The Influence of Surface Mycobiota on Sensory Properties of “Istarski pršut” and “Dalmatinski pršut”

 ,
,  , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Dry-Cured Ham Production and Sampling
2.2. Isolation and Traditional Identification of Surface Mycobiota
2.3. Molecular Identification of Surface Mycobiota
2.4. Sensory Evaluation
2.5. Statistical Analysis
3. Results and Discussion
3.1. The Presence of Mycobiota on Dry-Cured Ham Surface
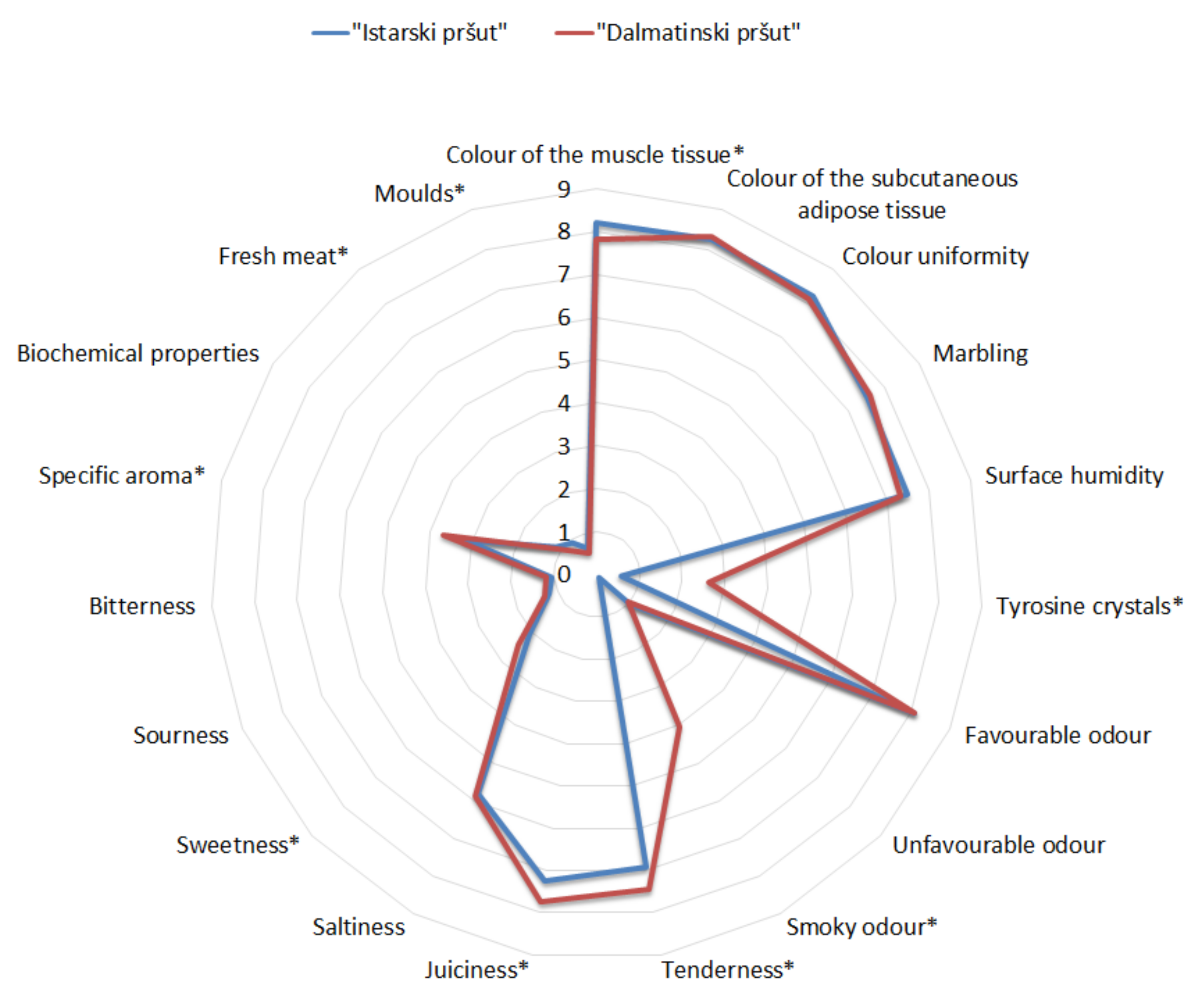
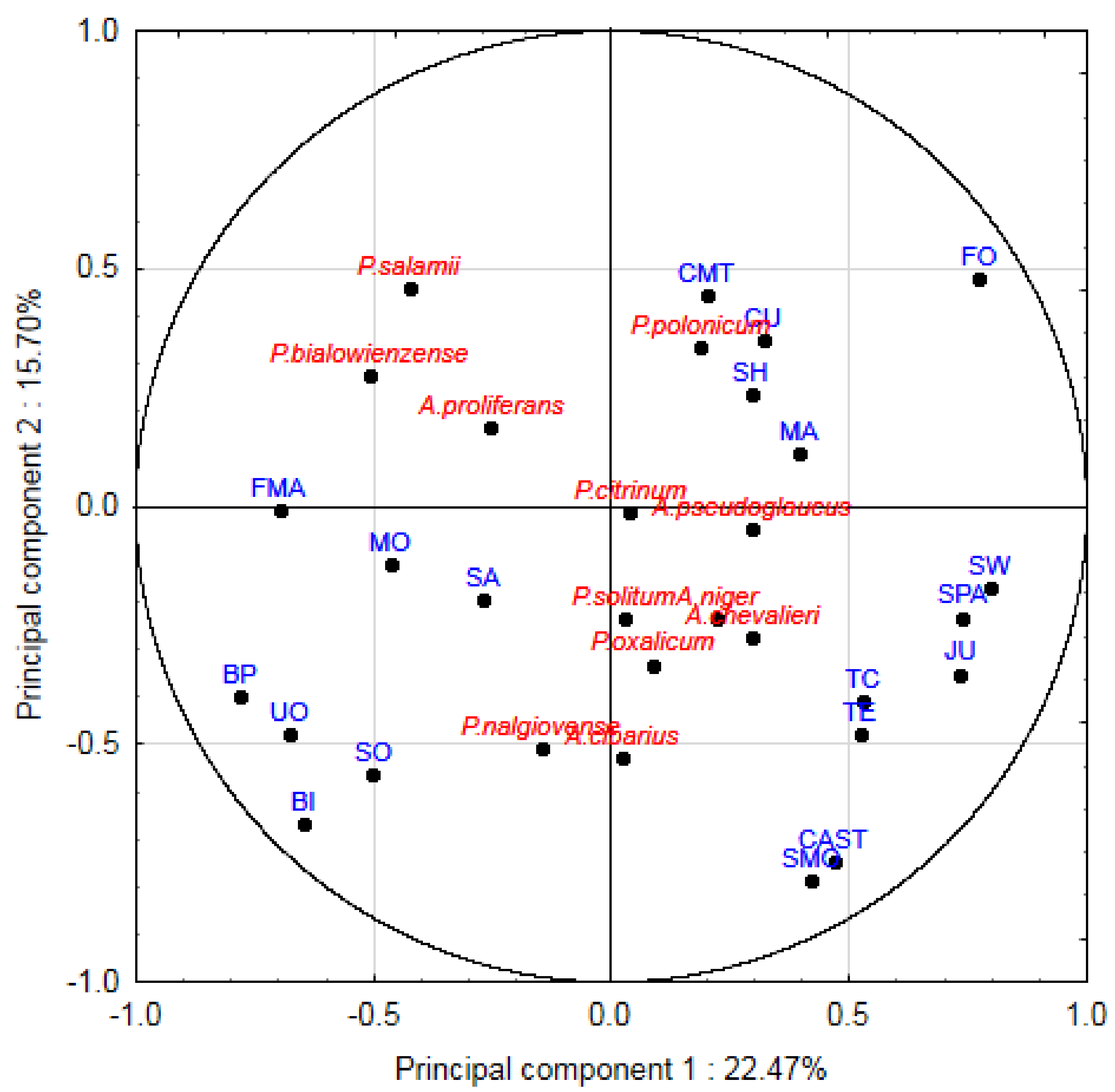
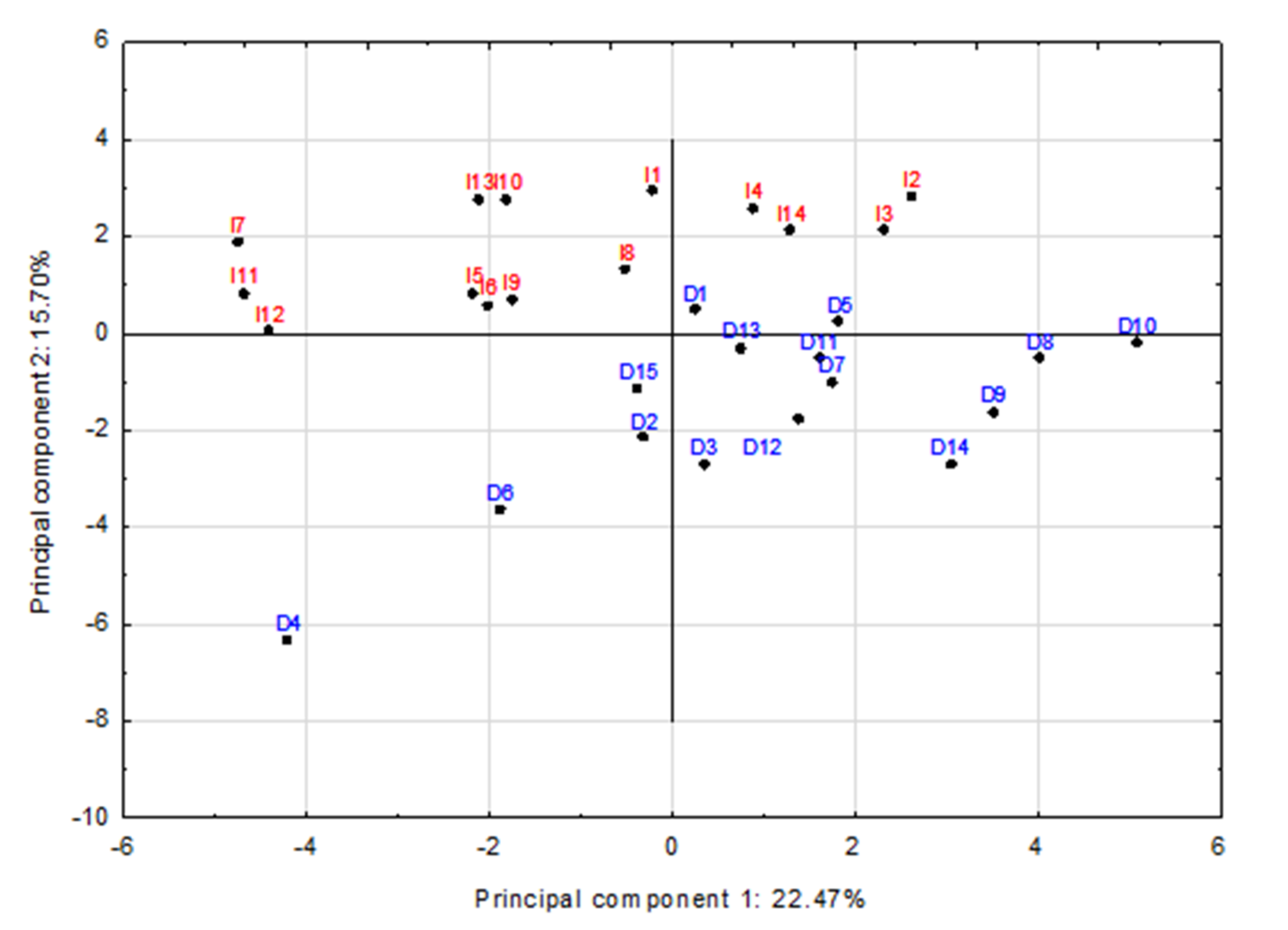
3.2. The Effect of Surface Mycobiota on Sensory Properties
