Role of Microalgae in the Recovery of Nutrients from Pig Manure

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgae and Culture Conditions
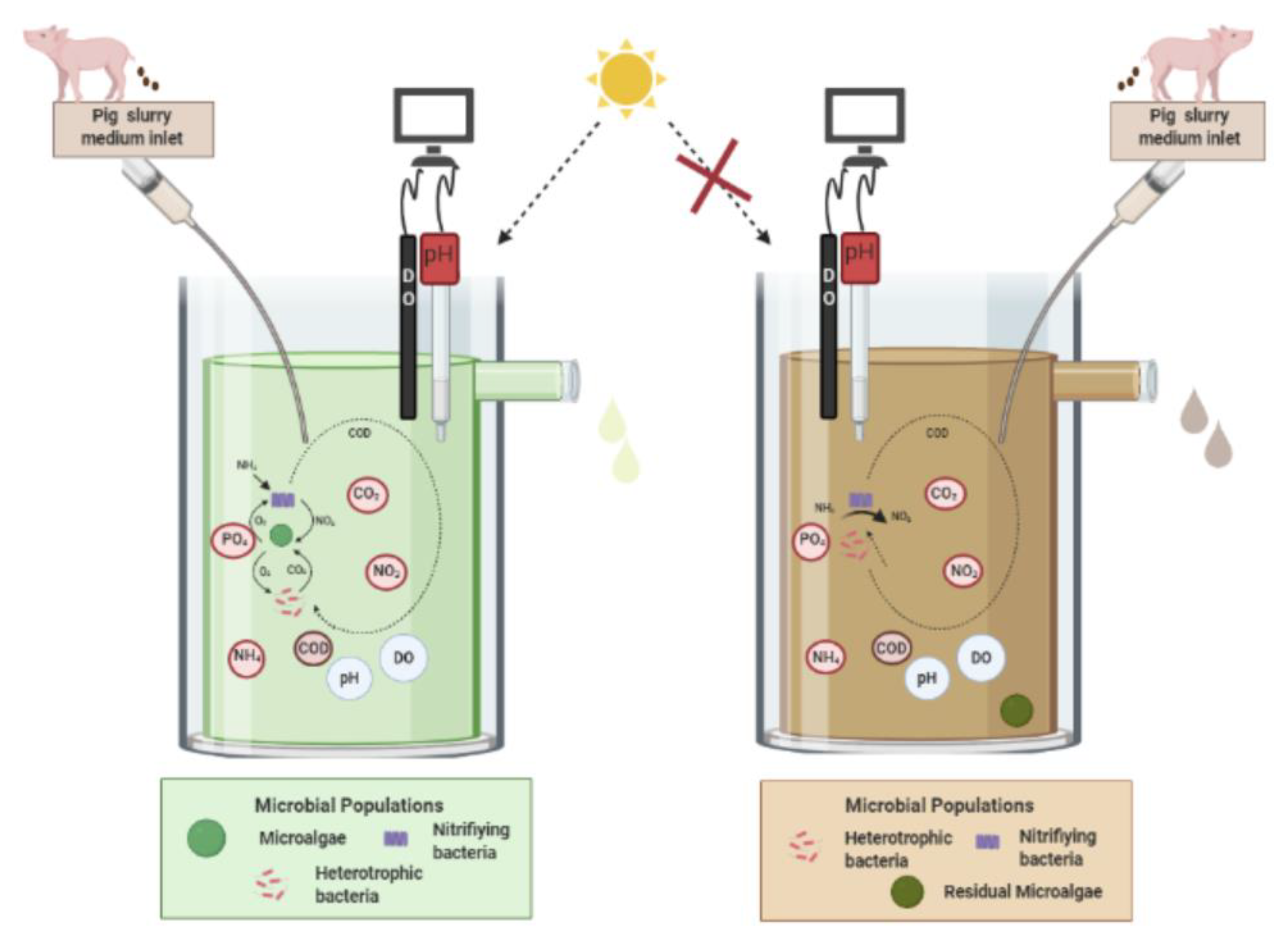
2.2. Photobioreactors
2.3. Photosynthesis and Respiration
2.4. Bacterial Counts
2.5. Statistical Analysis
3. Results
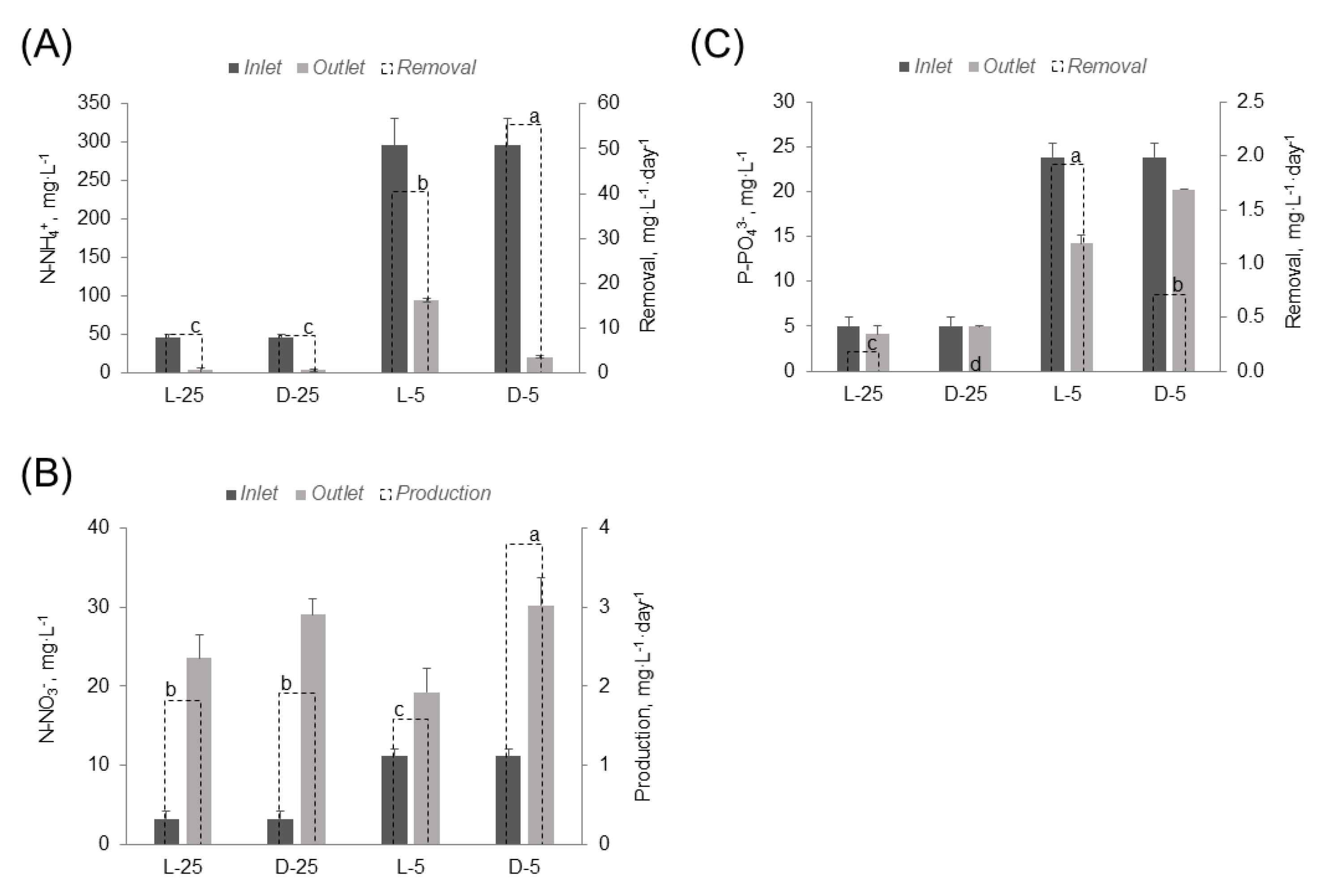
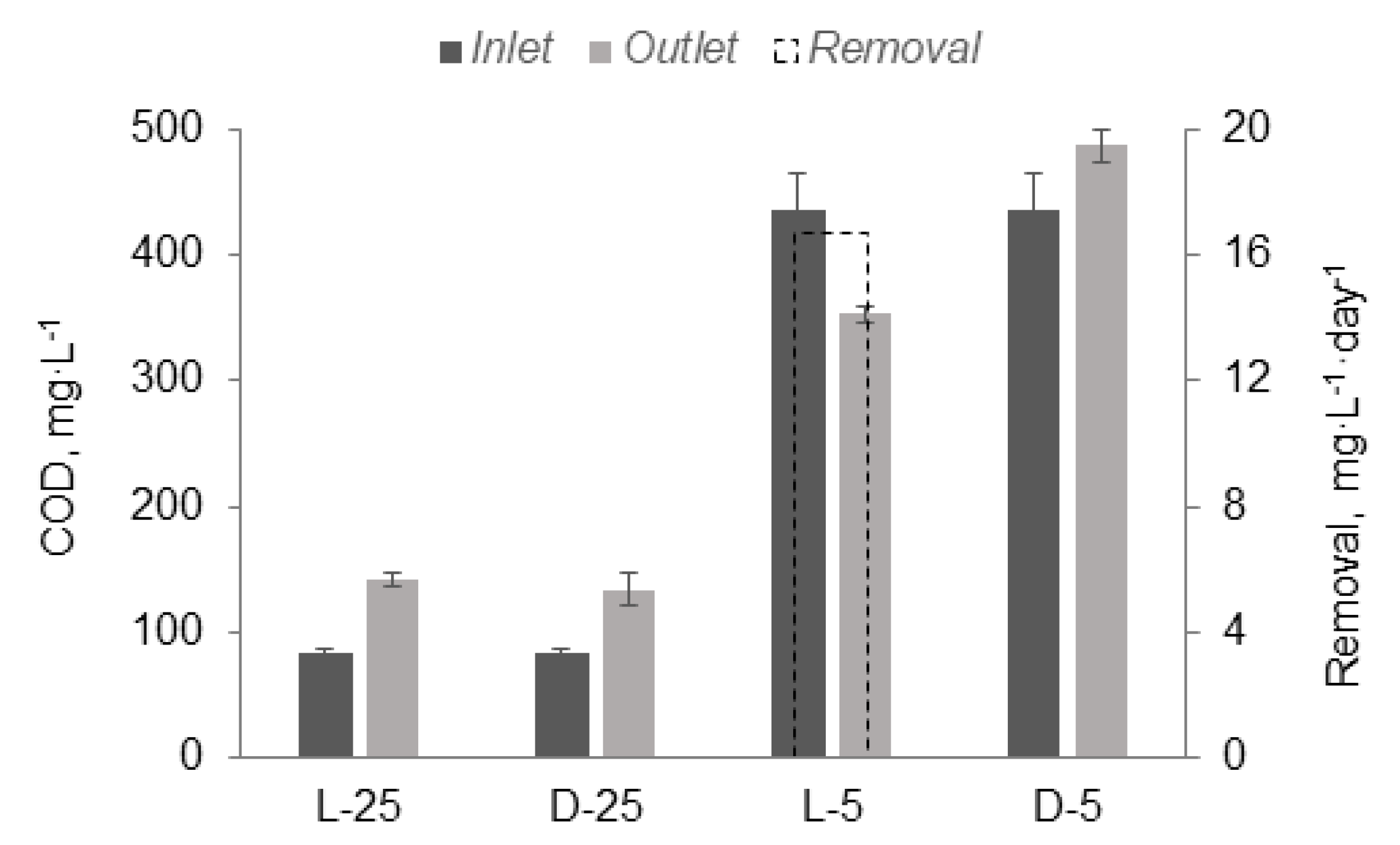
3.1. Nutrient Removal
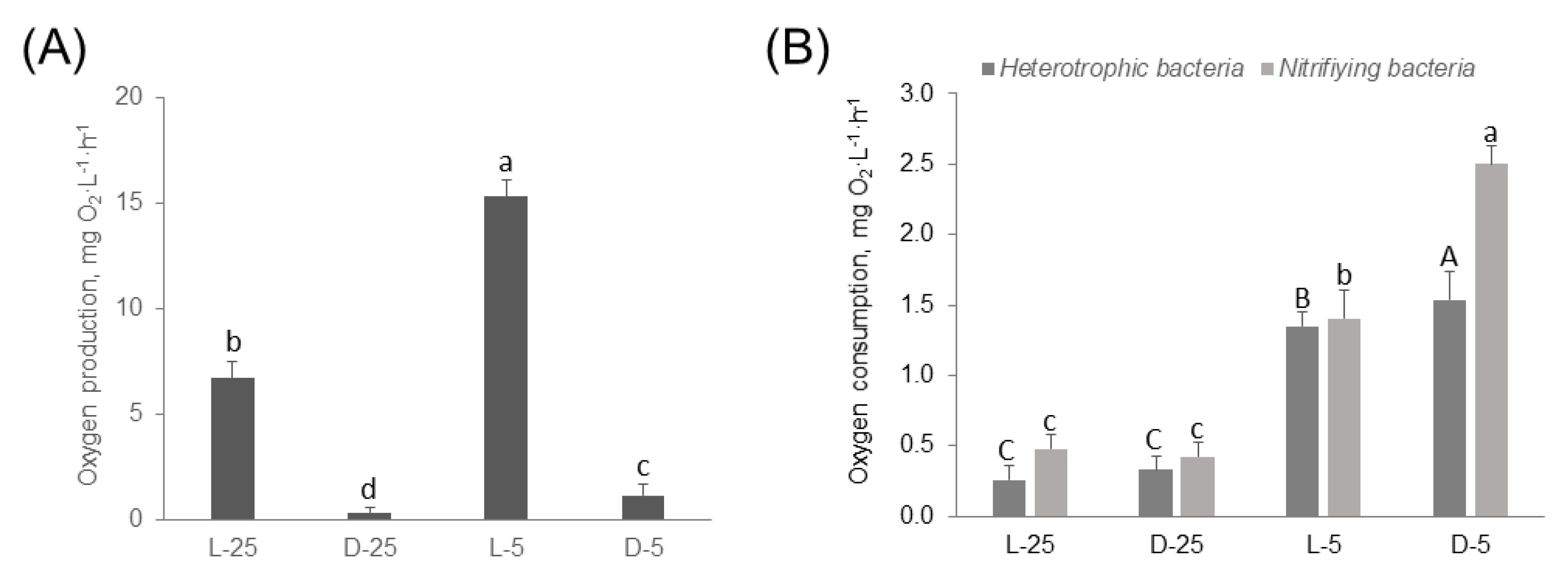
3.2. Respirometric Analysis
3.3. Microbiologic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, P. Malthus is still wrong: We can feed a world of 9–10 billion, but only by reducing food demand. In Nutrition Society; Cambridge University Press: Cambridge, UK, 2015; Volume 74, pp. 187–190. [Google Scholar]
- González, N.; Marquès, M.; Nadal, M.; Domingo, J.L. Meat consumption: Which are the current global risks? A review of recent (2010–2020) evidences. Food Res. Int. 2020, 137, 109341. [Google Scholar] [CrossRef] [PubMed]
- Faz, A.; Carmona, D.M.; Zanuzzi, A.; Mermut, A.R. Pig manure application for remediation of mine soils in Murcia Province, SE Spain. Sci. World J. 2008, 8, 819–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, D.; Posadas, E.; Grajeda, C.; Blanco, S.; Martínez-Páramo, S.; Acién, G.; García-Encina, P.; Bolado, S.; Muñoz, R. Comparative evaluation of piggery wastewater treatment in algal-bacterial photobioreactors under indoor and outdoor conditions. Bioresour. Technol. 2017, 245, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Acién Fernández, F.G.; Gómez-Serrano, C.; Fernández-Sevilla, J.M. Recovery of Nutrients From Wastewaters Using Microalgae. Front. Sustain. Food Syst. 2018, 2, 59. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Wu, X.; Zou, G.; Zhou, T.; Liu, Y.; Ruan, R. Cultivation of Chlorella vulgaris in manure-free piggery wastewater with high-strength ammonium for nutrients removal and biomass production: Effect of ammonium concentration, carbon/nitrogen ratio and pH. Bioresour. Technol. 2019, 273, 203–211. [Google Scholar] [CrossRef]
