
Production of Omega-3 Oil by Aurantiochytrium mangrovei Using Spent Osmotic Solution from Candied Fruit Industry as Sole Organic Carbon Source

 ,
,  ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Organism and Cultivation
2.2. Spent Osmotic Solution Samples
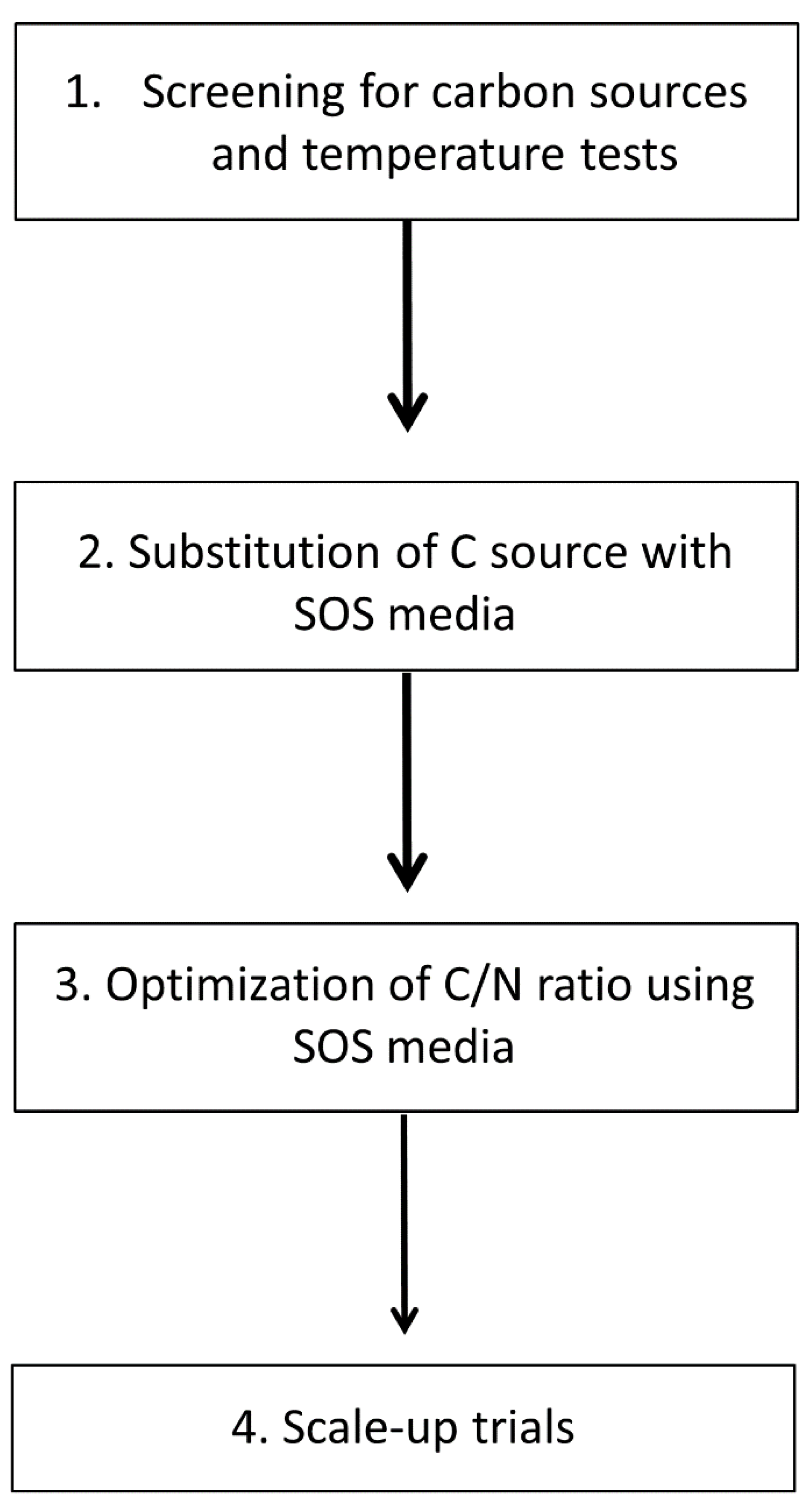
2.3. Experimental Design
- (1)
- Screening test to evaluate the growth of A. mangrovei RCC893 using different temperatures and different organic carbon sources in order to define the growth performances and the best operating parameters;
- (2)
- Substitution of C source in the standard media using SOS as an organic carbon source;
- (3)
- Optimization of biomass and DHA production through response surface methodologies (RSM) with different C/N ratios;
- (4)
- Scale-up trial in airlift reactor to evaluate the scalability of the new SOS medium.
