
Bacterial Succession Pattern during the Fermentation Process in Whole-Plant Corn Silage Processed in Different Geographical Areas of Northern China

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Silage Preparation
2.2. Analyses
2.2.1. Physicochemical Analysis
2.2.2. Microbial Analysis
2.3. Statistical Analyses
3. Results
3.1. Fermentation Quality and Microbial Counts
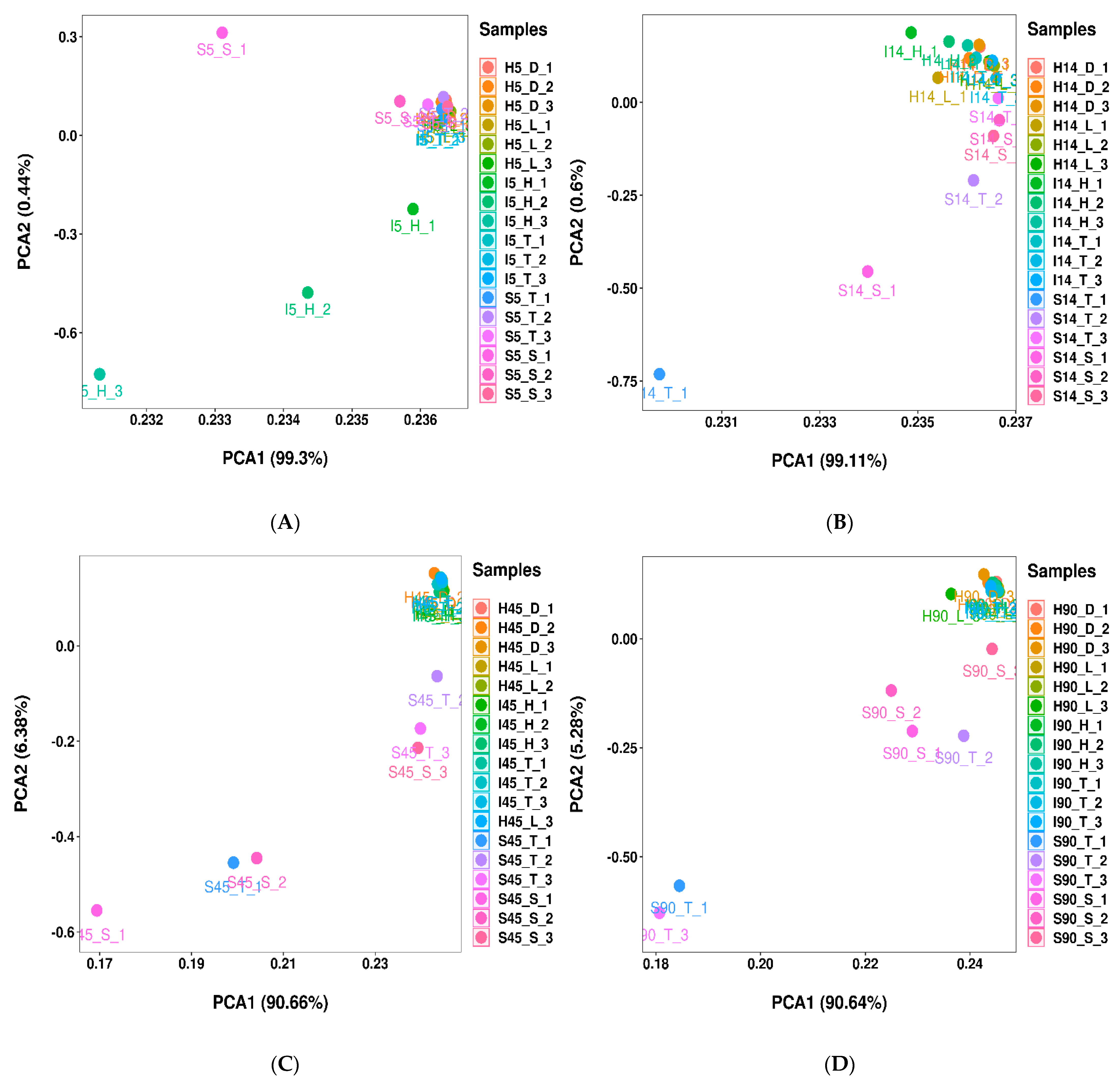
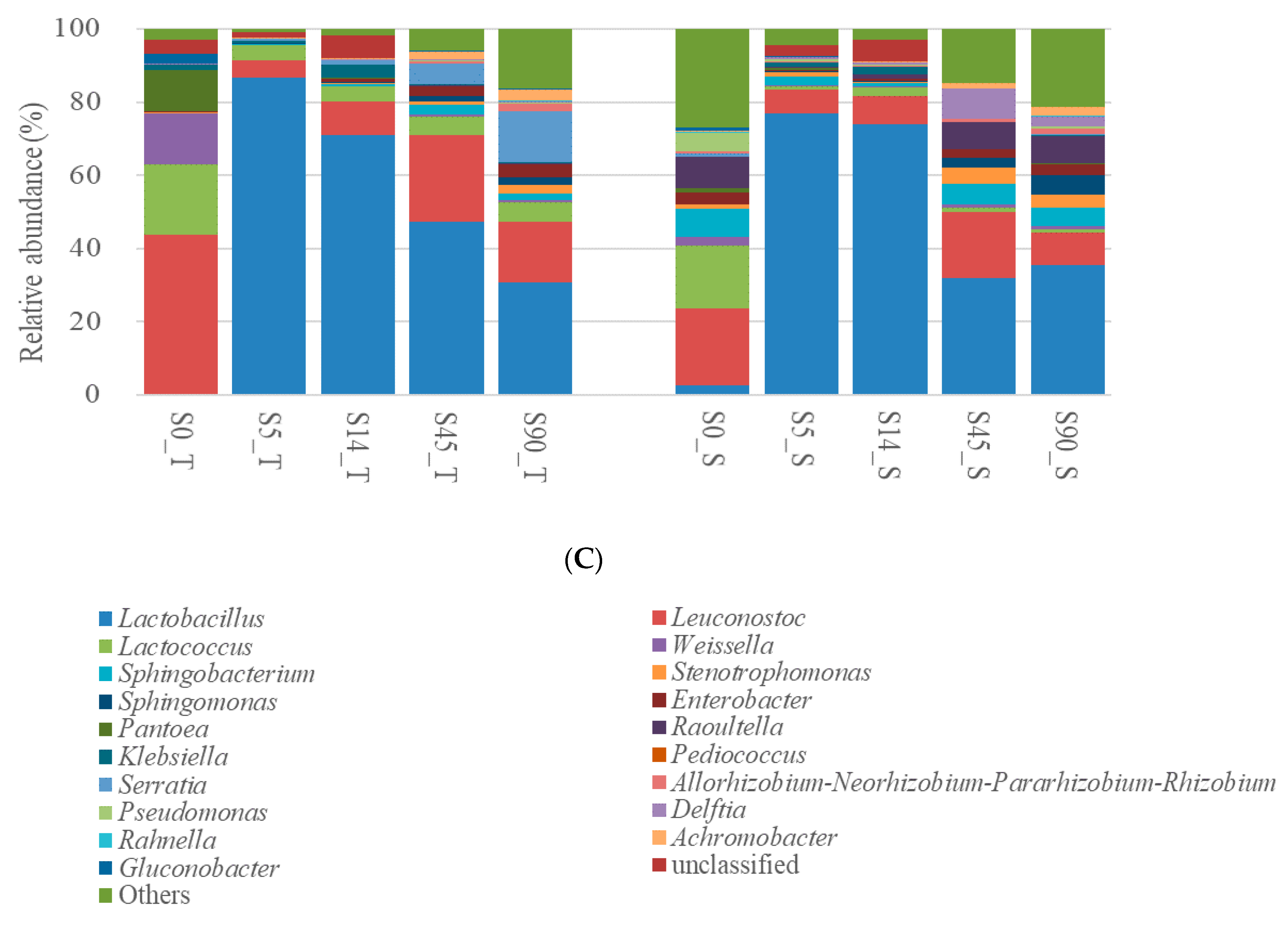
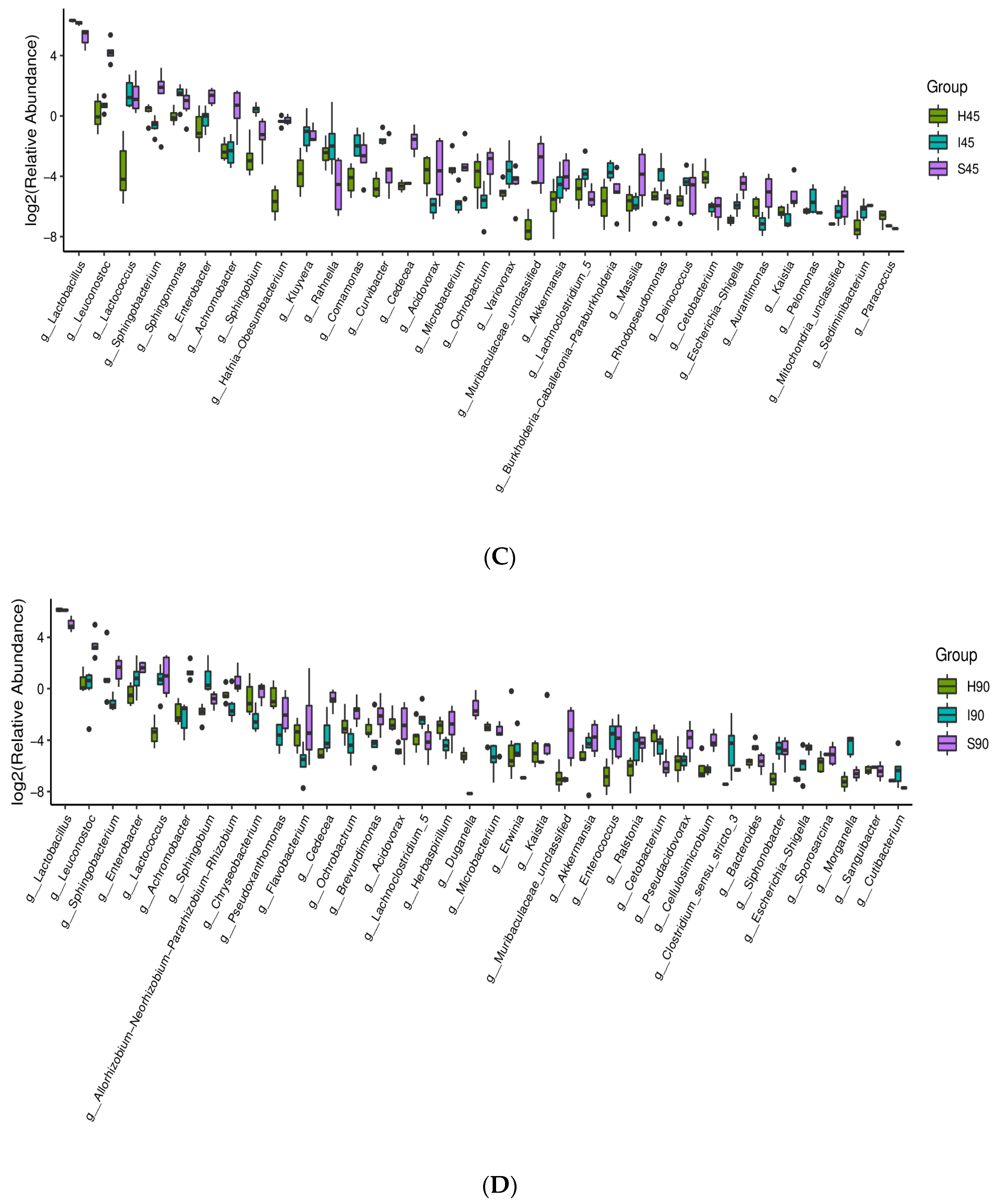
3.2. Bacterial Communities
3.3. Correlation between Fermentation Quality and Bacterial Genera
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grant, R.; Ferraretto, L. Silage review: Silage feeding management: Silage characteristics and dairy cow feeding behavior. J. Dairy Sci. 2018, 101, 4111–4121. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Yan, Y.; Li, X.; Li, X.; Shuai, Y.; Feng, G.; Ran, Q.; Cai, Y.; Li, Y.; Zhang, X. Microbial communities and natural fermentation of corn silages prepared with farm bunker-silo in Southwest China. Bioresour. Technol. 2018, 265, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, J.M.; Rinne, M. Highlights of progress in silage conservation and future perspectives. Grass Forage Sci. 2017, 73, 40–52. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, X.; Gu, Q.; Liang, M.; Mu, S.; Zhou, B.; Huang, F.; Lin, B.; Zou, C. Analysis of the correlation between bacteria and fungi in sugarcane tops silage prior to and after aerobic exposure. Bioresour. Technol. 2019, 291, 121835. [Google Scholar] [CrossRef]
- Kaiser, A.G.; Piltz, J.W.; Burns, H.M.; Griffiths, N.W. TOPFODDER Sucessful Silage, 2nd ed.; Dairt Australia and New South Wales Department of Primary Industries: Orange, NSW, Australia, 2004. [Google Scholar]
