.D.; Cui, J.; Maloney, M.; Olson, D.G.; Lynd, L.R. Metabolic engineering of Clostridium thermocellum for n-butanol production from cellulose. Biotechnol. Biofuels 2019, 12, 186. [Google Scholar] [CrossRef]
- Song, X.; Li, Y.; Wu, Y.; Cai, M.; Liu, Q.; Gao, K.; Zhang, X.; Bai, Y.; Xu, H.; Qiao, M. Metabolic engineering strategies for improvement of ethanol production in cellulolytic Saccharomyces cerevisiae. FEMS Yeast Res. 2018, 18, 18–26. [Google Scholar] [CrossRef]
- Kumar, R.; Tabatabaei, M.; Karimi, K.; Horváth, I.S. Recent updates on lignocellulosic biomass derived ethanol—A review. Biofuel Res. J. 2016, 3, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Shih-Hsin, H.; Kengo, S.; Riaan den, H.; Kentaro, I.; Chiaki, O.; Willem, H.Z.; Tomohisa, H. Engineering of a novel cellulose-adherent cellulolytic Saccharomyces cerevisiae for cellulolitic biofuel production. Sci. Rep. 2016, 6, 1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Wu, D.; Lin, Y.; Wang, X.; Kong, H.; Tanaka, S. Substrate and Product Inhibition on Yeast Performance in Ethanol Fermentation. Energy Fuels 2015, 29, 1019–1027. [Google Scholar] [CrossRef]
- Wen, Z.; Ledesma-Amaro, R.; Lin, J.; Jiang, Y.; Yang, S. Improved n-Butanol Production from Clostridium cellulovorans by Integrated Metabolic and Evolutionary Engineering. Appl. Environ. Microbiol. 2019, 85, 7–24. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Yun, J.; Zhang, H.; Magocha, T.A.; Zabed, H.M.; Xue, Y.; Fokum, E.B.; Sun, W.; Qi, X. Genetically Engineered Strains: Application and Advances for 1,3-Propanediol Production from Glycerol. Food Technol. Biotechnol. 2017, 56, 3–15. [Google Scholar] [CrossRef]
- Mezzina, M.P.; Álvarez, D.S.; Egoburo, D.E.; Peña, R.D.; Nikel, P.I.; Pettinari, M.J. A New Player in the Biorefineries Field: Phasin PhaP Enhances Tolerance to Solvents and Boosts Ethanol and 1,3-Propanediol Synthesis in Escherichia coli. Appl. Environ. Microbiol. 2017, 83, e00662-17. [Google Scholar] [CrossRef] [Green Version]
- Davy, A.M.; Kildegaard, H.F.; Andersen, M.R. Cell Factory Engineering. Cell Syst. 2017, 4, 262–275. [Google Scholar] [CrossRef] [Green Version]
- Karagiosis, S.A.; Baker, S.E. Fungal Cell Factories. Food and Industrial Bioproducts and Bioprocessing 2012, 205–219. [Google Scholar] [CrossRef]
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Bailey, J. Toward a science of metabolic engineering. Science 1991, 252, 1668–1675. [Google Scholar] [CrossRef]
- Baeshen, N.A.; Baeshen, M.N.; Sheikh, A.; Bora, R.S.; Ahmed, M.M.M.; Ramadan, H.A.; Saini, K.S.; Redwan, E.M. Cell factories for insulin production. Microb. Cell Factories 2014, 13, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bomgardner, M. Cargill, DSM start up stevia factory. C&EN Glob. Enterp. 2019, 97, 14–15. [Google Scholar] [CrossRef]
- Wickerham, L.J.; Kurtzman, C.P.; Herman, A.I. Sexual Reproduction in Candida lipolytica. Am. Assoc. Adv. Sci. 1970, 167, 1141. [Google Scholar] [CrossRef] [PubMed]
- Timoumi, A.; Cléret, M.; Bideaux, C.; Guillouet, S.E.; Allouche, Y.; Molina-Jouve, C.; Fillaudeau, L.; Gorret, N. Dynamic behavior of Yarrowia lipolytica in response to pH perturbations: Dependence of the stress response on the culture mode. Appl. Microbiol. Biotechnol. 2016, 101, 351–366. [Google Scholar] [CrossRef]
- Workman, M.; Holt, P.; Thykaer, J. Comparing cellular performance of Yarrowia lipolytica during growth on glucose and glycerol in submerged cultivations. AMB Express 2013, 3, 58. [Google Scholar] [CrossRef] [Green Version]