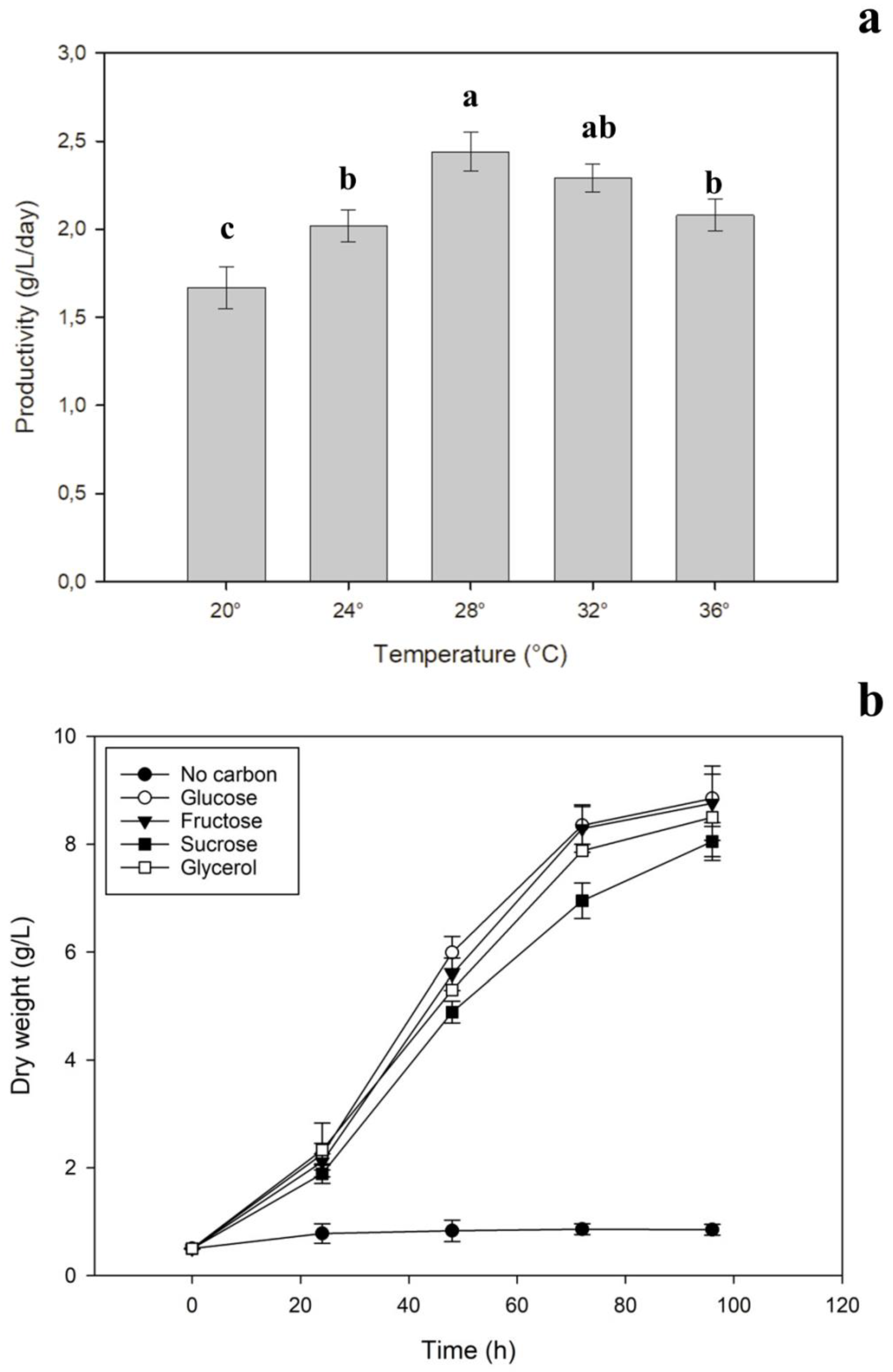
2.3.1. Best Growth Temperature and Organic Carbon Source
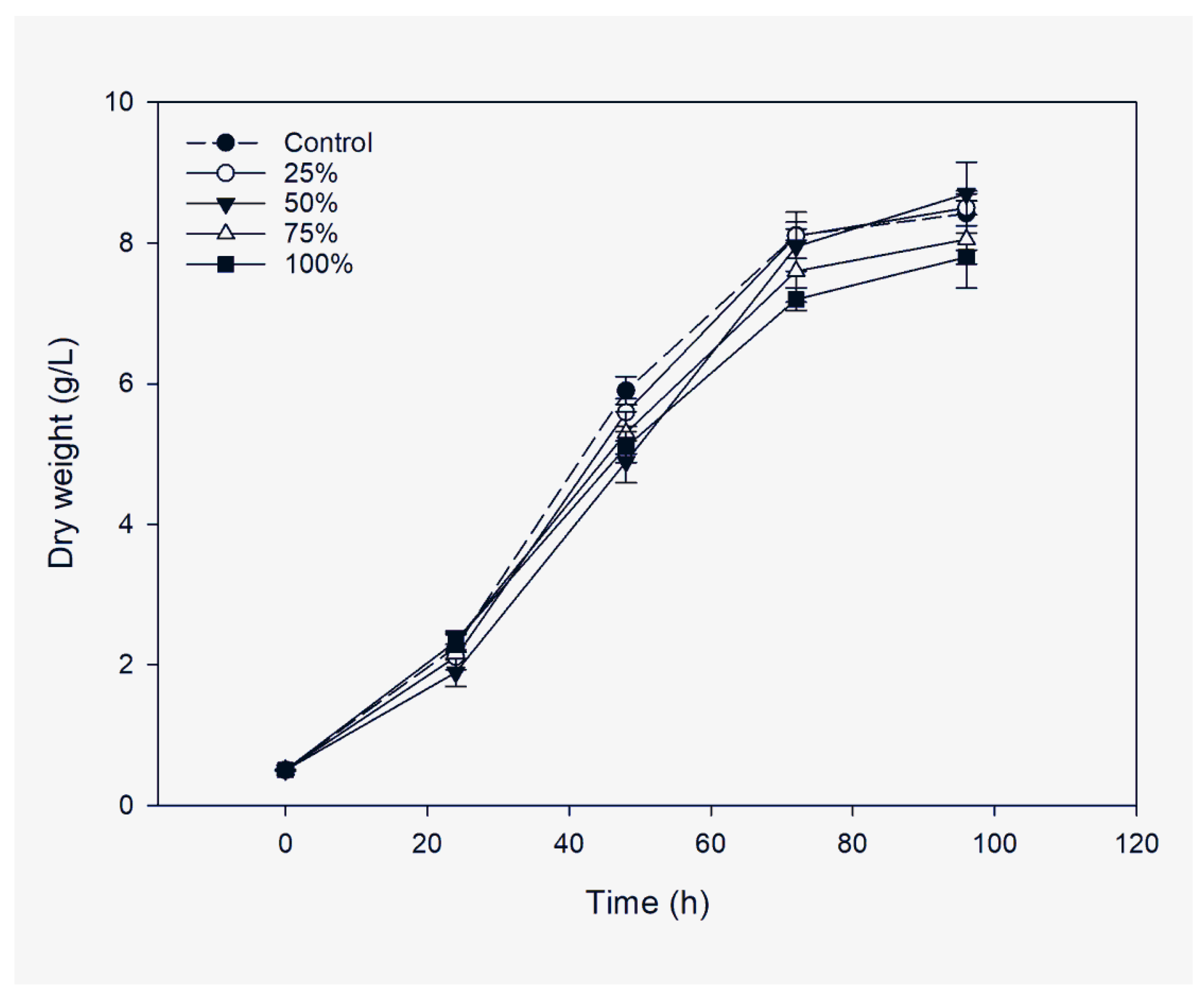
2.3.2. Substitution of Carbon Source with Spent Osmotic Solution
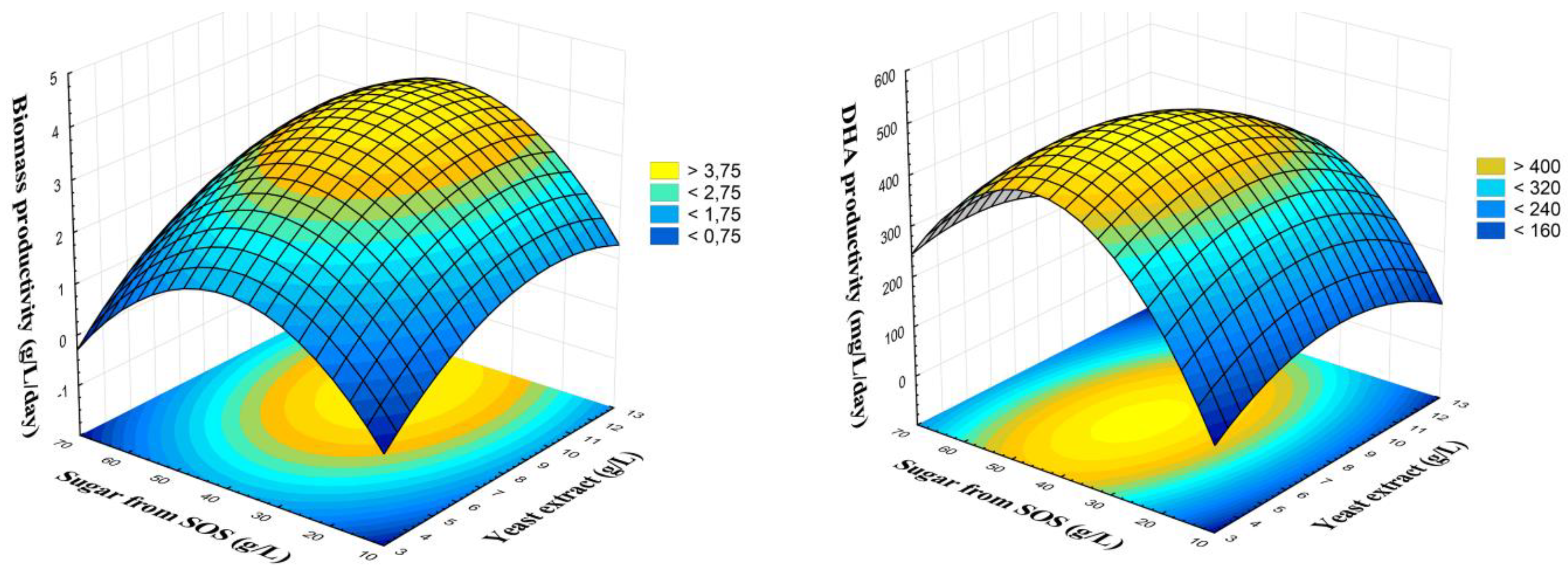
2.3.3. Response Surface Analysis and Formulation of Optimized Media
2.4. Analytical Methods
2.4.1. Measurement of Dry Cell Weight and Residual Nutrients
2.4.2. Fatty Acid Methyl Esters (FAMEs)
2.5. Statistical Analysis
3. Results and Discussions
3.1. Optimal Standard Conditions
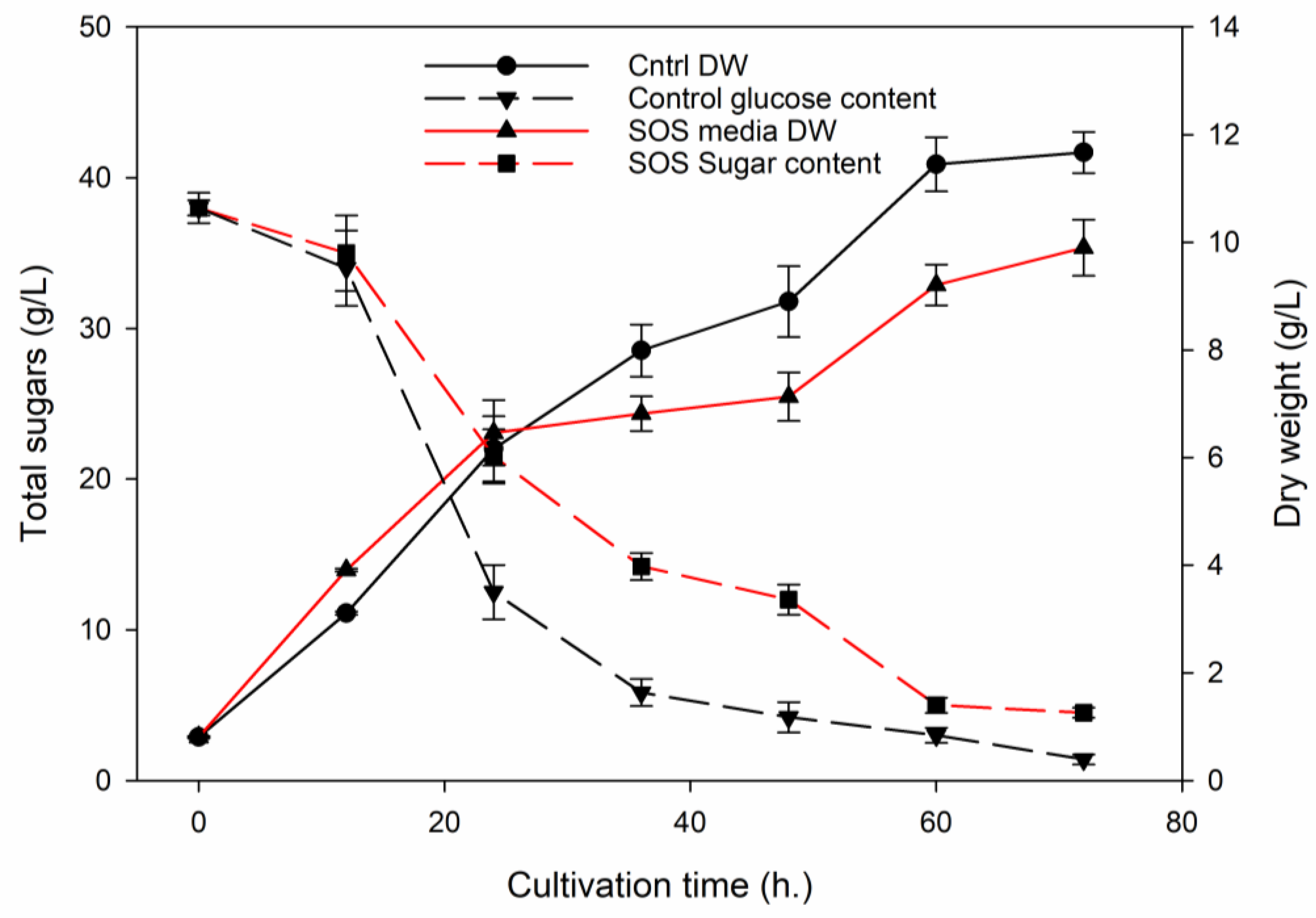
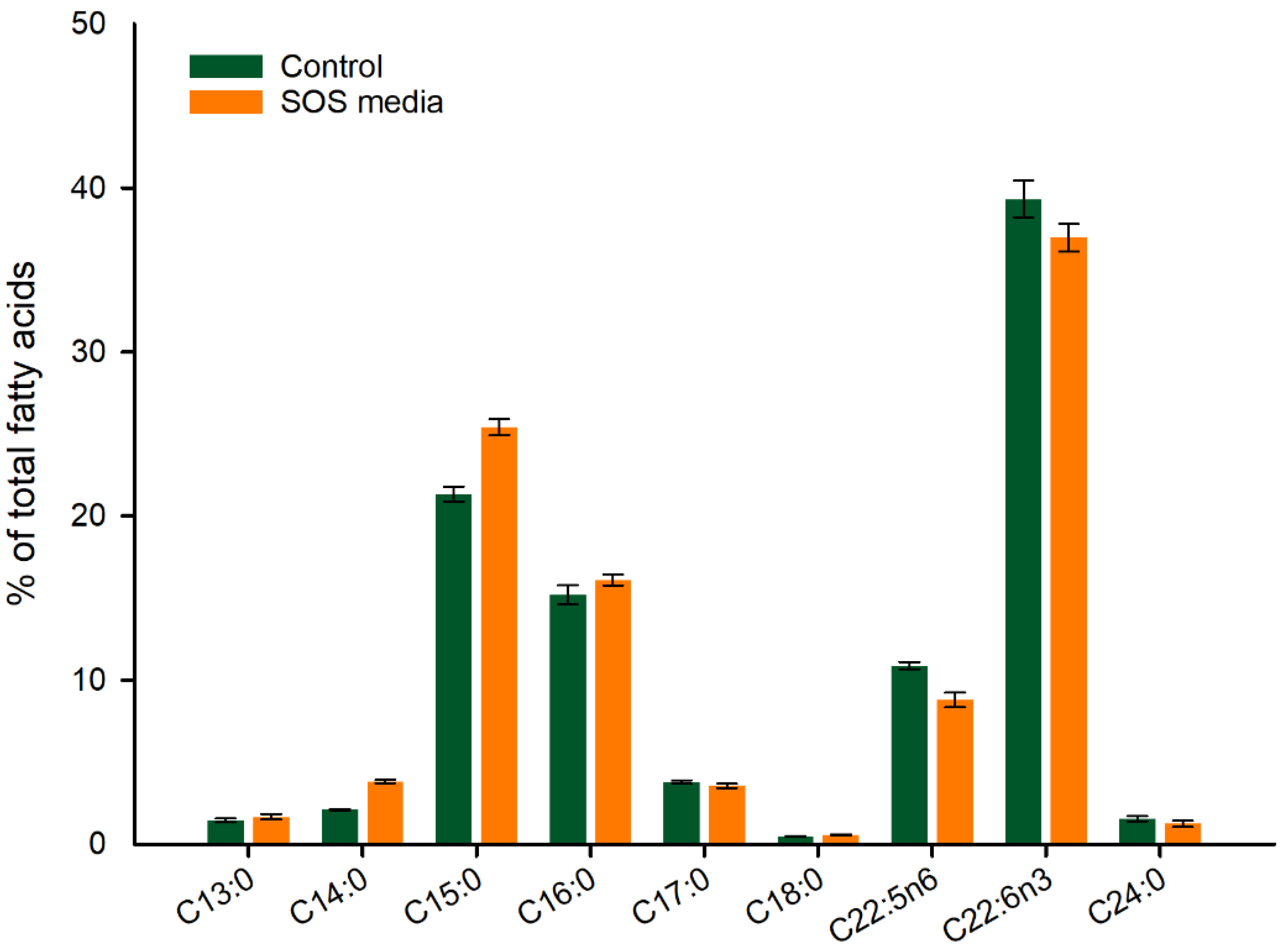
3.2. Utilization of SOS as a Carbon Source
3.3. Response Surface Results for Biomass and DHA Productivity
3.4. Scale-up Trial
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Birch, E.E.; Garfield, S.; Castañeda, Y.; Hughbanks-Wheaton, D.; Uauy, R.; Hoffman, D. Visual acuity and cognitive outcomes at 4 years of age in a double-blind, randomized trial of long-chain polyunsaturated fatty acid-supplemented infant formula. Early Hum. Dev. 2007, 83, 279–284. [Google Scholar] [CrossRef]
- Zeng, Y.; Ji, X.J.; Lian, M.; Ren, L.J.; Jin, L.J.; Ouyang, P.K.; Huang, H. Development of a temperature shift strategy for efficient docosahexaenoic acid production by a marine fungoid protist, Schizochytrium sp. HX-308. Appl. Biochem. Biotechnol. 2011, 164, 249–255. [Google Scholar] [CrossRef]
