
Impact of Indigenous Non-Saccharomyces Yeasts Isolated from Madeira Island Vineyards on the Formation of Ethyl Carbamate in the Aging of Fortified Wines

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Samples
2.1.1. Yeast Isolation and Identification
2.1.2. Inoculation and Wine Production
2.1.3. Alcoholic Fortification and Wine Accelerated Aging
2.2. Determination of Ethyl Carbamate, Urea, Citrulline and Arginine
3. Results and Discussion
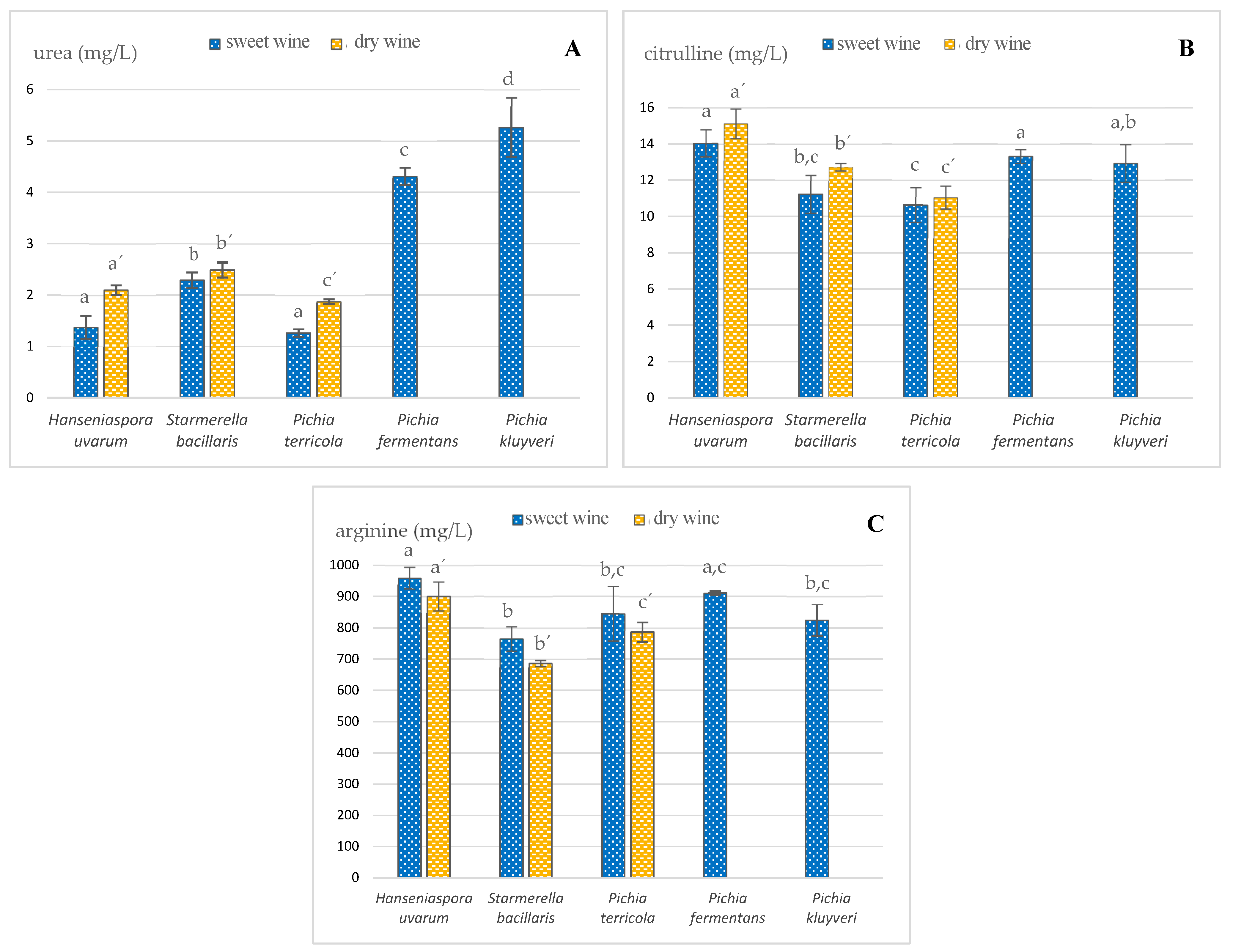
3.1. Development of Ethyl Carbamate Precursors
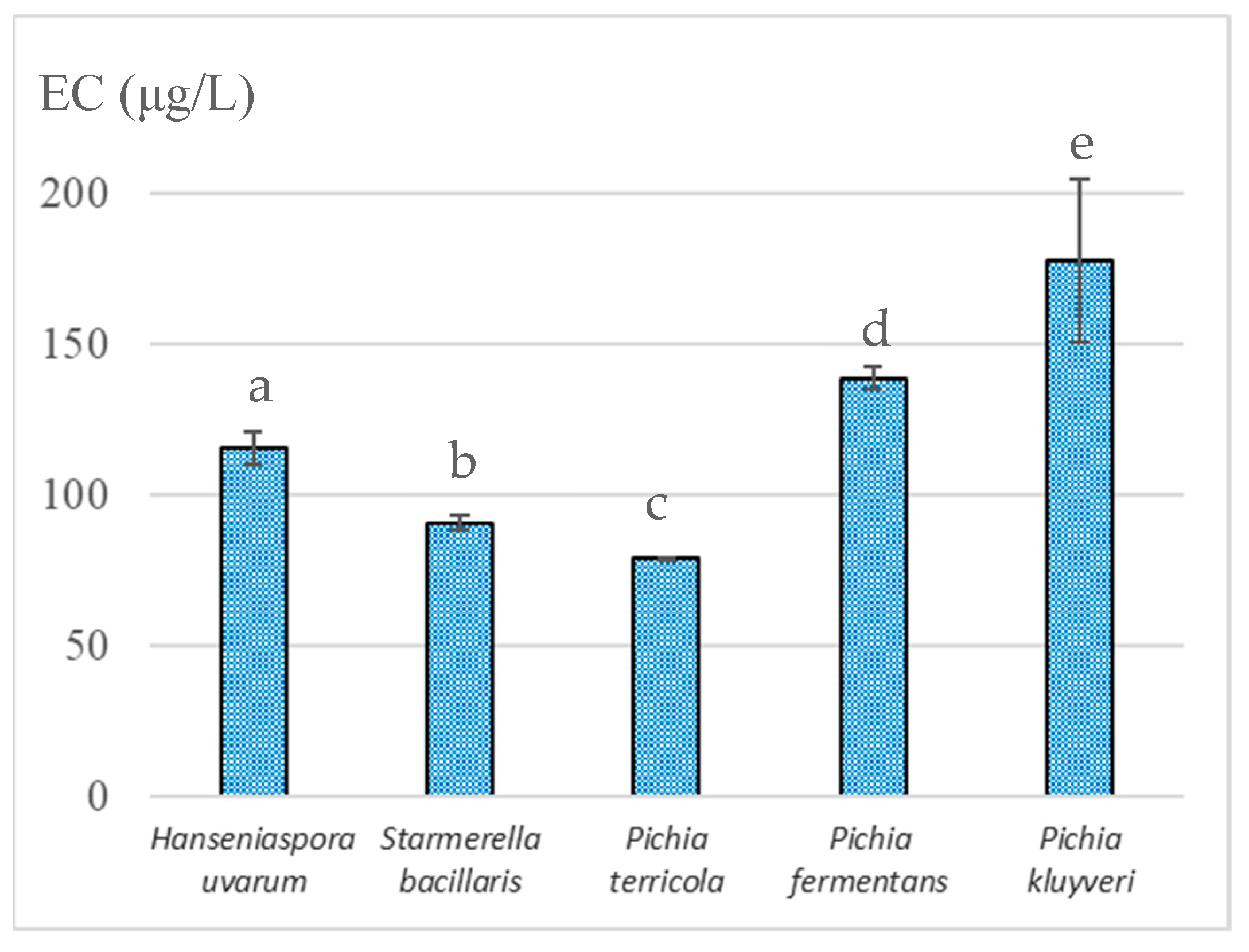
3.2. Ethyl Carbamate Formation Potential with Wine Aging
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Comitini, F.; Capece, A.; Ciani, M.; Romano, P. New insights on the use of wine yeasts. Curr. Opin. Food Sci. 2017, 13, 44–49. [Google Scholar] [CrossRef]