- Beopoulos, A.; Mrozova, Z.; Thevenieau, F.; Le Dall, M.-T.; Hapala, I.; Papanikolaou, S.; Chardot, T.; Nicaud, J.-M. Control of Lipid Accumulation in the Yeast Yarrowia lipolytica. Appl. Environ. Microbiol. 2008, 74, 7779–7789. [Google Scholar] [CrossRef] [Green Version]
- Larroude, M.; Celinska, E.; Back, A.; Thomas, S.; Nicaud, J.-M.; Ledesma-Amaro, R. A synthetic biology approach to transform Yarrowia lipolytica into a competitive biotechnological producer of β-carotene. Biotechnol. Bioeng. 2017, 115, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Xie, D.; Jackson, E.N.; Zhu, Q. Sustainable source of omega-3 eicosapentaenoic acid from metabolically engineered Yarrowia lipolytica: From fundamental research to commercial production. Appl. Microbiol. Biotechnol. 2015, 99, 1061–1062. [Google Scholar] [CrossRef] [Green Version]
- Juszczyk, P.; Rymowicz, W.; Kita, A.; Rywińska, A. Biomass production by Yarrowia lipolytica yeast using waste derived from the production of ethyl esters of polyunsaturated fatty acids of flaxseed oil. Ind. Crop. Prod. 2019, 138, 138. [Google Scholar] [CrossRef]
- Fell, J.W.; Boekhout, T.; Fonseca, A. Biodiversity and systematics of basidiomycetous yeasts as determined by large-subunit rDNA D1/D2 domain sequence analysis. Int. J. Syst. Evol. Micr. 2002, 50, 1351–1371. [Google Scholar] [CrossRef]
- Gujjari, P.; Coumes, K.; Zhou, J.J.; Suh, S.-O. Characterization of oleaginous yeasts revealed two novel species: Trichosporon cacaoliposimilis sp. nov. and Trichosporon oleaginosus sp. nov. Mycologia 2011, 103, 1110–1118. [Google Scholar] [CrossRef]
- Yaguchi, A.; Rives, D.; Blenner, M.A. New kids on the block: Emerging oleaginous yeast of biotechnological importance. AIMS Microbiol. 2017, 3, 227–247. [Google Scholar] [CrossRef] [PubMed]
- Moon, N.J.; Hammond, E.; Glatz, B.A. Conversion of Cheese Whey and Whey Permeate to Oil and Single-Cell Protein. J. Dairy Sci. 1978, 61, 1537–1547. [Google Scholar] [CrossRef]
- Bednarski, W.; Leman, J.; Tomasik, J. Utilization of beet molasses and whey for fat biosynthesis by a yeast. Agric. Wastes 1986, 18, 19–26. [Google Scholar] [CrossRef]
- Christophe, G.; Deo, J.L.; Kumar, V.; Nouaille, R.; Fontanille, P.; Larroche, C. Production of Oils from Acetic Acid by the Oleaginous Yeast Cryptococcus curvatus. Appl. Biochem. Biotechnol. 2011, 167, 1270–1279. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zheng, Y.; Dorgan, K.M.; Chen, S. Oil production by oleaginous yeasts using the hydrolysate from pretreatment of wheat straw with dilute sulfuric acid. Bioresour. Technol. 2011, 102, 6134–6140. [Google Scholar] [CrossRef]
- Zheng, Y.; Chi, Z.; Ahring, B.K.; Chen, S. Oleaginous yeast Cryptococcus curvatus for biofuel production: Ammonia’s effect. Biomass- Bioenergy 2012, 37, 114–121. [Google Scholar] [CrossRef]
- Park, G.W.; Chang, H.N.; Jung, K.; Seo, C.; Kim, Y.-C.; Choi, J.H.; Woo, H.C.; Hwang, I.-J. Production of microbial lipid by Cryptococcus curvatus on rice straw hydrolysates. Process. Biochem. 2017, 56, 147–153. [Google Scholar] [CrossRef]
- Yaguchi, A.; Robinson, A.; Mihealsick, E.; Blenner, M.A. Metabolism of aromatics by Trichosporon oleaginosus while remaining oleaginous. Microb. Cell Factories 2017, 16, 206. [Google Scholar] [CrossRef] [Green Version]
- Xiaolei, Z.; Jiaxin, C.; Di, W.; Ji, L.; Rajeshwar, D.T.; Rao, Y.S. Economical lipid production from Trichosporon oleaginosus via dissolved oxygen adjustment and crude glycerol addition. Bioresour. Technol. 2019, 273, 288–296. [Google Scholar]