- Chen, W.; Zhou, P.; Zhu, Y.; Xie, C.; Ma, L.; Wang, X.; Bao, Z.; Yu, L. Improvement in the docosahexaenoic acid production of Schizochytrium sp. S056 by replacement of sea salt. Bioprocess Biosyst. Eng. 2016, 39, 315–321. [Google Scholar] [CrossRef]
- Graham, I.A.; Larson, T.; Napier, J.A. Rational metabolic engineering of transgenic plants for biosynthesis of omega-3 polyunsaturates. Curr. Opin. Biotechnol. 2007, 18, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, T.; Yokochi, T.; Higashihara, T.; Tanaka, S.; Yaguchi, T.; Honda, D. Production of docosahexaenoic and docosapentaenoic acids by Schizochytrium sp. isolated from yap islands. JAOCS J. Am. Oil Chem. Soc. 1996, 73, 1421–1426. [Google Scholar] [CrossRef]
- Lewis, T.E.; Nichols, P.D.; McMeekin, T.A. The biotechnological potential of thraustochytrids. Mar. Biotechnol. 1999, 1, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Song, X.; Feng, Y.; Li, W.; Cui, Q. Isolation and characterization of Aurantiochytrium species: High docosahexaenoic acid (DHA) production by the newly isolated microalga, Aurantiochytrium sp. SD116. J. Oleo Sci. 2013, 62, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barclay, W.; Weaver, C.; Metz, J.; Hansen, J. Development of a Docosahexaenoic Acid Production Technology Using Schizochytrium: Historical Perspective and Update. In Single Cell Oils: Microbial and Algal Oils, 2nd ed.; Elsevier BV: Amsterdam, The Netherlands, 2010; ISBN 9781630670078. [Google Scholar]