- Ferraretto, L.; Shaver, R.; Luck, B. Silage review: Recent advances and future technologies for whole-plant and fractionated corn silage harvesting. J. Dairy Sci. 2018, 101, 3937–3951. [Google Scholar] [CrossRef]
- Buxton, D.R.; Muck, R.E.; Harrison, J.H. Silage Science and Technology; American Society of Agronomy: Madison, WI, USA, 2003. [Google Scholar]
- Khan, N.A.; Yu, P.; Ali, M.; Cone, J.W.; Hendriks, W.H. Nutritive value of maize silage in relation to dairy cow performance and milk quality. J. Sci. Food Agric. 2015, 95, 238–252. [Google Scholar] [CrossRef]
- Sun, L.; Bai, C.; Xu, H.; Na, N.; Jiang, Y.; Yin, G.; Liu, S.; Xue, Y. Succession of Bacterial Community During the Initial Aerobic, Intense Fermentation, and Stable Phases of Whole-Plant Corn Silages Treated with Lactic Acid Bacteria Suspensions Prepared From Other Silages. Front. Microbiol. 2021, 12, 591. [Google Scholar] [CrossRef]
- Jin, L.; Dunière, L.; Lynch, J.P.; Zaheer, R.; Turkington, K.; Blackshaw, R.E.; Lupwayi, N.Z.; O’Donovan, J.T.; Harker, K.N.; McAllister, T.; et al. Impact of ferulic acid esterase-producing lactobacilli and fibrolytic enzymes on ensiling and digestion kinetics of mixed small-grain silage. Grass Forage Sci. 2016, 72, 80–92. [Google Scholar] [CrossRef]
- Xu, D.; Wang, N.; Rinne, M.; Ke, W.; Weinberg, Z.G.; Da, M.; Bai, J.; Zhang, Y.; Li, F.; Guo, X. The bacterial community and metabolome dynamics and their interactions modulate fermentation process of whole crop corn silage prepared with or without inoculants. Microb. Biotechnol. 2021, 14, 561–576. [Google Scholar] [CrossRef]
- Gharechahi, J.; Kharazian, Z.A.; Sarikhan, S.; Jouzani, G.S.; Aghdasi, M.; Salekdeh, G.H. The dynamics of the bacterial communities developed in maize silage. Microb. Biotechnol. 2017, 10, 1663–1676. [Google Scholar] [CrossRef]