- García, D.; de Godos, I.; Domínguez, C.; Turiel, S.; Bolado, S.; Muñoz, R. A systematic comparison of the potential of microalgae-bacteria and purple phototrophic bacteria consortia for the treatment of piggery wastewater. Bioresour. Technol. 2019, 276, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Zurano, A.; Gómez-Serrano, C.; Acién-Fernández, F.G.; Fernández-Sevilla, J.M.; Molina-Grima, E. A novel photo-respirometry method to characterize consortia in microalgae-related wastewater treatment processes. Algal Res. 2020, 47. [Google Scholar] [CrossRef]
- Zambrano, J.; Krustok, I.; Nehrenheim, E.; Carlsson, B. A simple model for algae-bacteria interaction in photo-bioreactors. Algal Res. 2016, 19, 155–161. [Google Scholar] [CrossRef]
- Acién, F.G.; Gómez-Serrano, C.; Morales-Amaral, M.M.; Fernández-Sevilla, J.M.; Molina-Grima, E. Wastewater treatment using microalgae: How realistic a contribution might it be to significant urban wastewater treatment? Appl. Microbiol. Biotechnol. 2016, 100, 9013–9022. [Google Scholar] [CrossRef]
- Risgaard-Petersen, N.; Nicolaisen, M.H.; Revsbech, N.P.; Lomstein, B.A. Competition between ammonia-oxidizing bacteria and benthic microalgae. Appl. Environ. Microbiol. 2004, 70, 5528–5537. [Google Scholar] [CrossRef] [Green Version]
- González-Camejo, J.; Montero, P.; Aparicio, S.; Ruano, M.V.; Borrás, L.; Seco, A.; Barat, R. Nitrite inhibition of microalgae induced by the competition between microalgae and nitrifying bacteria. Water Res. 2020, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spanjers, H.; Vanrolleghem, P. Respirometry as a tool for rapid characterization of wastewater and activated sludge. Water Sci. Technol. 1995, 31, 105–114. [Google Scholar] [CrossRef]