- Nazir, Y.; Shuib, S.; Kalil, M.S.; Song, Y.; Hamid, A.A. Optimization of Culture Conditions for Enhanced Growth, Lipid and Docosahexaenoic Acid (DHA) Production of Aurantiochytrium SW1 by Response Surface Methodology. Sci. Rep. 2018, 8, 8909. [Google Scholar] [CrossRef]
- Humhal, T.; Kastanek, P.; Jezkova, Z.; Cadkova, A.; Kohoutkova, J.; Branyik, T. Use of saline waste water from demineralization of cheese whey for cultivation of Schizochytrium limacinum PA-968 and Japonochytrium marinum AN-4. Bioprocess Biosyst. Eng. 2017, 40, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Russo, G.L.; Langellotti, A.L.; Oliviero, M.; Sacchi, R.; Masi, P. Sustainable production of food grade omega-3 oil using aquatic protists: Reliability and future horizons. New Biotechnol. 2021, 62, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Aachary, A.A.; Prapulla, S.G. Value addition to spent osmotic sugar solution (SOS) by enzymatic conversion to fructooligosaccharides (FOS), a low calorie prebiotic. Innov. Food Sci. Emerg. Technol. 2009, 10, 284–288. [Google Scholar] [CrossRef]
- Massa, M.; Buono, S.; Langellotti, A.L.; Martello, A.; Russo, G.L.; Troise, D.A.; Sacchi, R.; Vitaglione, P.; Fogliano, V. Biochemical composition and in vitro digestibility of Galdieria sulphuraria grown on spent cherry-brine liquid. New Biotechnol. 2019, 53, 9–15. [Google Scholar] [CrossRef]
