
Toxicological and Epigenetic Studies of Two Types of Ale Beer, Tyrosol and Iso-Alpha Humulone

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation and Simple Compounds
2.2. In Vivo Assays
- mwh/mwh, a recessive mutation (multiple wing hairs) that induces multiple hairs per homozygous cell instead of one per cell in the wild-type phenotype [43].
- flr3/In (3LR) TM3, rippsep bx34eesBdS, where the flr3 (flare) mutation is homozygous recessive lethal and cells carrying both doses are able to produce deformed hairs in the late stages of larval development [44].
2.2.1. Toxicity and Antitoxicity Assays
2.2.2. Genotoxicity and Antigenotoxicity Assays (SMART)
2.2.3. Lifespan Assays
2.3. In Vitro Cytotoxicity Assays
2.3.1. Cell Culture
2.3.2. Cytotoxicity Assay
2.3.3. Determination of DNA Fragmentation
2.3.4. Comet Assay
2.3.5. Methylation Status
3. Results and Discussion
3.1. In Vivo
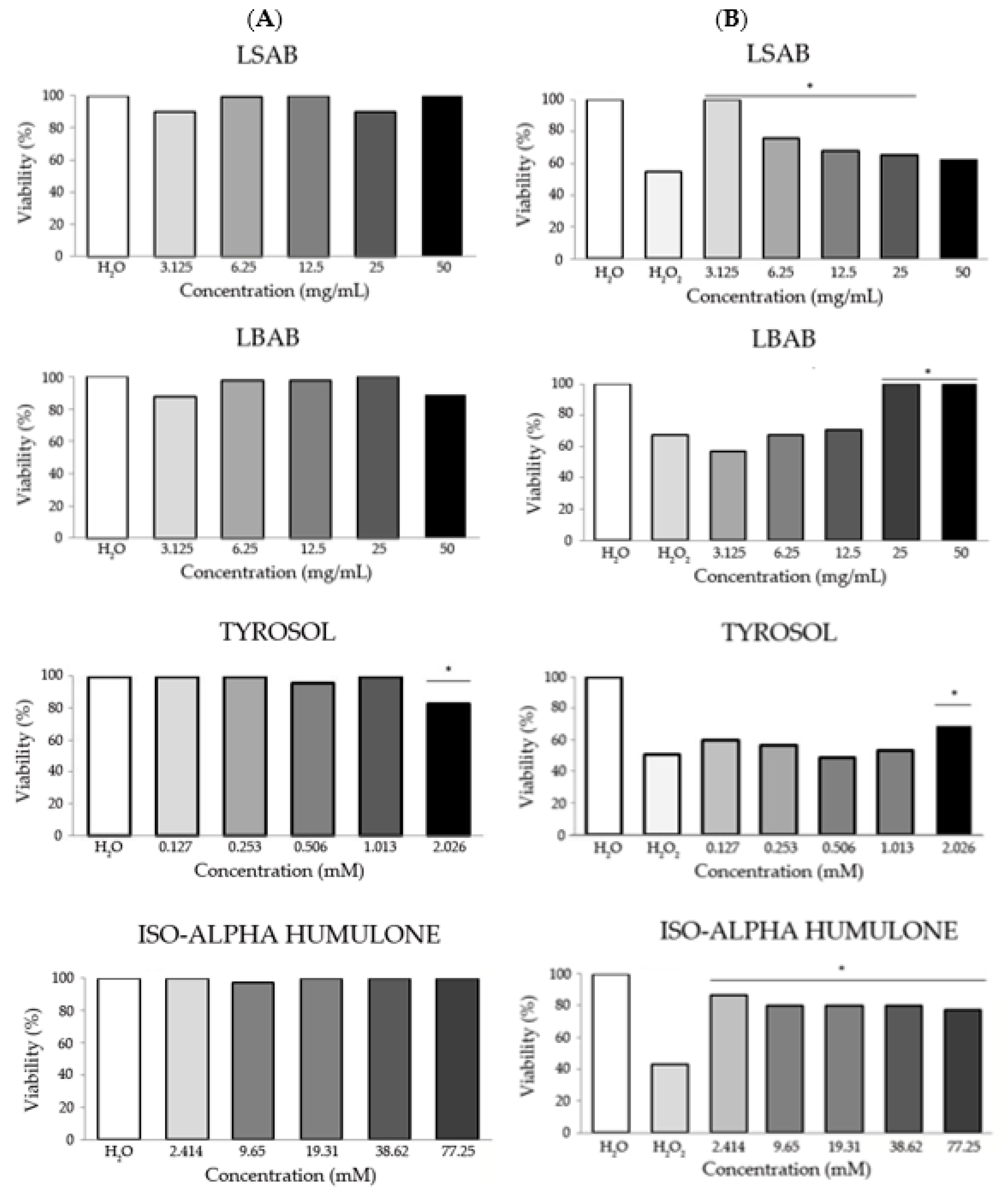
3.1.1. Toxicity and Antitoxicity
3.1.2. Genotoxicity and Antigenotoxicity
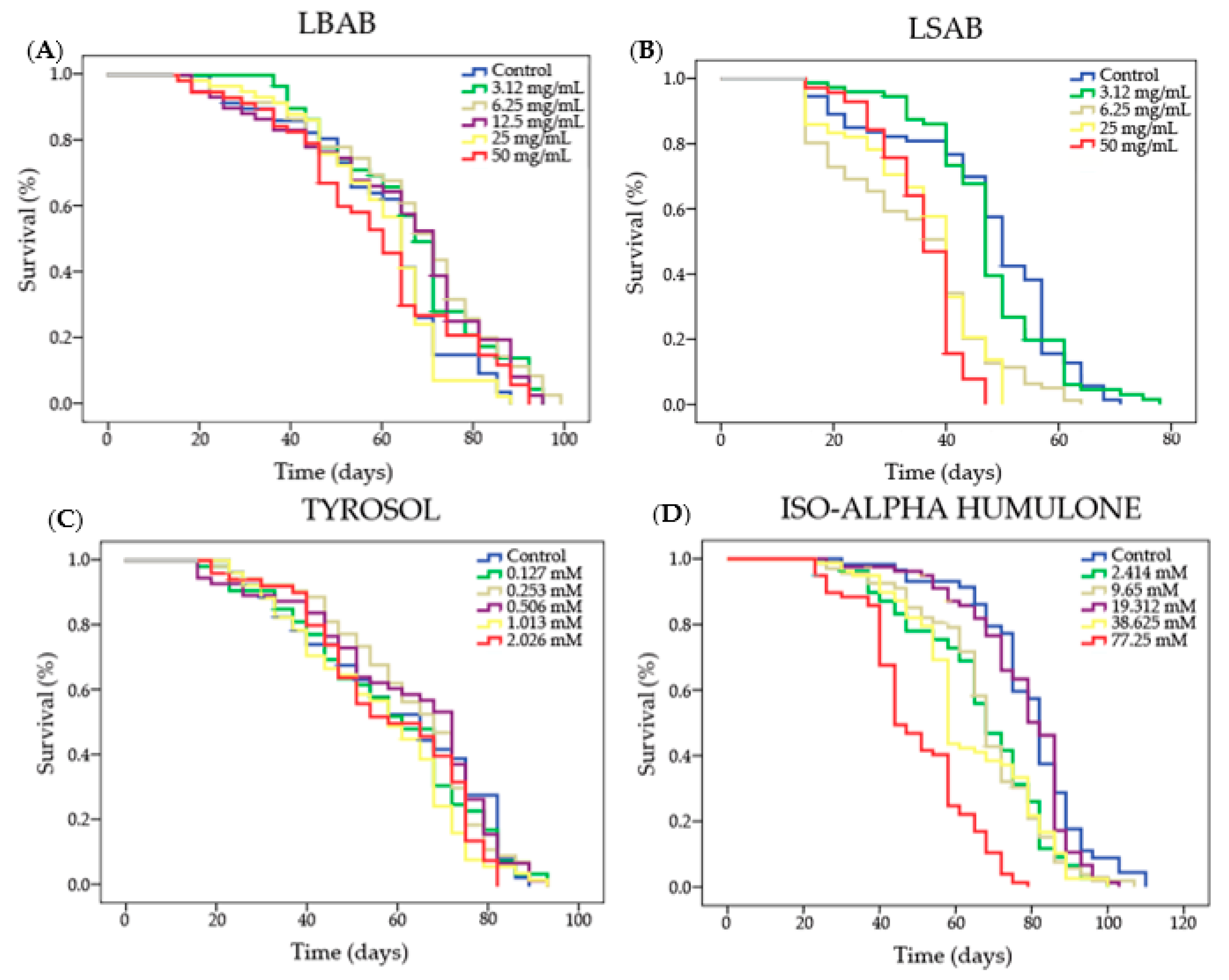
3.1.3. Lifespan
3.1.4. Healthspan
3.2. In Vitro
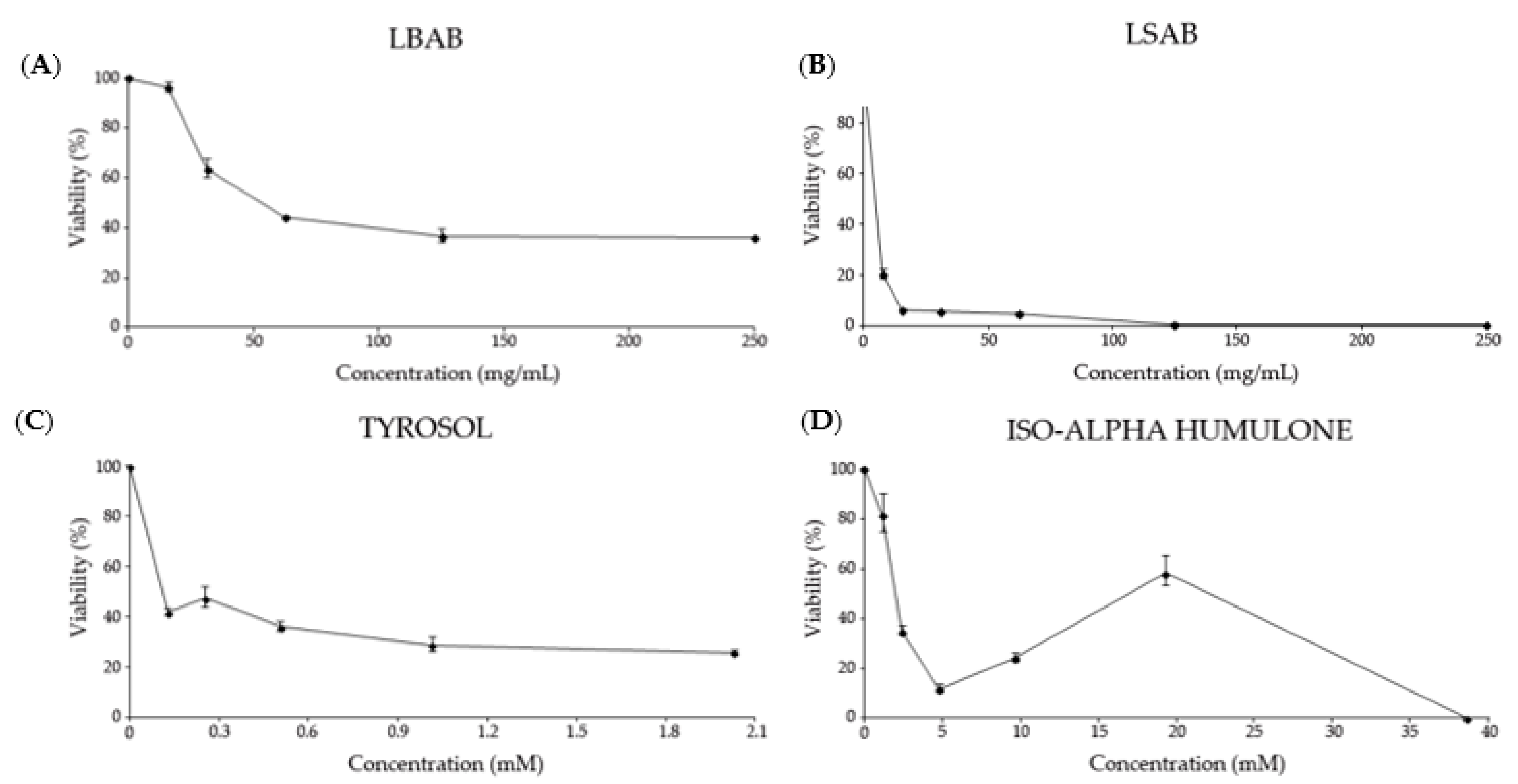
3.2.1. Cytotoxicity
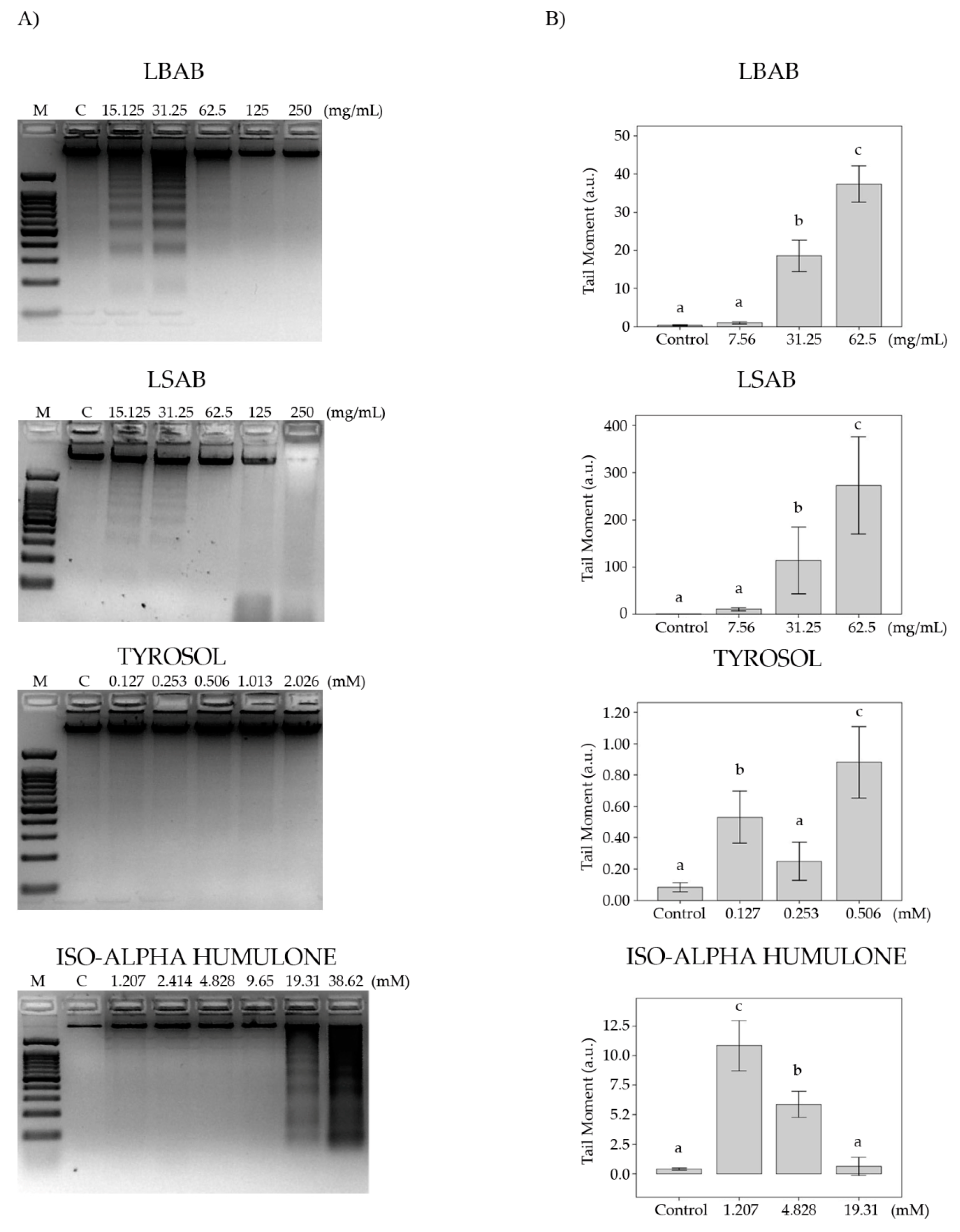
3.2.2. DNA Fragmentation
3.2.3. Comet Assay
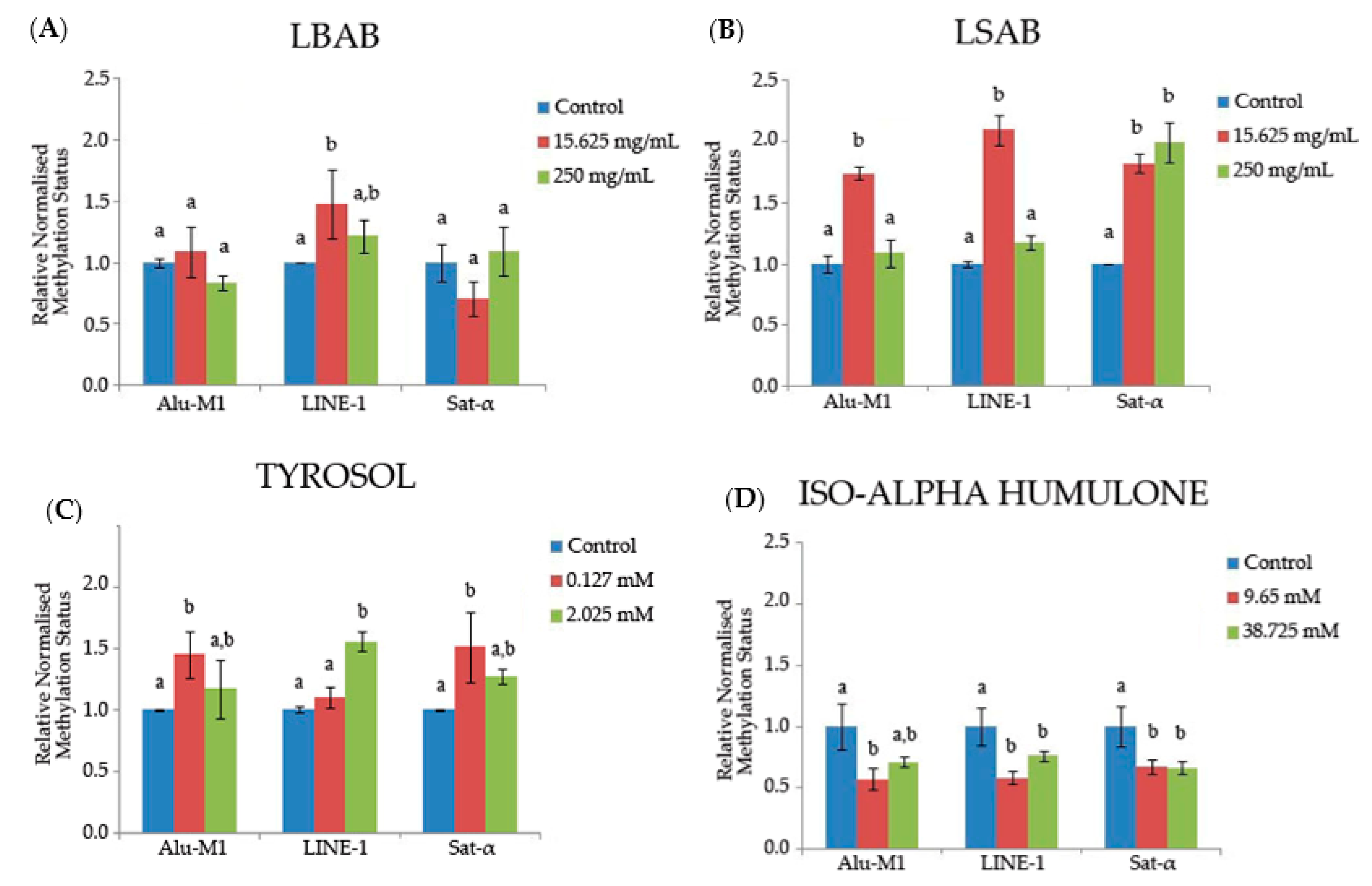
3.2.4. Methylation Status
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Martín-Moreno, J.M.; Soerjomataram, I.; Magnusson, G. Cancer causes and prevention: A condensed appraisal in Europe in 2008. Eur. J. Cancer 2008, 44, 1390–1403. [Google Scholar] [CrossRef]
- Reddy, L.; Odhav, B.; Bhoola, K. Natural products for cancer prevention: A global perspective. Pharmacol. Ther. 2003, 99, 1–13. [Google Scholar] [CrossRef]
- Doll, R.; Peto, R. The Causes of Cancer: Quantitative Estimates of Avoidable Risks of Cancer in the United States Today. J. Natl. Cancer Inst. 1981, 66, 1192–1308. [Google Scholar] [CrossRef]
- Clayton, D.; McKeigue, P.M. Epidemiological methods for studying genes and environmental factors in complex diseases. Lancet 2001, 358, 1356–1360. [Google Scholar] [CrossRef]
- Walker, J.; Hell, J.; Liszt, K.I.; Dresel, M.; Pignitter, M.; Hofmann, T.; Somoza, V. Identification of Beer Bitter Acids Regulating Mechanisms of Gastric Acid Secretion. J. Agric. Food Chem. 2012, 60, 1405–1412. [Google Scholar] [CrossRef]
- Piendl, A. On the importance of beer in nutrition today. Brauwelt 1989, 124, 546–552. [Google Scholar]
- Chen, W.; Becker, T.; Qian, F.; Ring, J. Beer and beer compounds: Physiological effects on skin health. J. Eur. Acad. Dermatol. Venereol. 2013, 28, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Buiatti, S. Chapter 20-Beer Composition: An Overview; Academic Press: London, UK, 2009; pp. 212–225. [Google Scholar]
- Rimm, E.; Giovannucci, E.; Willett, W.; Colditz, G.; Ascherio, A.; Rosner, B.; Stampfer, M. Prospective study of alcohol consumption and risk of coronary disease in men. Lancet 1991, 338, 464–468. [Google Scholar] [CrossRef]
- Muller, R.; Walker, S.; Brauer, J.; Junquera, M. Does Beer Contain Compounds That Might Interfere with Cholesterol Metabolism? J. Inst. Brew. 2007, 113, 102–109. [Google Scholar] [CrossRef]
- Curhan, G.C.; Willett, W.C.; Rimm, E.B.; Spiegelman, D.; Stampfer, M.J. Prospective Study of Beverage Use and the Risk of Kidney Stones. Am. J. Epidemiol. 1996, 143, 240–247. [Google Scholar] [CrossRef]
