Batch Syngas Fermentation by Clostridium carboxidivorans for Production of Acids and Alcohols

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism and Culture Media
2.2. Batch Tests
2.3. Analytical Methods
- CO conversion (ξCO), the ratio between the CO converted and the CO fed at the beginning of the test;
- CO-to-product “i” yield coefficient (Yi/CO), the ratio between the produced mass of product “i” (cells or acid/solvent), and the decrease of the CO mass during the same time interval;
- The specific cell growth rate (μ). It was estimated at the beginning of the exponential phase as the slope of the biomass concentration (X) vs. time curve, on a log scale.
3. Results and Discussion
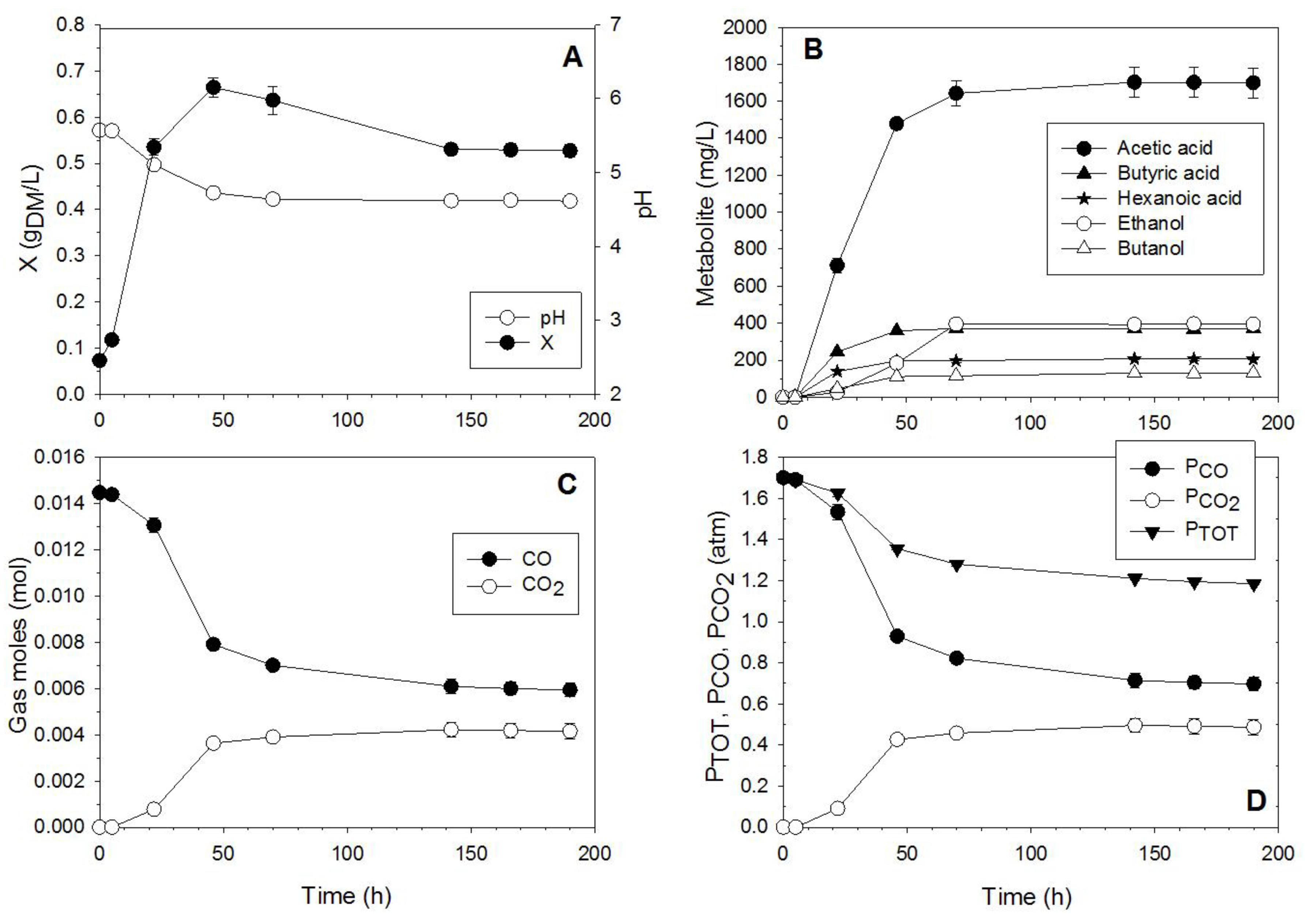
3.1. CO Fermentation
3.2. Effect of the CO Pressure
- PCO affected maximum cell concentration measured during fermentation tests. Maximum cell concentration was about 0.7 gDM/L for initial PCO between 0.5 and 2.2 atm, and decreased at higher initial PCO. Substrate inhibition might be responsible for the decrease of cell growth with PCO.