- Ryu, B.-G.; Kim, K.; Kim, J.; Han, J.-I.; Yang, J.-W. Use of organic waste from the brewery industry for high-density cultivation of the docosahexaenoic acid-rich microalga, Aurantiochytrium sp. KRS101. Bioresour. Technol. 2013, 129, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.-J.; Yu, Z.-Q.; Liu, Y.-L.; Sun, J.; Zheng, J.-Y.; Wang, Z. Utilization of High-Fructose Corn Syrup for Biomass Production Containing High Levels of Docosahexaenoic Acid by a Newly Isolated Aurantiochytrium sp. YLH70. Appl. Biochem. Biotechnol. 2015, 177, 1229–1240. [Google Scholar] [CrossRef]
- Lee Chang, K.J.; Dumsday, G.; Nichols, P.D.; Dunstan, G.A.; Blackburn, S.I.; Koutoulis, A. High cell density cultivation of a novel Aurantiochytrium sp. strain TC 20 in a fed-batch system using glycerol to produce feedstock for biodiesel and omega-3 oils. Appl. Microbiol. Biotechnol. 2013, 97, 6907–6918. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Morrison, W.R.; Smith, L.M. Preparation of fatty acid methyl esters and dimethylacetals from lipids with boron fluoride–methanol. J. Lipid Res. 1964, 5, 600–608. [Google Scholar] [CrossRef]
- Nakazawa, A.; Matsuura, H.; Kose, R.; Kato, S.; Honda, D.; Inouye, I.; Kaya, K.; Watanabe, M.M. Optimization of culture conditions of the thraustochytrid Aurantiochytrium sp. strain 18W-13a for squalene production. Bioresour. Technol. 2012, 109, 287–291. [Google Scholar] [CrossRef]