- Chen, J.; Zhang, X.; Drogui, P.; Tyagi, R. The pH-based fed-batch for lipid production from Trichosporon oleaginosus with crude glycerol. Bioresour. Technol. 2018, 259, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Feldbrügge, M.; Kellner, R.; Schipper, K. The biotechnological use and potential of plant pathogenic smut fungi. Appl. Microbiol. Biotechnol. 2013, 97, 3253–3265. [Google Scholar] [CrossRef] [PubMed]
- Guevarra, E.D.; Tabuchi, T. Accumulation of itaconic, 2-hydroxyparaconic, itatartaric, and malic acids by strains of the genus Ustilago. Agric. Biol. Chem. 1990, 54, 2353–2358. [Google Scholar] [CrossRef] [Green Version]
- Morita, T.; Konishi, M.; Fukuoka, T.; Imura, T.; Kitamoto, D. Identification of Ustilago cynodontis as a new producer of glycolipid biosurfactants, mannosylerythritol lipids, based on ribosomal DNA sequences. J. Oleo Sci. 2008, 57, 549–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, S. Biological amphiphiles (microbial surfactants). Curr. Opin. Colloid. Interface Sci. 2002, 7, 12–20. [Google Scholar] [CrossRef]
- Kitamoto, D.; Isoda, H.; Nakahara, T. Functional and potential application of glycolipids biosurfactants. J. Biosci. Bioeng. 2002, 94, 187–201. [Google Scholar] [CrossRef]
- Couturier, M.; Navarro, D.; Olivé, C.; Chevret, D.; Haon, M.; Favel, A.; Lesage-Meessen, L.; Henrissat, B.; Coutinho, P.M.; Berrin, J.-G. Post-genomic analyses of fungal lignocellulosic biomass degradation reveal the unexpected potential of the plant pathogen Ustilago maydis. BMC Genom. 2012, 13, 57. [Google Scholar] [CrossRef] [Green Version]
- Geiser, E.; Wierckx, N.; Zimmermann, M.; Blank, L.M. Identification of an endo-1,4-beta-xylanase of Ustilago maydis. BMC Biotechnol. 2013, 13, 59. [Google Scholar] [CrossRef] [Green Version]
- Willke, T.; Vorlop, K.-D. Biotechnological production of itaconic acid. Appl. Microbiol. Biotechnol. 2001, 56, 289–295. [Google Scholar] [CrossRef]
- Geilen, F.M.A.; Engendahl, B.; Harwardt, A.; Marquardt, W.; Klankermayer, J.; Leitner, W. Selective and Flexible Transformation of Biomass-Derived Platform Chemicals by a Multifunctional Catalytic System. Angew. Chem. 2010, 122, 5642–5646. [Google Scholar] [CrossRef]
- Klement, T.; Büchs, J. Itaconic acid – A biotechnological process in change. Bioresour. Technol. 2013, 135, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Tehrani, H.H.; Saur, K.; Tharmasothirajan, A.; Blank, L.M.; Wierckx, N. Process engineering of pH tolerant Ustilago cynodontis for efficient itaconic acid production. Microb. Cell Factories 2019, 18, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tehrani, H.H.; Tharmasothirajan, A.; Track, E.; Blank, L.M.; Wierckx, N. Engineering the morphology and metabolism of pH tolerant Ustilago cynodontis for efficient itaconic acid production. Metab. Eng. 2019, 54, 293–300. [Google Scholar] [CrossRef]
- Norkrans, B. Studies on marine occurring yeasts: Growth related to pH, NaCl concentration and temperature. Arch. Microbiol. 1966, 54, 374–392. [Google Scholar] [CrossRef]
- Breuer, U.; Harms, H. Debaryomyces hansenii—An extremophilic yeast with biotechnological potential. Yeast 2006, 23, 415–437. [Google Scholar] [CrossRef] [PubMed]
- Gunde-Cimerman, N.; Ramos, J.; Plemenitaš, A. Halotolerant and halophilic fungi. Mycol. Res. 2009, 113, 1231–1241. [Google Scholar] [CrossRef]
- Almagro, A.; Prista, C.; Castro, S.; Quintas, C.; Madeira-Lopes, A.; Ramos, J.; Loureiro-Dias, M. Effects of salts on Debaryomyces hansenii and Saccharomyces cerevisiae under stress conditions. Int. J. Food Microbiol. 2000, 56, 191–197. [Google Scholar] [CrossRef]