- CO conversion measured during fermentation tests carried out at VL/VG = 0.28, decreased with initial PCO. CO conversion was almost total at low initial PCO (0.5 and 1 atm), and it significantly decreased as initial PCO was higher than 1 atm.
- The best performance in terms of ethanol/butanol production was obtained with VL/VG = 0.28 at initial PCO = 1.7 atm.
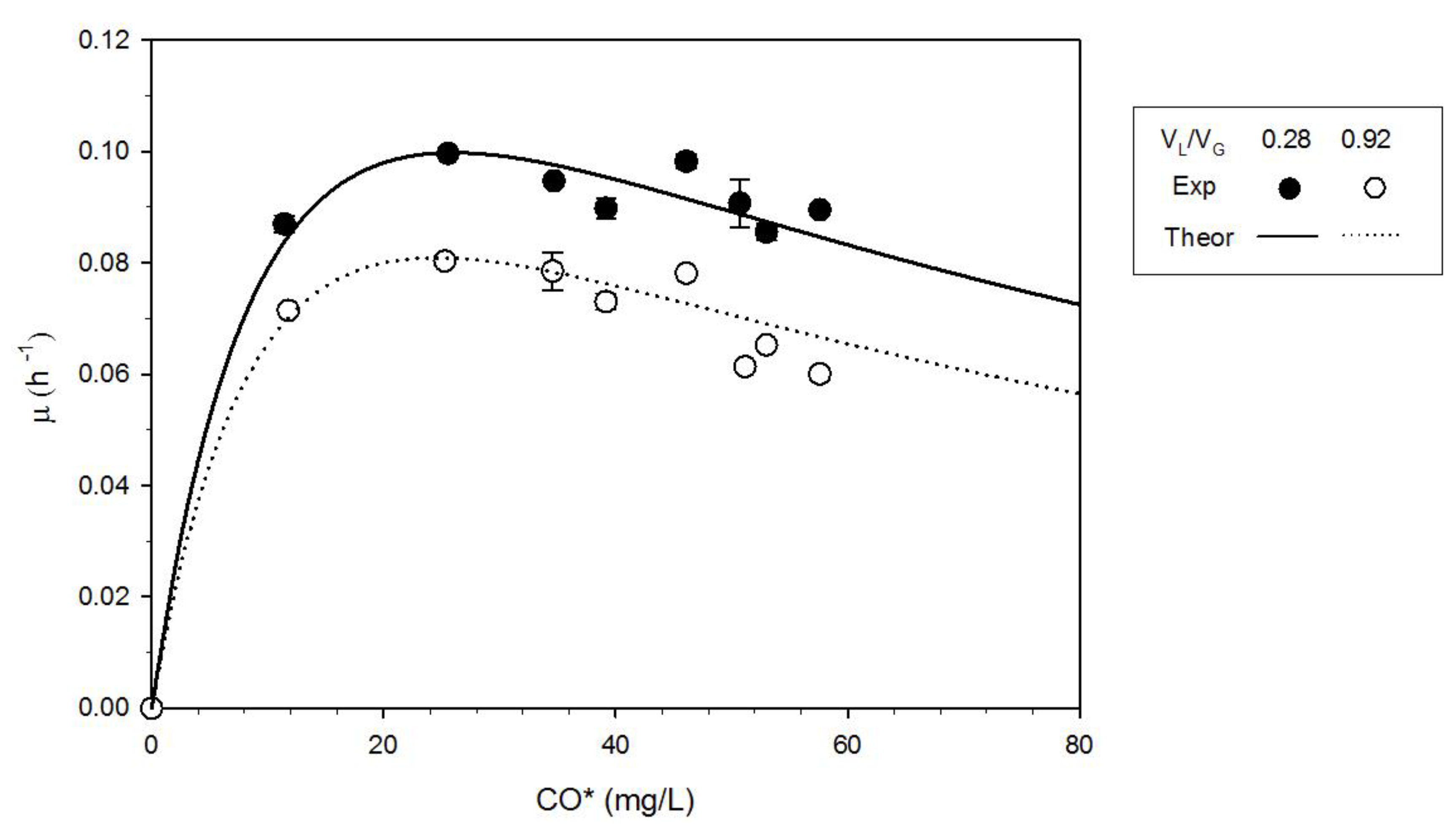
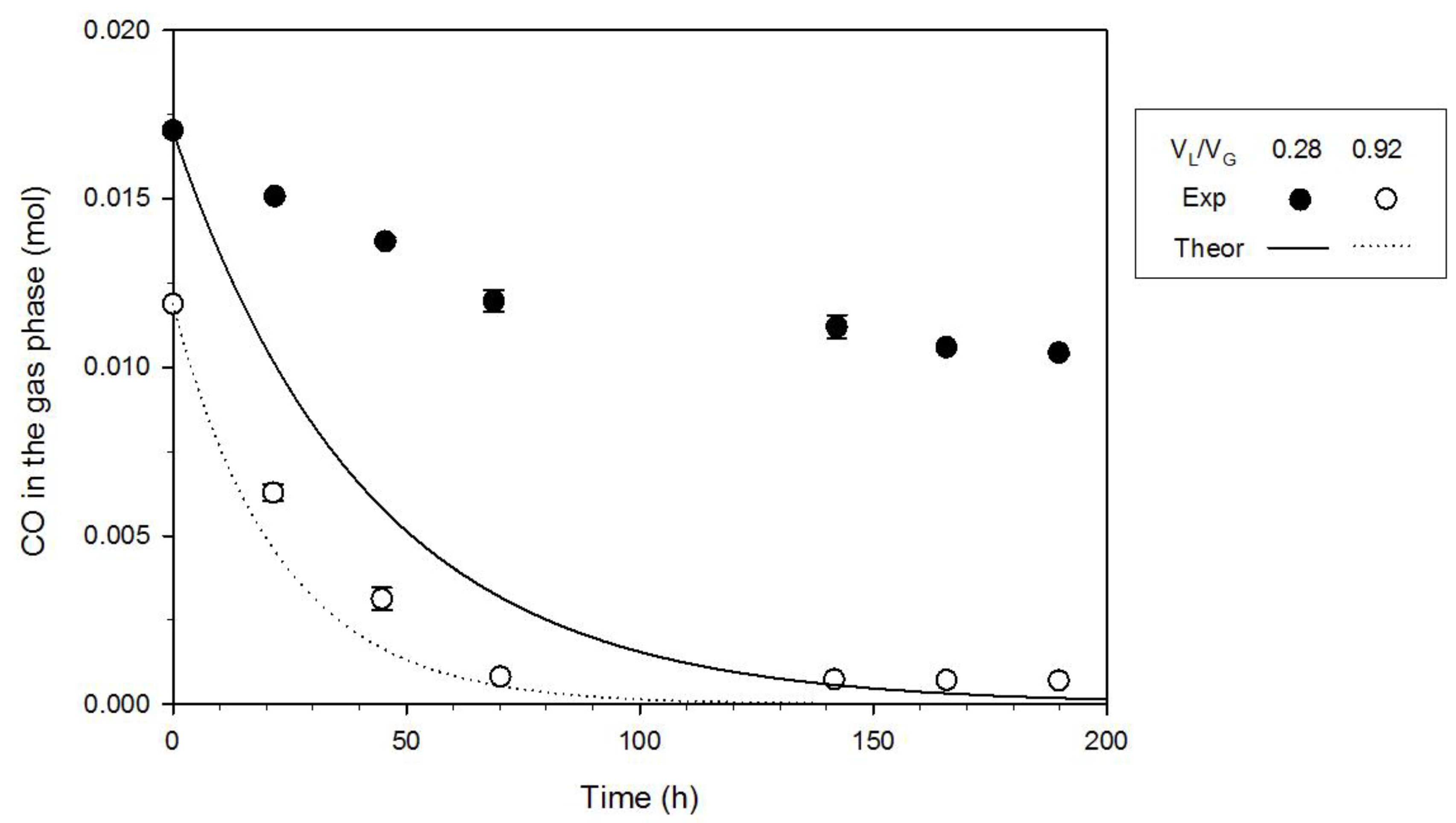
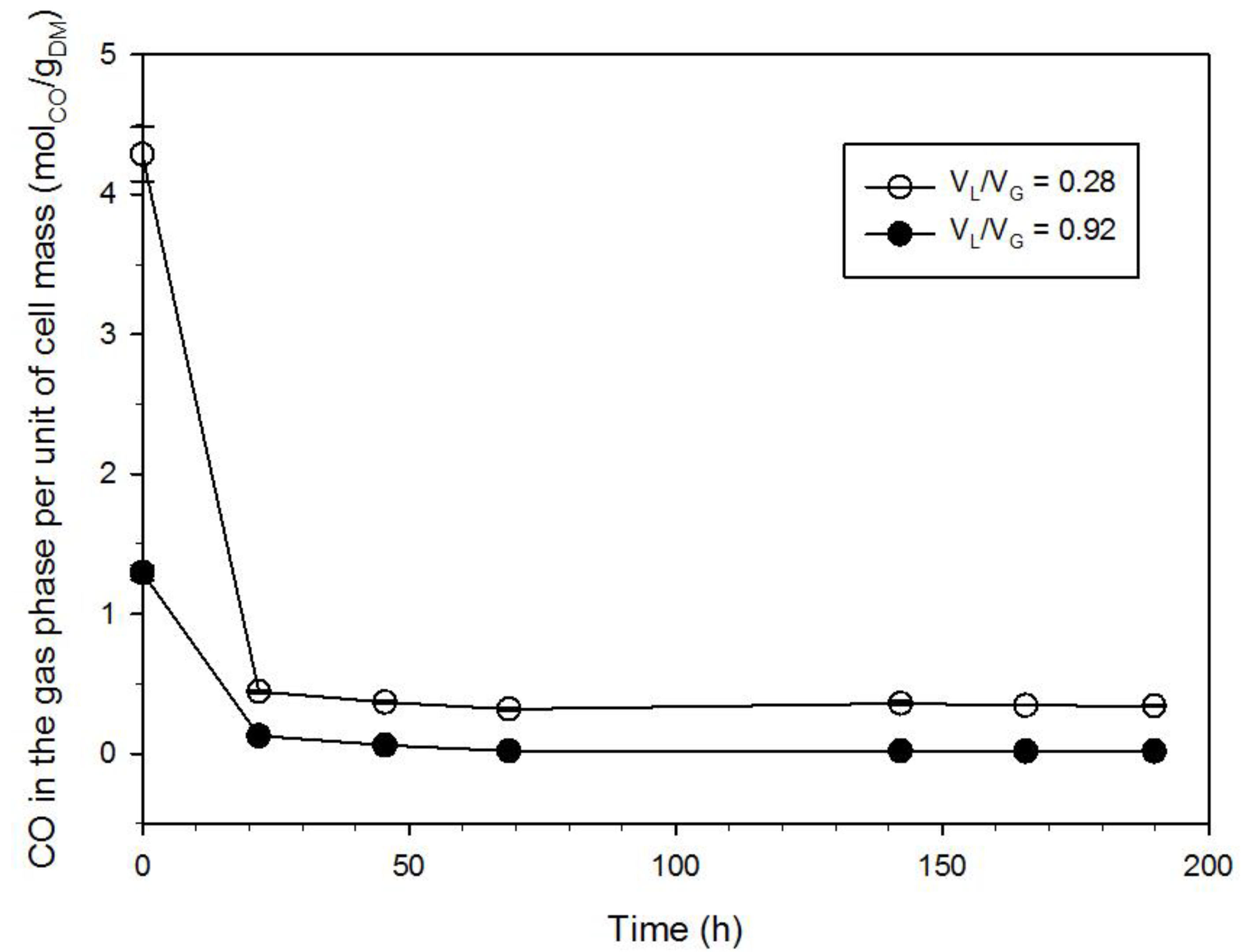
3.3. Effect of the Gas-to-Liquid Volume Ratio
4. Conclusions
- PCO affected microorganism growth kinetics; indeed, C. carboxidivorans growth rate was characterized by CO inhibition within the investigated range of CO concentration, and the optimal PCO was 1.1 atm (corresponding to a dissolved CO concentration of about 25 mg/L) for both the VL/VG used.
- Growth differences were observed when the gas-to-liquid volume ratio was changed; the mass-transport phenomena did not control CO uptake for VL/VG = 0.28; on the contrary, the experimental CO depletion rate was about equal to the transport rate in the case of VL/VG = 0.92. Therefore, the characterization of kinetics and fermentation stoichiometry carried out at VL/VG = 0.28 was more reliable than those carried out at VL/VG = 0.92.
Author Contributions
Funding
Conflicts of Interest
Nomenclature
| AA | Acetic acid concentration (mg/L) |
| B | butanol concentration (mg/L); |
| BA | Butyric acid concentration (mg/L); |
| CO | concentration of CO in the liquid phase (mg/L); |