- Capozzi, V.; Di Toro, M.R.; Grieco, F.; Michelotti, V.; Salma, M.; Lamontanara, A.; Russo, P.; Orrù, L.; Alexandre, H.; Spano, G. Viable but Not Culturable (VBNC) state of Brettanomyces bruxellensis in wine: New insights on molecular basis of VBNC behaviour using a transcriptomic approach. Food Microbiol. 2016, 59, 196–204. [Google Scholar] [CrossRef]
- Zarraonaindia, I.; Owens, S.M.; Weisenhorn, P.; West, K.; Hampton-Marcell, J.; Lax, S.; Bokulich, N.A.; Mills, D.A.; Martin, G.; Taghavi, S.; et al. The Soil Microbiome Influences Grapevine-Associated Microbiota. mBio 2015, 6, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and Future of Non-Saccharomyces Yeasts: From Spoilage Microorganisms to Biotechnological Tools for Improving Wine Aroma Complexity. Front. Microbiol. 2016, 7, 7–20. [Google Scholar] [CrossRef] [Green Version]
- IARC. Alcohol Consumption and Ethyl Carbamate; IARC Press, International Agency for Research on Cancer: Lyons, France, 2010; Volume 96. [Google Scholar]
- Conacher, H.; Page, B. Ethyl carbamate in alcoholic beverages: A canadian case history. In Proceedings of Euro Food Tox II; European Society of Toxicology: Schwerzenbach, Switzerland, 1986; pp. 237–242. [Google Scholar]
- Government of Canada. Health Canada’s Maximum Levels for Chemical Contaminants in Foods. Available online: https://www.canada.ca/en/health-canada/corporate/contact-us.html (accessed on 17 March 2021).
- Jiao, Z.; Dong, Y.; Chen, Q. Ethyl Carbamate in Fermented Beverages: Presence, Analytical Chemistry, Formation Mechanism, and Mitigation Proposals. Compr. Rev. Food Sci. Food Saf. 2014, 13, 611–626. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.H. Compartmental and regulatory mechanisms in the arginine pathways of Neurospora crassa and Saccharomyces cerevisiae. Microbiol. Rev. 1986, 50, 280–313. [Google Scholar] [CrossRef] [PubMed]
- Butzke, C.E.; Bisson, L.F. Ethyl Carbamate Preventative Action Manual; US Food and Drug Administration, Center for food safety and Applied nutrition: Washington, DC, USA, 1997.