- Papouskova, K.; Sychrová, H. The co-action of osmotic and high temperature stresses results in a growth improvement of Debaryomyces hansenii cells. Int. J. Food Microbiol. 2007, 118, 1–7. [Google Scholar] [CrossRef]
- Navarrete, C.; Siles, A.; Martãnez, J.L.; Calero, F.; Ramos, J.; Martãnez, J.L.; Ramos, J. Oxidative stress sensitivity inDebaryomyces hansenii. FEMS Yeast Res. 2009, 9, 582–590. [Google Scholar] [CrossRef] [Green Version]
- Gustafsson, L.; Norkrans, B. On the mechanism of salt tolerance. Production of glycerol and heat during growth of Debaryomyces hansenii. Arch Microbiol 1976, 110, 177–183. [Google Scholar] [CrossRef]
- Adler, L.; Blomberg, A.; Nilsson, A. Glycerol metabolism and osmoregulation in the salt-tolerant yeast Debaryomyces hansenii. J. Bacteriol. 1985, 162, 300–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadanho, M.; Almeida, J.; Sampaio, J. Assessment of yeast diversity in a marine environment in the south of Portugal by microsatellite-primed PCR. Antonie van Leeuwenhoek 2003, 84, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Butinar, L.; Zalar, P.; Frisvad, J.C.; Gunde-Cimerman, N. The genus Eurotium: Members of indigenous fungal community in hypersaline waters of salterns. FEMS Microbiol. Ecol. 2005, 51, 155–166. [Google Scholar] [CrossRef]
- Ramos, J.; Melero, Y.; Ramos-Moreno, L.; Michán, C.; Cabezas, L. Debaryomyces hansenii Strains from Valle De Los Pedroches Iberian Dry Meat Products: Isolation, Identification, Characterization, and Selection for Starter Cultures. J. Microbiol. Biotechnol. 2017, 27, 1576–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, M.L.D.C.; Delile, A.; Spinnler, H.-E.; Bonnarme, P.; Landaud, S. Comparison of volatile sulphur compound production by cheese-ripening yeasts from methionine and methionine–cysteine mixtures. Appl. Microbiol. Biotechnol. 2007, 75, 1447–1454. [Google Scholar] [CrossRef] [PubMed]
- Cano-García, L.; Belloch, C.; Flores, M. Impact of Debaryomyces hansenii strains inoculation on the quality of slow dry-cured fermented sausages. Meat Sci. 2014, 96, 1469–1477. [Google Scholar] [CrossRef] [Green Version]
- Prista, C.; Loureiro-Dias, M.; Montiel, V.; García, R.; Ramos, J. Mechanisms underlying the halotolerant way of Debaryomyces hansenii. FEMS Yeast Res. 2005, 5, 693–701. [Google Scholar] [CrossRef] [Green Version]
- Velkova, K.; Sychrová, H. The Debaryomyces hansenii NHA1 gene encodes a plasma membrane alkali-metal-cation antiporter with broad substrate specificity. Gene 2006, 369, 27–34. [Google Scholar] [CrossRef]
- Almagro, A.; Prista, C.; Benito, B.; Loureiro-Dias, M.C.; Ramos, J. Cloning and expression of two genes coding for sodium pumps in the sal-tolerant yeast Debaryomyces hansenii. J. Bacteriol. 2001, 183, 3251–3255. [Google Scholar] [CrossRef] [Green Version]
- Montiel, V.; Ramos, J.; Ramos, J. Intracellular Na+ and K+ distribution in Debaryomyces hansenii. Cloning and expression in Saccharomyces cerevisiae of DhNHX1. FEMS Yeast Res. 2007, 7, 102–109. [Google Scholar] [CrossRef]
- Carcãa-Salcedo, R.; Montiel, V.; Calero, F.; Ramos, J.; Ramos, J.; Salcedo, R.G. Characterization ofDhKHA1, a gene coding for a putative Na+transporter fromDebaryomyces hansenii. FEMS Yeast Res. 2007, 7, 905–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, J.L.; Sychrová, H.; Ramos, J. Monovalent cations regulate expression and activity of the Hak1 potassium transporter in Debaryomyces hansenii. Fungal Genet. Boil. 2011, 48, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Prista, C.; González-Hernández, J.C.; Ramos, J.; Loureiro-Dias, M. Cloning and characterization of two K+ transporters of Debaryomyces hansenii. Microbiol. 2007, 153, 3034–3043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, C.; Da Costa, M.S.; Van Uden, N. Osmoregulatory active sodium-glycerol co-transport in the halotolerant yeast Debaryomyces hansenii. Yeast 1990, 6, 187–191. [Google Scholar] [CrossRef]