| CO * | liquid concentration of CO under equilibrium conditions with the gas phase (mg/L) |
| E | Ethanol concentration (mg/L) |
| HA | Hexanoic acid concentration (mg/L) |
| HCO | Henry constant referred to CO (atm *L/mol) |
| kLaCO | volumetric mass transport coefficient for CO (h−1) |
| KM, KI | parameters of the Haldane model (mg/L) |
| nCO | CO mole number (mol) |
| PCO | CO partial pressure (atm) |
| R | universal gas constant (L *atm/K/mol) |
| T | Fermentation temperature (K) |
| VG | Gas volume in serum bottles (mL) |
| VL | Liquid volume in serum bottles (mL; |
| X | cell concentration (gDM/L); |
| Yi/CO | CO-to-product “i” yield coefficient (g/g); |
| μ | specific cell growth rate (h−1) |
| μmax | maximum specific cell growth rate (h−1) |
| ξCO | CO conversion degree (-) |
References
- Gowen, C.M.; Fong, S.S. Applications of systems biology towards microbial fuel production. Trends Microbiol. 2011, 10, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Abdehagh, N.; Tezel, F.H.; Thibault, J. Separation techniques in butanol production: Challenges and developments. Biomass Bioenergy 2014, 60, 222–246. [Google Scholar] [CrossRef]
- Fernández-Naveira, Á.; Abubackar, H.N.; Veiga, M.C.; Kennes, C. Efficient butanol-ethanol (B-E) production from carbon monoxide fermentation by Clostridium carboxidivorans. Appl. Microbiol. Biotechnol. 2016, 100, 3361–3370. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Najafpour, G.D.; Younesi, H.; Lahijani, P.; Uzir, M.H.; Mohamed, A.R. Bioconversion of synthesis gas to second generation biofuels: A review. Renew. Sustain. Energy Rev. 2011, 15, 4255–4257. [Google Scholar] [CrossRef]