- Munz, G.; Gori, R.; Mori, G.; Lubello, C. Monitoring biological sulphide oxidation processes using combined respirometric and titrimetric techniques. Chemosphere 2009, 76, 644–650. [Google Scholar] [CrossRef] [PubMed]
- Pambrun, V.; Marquot, A.; Racault, Y. Characterization of the toxic effects of cadmium and 3.5-dichlorophenol on nitrifying activity and mortality in biologically activated sludge systems—Effect of low temperature. Environ. Sci. Pollut. Res. 2008, 15, 592–599. [Google Scholar]
- Rossi, S.; Sforza, E.; Pastore, M.; Bellucci, M.; Casagli, F.; Marazzi, F.; Ficara, E. Photo-respirometry to shed light on microalgae-bacteria consortia—a review. Rev. Environ. Sci. Biotechnol. 2020, 19, 43–72. [Google Scholar] [CrossRef]
- Dubinsky, Z.; Falkowski, P.G.; Post, A.F.; Van Hes, U.M. A system for measuring phytoplankton photosynthesis in a defined light field with an oxygen electrode. J. Plankton Res. 1987, 9, 607–612. [Google Scholar] [CrossRef]
- Decostere, B.; Janssens, N.; Alvarado, A.; Maere, T.; Goethals, P.; Van Hulle, S.W.H.; Nopens, I. A combined respirometer–titrimeter for the determination of microalgae kinetics: Experimental data collection and modelling. Chem. Eng. J. 2013, 222, 85–93. [Google Scholar] [CrossRef]
- Sforza, E.; Pastore, M.; Barbera, E.; Bertucco, A. Respirometry as a tool to quantify kinetic parameters of microalgal mixotrophic growth. Bioprocess Biosyst. Eng. 2019, 42, 839–851. [Google Scholar] [CrossRef]