- Gerhäuser, C. Beer constituents as potential cancer chemopreventive agents. Eur. J. Cancer 2005, 41, 1941–1954. [Google Scholar] [CrossRef] [PubMed]
- Preedy, V.R. Beer in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Dewapriya, P.; Himaya, S.; Li, Y.-X.; Kim, S.-K.; Hewage, H.S.W. Tyrosol exerts a protective effect against dopaminergic neuronal cell death in in vitro model of Parkinson’s disease. Food Chem. 2013, 141, 1147–1157. [Google Scholar] [CrossRef] [PubMed]
- Tressl, R.; Renner, R.; Apetz, M. Volatile phenolic components in beer, smoked beer, and sherry (author’s transl). Zeitschrift fuer Lebensmittel-Untersuchung und-Forschung 1976, 162, 115. [Google Scholar] [CrossRef] [PubMed]
- Mateos, R.; Domínguez, M.M.; Espartero, A.J.L.; Cert, A. Antioxidant Effect of Phenolic Compounds, α-Tocopherol, and Other Minor Components in Virgin Olive Oil. J. Agric. Food Chem. 2003, 51, 7170–7175. [Google Scholar] [CrossRef] [PubMed]
- Rietjens, S.J.; Bast, A.; De Vente, J.; Haenen, G.R.M.M. The olive oil antioxidant hydroxytyrosol efficiently protects against the oxidative stress-induced impairment of the NO response of isolated rat aorta. Am. J. Physiol. Circ. Physiol. 2007, 292, H1931–H1936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannini, C.; Straface, E.; Modesti, D.; Coni, E.; Cantafora, A.; De Vincenzi, M.; Malorni, W.; Masella, R. Tyrosol, the Major Olive Oil Biophenol, Protects Against Oxidized-LDL-Induced Injury in Caco-2 Cells. J. Nutr. 1999, 129, 1269–1277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuel, S.M.; Thirunavukkarasu, M.; Penumathsa, S.V.; Paul, D.; Maulik, N. Akt/FOXO3a/SIRT1-mediated cardioprotection by n-tyrosol against ischemic stress in rat in vivo model of myocardial infarction: Switching gears toward survival and longevity. J. Agric. Food Chem. 2008, 56, 9692–9698. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Isaak, C.K.; Zhou, Y.; Petkau, J.C.; Liu, Y.; Siow, Y.L. Salidroside and tyrosol from Rhodiola protect H9c2 cells from ischemia/reperfusion-induced apoptosis. Life Sci. 2012, 91, 151–158. [Google Scholar] [CrossRef]
- Cañuelo, A.; Gilbert-López, B.; Pacheco-Liñán, P.; Martínez-Lara, E.; Siles-Rivas, E.; Miranda-Vizuete, A. Tyrosol, a main phenol present in extra virgin olive oil, increases lifespan and stress resistance in Caenorhabditis elegans. Mech. Ageing Dev. 2012, 133, 563–574. [Google Scholar] [CrossRef]
- Bu, Y.; Rho, S.; Kim, J.; Kim, M.Y.; Lee, D.H.; Kim, S.Y.; Choi, H.; Kim, H. Neuroprotective effect of tyrosol on transient focal cerebral ischemia in rats. Neurosci. Lett. 2007, 414, 218–221. [Google Scholar] [CrossRef]
- De Keukeleire, J.; Ooms, G.; Heyerick, A.; Roldán-Ruiz, I.; Van Bockstaele, E.; De Keukeleire, D. Formation and Accumulation of α-Acids, β-Acids, Desmethylxanthohumol, and Xanthohumol during Flowering of Hops (Humulus lupulusL.). J. Agric. Food Chem. 2003, 51, 4436–4441. [Google Scholar] [CrossRef]
- De Keukeleire, D. Fundamentals of beer and hop chemistry. Química Nova 2000, 23, 108–112. [Google Scholar] [CrossRef]
- Verzele, M.; De Keukeleire, D. Chemistry and Analysis of Hop and Beer Bitter Acids; Elsevier: Amsterdam, The Netherlands, 2013; Volume 27. [Google Scholar]
- Lu, Y.; Bergenståhl, B. Condensation of iso-humulone in solution and at hydrophobic surfaces. Colloids Surf. A Physicochem. Eng. Asp. 2021, 613, 126102. [Google Scholar] [CrossRef]
- Lu, Y.; Bergenståhl, B.; Nilsson, L. Interfacial properties and interaction between beer wort protein fractions and iso-humulone. Food Hydrocoll. 2020, 103, 105648. [Google Scholar] [CrossRef]