- Fernandez-Naveira, A.; Veiga, M.C.; Kennes, C. Production of higher alcohols through anaerobic H–B–E fermentation of syngas or waste gas. Chem. Technol. Biotechnol. 2017, 92, 712–713. [Google Scholar] [CrossRef]
- Fernandez-Naveira, A.; Abubackar, H.N.; Veiga, M.C.; Kennes, C. Production of chemicals from C1 gases (CO, CO2) by Clostridium carboxidivorans. World J. Microbiol. Biotechnol. 2017, 33, 43. [Google Scholar] [CrossRef]
- Stoll, K.I.; Herbig, S.; Sauer, J.; Neumann, A.; Oswald, F. Fermentation of H2 and CO2 with Clostridium ljungdahlii at elevated process pressure—first experimental results. Chem. Eng. Trans. 2018, 64, 2–7. [Google Scholar]
- Jin, Y.; Guo, L.; Veiga, M.C.; Kennes, C. Optimization ofthe treatment ofcarbonmonoxide-polluted air in biofilters. Chemosphere 2009, 74, 332–337. [Google Scholar] [CrossRef] [Green Version]
- Bredwell, M.D.; Worden, R.M. Mass-transfer properties of microbubbles. 1. Experimental studies. Biotechnol Prog 1998, 14, 31–38. [Google Scholar] [CrossRef]
- Raganati, F.; Procentese, A.; Olivieri, G.; Russo, M.E.; Salatino, P.; Marzocchella, A. Biobutanol production from hexose and pentose sugars. Chem. Eng. Trans. 2014, 38, 193–198. [Google Scholar]
- Procentese, A.; Raganati, F.; Olivieri, G.; Russo, M.E.; Salatino, P.; Marzocchella, A. Continuous lactose fermentation by Clostridium acetobutylicum—Assessment of solventogenic kinetics. Bioresour. Technol. 