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Benedini, R.; Parolari, G.; Toscani, T.; Virgili, R. Sensory and texture properties of Italian typical dry-cured hams as related to maturation time and salt content. Meat Sci. 2012, 90, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Flores, M. Sensory descriptors for dry-cured meat products. In Sensory Analysis of Foods of Animal Origin; Nolet, L.M., Toldra, F., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 173–192. [Google Scholar]
- García-González, D.L.; Roncales, P.; Cilla, I.; Del Río, S.; Poma, J.P.; Aparicio, R. Interlaboratory evaluation of dry-cured hams (from France and Spain) by assessors from two different nationalities. Meat Sci. 2006, 73, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Petričević, S.; Marušić Radovčić, N.; Lukić, K.; Listeš, E.; Medić, H. Differentiation of dry-cured hams from different processing methods by means of volatile compounds, physico-chemical and sensory analysis. Meat Sci. 2018, 137, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Agriculture. Product Specification “Istarski Pršut”/“Istrski Pršut” for Registration of Protected Designation of Origin. 2014. Available online: https://poljoprivreda.gov.hr/UserDocsImages/arhiva/datastore/filestore/97/Izmijenjena_Specifikacija_proizvoda_Istarski_prsut-Istrski_prsut.pdf (accessed on 4 January 2021).