- Simpson, W.J. Ionophoric action of trans-isohumulone on Lactobacillus brevis. J. Gen. Microbiol. 1993, 139, 1041–1045. [Google Scholar] [CrossRef] [Green Version]
- Merinas-Amo, T.; Merinas-Amo, R.; García-Zorrilla, V.; Velasco-Ruiz, A.; Chladek, L.; Plachy, V.; Del Río-Celestino, M.; Font, R.; Kokoska, L.; Alonso-Moraga, Á. Toxicological Studies of Czech Beers and Their Constituents. Foods 2019, 8, 328. [Google Scholar] [CrossRef] [Green Version]
- Bohra, G.; Klimob, K.; Beckera, H.; Gerhäuserb, C. Cancer chemopreventive potential of humulones and isohumulones (hops α-and iso-α-acids): Induction of NAD (P) H: Quinone reductase as a novel mechanism. Nat. Prod. Commun. 2008, 3, 1934578X0800301206. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Wang, J.; Yamamoto, S.; Tobe, H. Suppression of cyclooxygenase-2 gene transcription by humulon of beer hop extract studied with reference to glucocorticoid. FEBS Lett. 2000, 465, 103–106. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-C.; Kundu, J.K.; Hwang, D.-M.; Na, H.-K.; Surh, Y.-J. Humulone inhibits phorbol ester-induced COX-2 expression in mouse skin by blocking activation of NF-κB and AP-1: IκB kinase and c-Jun-N-terminal kinase as respective potential upstream targets. Carcinogenesis 2007, 28, 1491–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimamura, M.; Hazato, T.; Ashino, H.; Yamamoto, Y.; Iwasaki, E.; Tobe, H.; Yamamoto, K.; Yamamoto, S. Inhibition of Angiogenesis by Humulone, a Bitter Acid from Beer Hop. Biochem. Biophys. Res. Commun. 2001, 289, 220–224. [Google Scholar] [CrossRef]
- Sánchez, C.L.; Franco, L.; Bravo, R.; Rubio, C.; Rodríguez, A.B.; Barriga, C.; Cubero, J. Cerveza y salud, beneficios en el sueño. Rev. Esp. Nutr. Comunitaria. 2010, 16, 160–163. [Google Scholar] [CrossRef]
- Anter, J.; Campos-Sánchez, J.; El Hamss, R.; Rojas-Molina, M.; Muñoz-Serrano, A.; Analla, M.; Alonso-Moraga, Á. Modulation of genotoxicity by extra-virgin olive oil and some of its distinctive components assessed by use of the Drosophila wing-spot test. Mutat. Res. Toxicol. Environ. Mutagen. 2010, 703, 137–142. [Google Scholar] [CrossRef]
- Richardson, H.E.; Willoughby, L.; Humbert, P.O. Screening for Anti-cancer Drugs in Drosophila. eLS 2015, 1–14. [Google Scholar]
- Graf, U.; Würgler, F.E.; Katz, A.J.; Frei, H.; Juon, H.; Hall, C.B.; Kale, P.G. Somatic mutation and recombination test in Drosophila melanogaster. Environ. Mutagen. 1984, 6, 153–188. [Google Scholar] [CrossRef] [PubMed]
- Ja, W.W.; Carvalho, G.B.; Mak, E.M.; De La Rosa, N.N.; Fang, A.Y.; Liong, J.C.; Brummel, T.; Benzer, S. Prandiology of Drosophila and the CAFE assay. Proc. Natl. Acad. Sci. USA 2007, 104, 8253–8256. [Google Scholar] [CrossRef] [Green Version]
- Kirin. Globar Beer Consumption by Country in 2011. Available online: http://www.kirinholdings.co.jp/english/news/2015/1224_01.html (accessed on 24 December 2015).
- Gonzalez, C. Drosophila melanogaster: A model and a tool to investigate malignancy and identify new therapeutics. Nat. Rev. Cancer 2013, 13, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Lints, F.A.; Soliman, M.H. Drosophila as a Model Organism for Ageing Studies; Springer: Berlin/Heidelberg, Germany, 1988. [Google Scholar]