- Rossi, S.; Bellucci, M.; Marazzi, F.; Mezzanotte, V.; Ficara, E. Activity assessment of microalgal-bacterial consortia based on respirometric tests. Water Sci. Technol. 2018, 78, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Petrini, S.; Foladori, P.; Donati, L.; Andreottola, G. Comprehensive respirometric approach to assess photosynthetic, heterotrophic and nitrifying activity in microalgal-bacterial consortia treating real municipal wastewater. Biochem. Eng. J. 2020, 161, 107697. [Google Scholar] [CrossRef]
- Morillas-España, A.; Lafarga, T.; Gómez-Serrano, C.; Acién-Fernández, F.G.; González-López, C.V. Year-long production of Scenedesmus almeriensis in pilot-scale raceway and thin-layer cascade photobioreactors. Algal Res. 2020, 51, 102069. [Google Scholar] [CrossRef]
- Allen, M.B.; Arnon, D.I. Studies on Nitrogen-fixing Blue-green Algae. Physiol. Plant. 1955, 8, 653–660. [Google Scholar] [CrossRef]
- Garrido-Cardenas, J.A.; Manzano-Agugliaro, F.; Acien-Fernandez, F.G.; Molina-Grima, E. Microalgae research worldwide. Algal Res. 2018, 35, 50–60. [Google Scholar] [CrossRef]
- Liu, J.; Wu, Y.; Wu, C.; Muylaert, K.; Vyverman, W.; Yu, H.Q.; Muñoz, R.; Rittmann, B. Advanced nutrient removal from surface water by a consortium of attached microalgae and bacteria: A review. Bioresour. Technol. 2017, 241, 1127–1137. [Google Scholar] [CrossRef] [PubMed]
- Mengesha, S.; Dehairs, F.; Elskens, M.; Goeyens, L. Phytoplankton Nitrogen Nutrition in the Western Indian Ocean: Ecophysiological Adaptations of Neritic and Oceanic Assemblages to Ammonium Supply. Estuar. Coast. Shelf Sci. 1999, 48, 589–598. [Google Scholar] [CrossRef]
- Solimeno, A.; Samsó, R.; Uggetti, E.; Sialve, B.; Steyer, J.-P.; Gabarró, A.; García, J. New mechanistic model to simulate microalgae growth. Algal Res. 2015, 12, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Delgadillo-Mirquez, L.; Lopes, F.; Taidi, B.; Pareau, D. Nitrogen and phosphate removal from wastewater with a mixed microalgae and bacteria culture. Biotechnol. Rep. 2016. [Google Scholar] [CrossRef]