- Ministry of Agriculture. Product Specification “Dalmatinski Pršut” for Registration of Protected Geographical Indications. 2015. Available online: https://www.prsut-vostane.hr/hr/specifikacija-dalmatinski-prsut.pdf (accessed on 11 January 2021).
- Prado, N.; Sampayo, M.; González, P.; Lombó, F.; Díaz, J. Physicochemical, sensory and microbiological characterization of Asturian Chorizo, a traditional fermented sausage manufactured in Northern Spain. Meat Sci. 2019, 156, 118–124. [Google Scholar] [CrossRef]
- Alapont, C.; Martinez-Culebras, P.V.; Lopez-Mendoza, M.C. Determination of lipolytic and proteolytic activities of mycoflora isolated from dry-cured teruel ham. J. Food Sci. Technol. 2014, 52, 5250–5256. [Google Scholar] [CrossRef] [Green Version]
- Comi, G.; Orlić, S.; Redžepović, S.; Urso, R.; Iacumin, L. Moulds isolated from Istrian dried ham at the pre-ripening and ripening level. Int. J. Food Microbiol. 2004, 96, 29–34. [Google Scholar] [CrossRef]
- Comi, G.; Iacumin, L. Ecology of moulds during the pre-ripening and ripening of San Daniele dry cured ham. Food Res. Int. 2013, 54, 1113–1119. [Google Scholar] [CrossRef]
- Sonjak, S.; Ličen, M.; Frisvald, J.C.; Gunde-Cimerman, N. The mycobiota of three dry-cured meat products from Slovenia. Food Microbiol. 2011, 28, 373–376. [Google Scholar] [CrossRef]
- Zadravec, M.; Vahčić, N.; Brnić, D.; Markov, K.; Frece, J.; Beck, R.; Lešić, T.; Pleadin, J. A study of surface moulds and mycotoxins in Croatian traditional dry-cured meat products. Int. J. Food Microbiol. 2020, 317, 108459. [Google Scholar] [CrossRef]
- Magista, D.; Susca, A.; Ferrara, M.; Logrieco, A.F.; Perrone, G. Penicillium species: Crossroad between quality and safety of cured meat production. Curr. Opin. Food Sci. 2017, 17, 36–40. [Google Scholar] [CrossRef]
- Martıín, A.; Córdoba, J.J.; Aranda, E.; Córdoba, M.G.; Asensio, M.A. Contribution of a selected fungal population to the volatile compounds on dry-cured ham. Int. J. Food Microbiol. 2006, 110, 8–18. [Google Scholar] [CrossRef]
- Marušić, N.; Petrović, M.; Vidaček, S.; Petrak, T.; Medić, H. Characterization of traditional Istrian dry-cured ham by means of physical and chemical analyses and volatile compounds. Meat Sci. 2011, 88, 786–790. [Google Scholar] [CrossRef]
- Martıín, A.; Córdoba, J.J.; Núñez, F.; Benito, M.J.; Asensio, M.A. Contribution of a selected fungal population to proteolysis on dry-cured ham. Int. J. Food Microbiol. 2004, 94, 55–66. [Google Scholar] [CrossRef]
- Garcia, M.; Casas, C.; Toledo, V.; Selgas, M.D. Effect of selected mould strains on the sensory properties of dry fermented sausages. Eur. Food Res. Technol. 2001, 212, 287–291. [Google Scholar] [CrossRef]
- Ockerman, H.W.; Céspedes Sánchez, F.J.; León Crespo, F. Influence of molds on flavor quality of Spanish ham. J. Muscle Foods 2000, 11, 247–259. [Google Scholar] [CrossRef]
- Sunesen, L.O.; Stahnke, L.H. Mould starter cultures for dry sausages-selection, application and effects. Meat Sci. 2003, 65, 935–948. [Google Scholar] [CrossRef]
- Pleadin, J.; Zadravec, M.; Brnić, D.; Perković, I.; Škrivanko, M.; Kovačević, D. Moulds and mycotoxins detected in the regional speciality fermented sausage ’slavonski kulen’ during a 1-year production period. Food Addit. Contam. A 2017, 34, 282–290. [Google Scholar] [CrossRef]
- Sunesen, L.O.; Trihaas, J.; Stahnke, L.H. Volatiles in a sausage surface model-influence of Penicillium nalgiovense, Pediococcus pentosaceus, ascorbate, nitrate and temperature. Meat Sci. 2004, 66, 447–456. [Google Scholar] [CrossRef]