- Rudrapatna, V.A.; Cagan, R.L.; Das, T.K. Drosophila cancer models. Dev. Dyn. 2011, 241, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Huen, D.; Morely, T.; Johnson, G.; Gubb, D.; Roote, J.; Adler, P.N. The multiple-wing-hairs Gene Encodes a Novel GBD–FH3 Domain-Containing Protein That Functions Both Prior to and After Wing Hair Initiation. Genetics 2008, 180, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Ren, N.; Charlton, J.; Adler, P.N. The flare Gene, Which Encodes the AIP1 Protein of Drosophila, Functions to Regulate F-Actin Disassembly in Pupal Epidermal Cells. Genetics 2007, 176, 2223–2234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasset-Cuevas, I.; Fernández-Bedmar, Z.; Lozano-Baena, M.D.; Campos-Sánchez, J.; De Haro-Bailón, A.; Muñoz-Serrano, A.; Alonso-Moraga, Á. Protective Effect of Borage Seed Oil and Gamma Linolenic Acid on DNA: In Vivo and In Vitro Studies. PLoS ONE 2013, 8, e56986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graf, U.; Abraham, S.K.; Guzmán-Rincón, J.; Würgler, F.E. Antigenotoxicity studies in Drosophila melanogaster. Mutat. Res. Mol. Mech. Mutagen. 1998, 402, 203–209. [Google Scholar] [CrossRef]
- Frei, H.; Würgler, F. Statistical methods to decide whether mutagenicity test data from Drosophila assays indicate a positive, negative, or inconclusive result. Mutat. Res. Mutagen. Relat. Subj. 1988, 203, 297–308. [Google Scholar] [CrossRef]
- Frei, H.; Würgler, F.E. Optimal experimental design and sample size for the statistical evaluation of data from somatic mutation and recombination tests (SMART) in Drosophila. Mutat. Res. Mutagen. Relat. Subj. 1995, 334, 247–258. [Google Scholar] [CrossRef]
- Abraham, S.K. Antigenotoxicity of coffee in the Drosophila assay for somatic mutation and recombination. Mutagenesis 1994, 9, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Soh, J.W.; Hotic, S.; Arking, R. Dietary restriction in Drosophila is dependent on mitochondrial efficiency and constrained by pre-existing extended longevity. Mech. Ageing Dev. 2007, 128, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, R.; Collins, S.; Trujillo, J.; McCredie, K.; Ahearn, M.; Tsai, S.; Metzgar, R.; Aulakh, G.; Ting, R.; Ruscetti, F.; et al. Characterization of the continuous, differentiating myeloid cell line (HL-60) from a patient with acute promyelocytic leukemia. Blood 1979, 54, 713–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merinas-Amo, T.; Tasset-Cuevas, I.; Díaz-Carretero, A.M.; Alonso-Moraga, Á.; Calahorro, F. In vivo and in vitro studies of the role of lyophilised blond Lager beer and some bioactive components in the modulation of degenerative processes. J. Funct. Foods 2016, 27, 274–294. [Google Scholar] [CrossRef]
- Husseini, G.A.; El-Fayoumi, R.I.; O’Neill, K.L.; Rapoport, N.Y.; Pitt, W.G. DNA damage induced by micellar-delivered doxorubicin and ultrasound: Comet assay study. Cancer Lett. 2000, 154, 211–216. [Google Scholar] [CrossRef]
- Olive, P.L.; Banáth, J.P. The comet assay: A method to measure DNA damage in individual cells. Nat. Protoc. 2006, 1, 23–29. [Google Scholar] [CrossRef]
- Prosperini, A.; Juan-García, A.; Font, G.; Ruiz, M. Beauvericin-induced cytotoxicity via ROS production and mitochondrial damage in Caco-2 cells. Toxicol. Lett. 2013, 222, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Mateo-Fernández, M.; Merinas-Amo, T.; Moreno-Millán, M.; Alonso-Moraga, Á.; Demyda-Peyrás, S. In vivo and in vitro genotoxic and epigenetic effects of two types of cola beverages and caffeine: A multi-assay approach. Bio. Med. Res. Int. 2016, 2016, 15. [Google Scholar]