- Azevedo, Z.; Couto, J.A.; Hogg, T. Citrulline as the main precursor of ethyl carbamate in model fortified wines inoculated with Lactobacillus hilgardii: A marker of the levels in a spoiled fortified wine. Lett. Appl. Microbiol. 2002, 34, 32–36. [Google Scholar] [CrossRef]
- Kodama, S.; Suzuki, T.; Fujinawa, S.; Teja, P.; Yotsuzuka, F. Urea Contribution to Ethyl Carbamate Formation in Commercial Wines During Storage. Am. J. Enol. Vitic. 1994, 45, 17–24. [Google Scholar]
- Stevens, D.F.; Ough, C.S. Ethyl Carbamate Formation: Reaction of Urea and Citrulline with Ethanol in Wine Under Low to Normal Temperature Conditions. Am. J. Enol. Vitic. 1993, 44, 309–312. [Google Scholar]
- An, D.; Ough, C.S. Urea Excretion and Uptake by Wine Yeasts as Affected by Various Factors. Am. J. Enol. Vitic. 1993, 44, 35–40. [Google Scholar]
- Henschke, P.A.; Ough, C.S. Urea Accumulation in Fermenting Grape Juice. Am. J. Enol. Vitic. 1991, 42, 317–321. [Google Scholar]
- Coulon, J.; Husnik, J.I.; Inglis, D.L.; van der Merwe, G.K.; Lonvaud, A.; Erasmus, D.J.; van Vuuren, H.J.J. Metabolic Engineering of Saccharomyces cerevisiae to Minimize the Production of Ethyl Carbamate in Wine. Am. J. Enol. Vitic. 2006, 57, 113. [Google Scholar]
- Pereira, V.; Pereira, A.C.; Marques, J.C. 13-Emerging Trends in Fortified Wines: A Scientific Perspective. In Alcoholic Beverages; Grumezescu, A.M., Holban, A.M., Eds.; Woodhead Publishing: Amsterdam, The Netherlands, 2019; pp. 419–470. [Google Scholar]
- Perestrelo, R.; Silva, C.; Pereira, J.; Câmara, J.S. Wines: Madeira, Port and Sherry Fortified Wines–The Sui Generis and Notable Peculiarities. Major Differences and Chemical Patterns. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: Oxford, UK, 2016; pp. 534–555. [Google Scholar]
- Elliott, T. Viticulture. In The Wines of Madeira: An Indispensable Guide to the Wines, Grapes and Producers, 1st ed.; Trevor Elliott Publishing: Hampshire, UK, 2010. [Google Scholar]
- Jackson, R.S. Chemical Constituents of Grapes and Wine. Wine Science, 2nd ed.; Academic Press: San Diego, CA, USA, 2000. [Google Scholar]
- Esteve-Zarzoso, B.; Belloch, C.; Uruburu, F.; Querol, A. Identification of yeasts by RFLP analysis of the 5.8S rRNA gene and the two ribosomal internal transcribed spacers. Int. J. Syst. Evol. Microbiol. 1999, 49, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Benito, S.; Calderón, F.; Benito, Á. The Combined Use of Schizosaccharomyces pombe and Lachancea thermotolerans—Effect on the Anthocyanin Wine Composition. Molecules 2017, 22, 739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIV. Alcoholic Strength by Volume, OIV-MA-AS312-01B, Compendium of International Methods of Analysis. 2009. Resolution Oeno 377/2009. Available online: https://www.oiv.int/public/medias/2490/oiv-ma-as312-01b.pdf (accessed on 28 April 2021).
- Pereira, V.; Cacho, J.; Marques, J.C. Volatile profile of Madeira wines submitted to traditional accelerated ageing. Food Chem. 2014, 162, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Pereira, V.; Santos, M.; Cacho, J.; Marques, J.C. Assessment of the development of browning, antioxidant activity and volatile organic compounds in thermally processed sugar model wines. LWT Food Sci. Technol. 2017, 75, 719–726. [Google Scholar] [CrossRef]
- Leça, J.M.; Pereira, V.; Pereira, A.C.; Marques, J.C. A Sensitive Method for the Rapid Determination of Underivatized Ethyl Carbamate in Fortified Wine by Liquid Chromatography-Electrospray Tandem Mass Spectrometry. Food Anal. Methods 2017, 11, 327–333. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, G.; Zhang, Y.; Gao, Q.; Wang, D.; Liu, H. Simultaneous Determination of Ethyl Carbamate and Urea in Alcoholic Beverages by High-Performance Liquid Chromatography Coupled with Fluorescence Detection. J. Agric. Food Chem. 2014, 62, 2797–2802. [Google Scholar] [CrossRef] [PubMed]