- Guan, H.; Shuai, Y.; Yan, Y.; Ran, Q.; Wang, X.; Li, D.; Cai, Y.; Zhang, X. Microbial Community and Fermentation Dynamics of Corn Silage Prepared with Heat-Resistant Lactic Acid Bacteria in a Hot Environment. Microorganisms 2020, 8, 719. [Google Scholar] [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela, S. Microbiome dynamics during ensiling of corn with and without Lactobacillus plantarum inoculant. Appl. Microbiol. Biotechnol. 2018, 102, 4025–4037. [Google Scholar] [CrossRef]
- Jiang, F.-G.; Cheng, H.-J.; Liu, D.; Wei, C.; An, W.-J.; Wang, Y.-F.; Sun, H.-T.; Song, E.-L. Treatment of Whole-Plant Corn Silage with Lactic Acid Bacteria and Organic Acid Enhances Quality by Elevating Acid Content, Reducing pH, and Inhibiting Undesirable Microorganisms. Front. Microbiol. 2020, 11, 593088. [Google Scholar] [CrossRef]
- Wang, M.; Franco, M.; Cai, Y.; Yu, Z. Dynamics of fermentation profile and bacterial community of silage prepared with alfalfa, whole-plant corn and their mixture. Anim. Feed. Sci. Technol. 2020, 270, 114702. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Meng, Q.; Zhou, Z.; Wu, H. A mixture of potassium sorbate and sodium benzoate improved fermentation quality of whole-plant corn silage by shifting bacterial communities. J. Appl. Microbiol. 2020, 128, 1312–1323. [Google Scholar] [CrossRef]
- Ni, Y.; Wang, M. Spatiotemporal evolution of China’s silage corn industry and the factors driving its develop-ment. Pratacult. Sci. 2019, 7, 1915–1924. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, X.; Zhao, M.; Yu, Z. Isolating and evaluating lactic acid bacteria strains for effectiveness of Leymus chinensis silage fermentation. Lett. Appl. Microbiol. 2014, 59, 391–397. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Cai, Y. Identification and Characterization of Enterococcus Species Isolated from Forage Crops and Their Influence on Silage Fermentation. J. Dairy Sci. 1999, 82, 2466–2471. [Google Scholar] [CrossRef]