- Kumaravel, T.S.; Vilhar, B.; Faux, S.P.; Jha, A.N. Comet Assay measurements: A perspective. Cell Biol. Toxicol. 2007, 25, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Deininger, P.L.; Moran, J.V.; Batzer, M.A.; Kazazian, H.H. Mobile elements and mammalian genome evolution. Curr. Opin. Genet. Dev. 2003, 13, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, M. DNA Hypomethylation, Cancer, the Immunodeficiency, Centromeric Region Instability, Facial Anomalies Syndrome and Chromosomal Rearrangements. J. Nutr. 2002, 132, 2424S–2429S. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.; Wevrick, R.; Fisher, R.B.; Ferguson-Smith, M.A.; Lin, C.C. Human centromeric DNAs. Qual. Life Res. 1997, 100, 291–304. [Google Scholar] [CrossRef]
- Weiner, A.M. SINEs and LINEs: The art of biting the hand that feeds you. Curr. Opin. Cell Biol. 2002, 14, 343–350. [Google Scholar] [CrossRef]
- Nikolaidis, G.; Raji, O.Y.; Markopoulou, S.; Gosney, J.R.; Bryan, J.; Warburton, C.; Walshaw, M.; Sheard, J.; Field, J.K.; Liloglou, T. DNA Methylation Biomarkers Offer Improved Diagnostic Efficiency in Lung Cancer. Cancer Res. 2012, 72, 5692–5701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liloglou, T.; Bediaga, N.G.; Brown, B.R.; Field, J.K.; Davies, M.P. Epigenetic biomarkers in lung cancer. Cancer Lett. 2014, 342, 200–212. [Google Scholar] [CrossRef]
- Weisenberger, D.J.; Campan, M.; Long, T.I.; Kim, M.; Woods, C.; Fiala, E.; Ehrlich, M.; Laird, P.W. Analysis of repetitive element DNA methylation by MethyLight. Nucleic Acids Res. 2005, 33, 6823–6836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Jiménez, M.; Campos-Sánchez, J.; Analla, M.; Muñoz-Serrano, A.; Alonso-Moraga, Á. Genotoxicity and anti-genotoxicity of some traditional medicinal herbs. Mutat. Res. Toxicol. Environ. Mutagen. 2005, 585, 147–155. [Google Scholar] [CrossRef]
- Gorjanovic, S.Z.; Novaković, M.M.; Vukosavljevic, P.V.; Pastor, F.T.; Tesevic, V.V.; Sužnjević, D.Z. Polarographic Assay Based on Hydrogen Peroxide Scavenging in Determination of Antioxidant Activity of Strong Alcohol Beverages. J. Agric. Food Chem. 2010, 58, 8400–8406. [Google Scholar] [CrossRef]
- Medina, E.; De Castro, A.; Romero, C.; Brenes, M. Comparison of the Concentrations of Phenolic Compounds in Olive Oils and Other Plant Oils: Correlation with Antimicrobial Activity. J. Agric. Food Chem. 2006, 54, 4954–4961. [Google Scholar] [CrossRef]
- De Keukeleire, D.; De Cooman, L.; Rong, H.; Heyerick, A.; Kalita, J.; Milligan, S.R. Functional properties of hop polyphenols. In Basic Life Sciences; Springer International Publishing: Berlin/Heidelberg, Germany, 1999; Volume 66, pp. 739–760. [Google Scholar]
- Halliwell, B.; Gutteridge, J. Cellular responses to oxidative stress: Adaptation, damage, repair, senescence and death. Free Radic. Biol. Med. 2007, 4, 187–267. [Google Scholar]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free. Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Yamanaka, N.; Oda, O.; Nagao, S. Green tea catechins such as (−)-epicatechin and (−)-epigallocatechin accelerate Cu2+ -induced low density lipoprotein oxidation in propagation phase. FEBS Lett. 1997, 401, 230–234. [Google Scholar] [CrossRef] [Green Version]