- Jerković, I.; Mastelić, J.; Tartaglia, S. A study of volatile flavour substances in Dalmatian traditional smoked ham: Impact of dry-curing and frying. Food Chem. 2007, 104, 1030–1039. [Google Scholar] [CrossRef]
- Marušić, N.; Vidaček, S.; Janči, T.; Petrak, T.; Medić, H. Determination of volatile compounds and quality parameters of traditional Istrian dry-cured ham. Meat Sci. 2014, 96, 1409–1416. [Google Scholar] [CrossRef]
- Marušić Radovčić, N.; Vidaček, S.; Janči, T.; Medić, H. Characterization of volatile compounds, physico-chemical and sensory characteristics of smoked dry-cured ham. J. Food Sci. Technol. 2016, 53, 4093–4105. [Google Scholar] [CrossRef] [Green Version]
- Pleadin, J.; Lešić, T.; Vahčić, N.; Raspović, I.; Malenica Staver, M.; Krešić, G.; Bogdanović, T.; Kovačević, D. Seasonal variations in fatty acids composition of Istrian and Dalmatian prosciutto. Meso 2015, 5, 449–454. [Google Scholar]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage; Springer: New York, NY, USA, 2009. [Google Scholar]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi, 2nd ed.; Westerdijk Fungal Biodiversity Institute: Utrecht, The Netherlands, 2019. [Google Scholar]
- International Organization for Standardization (ISO). Sensory Analysis-Methodology-Guidelines for Monitoring the Performance of a Quantitative Sensory Panel; ISO Standard 11132:2012; ISO: Geneva, Switzerland, 2012. [Google Scholar]
- Pham, A.J.; Schilling, M.W.; Mikel, W.B.; Williams, J.B.; Martin, J.M.; Coggins, P.C. Relationships between sensory descriptors, consumer acceptability and volatile flavor compounds of American dry-cured ham. Meat Sci. 2008, 80, 728–737. [Google Scholar] [CrossRef]
- ISO. Guidance for the Design of Test Rooms; ISO Standard 8589:2007; ISO: Geneva, Switzerland, 2007. [Google Scholar]
- Asefa, D.T.; Gjerde, R.O.; Sidhu, M.S.; Langsrud, S.; Kure, C.F.; Nesbakken, T.; Skaar, I. Moulds contaminants on Norwegian dry-cured meat products. Int. J. Food Microbiol. 2009, 128, 435–439. [Google Scholar] [CrossRef]
- Kovačević, D. Kemija i Tehnologija Šunki i Pršuta; Prehrambeno-Tehnološki Fakultet: Osijek, Croatia, 2017. [Google Scholar]
- Geisen, R.; Schmidt-Heydt, M.; Touhami, N.; Himmelsbach, A. New aspects of ochratoxin A and citrinin biosynthesis in Penicillium. Curr. Opin. Food Sci. 2018, 23, 23–31. [Google Scholar] [CrossRef]
- Lešić, T.; Vahčić, N.; Kos, I.; Zadravec, M.; Sinčić Pulić, B.; Bogdanović, T.; Petričević, S.; Listeš, E.; Škrivanko, M.; Pleadin, J. Characterization of Traditional Croatian Household-Produced Dry-Fermented Sausages. Foods 2020, 9, 990. [Google Scholar] [CrossRef]
- Perrone, G.; Samson, R.A.; Frisvad, J.C.; Susca, A.; Gunde-Cimerman, N.; Epifani, F.; Houbraken, J. Penicillium salamii, a new species occurring during seasoning of dry-cured meat. Int. J. Food Microbiol. 2015, 193, 91–98. [Google Scholar] [CrossRef]
- Alapont, C.; López-Mendoza, M.C.; Gil, J.V.; Martínez-Culebras, P.V. Mycobiota and toxigenic Penicillium species on two Spanish dry-cured ham manufacturing plants. Food Addit. Contam. A 2014, 31, 93–104. [Google Scholar] [CrossRef]
- Núñez, F.; Rodríguez, M.M.; Bermúdez, M.E.; Córdoba, J.J.; Asensio, M.A. Composition and toxigenic potential of the mould population on dry-cured Iberian ham. Int. J. Food Microbiol. 1996, 32, 185–197. [Google Scholar] [CrossRef]
- Markov, K.; Pleadin, J.; Bevardi, M.; Vahčić, N.; Sokolić-Mihalek, D.; Frece, J. Natural occurrence of aflatoxin B1,ochratoxin A and citrinin in Croatian fermented meat products. Food Control 2013, 34, 312–317. [Google Scholar] [CrossRef]