- Taoka, Y.; Nagano, N.; Okita, Y.; Izumida, H.; Sugimoto, S.; Hayashi, M. Influences of culture temperature on the growth, lipid content and fatty acid composition of Aurantiochytrium sp. strain mh0186. Mar. Biotechnol. 2009, 11, 368–374. [Google Scholar] [CrossRef]
- Mariam, I.; Kareya, M.S.; Nesamma, A.A.; Jutur, P.P. Delineating metabolomic changes in native isolate Aurantiochytrium for production of docosahexaenoic acid in presence of varying carbon substrates. Algal Res. 2021, 55, 102285. [Google Scholar] [CrossRef]
- Pahlavanyali, M.; Jalili, H.; Noroozi, M.; Moradi, Y.; Hallajisani, A. The effect of temperature and different carbon and nitrogen sources on the growth and fatty acid profile of a newly isolated microorganism Aurantiochytrium sp. strain SHY. Iran. J. Fish. Sci. 2020, 19, 3112–3126. [Google Scholar] [CrossRef]
- Hong, W.K.; Rairakhwada, D.; Seo, P.S.; Park, S.Y.; Hur, B.K.; Kim, C.H.; Seo, J.W. Production of lipids containing high levels of docosahexaenoic acid by a newly isolated microalga, Aurantiochytrium sp. KRS101. Appl. Biochem. Biotechnol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.-W.; Zhu, S.-Y.; Guo, D.-S.; Ren, L.-J.; Ji, X.-J.; Huang, H.; Gao, Z. Development of a strategy for the production of docosahexaenoic acid by Schizochytrium sp. from cane molasses and algae-residue. Bioresour. Technol. 2019, 271, 118–124. [Google Scholar] [CrossRef]