- Loru, D.; Incani, A.; Deiana, M.; Corona, G.; Atzeri, A.; Melis, M.; Rosa, A.; Dessì, M. Protective effect of hydroxytyrosol and tyrosol against oxidative stress in kidney cells. Toxicol. Ind. Health 2009, 25, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Liégeois, C.; Lermusieau, G.; Collin, S. Measuring Antioxidant Efficiency of Wort, Malt, and Hops against the 2,2‘-Azobis(2-amidinopropane) Dihydrochloride-Induced Oxidation of an Aqueous Dispersion of Linoleic Acid. J. Agric. Food Chem. 2000, 48, 1129–1134. [Google Scholar] [CrossRef] [PubMed]
- Tafulo, P.A.R.; Queirós, R.B.; Delerue-Matos, C.M.; Sales, M.G.F. Control and comparison of the antioxidant capacity of beers. Food Res. Int. 2010, 43, 1702–1709. [Google Scholar] [CrossRef] [Green Version]
- Granato, D.; Branco, G.F.; Faria, J.D.A.F.; Cruz, A.G. Characterization of Brazilian lager and brown ale beers based on color, phenolic compounds, and antioxidant activity using chemometrics. J. Sci. Food Agric. 2010, 91, 563–571. [Google Scholar] [CrossRef]
- Anter, J.; Romero-Jiménez, M.; Fernández-Bedmar, Z.; Villatoro-Pulido, M.; Analla, M.; Alonso-Moraga, Á.; Muñoz-Serrano, A. Antigenotoxicity, Cytotoxicity, and Apoptosis Induction by Apigenin, Bisabolol, and Protocatechuic Acid. J. Med. Food 2011, 14, 276–283. [Google Scholar] [CrossRef]
- Arimoto-Kobayashi, S.; Ishida, R.; Nakai, Y.; Idei, C.; Takata, J.; Takahashi, E.; Okamoto, K.; Negishi, T.; Konuma, T. Inhibitory Effects of Beer on Mutation in the Ames Test and DNA Adduct Formation in Mouse Organs Induced by 2-Amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP). Biol. Pharm. Bull. 2006, 29, 67–70. [Google Scholar] [CrossRef] [Green Version]
- Fleming, J.; Reveillaud, I.; Niedzwiecki, A. Role of oxidative stress in Drosophila aging. Mutat. Res. 1992, 275, 267–279. [Google Scholar] [CrossRef]
- Guarente, L.; Kenyon, C. Genetic pathways that regulate ageing in model organisms. Nat. Cell Biol. 2000, 408, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, S.A.; Carvalho, G.B.; Amador, A.; Phillips, A.M.; Hoxha, S.; Lizotte, K.J.; Ja, W.W. Quantifying Drosophila food intake: Comparative analysis of current methodology. Nat. Methods 2014, 11, 535–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riboli, E.; Cornée, J.; Macquart-Moulin, G.; Kaaks, R.; Casagrande, C.; Guyader, M. Cancer and Polyps of the Colorectum and Lifetime Consumption of Beer and Other Alcoholic Beverages. Am. J. Epidemiology 1991, 134, 157–166. [Google Scholar] [CrossRef]
- Kato, I.; Tominaga, S.; Ikari, A. A Case-Control Study of Male Colorectal Cancer in Aichi Prefecture, Japan: With Special Reference to Occupational Activity Level, Drinking Habits and Family History. Jpn. J. Cancer Res. 1990, 81, 115–121. [Google Scholar] [CrossRef]
- Swanson, C.A.; Wilbanks, G.D.; Twiggs, L.B.; Monet, R.; Berman, M.L.; Barrett, R.J.; Brintonl, L.A. Moderate Alcohol Consumption and the Risk of Endometrial Cancer. Epidemiology 1993, 4, 530–536. [Google Scholar] [CrossRef]
- Potter, J.D.; Sellers, T.A.; Folsom, A.R.; McGovern, P.G. Alcohol, beer, and lung cancer in postmenopausal women The iowa women’s health study. Ann. Epidemiol. 1992, 2, 587–595. [Google Scholar] [CrossRef]
- Hevia, D.; Mayo, J.C.; Quiros, I.; Sainz, R. Beer constituents inhibit prostate cancer cells proliferation. Eur. J. Cancer Suppl. 2008, 6, 142. [Google Scholar] [CrossRef]
- Babich, H.; Visioli, F. In vitro cytotoxicity to human cells in culture of some phenolics from olive oil. Il Farm. 2003, 58, 403–407. [Google Scholar] [CrossRef]