- Logue, J.B.; Stedmon, C.A.; Kellerman, A.M.; Nielsen, N.J.; Andersson, A.F.; Laudon, H.; Lindström, E.; Kritzberg, E.S. Experimental insights into the importance of aquatic bacterial community composition to the degradation of dissolved organic matter. ISME J. 2016, 10, 533–545. [Google Scholar] [CrossRef]
- McDonald, P.; Henderson, A.R.; Heron, S.J.E. The Biochemistry of Silage, 2nd ed.; Cambrian Printers, Ltd.: Merlow, UK; Bucks, UK; Aberystwyth, UK; Wales, UK, 1991. [Google Scholar]
- Ferrero, F.; Tabacco, E.; Piano, S.; Casale, M.; Borreani, G. Temperature during conservation in laboratory silos affects fermentation profile and aerobic stability of corn silage treated with Lactobacillus buchneri, Lactobacillus hilgardii, and their combination. J. Dairy Sci. 2021, 104, 1696–1713. [Google Scholar] [CrossRef]
- McGarvey, J.; Franco, R.; Palumbo, J.; Hnasko, R.; Stanker, L.; Mitloehner, F. Bacterial population dynamics during the ensiling of Medicago sativa (alfalfa) and subsequent exposure to air. J. Appl. Microbiol. 2013, 114, 1661–1670. [Google Scholar] [CrossRef]
- Xu, D.; Ding, W.; Ke, W.; Li, F.; Zhang, P.; Guo, X. Modulation of Metabolome and Bacterial Community in Whole Crop Corn Silage by Inoculating Homofermentative Lactobacillus plantarum and Heterofermentative Lactobacillus buchneri. Front. Microbiol. 2019, 9, 3299. [Google Scholar] [CrossRef] [Green Version]
- Drouin, P.; Tremblay, J.; Chaucheyras-Durand, F. Dynamic Succession of Microbiota during Ensiling of Whole Plant Corn Following Inoculation with Lactobacillus buchneri and Lactobacillus hilgardii Alone or in Combination. Microorganisms 2019, 7, 595. [Google Scholar] [CrossRef] [Green Version]