- Larsdotter, K.; la Cour Jansen, J.; Dalhammar, G. Phosphorus removal from wastewater by microalgae in Sweden—A year-round perspective. Environ. Technol. 2010, 31, 117–123. [Google Scholar] [CrossRef]
- Powell, N.; Shilton, A.; Chisti, Y.; Pratt, S. Towards a luxury uptake process via microalgae—Defining the polyphosphate dynamics. Water Res. 2009, 43, 4207–4213. [Google Scholar] [CrossRef]
- De Godos, I.; Vargas, V.A.; Blanco, S.; González, M.C.G.; Soto, R.; García-Encina, P.A.; Becares, E.; Muñoz, R. A comparative evaluation of microalgae for the degradation of piggery wastewater under photosynthetic oxygenation. Bioresour. Technol. 2010, 101, 5150–5158. [Google Scholar] [CrossRef]
- García, D.; Posadas, E.; Blanco, S.; Acién, G.; García-Encina, P.; Bolado, S.; Muñoz, R. Evaluation of the dynamics of microalgae population structure and process performance during piggery wastewater treatment in algal-bacterial photobioreactors. Bioresour. Technol. 2018, 248, 120–126. [Google Scholar] [CrossRef] [Green Version]
- González, C.; Marciniak, J.; Villaverde, S.; García-Encina, P.A.; Muñoz, R. Microalgae-based processes for the biodegradation of pretreated piggery wastewaters. Appl. Microbiol. Biotechnol. 2008, 80, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Cabanelas, I.T.D.; Ruiz, J.; Arbib, Z.; Chinalia, F.A.; Garrido-Pérez, C.; Rogalla, F.; Nascimento, I.A.; Perales, J.A.; Garrido-P??rez, C.; Rogalla, F.; et al. Comparing the use of different domestic wastewaters for coupling microalgal production and nutrient removal. Bioresour. Technol. 2013, 131, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Tan, K.; He, L.; Qiu, Y.; Tan, W.; Guo, Y.; Wang, Z.; Sun, W. Effect of limitation of iron and manganese on microalgae growth in fresh water. Microbiology 2018, 164, 1514–1521. [Google Scholar] [CrossRef] [PubMed]