- Tyrrell, E.; Archer, R.; Skinner, G.; Singh, K.; Colston, K.; Driver, C. Structure elucidation and an investigation into the in vitro effects of hop acids on human cancer cells. Phytochem. Lett. 2010, 3, 17–23. [Google Scholar] [CrossRef]
- Honma, Y.; Tobe, H.; Makishima, M.; Yokoyama, A.; Okabe-Kado, J. Induction of differentiation of myelogenous leukemia cells by humulone, a bitter in the hop. Leuk. Res. 1998, 22, 605–610. [Google Scholar] [CrossRef]
- Salucci, S.; Burattini, S.; Curzi, D.; Buontempo, F.; Martelli, A.M.; Zappia, G.; Falcieri, E.; Battistelli, M. Antioxidants in the prevention of UVB-induced keratynocyte apoptosis. J. Photochem. Photobiol. B Biol. 2014, 141, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tobe, H.; Kubota, M.; Yamaguchi, M.; Kocha, T.; Aoyagi, T. Apoptosis to HL-60 by Humulone. Biosci. Biotechnol. Biochem. 1997, 61, 1027–1029. [Google Scholar] [CrossRef] [PubMed]
- Poe, B.S.; O’Neill, K.L. Caffeine modulates heat shock induced apoptosis in the human promyelocytic leukemia cell line HL-60. Cancer Lett. 1997, 121, 1–6. [Google Scholar] [CrossRef]
- Fabiani, R.; Rosignoli, P.; De Bartolomeo, A.; Fuccelli, R.; Morozzi, G. Genotoxicity of alkene epoxides in human peripheral blood mononuclear cells and HL60 leukaemia cells evaluated with the comet assay. Mutat. Res. Toxicol. Environ. Mutagen. 2012, 747, 1–6. [Google Scholar] [CrossRef]
- Fairbairn, D.W.; Walburger, D.K.; Fairbairn, J.J.; O’Neill, K.L. Key morphologic changes and DNA strand breaks in human lymphoid cells: Discriminating apoptosis from necrosis. Scanning 2006, 18, 407–416. [Google Scholar] [CrossRef]
- Fairbairn, D.W.; Olive, P.L.; O’Neill, K.L. The comet assay: A comprehensive review. Mutat. Res. Genet. Toxicol. 1995, 339, 37–59. [Google Scholar] [CrossRef]
- Herman, J.G.; Baylin, S.B. Gene Silencing in Cancer in Association with Promoter Hypermethylation. New Engl. J. Med. 2003, 349, 2042–2054. [Google Scholar] [CrossRef]
- Esteller, M. DNA methylation and cancer therapy: New developments and expectations. Curr. Opin. Oncol. 2005, 17, 55–60. [Google Scholar] [CrossRef]
- Román-Gómez, J.; Jiménez-Velasco, A.; Agirre, X.; Castillejo, J.A.; Navarro, G.; José-Enériz, E.S.; Garate, L.; Cordeu, L.; Cervantes, F.; Prosper, F.; et al. Repetitive DNA hypomethylation in the advanced phase of chronic myeloid leukemia. Leuk. Res. 2008, 32, 487–490. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reaction. ID | GenBank Number | Amplicon Start End | Forward Primer Sequence 5′ to 3′ (N) | Reverse Primer Sequence 5′ to 3′ (N) | GC Content (%) Forward Reverse | ||
|---|---|---|---|---|---|---|---|
| Alu C4 | Consensus Sequence | 1 | 98 | GGTTAGGTA TAGTGGTTT AT ATTTGTA ATTTTAGTA (36) | ATTAACT AA ACT AATCTT AAACTCCTA ACCTCA (33) | 25 | 27.3 |
| Alu M1 | Y07755 | 5059 | 5164 | ATTATGTTA GTTAGGATG GTTTCGATT TT (29) | CAATCGACC GAACGCGA (17) | 27.6 | 58.8 |
| LINE-1 | X52235 | 251 | 331 | GGACGT ATT TGGAAAATC GGG (21) | AATCTCGCG AT ACGCCGT T (19) | 47.6 | 52.6 |
| Sat-α | M38468 | 139 | 260 | TGATGGAGT ATTTTTAAA AT ATACGTT TTGTAGT (34) | AATTCTAAA AAT ATTCCT CTTCAATTA CGTAAA (33) | 23.5 | 21.2 |
| Clones per Wing (No. of Spots) (1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Compound | Number of Wings | Small Single Clones (1–2 Cells) m = 2 | Large Simple Clones (>2 Cells) m = 5 | Twin Clones m = 5 | Total Clones m = 2 | Mann –Whitney U test (2) | Inhibition Percentage (%) (3) | ||
| H2O | 38 | 0.157 | (6) | 0 | 0 | 0.157 | (6) | ||
| H2O2 | 36 | 0.305 | (11) | 0.083 (3) | 0 | 0.388 | (14) + | ||