- Dujon, B.; Sherman, D.J.; Fischer, G.; Durrens, P.; Casaregola, S.; Lafontaine, I.; De Montigny, J.; Marck, C.; Neuvéglise, C.; Talla, E.; et al. Genome evolution in yeasts. Nature 2004, 430, 35–44. [Google Scholar] [CrossRef]
- Spier, M.R.; Vandenberghe, L.P.; Medeiros, A.B.; Soccol, R. Application of different types of bioreactor in bioprocesses. Bioreactors: Design, Properties and Applications. P.G. Antolli and Z. Liu. Nova Science Publishers 2011. pp. 55-90; Nova Science Publishers: New York, NY, USA, 2011; pp. 55–90. [Google Scholar]
- Guerriero, G.; Berni, R.; Muñoz-Sánchez, J.A.; Apone, F.; Abdel-Salam, E.M.; Qahtan, A.A.; Alatar, A.; Cantini, C.; Cai, G.; Hausman, J.-F.; et al. Production of Plant Secondary Metabolites: Examples, Tips and Suggestions for Biotechnologists. Genes 2018, 9, 309. [Google Scholar] [CrossRef] [Green Version]
- Bharathiraja, B.; Sudharsana, T.; Jayamuthunagai, J.; Pravenkumar, R.; Chozhavendhan, S.; Iyyappan, J. Biogas production—A review on composition, fuel properties, feed stock and principles of anaerobic digestion. Renew Sust Ener. Rev. 2018, 90, 570–582. [Google Scholar] [CrossRef]
- Raganati, F.; Olivieri, G.; Procentese, A.; Russo, M.E.; Salatino, P.; Marzocchella, A. Butanol production by bioconversion of cheese whey in a continuous packed bed reactor. Bioresour. Technol. 2013, 138, 259–265. [Google Scholar] [CrossRef]
- Raganati, F.; Procentese, A.; Olivieri, G.; Russo, M.E.; Gotz, P.; Salatino, P.; Marzocchella, A. Butanol production by Clostridium acetobutylicum in a series of packed bed biofilm reactors. Chem. Eng. Sci. 2016, 152, 678–688. [Google Scholar] [CrossRef]
- Dauglis, A.J. Integreted fermentation and recovery processes. Curr. Opin. Biotechnol. 1994, 5, 192–195. [Google Scholar] [CrossRef]
- Ezeji, T.C.; Karcher, P.M.; Qureshi, N.; Blaschek, H.P. Improving performance of a gas stripping-based recovery system to remove butanol from Clostridium beijerinckii fermentation. Bioprocess Biosyst. Eng. 2005, 27, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Rochón, E.; Ferrari, M.; Lareo, C. Integrated ABE fermentation-gas stripping process for enhanced butanol production from sugarcane-sweet sorghum juices. Biomass- Bioenergy 2017, 98, 153–160. [Google Scholar] [CrossRef]
- Gaykawad, S.S.; Zha, Y.; Punt, P.J.; Van Groenestijn, J.W.; Van Der Wielen, L.A.; Straathof, A.J.J. Pervaporation of ethanol from lignocellulosic fermentation broth. Bioresour. Technol. 2013, 129, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Hu, S.; Chen, C.; Wang, Y.; Zhang, C.; Miao, Q.; Qin, P.; Tan, T. Immobilized ethanol fermentation coupled to pervaporation with silicalite-1/polydimethylsiloxane/polyvinylidene fluoride composite membrane. Bioresour. Technol. 2016, 220, 124–131. [Google Scholar] [CrossRef]
- Aljundi, I.H.; Belovich, J.M.; Talu, O. Adsorption of lactic acid from fermentation broth and aqueous solutions on Zeolite molecular sieves. Chem. Eng. Sci. 2005, 60, 5004–5009. [Google Scholar] [CrossRef] [Green Version]
- Raganati, F.; Procentese, A.; Olivieri, G.; Russo, M.E.; Salatino, P.; Marzocchella, A. Bio-butanol separation by adsorption on various materials: Assessment of isotherms and effects of other ABE-fermentation compounds. Sep. Purif. Technol. 2018, 191, 328–339. [Google Scholar] [CrossRef]
Figure 1.