- Vulić, A.; Lešić, T.; Kudumija, N.; Zadravec, M.; Kiš, M.; Vahčić, N.; Pleadin, J. The development of LC-MS/MS method of determination of cyclopiazonic acid in dry-fermented meat products. Food Control 2020, 123, 107814. [Google Scholar] [CrossRef]
- Pleadin, J.; Malenica Staver, M.; Vahčić, N.; Kovačević, D.; Milone, S.; Saftić, L.; Scortichini, G. Survey of aflatoxin B1 and ochratoxin A occurrence in traditional meat products coming from Croatian households and markets. Food Control 2015, 52, 71–77. [Google Scholar] [CrossRef]
- Iacumin, L.; Milesi, S.; Pirani, S.; Comi, G.; Chiesa, L.M. Ochratoxigenic mold and ochratoxin a in fermented sausages from different areas in Northern Italy: Occurrence, reduction or prevention with ozonated air. J. Food Saf. 2011, 31, 538–545. [Google Scholar] [CrossRef]
- Montanha, F.M.; Anater, A.; Burchard, J.F.; Luciano, F.L.; Meca, G.; Manyes, L.; Pimpão, C.T. Mycotoxins in dry-cured meats: A review. Food Chem. Toxicol. 2018, 111, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Peromingo, B.; Rodríguez, M.; Núñez, F.; Silva, A.; Rodríguez, A. Sensitive determination of cyclopiazonic acid in dry-cured ham using a QuEChERS method and UHPLC–MS/MS. Food Chem. 2018, 263, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, P.; Silva, D.; Costa, P.; Abrunhosa, L.; Venancio, A.; Teixeira, A. Mycobiota and mycotoxins in Portuguese pork, goat and sheep dry-cured hams. Mycotoxin Res. 2019, 35, 405–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz, A.; García-Torres, S.; González, E.; De Pedro-Sanz, E.J.; Gaspar, P.; Tejerina, D. Quality traits of fresh and dry-cured loin from Iberian × Duroc crossbred pig in the Montanera system according to slaughtering age. Meat Sci. 2020, 170, 108242. [Google Scholar] [CrossRef]
- Perez-Alvarez, J.A.; Fernandez-Lopez, J. Color Characteristic of Meat and poultry processing. In Sensory Analysis of Foods of Animal Origin; Nolet, L.M., Toldra, F., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 101–113. [Google Scholar]
- Toldra, F. Dry-Cured Meat Products; Food and Nutrition Press: Trumbull, CT, USA, 2002. [Google Scholar]
- Ludemann, V.; Pose, G.; Pollio, M.L.; Segura, J. Determination of growth characteristics and lipolytic and proteolytic activities of Penicillium strains isolated from Argentinean salami. Int. J. Food Microbiol. 2004, 96, 13–18. [Google Scholar] [CrossRef]
- García-González, D.L.; Tena, N.; Aparicio-Ruiz, R.; Morales, M.T. Relationship between sensory attributes and volatile compounds qualifying dry-cured hams. Meat Sci. 2008, 80, 315–325. [Google Scholar] [CrossRef]
- Bruna, J.M.; Hierro, E.M.; De la Hoz, L.; Mottram, D.S.; Fernández, M.; Ordóñez, J.A. The contribution of Penicillium aurantiogriseum to the volatile composition and sensory quality of dry fermented sausages. Meat Sci. 2001, 59, 97–107. [Google Scholar] [CrossRef]
- Flores, M. Flavour of meat products. In Sensory Analysis of Foods of Animal Origin; Nolet, L.M., Toldra, F., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 131–143. [Google Scholar]
- Bruna, J.M.; Hierro, E.M.; De la Hoz, L.; Mottram, D.S.; Fernández, M.; Ordóñez, J.A. Changes in selected biochemical and sensory parameters as affected by the superficial inoculation of Penicillium camemberti on dry fermented sausages. Int. J. Food Microbiol. 2003, 85, 111–125. [Google Scholar] [CrossRef]
- Kos, I.; Sinčić Pulić, B.; Gorup, D.; Kaić, A. Sensory profiles of artisanal smoked dry-cured ham as affected by production season. J. Cent. Eur. Agric. 2019, 20, 1089–1098. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sensory Traits | Definition (Range) |