- Iwasaka, H.; Aki, T.; Adachi, H.; Watanabe, K.; Kawamoto, S.; Ono, K. Utilization of waste syrup for production of polyunsaturated fatty acids and xanthophylls by Aurantiochytrium. J. Oleo Sci. 2013, 62, 729–736. [Google Scholar] [CrossRef] [Green Version]
- Wong, M.K.M.; Tsui, C.K.M.; Au, D.W.T.; Vrijmoed, L.L.P. Docosahexaenoic acid production and ultrastructure of the thraustochytrid Aurantiochytrium mangrovei MP2 under high glucose concentrations. Mycoscience 2008, 49, 266–270. [Google Scholar] [CrossRef]
- Yokochi, T.; Honda, D.; Higashihara, T.; Nakahara, T. Optimization of docosahexaenoic acid production by Schizochytrium limacinum SR21. Appl. Microbiol. Biotechnol. 1998, 49, 72–76. [Google Scholar] [CrossRef]
- Chaung, K.C.; Chu, C.Y.; Su, Y.M.; Chen, Y.M. Effect of culture conditions on growth, lipid content, and fatty acid composition of Aurantiochytrium mangrovei strain BL10. AMB Express 2012, 2, 42. [Google Scholar] [CrossRef] [Green Version]
- Ju, J.H.; Oh, B.R.; Ryu, S.K.; Heo, S.Y.; Kim, S.Y.; Hong, W.K.; Kim, C.H.; Seo, J.W. Production of Lipid Containing High Levels of Docosahexaenoic Acid by Cultivation of Aurantiochytrium sp. KRS101 Using Jerusalem Artichoke Extract. Biotechnol. Bioprocess Eng. 2018, 23, 726–732. [Google Scholar] [CrossRef]