Simplified overview of glucose and glycerol metabolism in Yarrowia lipolytica with key metabolites highlighted. GUT2 = glycerol-3-phosphate dehydrogenase. Figure based on results from: [58,59].

Figure 2.
Schematic depiction of the three main funneling pathways for aromatic metabolism: hydroxyquinol (blue), catechol (green), and protocatechuate (red). Trichosporon oleaginosus is able to tolerate and metabolize model lignin monoaromatics, and several intermediates derived from these pathways. Figure based on results from [72].
Figure 2.
Schematic depiction of the three main funneling pathways for aromatic metabolism: hydroxyquinol (blue), catechol (green), and protocatechuate (red). Trichosporon oleaginosus is able to tolerate and metabolize model lignin monoaromatics, and several intermediates derived from these pathways. Figure based on results from [72].

Figure 3.
Osmoregulation in Debaryomyces hansenii. Main transport systems and pumps located in the cell membrane (CM), vacuole and Golgi apparatus, and involved in the maintenance of the intracellular osmolarity, are represented in the figure. ATP: adenosine triphosphate. ADP: adenosine diphosphate.
Figure 3.
Osmoregulation in Debaryomyces hansenii. Main transport systems and pumps located in the cell membrane (CM), vacuole and Golgi apparatus, and involved in the maintenance of the intracellular osmolarity, are represented in the figure. ATP: adenosine triphosphate. ADP: adenosine diphosphate.

Figure 4.
Sequential bioreactor configuration.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Biotechnological processes running at industrial level.
| Company | Feedstock | Organism | Process | Product | Reference |
|---|---|---|---|---|---|
| Novo Nordisk | Glucose | S. cerevisiae | Fermentation | Human insulin | [5] |
| Biosynthia | Renewable sugars * | E. coli | Fermentation | Vitamins | [6,7] |
| Novozymes | * | Aspergillus oryzae Fungi Bacteria | Enzyme production | Bulk enzymes for food, beverages, agriculture, bioenergy and household care | [8] |
| LanzaTech | Dihydrogen, carbon monoxide and carbon dioxide | Clostridium autoethanogenum | Syngas fermentation | Ethanol | [9,10] |
| Avansya (DSM/Cargill) | Crude sugars * | S. cerevisiae | Recombinant yeast fermentation | Steviol glycosides | [11,12] |
| Lonza | * | Mammalian cells Pichia, Bacillus, E. coli | * | Pharmaceuticals, biopharmaceuticals, fine chemicals, enzymes | [13] |
* no further details disclosed by company.
Table 2.
Features of the main feedstock currently investigated.
| Feedstock | Lignin Content (%) | Availability | Amount (Mt y−1) | Transport Cost | Reference |
|---|---|---|---|---|---|
| Brewer’ Spent Grains | 15–27 | All year round | 8 | − 1 | [15,16] |
| Agricultural residues | 5–15 | Seasonal | 150 | + 2 | [17,19] |
| Potato peel | 30 | All year round | 0.5 | − | [17,19] |
| Fresh cut sector | 15–20 | All year round | 1.4 | − | [21,22] |
| Sugar beet pulp | 2 | Seasonal | 7 | − | [17,18] |
| Sugarcane bagasse | 25–28 | Seasonal | 10 | − | [20] |
| Coffee silverskin | 27 | All year round | 0.1 | − | [16,21] |
| OFMSW | 5 | All year round | 1000 | − | [23,24] |
1 The production is focused in a particular area thus low transportation costs are expected. 2 The production is spread in an extended area thus high transportation costs are expected.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Navarrete, C.; Jacobsen, I.H.; Martínez, J.L.; Procentese, A. Cell Factories for Industrial Production Processes: Current Issues and Emerging Solutions. Processes 2020, 8, 768. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8070768
AMA Style
Navarrete C, Jacobsen IH, Martínez JL, Procentese A. Cell Factories for Industrial Production Processes: Current Issues and Emerging Solutions. Processes. 2020; 8(7):768. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8070768
Chicago/Turabian StyleNavarrete, Clara, Irene Hjorth Jacobsen, José Luis Martínez, and Alessandra Procentese. 2020. "Cell Factories for Industrial Production Processes: Current Issues and Emerging Solutions" Processes 8, no. 7: 768. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8070768
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.