- Ushizaka, S.; Kuma, K.; Suzuki, K. Effects of Mn and Fe on growth of a coastal marine diatom Talassiosira weissflogii in the presence of precipitated Fe(III) hydroxide and EDTA-Fe(III) complex. Fish. Sci. 2011, 77, 411–424. [Google Scholar] [CrossRef]
- Tortell, P.D.; Maldonado, M.T.; Granger, J.; Price, N.M. Marine bacteria and biogeochemical cycling of iron in the oceans. FEMS Microbiol. Ecol. 1999, 29, 1–11. [Google Scholar] [CrossRef]
- Yao, S.; Lyu, S.; An, Y.; Lu, J.; Gjermansen, C.; Schramm, A. Microalgae-bacteria symbiosis in microalgal growth and biofuel production: A review. J. Appl. Microbiol. 2019, 126, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Babu, M.A.; Hes, E.M.A.; van der Steen, N.P.; Hooijmans, C.M.; Gijzen, H.J. Nitrification rates of algal-bacterial biofilms in wastewater stabilization ponds under light and dark conditions. Ecol. Eng. 2010, 36, 1741–1746. [Google Scholar] [CrossRef]
- Allen, M.J.; Edberg, S.C.; Reasoner, D.J. Heterotrophic plate count bacteria—What is their significance in drinking water? Int. J. Food Microbiol. 2004, 92, 265–274. [Google Scholar] [CrossRef] [Green Version]
- Smriga, S.; Fernandez, V.I.; Mitchell, J.G.; Stocker, R. Chemotaxis toward phytoplankton drives organic matter partitioning among marine bacteria. Proc. Natl. Acad. Sci. USA 2016. [Google Scholar] [CrossRef] [Green Version]
- Ramanan, R.; Kim, B.H.; Cho, D.H.; Oh, H.M.; Kim, H.S. Algae-bacteria interactions: Evolution, ecology and emerging applications. Biotechnol. Adv. 2016, 34, 14–29. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Raouf, N.; Al-Homaidan, A.A.; Ibraheem, I.B.M. Microalgae and wastewater treatment. Saudi J. Biol. Sci. 2012, 19, 257–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Piggery Wastewater | Arnon |