- Drouin, P.; Tremblay, J.; Renaud, J.; Apper, E. Microbiota succession during aerobic stability of maize silage inoculated with Lentilactobacillus buchneri NCIMB 40788 and Lentilactobacillus hilgardii CNCM-I. MicrobiologyOpen 2021, 10, e1153. [Google Scholar] [CrossRef]
- Wang, C.; Sun, L.; Xu, H.; Na, N.; Yin, G.; Liu, S.; Jiang, Y.; Xue, Y. Microbial Communities, Metabolites, Fermentation Quality and Aerobic Stability of Whole-Plant Corn Silage Collected from Family Farms in Desert Steppe of North China. Processes 2021, 9, 784. [Google Scholar] [CrossRef]
- Cai, Y.; Benno, Y.; Ogawa, M.; Ohmomo, S.; Kumai, S.; Nakase, T. Influence of Lactobacillus spp. from an Inoculant and of Weissella and Leuconostoc spp. from Forage Crops on Silage Fermentation. Appl. Environ. Microbiol. 1998, 64, 2982–2987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ávila, C.; Carvalho, B. Silage fermentation—Updates focusing on the performance of micro-organisms. J. Appl. Microbiol. 2019, 128, 966–984. [Google Scholar] [CrossRef] [Green Version]
- da Silva, E.; Smith, M.; Savage, R.; Polukis, S.; Drouin, P.; Kung, L. Effects of Lactobacillus hilgardii 4785 and Lactobacillus buchneri 40788 on the bacterial community, fermentation and aerobic stability of high-moisture corn silage. J. Appl. Microbiol. 2021, 130, 1481–1493. [Google Scholar] [CrossRef]
- Liu, B.; Huan, H.; Gu, H.; Xu, N.; Shen, Q.; Ding, C. Dynamics of a microbial community during ensiling and upon aerobic exposure in lactic acid bacteria inoculation-treated and untreated barley silages. Bioresour. Technol. 2019, 273, 212–219. [Google Scholar] [CrossRef]