2015, 180, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Liou, J.S.C.; Balkwill, D.L.; Drake, G.R.; Tanner, R.S. Clostridium carboxidivorans sp. nov., a solvent-producing clostridium isolated from an agricultural settling lagoon, and reclassification of the acetogen Clostridium scatologenesstrain SL1 as Clostridium drakei sp. nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 2085–2091. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Jiaqiang, E.; Yang, J.W.; Hui, A.; Cai, H. Development of a skeletal mechanism for biodiesel blend surrogates with varying fatty acid methyl esters proportion. Appl. Energy 2016, 162, 278–288. [Google Scholar] [CrossRef]
- Phillips, J.R.; Atiyeh, H.K.; Tanner, R.S.; Torres, J.R.; Saxena, J.; Wilkins, M.R.; Huhnke, R.L. Butanol and hexanol production in Clostridium carboxidivorans syngas fermentation: Medium development and culture techniques. Bioresour. Technol. 2015, 190, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Abubackar, H.N.; Veiga, M.C.; Kennes, C. Carbon monoxide fermentation to ethanol by Clostridium autoethanogenum in a bioreactor with no accumulation of acetic acid. Bioresour. Technol. 2015, 186, 122–127. [Google Scholar] [CrossRef] [Green Version]
- Rittmann, B.E.; McCarty, P.L. Environmental Technology: Principles and Applications; McGraw-Hill: New York, NY, USA, 2005. [Google Scholar]
- Ragsdale, S.W. Life with carbon monoxide. Crit. Rev. Biochem. Mol. Biol. 2004, 39, 165–195. [Google Scholar] [CrossRef]