|---|---|
| Appearance | |
| Colour of the muscle tissue | Intensity of red colour in the lean (pale pink to dark red) |
| Colour uniformity | Presence of colour homogeneity in the different muscles of a transversal cut (very low to very high) |
| Colour of the subcutaneous adipose tissue | Intensity of yellow colour of the fat (white to intense yellow) |
| Marbling | Level of visible intramuscular fat (very lean to intense marbled) |
| Surface humidity | Impresion of the surface wetness (very dry to very wet) |
| Tyrosine crystals | Presence of tyrosine crystals on the surface of a transversal cut (none to more than 10 crystals) |
| Odour | |
| Favourable odour | Intensity of the typical odour from cured meat products (very low to very high) |
| Unfavourable odour | Intensity of the off-odours (very low to very high) |
| Smoky odour | Perception of any type of smoke aroma (very low to very high) |
| Texture | |
| Tenderness | Effort required to bite thorough lean and to convert the sample to a swallowable state (very firm to very tender) |
| Juiciness | Impression of the release of juice during mastication (not to very juicy) |
| Taste | |
| Saltiness | Intensity of taste associated with sodium ions (not to very salty) |
| Sweetness | Intensity of sweetness (not to very sweet) |
| Sourness | Intensity of taste associated with citric acid (not to very sour) |
| Bitterness | Intensity of taste associated with caffeine (not to very bitter) |
| Aroma | |
| Specific aroma | Intensity of aroma associated with aromatic herbs (Istrian ham) and buttery aroma (Dalmatian ham) (very low to very high) |
| Biochemical aroma | Intensity of aroma associated with oxidised fat (very low to very high) |
| Fresh meat aroma | Intensity of aroma associated with fresh pork meat (very low to very high) |
| Moulds | Intensity of aroma associated with moulds (very low to very high) |
| Genus | Species | Number of Isolates | “Istarski Pršut” (n = 15) | “Dalmatinski Pršut” (n = 20) | ||
|---|---|---|---|---|---|---|
| Dr (%) | Fr (%) | Dr (%) | Fr (%) | |||
| Penicillium | P. salamii | 6 | 9 | 43 | - | - |
| P. polonicum | 2 | 3 | 14 | - | - | |
| P. citrinum | 14 | 9 | 43 | 12 | 40 | |
| P. bialowienzense | 4 | 6 | 29 | - | - | |
| P. solitum | 1 | - | - | 2 | 5 | |
| P. nalgiovense | 2 | - | - | 3 | 10 | |
| P. oxalicum | 2 | - | - | 3 | 10 | |
| Aspergillus | A. proliferans | 14 | 13 | 64 | 8 | 25 |
| A. chevalieri | 13 | 5 | 21 | 15 | 50 | |
| A. pseudoglaucus | 1 | - | - | 2 | 5 | |
| A. niger | 1 | - | - | 2 | 5 | |
| A. cibarius | 7 | - | - | 10 | 35 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lešić, T.; Vahčić, N.; Kos, I.; Zadravec, M.; Milićević, D.; Perković, I.; Listeš, E.; Pleadin, J. The Influence of Surface Mycobiota on Sensory Properties of “Istarski pršut” and “Dalmatinski pršut”. Processes 2021, 9, 2287. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9122287
Lešić T, Vahčić N, Kos I, Zadravec M, Milićević D, Perković I, Listeš E, Pleadin J. The Influence of Surface Mycobiota on Sensory Properties of “Istarski pršut” and “Dalmatinski pršut”. Processes. 2021; 9(12):2287. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9122287
Chicago/Turabian StyleLešić, Tina, Nada Vahčić, Ivica Kos, Manuela Zadravec, Dragan Milićević, Irena Perković, Eddy Listeš, and Jelka Pleadin. 2021. "The Influence of Surface Mycobiota on Sensory Properties of “Istarski pršut” and “Dalmatinski pršut”" Processes 9, no. 12: 2287. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9122287