- Ogunade, I.; Jiang, Y.; Cervantes, A.P.; Kim, D.; Oliveira, A.; Vyas, D.; Weinberg, Z.; Jeong, K.; Adesogan, A. Bacterial diversity and composition of alfalfa silage as analyzed by Illumina MiSeq sequencing: Effects of Escherichia coli O157:H7 and silage additives. J. Dairy Sci. 2018, 101, 2048–2059. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Day | H_D | H_L | I_H | I_T | S_T | S_S | SEM | p-Value | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | T | A × T | ||||||||||
| Dry matter | 0 | 254 c | 245 c | 285 b | 288 b | 303 a | 302 ABa | 4.14 | <0.001 | <0.001 | 0.007 | 0.0694 |
| 5 | 256 b | 254 b | 312 a | 301 a | 307 a | 303 ABa | 8.53 | <0.001 | ||||
| 14 | 248 c | 253 c | 289 b | 298 ab | 304 ab | 317 ABa | 6.58 | <0.001 | ||||
| 45 | 256 b | 240 b | 280 a | 302 a | 302 a | 287 Ba | 5.72 | <0.001 | ||||
| 90 | 257 d | 250 d | 291 c | 294 c | 309 b | 331 Aa | 4.02 | <0.001 | ||||
| SEM | 4.56 | 5.67 | 6.93 | 5.62 | 6.04 | 7.02 | ||||||
| p-value | 0.645 | 0.407 | 0.068 | 0.385 | 0.893 | 0.012 | ||||||
| pH | 0 | 6.03 A | 5.96 A | 6.06 A | 6.07 A | 5.98 A | 5.99 A | 0.029 | 0.106 | <0.001 | <0.001 | <0.001 |
| 5 | 3.91 Ba | 3.87 Ba | 3.74 Bb | 3.72 Bb | 3.75 Bb | 3.85 Ba | 0.020 | <0.001 | ||||
| 14 | 3.87 BCa | 3.86 Ba | 3.58 Cc | 3.62 Cc | 3.75 Bb | 3.83 Ba | 0.018 | <0.001 | ||||
| 45 | 3.81 Ca | 3.78 Cab | 3.74 Bbc | 3.67 BCd | 3.69 Ccd | 3.76 Cab | 0.016 | <0.001 | ||||
| 90 | 3.80 Ca | 3.62 Db | 3.52 Cc | 3.53 Dc | 3.58 Db | 3.60 Db | 0.012 | <0.001 | ||||
| SEM | 0.019 | 0.018 | 0.023 | 0.025 | 0.013 | 0.019 | ||||||
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||||||
| Lactic acid | 0 | ND | ND | ND | ND | ND | ND | - | - | 0.148 | <0.001 | 0.092 |
| 5 | 111 BCa | 72.6 Bb | 72.9 Cb | 93.6 Bb | 76.5 Cb | 80.8 Cb | 4.81 | <0.001 | ||||
| 14 | 99.6 Cbc | 72.8 Bc | 92.5 Bbc | 174 Aa | 100 Bbc | 117 Ab | 6.48 | <0.001 | ||||
| 45 | 123 ABb | 128 Ab | 152 Aa | 94.3 Bc | 119 Ab | 100 Bc | 4.67 | <0.001 | ||||