- Yasin, M.; Jeong, Y.; Park, S.; Jeong, J.; Lee, E.Y.; Lovitt, R.W.; Kim, B.H.; Lee, J.; Chang, I.S. Microbial synthesis gas utilization and ways to resolve kinetic and mass-transfer limitations. Bioresour. Technol. 2015, 177, 361–374. [Google Scholar] [CrossRef]
- Hurst, K.M.; Lewis, R.S. Carbon monoxide partial pressure effects on the metabolic process of syngas fermentation. Biochem. Eng. J. 2010, 48, 159–165. [Google Scholar] [CrossRef]
- Fernandez-Naveira, A.; Abubackar, H.N.; Veiga, M.C.; Kennes, C. Carbon monoxide bioconversion to butanol-ethanol by Clostridium carboxidivorans: Kinetics and toxicity of alcohols. Appl. Microbiol. Biotechnol. 2016, 100, 4231–4240. [Google Scholar] [CrossRef]
- Younesi, H.; Najafpour, G.; Mohamed, A.R. Ethanol and acetate production from synthesis gas via fermentation processes using anaerobic bacterium, Clostridium ljungdahlii. Biochem. Eng. J. 2005, 27, 110–119. [Google Scholar] [CrossRef]
- Mohammadi, M.; Mohamed, A.R.; Najafpour, G.D.; Younesi, H.; Uzir, M.H. Kinetic studies on fermentative production of biofuel from synthesis gas using Clostridium ljungdahlii. Sci. World J. 2014, 2014, 910590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frankman, A.W. Redox, Pressure and Mass Transfer Effects on Syngas Fermentation. Master’s Thesis, Brigham Young University, Provo, UT, USA, 2009. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCO atm | X gDM/L | AA mg/L | BA mg/L | HA mg/L | E mg/L | B mg/L | YE/CO g/g | ξCO% | μ h−1 |
|---|---|---|---|---|---|---|---|---|---|
| 0.5 | 0.67 ± 1.2 × 10−2 | 1280 ± 20 | 325 ± 6 | 145 ± 2 | 130 ± 2 | 70 ± 1 | 0.063 ± 1.2 × 10−3 | 97.9 ± 0.7 | 0.088 ± 1.5 × 10−3 |