|---|---|---|
| pH | 8.1 ± 0.3 | 7.5 ± 0.2 |
| COD | 2181.7 ± 100.9 | 16.0 ± 1.2 |
| Nitrogen-Nitrate | 56.4 ± 2.7 | 140.0 ± 4.5 |
| Chloride | 2060.2 ± 23.5 | 78.9 ± 2.1 |
| Potassium | 1800 ± 1.6 | 325.1 ± 6.3 |
| Calcium | 350.1 ± 0.2 | 364.9 ± 5.5 |
| Magnesium | 108.2 ± 14.1 | 12.2 ± 0.6 |
| Phosphorus-Phosphate | 119.2 ± 5.1 | 39.3 ± 3.1 |
| Nitrogen-Ammonium | 1485.6 ± 17.7 | 0.0 ± 0.1 |
| Iron | 4.8 ± 0.01 | 5.0 ± 0.3 |
| Copper | 1.1 ± 0.1 | 0.02 ± 0.00 |
| Manganese | 2.6 ± 0.0 | 0.5 ± 0.02 |
| Zinc | 20.1 ± 0.2 | 0.06 ± 0.01 |
| Boron | 5.3 ± 0.1 | 0.4 ± 0.0 |
| L-25 | D-25 | L-5 | D-5 | |
|---|---|---|---|---|
| Heterotrophic bacteria | 2.50 × 105 | 1.75 × 104 | 2.35 × 105 | 1.35 × 105 |
| Coliforms bacteria | 2.60 × 101 | 2.05 × 102 | 7.80 × 101 | 4.11 × 102 |
| E. coli | ND | ND | ND | ND |
| Salmonella sp. | ND | ND | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Zurano, A.; Ciardi, M.; Lafarga, T.; Fernández-Sevilla, J.M.; Bermejo, R.; Molina-Grima, E. Role of Microalgae in the Recovery of Nutrients from Pig Manure. Processes 2021, 9, 203. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9020203
Sánchez-Zurano A, Ciardi M, Lafarga T, Fernández-Sevilla JM, Bermejo R, Molina-Grima E. Role of Microalgae in the Recovery of Nutrients from Pig Manure. Processes. 2021; 9(2):203. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9020203
Chicago/Turabian StyleSánchez-Zurano, Ana, Martina Ciardi, Tomás Lafarga, José María Fernández-Sevilla, Ruperto Bermejo, and Emilio Molina-Grima. 2021. "Role of Microalgae in the Recovery of Nutrients from Pig Manure" Processes 9, no. 2: 203. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9020203