| SIMPLE TREATMENT | |||||||||
| LBAB (mg/mL) | |||||||||
| 3.125 | 39 | 0.154 | (6) | 0 | 0 | 0.154 | (6) i | ∆ | |
| 50 | 38 | 0.263 | (10) | 0.026 (1) | 0 | 0.289 | (11) i | ∆ | |
| LSAB (mg/mL) | |||||||||
| 3.125 | 42 | 0.190 | (8) | 0.047 (2) | 0 | 0.238 | (10) i | ∆ | |
| 50 | 44 | 0.159 | (7) | 0.091 (4) | 0.022 (1) | 0.273 | (12) i | ∆ | |
| Tyrosol (mM) | |||||||||
| 0.127 | 40 | 0.263 | (10) | 0.105 (4) | 0.026 (1) | 0.395 | (15) i | ∆ | |
| 2.026 | 40 | 0.175 | (7) | 0 | 0 | 0.175 | (7) i | ∆ | |
| Iso-alpha humulone (mM) | |||||||||
| 2.414 | 40 | 0.150 | (6) | 0 | 0 | 0.150 | (6) i | ∆ | |
| 77.25 | 40 | 0.125 | (5) | 0.125 (5) | 0 | 0.250 | (10) i | ∆ | |
| COMBINED TREATMENT WITH H2O2 (0.12 M) | |||||||||
| LBAB (mg/mL) | |||||||||
| 3.125 | 40 | 0.275 | (11) | 0 | 0.025 (1) | 0.300 | (12) γ | 22.68 | |
| 50 | 42 | 0.095 | (4) | 0.071 (3) | 0 | 0.166 | (7) γ | 57.21 | |
| LSAB (mg/mL) | |||||||||
| 3.125 | 40 | 0.175 | (7) | 0.025 (1) | 0 | 0.200 | (8) γ | 48.45 | |
| 50 | 44 | 0.159 | (7) | 0 | 0 | 0.159 | (7) γ | 59.02 | |
| Tyrosol (mM) | |||||||||
| 0.127 | 40 | 0.325 | (13) | 0.075 (3) | 0 | 0.400 | (16) λ | ∆ | −3.09 |
| 2.026 | 40 | 0.325 | (13) | 0.100 (4) | 0 | 0.425 | (17) λ | ∆ | −9.53 |
| Iso-alpha humulone (mM) | |||||||||
| 2.414 | 38 | 0.342 | (13) | 0 | 0 | 0.342 | (13) γ | 11.85 | |
| 77.25 | 38 | 0.289 | (11) | 0 | 0 | 0.289 | (11) γ | 25.51 | |
| Compound | Concentration | Mean Lifespan (1) | Mean Healthspan (1) | ||
|---|---|---|---|---|---|
| (Days) | (Days) | ||||
| LBAB | Control | 60.104 | 33.229 | ||
| 3.125 mg/mL | 66.176 | * | 42.500 | ** | |
| 6.25 mg/mL | 66.149 | * | 36.378 | ns | |
| 12.5 mg/mL | 66.688 | * | 31.750 | ns | |
| 25 mg/mL | 59.474 | ns | 34.634 | ns | |
| 50 mg/mL | 58.600 | ns | 31.756 | ns | |
| Control | 47.242 | 19.769 | |||
| 3.125 mg/mL | 47.636 | ns | 34.728 | *** | |
| LSAB | 6.25 mg/mL | 34.323 | *** | 16.348 | ** |
| 25 mg/mL | 35.918 | *** | 17.313 | ns | |
| 50 mg/mL | 35.607 | *** | 24.671 | ** | |
| Tyrosol | Control | 60.333 | 33.154 | ||
| 0.127 mM | 58.664 | ns | 20.000 | ** | |
| 0.253 mM | 63.046 | ns | 47.923 | *** | |
| 0.506 mM | 62.018 | ns | 43.857 | *** | |
| 1.013 mM | 56.273 | ns | 38.364 | ns | |
| 2.026 mM | 58.588 | ns | 38.083 | ns | |
| Iso-alpha humulone | Control | 79.853 | 50.156 | ||
| 2.414 mM | 66.779 | *** | 40.510 | ** | |
| 9.65 mM | 67.702 | *** | 43.529 | ** | |
| 19.312 mM | 77.522 | ns | 54.035 | ns | |
| 38.625 mM | 64.205 | *** | 41.017 | ** | |
| 77.25 mM | 49.824 | *** | 30.323 | *** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Merinas-Amo, T.; Merinas-Amo, R.; Font, R.; del Río Celestino, M.; Alonso-Moraga, Á. Toxicological and Epigenetic Studies of Two Types of Ale Beer, Tyrosol and Iso-Alpha Humulone. Processes 2021, 9, 485. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9030485
Merinas-Amo T, Merinas-Amo R, Font R, del Río Celestino M, Alonso-Moraga Á. Toxicological and Epigenetic Studies of Two Types of Ale Beer, Tyrosol and Iso-Alpha Humulone. Processes. 2021; 9(3):485. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9030485
Chicago/Turabian StyleMerinas-Amo, Tania, Rocío Merinas-Amo, Rafael Font, Mercedes del Río Celestino, and Ángeles Alonso-Moraga. 2021. "Toxicological and Epigenetic Studies of Two Types of Ale Beer, Tyrosol and Iso-Alpha Humulone" Processes 9, no. 3: 485. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9030485