| 1.0 | 0.61 ± 1.1 × 10−2 | 1740 ± 25 | 320 ± 4 | 140 ± 1 | 220 ± 4 | 110 ± 2 | 0.049 ± 9 × 10−4 | 94.2 ± 0.5 | 0.100 ± 5.5 × 10−4 |
| 1.5 | 0.68 ± 1.3 × 10−2 | 1550 ± 15 | 345 ± 5 | 250 ± 4 | 315 ± 5 | 120 ± 2 | 0.097 ± 1.7 × 10−3 | 49.5 ± 0.8 | 0.094 ± 7.2 × 10−4 |
| 1.7 | 0.68 ± 1.4 × 10−2 | 1620 ± 30 | 380 ± 4 | 210 ± 1 | 400 ± 7 | 130 ± 1 | 0.094 ± 1.5 × 10−3 | 57.0 ± 0.6 | 0.092 ± 1.8 × 10−3 |
| 2.0 | 0.68 ± 1.2 × 10−2 | 1200 ± 23 | 390 ± 5 | 285 ± 3 | 380 ± 6 | 100 ± 2 | 0.11 ± 2 × 10−3 | 39.0 ± 0.7 | 0.099 ± 1.2 × 10−3 |
| 2.2 | 0.73 ± 1.4 × 10−2 | 1600 ± 20 | 405 ± 6 | 230 ± 4 | 230 ± 4 | 105 ± 2 | 0.061 ± 1.1 × 10−3 | 44.1 ± 0.8 | 0.086 ± 4.2 × 10−3 |
| 2.3 | 0.53 ± 1 × 10−2 | 970 ± 12 | 390 ± 6 | 250 ± 3 | 180 ± 3 | 110 ± 2 | 0.047 ± 9 × 10−4 | 29.3 ± 0.5 | 0.086 ± 1.5 × 10−3 |
| 2.5 | 0.62 ± 1.1 × 10−2 | 950 ± 10 | 380 ± 4 | 290 ± 2 | 150 ± 2 | 105 ± 1 | 0.043 ± 8 × 10−4 | 31.7 ± 0.6 | 0.089 ± 9 × 10−4 |
| PCO atm | X gDM/L | AA mg/L | BA mg/L | HA mg/L | E mg/L | B mg/L | YE/CO g/g | ξCO% | μ h−1 |
|---|---|---|---|---|---|---|---|---|---|
| 0.5 | 0.38 ± 7 × 10−3 | 715 ± 10 | 85 ± 1 | 70 ± 1 | 120 ± 2 | 25 ± 0.5 | 0.17 ± 3 × 10−3 | 100 ± 0 | 0.072 ± 3 × 10−4 |
| 1.0 | 0.34 ± 6 × 10−3 | 800 ± 15 | 100 ± 2 | 60 ± 1.1 | 115 ± 2 | 25 ± 0.4 | 0.076 ± 1.5 × 10−3 | 99.9 ± 0.1 | 0.081 ± 2 × 10−4 |
| 1.5 | 0.40 ± 7 × 10−3 | 800 ± 10 | 110 ± 2 | 80 ± 1.5 | 120 ± 2 | 35 ± 0.3 | 0.059 ± 1.1 × 10−3 | 98.4 ± 0.5 | 0.075 ± 3 × 10−3 |
| 1.7 | 0.35 ± 7 × 10−3 | 1110 ± 20 | 124 ± 2 | 80 ± 1.4 | 130 ± 2 | 45 ± 0.7 | 0.056 ± 1.1 × 10−3 | 97.2 ± 0.4 | 0.072 ± 1.3 × 10−3 |