- Park, W.K.; Moon, M.; Shin, S.E.; Cho, J.M.; Suh, W.I.; Chang, Y.K.; Lee, B. Economical DHA (Docosahexaenoic acid) production from Aurantiochytrium sp. KRS101 using orange peel extract and low cost nitrogen sources. Algal Res. 2018, 29, 71–79. [Google Scholar] [CrossRef]
- Liang, Y.; Sarkany, N.; Cui, Y.; Yesuf, J.; Trushenski, J.; Blackburn, J.W. Use of sweet sorghum juice for lipid production by Schizochytrium limacinum SR21. Bioresour. Technol. 2010, 101, 3623–3627. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russo, G.L.; Langellotti, A.L.; Blasco, T.; Oliviero, M.; Sacchi, R.; Masi, P. Production of Omega-3 Oil by Aurantiochytrium mangrovei Using Spent Osmotic Solution from Candied Fruit Industry as Sole Organic Carbon Source. Processes 2021, 9, 1834. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9101834
Russo GL, Langellotti AL, Blasco T, Oliviero M, Sacchi R, Masi P. Production of Omega-3 Oil by Aurantiochytrium mangrovei Using Spent Osmotic Solution from Candied Fruit Industry as Sole Organic Carbon Source. Processes. 2021; 9(10):1834. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9101834
Chicago/Turabian StyleRusso, Giovanni L., Antonio L. Langellotti, Thierry Blasco, Maria Oliviero, Raffaele Sacchi, and Paolo Masi. 2021. "Production of Omega-3 Oil by Aurantiochytrium mangrovei Using Spent Osmotic Solution from Candied Fruit Industry as Sole Organic Carbon Source" Processes 9, no. 10: 1834. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9101834