- Pereira, V.; Pontes, M.; Câmara, J.S.; Marques, J.C. Simultaneous analysis of free amino acids and biogenic amines in honey and wine samples using in loop orthophthalaldeyde derivatization procedure. J. Chromatogr. A 2008, 1189, 435–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boeira, L.S.; Pereira, C.N.; Daudt, C.E. Arginine and urea evolution during musts fermentation of cv gewürztraminer from different nitrogen fertilizer levels in the soil. Ciência Rural 1995, 25, 295–298. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, F.F.; Bisson, L.F. Amino Acid Utilization and Urea Formation During Vinification Fermentations. Am. J. Enol. Vitic. 1991, 42, 199. [Google Scholar]
- Arena, M.E.; Saguir, F.M.; Manca de Nadra, M.C. Arginine, citrulline and ornithine metabolism by lactic acid bacteria from wine. Int. J. Food Microbiol. 1999, 52, 155–161. [Google Scholar] [CrossRef]
- Mira de Orduña, R.; Liu, S.-Q.; Patchett, M.L.; Pilone, G.J. Ethyl carbamate precursor citrulline formation from arginine degradation by malolactic wine lactic acid bacteria. FEMS Microbiol. Lett. 2000, 183, 31–35. [Google Scholar] [CrossRef]
- Terrade, N.; Mira de Orduña, R. Impact of winemaking practices on arginine and citrulline metabolism during and after malolactic fermentation. J. Appl. Microbiol. 2006, 101, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Pereira, V.; Pereira, A.C.; Trujillo, J.P.P.; Cacho, J.; Marques, J.C. Amino Acids and Biogenic Amines Evolution during the Estufagem of Fortified Wines. J. Chem. 2015, 2015, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Bisson, L.F. Influence of nitrogen on yeast and fermentation of grapes. In Proceedings of the International Symposium on Nitrogen in Grapes and Wine Seattle, Washington, DC, USA, 18–19 June 1991; pp. 78–89. [Google Scholar]
- Valero, E.; Millán, C.; Ortega, J.M.; Mauricio, J.C. Concentration of amino acids in wine after the end of fermentation by Saccharomyces cerevisiae strains. J. Sci. Food Agric. 2003, 83, 830–835. [Google Scholar] [CrossRef]
- Englezos, V.; Giacosa, S.; Rantsiou, K.; Rolle, L.; Cocolin, L. Starmerella bacillaris in winemaking: Opportunities and risks. Curr. Opin. Food Sci. 2017, 17, 30–35. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Yeast | Sweet Wines (g/L) | Dry Wines (g/L) |
|---|---|---|
| Hanseniaspora uvarum | 1054 ± 3 | 1020 ± 5 |
| Starmerella bacillari | 1059 ± 1 | 1021 ± 1 |
| Pichia terricola | 1052 ± 1 | 1026 ± 1 |
| Pichia kluyveri | 1051 ± 1 | - |
| Pichia terricola | 1045 ± 1 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leça, J.M.; Pereira, V.; Miranda, A.; Vilchez, J.L.; Malfeito-Ferreira, M.; Marques, J.C. Impact of Indigenous Non-Saccharomyces Yeasts Isolated from Madeira Island Vineyards on the Formation of Ethyl Carbamate in the Aging of Fortified Wines. Processes 2021, 9, 799. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9050799
Leça JM, Pereira V, Miranda A, Vilchez JL, Malfeito-Ferreira M, Marques JC. Impact of Indigenous Non-Saccharomyces Yeasts Isolated from Madeira Island Vineyards on the Formation of Ethyl Carbamate in the Aging of Fortified Wines. Processes. 2021; 9(5):799. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9050799
Chicago/Turabian StyleLeça, João Micael, Vanda Pereira, Andreia Miranda, José Luis Vilchez, Manuel Malfeito-Ferreira, and José Carlos Marques. 2021. "Impact of Indigenous Non-Saccharomyces Yeasts Isolated from Madeira Island Vineyards on the Formation of Ethyl Carbamate in the Aging of Fortified Wines" Processes 9, no. 5: 799. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9050799