| 2.0 | 0.42 ± 8 × 10−3 | 1580 ± 25 | 176 ± 3 | 85 ± 1.2 | 225 ± 3 | 70 ± 0.5 | 0.067 ± 1.3 × 10−3 | 94.2 ± 0.5 | 0.086 ± 8 × 10−4 |
| 2.2 | 0.47 ± 8 × 10−3 | 1870 ± 25 | 230 ± 4 | 90 ± 1.4 | 190 ± 2 | 75 ± 1 | 0.062 ± 1.2 × 10−3 | 100 ± 0 | 0.062 ± 7 × 10−4 |
| 2.3 | 0.39 ± 7 × 10−3 | 1500 ± 20 | 130 ± 2 | 80 ± 1.3 | 90 ± 2 | 60 ± 1.2 | 0.031 ± 6 × 10−4 | 92.6 ± 0.7 | 0.065 ± 5 × 10−4 |
| 2.5 | 0.38 ± 7 × 10−3 | 1520 ± 10 | 100 ± 1 | 60 ± 1 | 90 ± 1 | 80 ± 1.1 | 0.030 ± 6 × 10−4 | 86.8 ± 0.8 | 0.060 ± 1.1 × 10−3 |
| VL/VG | μmax h−1 | KM mg/L | KI mg/L |
|---|---|---|---|
| 0.28 | 0.22 | 15 | 45 |
| 0.92 | 0.18 | 15 | 40 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lanzillo, F.; Ruggiero, G.; Raganati, F.; Russo, M.E.; Marzocchella, A. Batch Syngas Fermentation by Clostridium carboxidivorans for Production of Acids and Alcohols. Processes 2020, 8, 1075. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8091075
Lanzillo F, Ruggiero G, Raganati F, Russo ME, Marzocchella A. Batch Syngas Fermentation by Clostridium carboxidivorans for Production of Acids and Alcohols. Processes. 2020; 8(9):1075. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8091075
Chicago/Turabian StyleLanzillo, Fabiana, Giacomo Ruggiero, Francesca Raganati, Maria Elena Russo, and Antonio Marzocchella. 2020. "Batch Syngas Fermentation by Clostridium carboxidivorans for Production of Acids and Alcohols" Processes 8, no. 9: 1075. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8091075