| 90 | 139 Aa | 73.0 Bc | 138 Aa | 96.0 Bb | 122 Aa | 100 Bb | 4.87 | <0.001 | ||||
| SEM | 5.35 | 4.46 | 5.07 | 5.06 | 4.03 | 4.13 | ||||||
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||||||
| Acetic acid | 0 | ND | ND | ND | ND | ND | ND | - | - | <0.001 | <0.001 | 0.038 |
| 5 | 23.9 BCa | 14.2 Bb | 12.0 Bb | 6.11 Ac | 13.4 Bb | 12.9 Bb | 1.09 | <0.001 | ||||
| 14 | 19.8 Ca | 10.8 Bbc | 10.5 Bbc | 8.70 Ac | 14.7 Babc | 16.8 ABab | 1.71 | 0.006 | ||||
| 45 | 27.2 Ba | 32.3 Aa | 20.0 Aab | 7.54 Ac | 23.4 Aab | 20.4 Aab | 3.80 | 0.012 | ||||
| 90 | 33.5 Aa | 17.5 Bc | 21.2 Abc | 9.57 Ad | 24.0 Ab | 18.6 ABc | 1.41 | <0.001 | ||||
| SEM | 1.57 | 4.12 | 0.934 | 1.30 | 0.630 | 1.52 | ||||||
| p-value | <0.001 | 0.004 | <0.001 | 0.003 | <0.001 | <0.001 | ||||||
| Propionic acid | 0 | ND | ND | ND | ND | ND | ND | - | - | <0.001 | <0.001 | 0.076 |
| 5 | ND | 3.39 Ba | 2.93 Da | 2.24 Ca | 3.08 Ca | ND | 0.2880 | <0.001 | ||||
| 14 | 6.39 Cb | 3.99 Bbc | ND | 1.22 Cc | 14.6 Ba | ND | 1.1440 | <0.001 | ||||
| 45 | 10.9 Bb | 13.9 Ab | 6.17 Bc | 6.86 Bc | 17.3 Ba | 6.41 Bc | 0.9800 | <0.001 | ||||
| 90 | 16.3 Ab | 11.2 Ac | 9.61 Ac | 10.6 Ac | 20.5 Aa | 10.3 Ac | 0.9965 | <0.001 | ||||
| SEM | 0.947 | 1.10 | 0.565 | 0.717 | 0.891 | 0.521 | ||||||
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||||||
| Ammonia nitrogen | 0 | 8.47 Da | 9.70 Ca | 7.67 Ca | 2.59 Db | 5.98 Dab | 10.1 Ca | 1.2479 | 0.010 | <0.001 | <0.001 | <0.001 |
| 5 | 39.4 Ca | 33.8 Bb | 33.1 Bb | 23.1 Cc | 33.9 Cb | 25.8 Bc | 1.2595 | <0.001 | ||||
| 14 | 61.4 Ba | 35.2 Bc | 51.0 Ab | 26.4 Bc | 29.0 Bc | 31.5 Bc | 2.5968 | <0.001 | ||||
| 45 | 99.7 Aa | 62.8 Ab | 50.3 Ac | 36.1 Ad | 59.3 Ab | 46.9 Ac | 1.9001 | <0.001 | ||||
| 90 | 91.4 Aa | 66.2 Ab | 50.8 Acd | 36.3 Ae | 61.0 Abc | 44.9 Ade | 3.4829 | <0.001 | ||||
| SEM | 2.65 | 1.61 | 2.02 | 0.995 | 1.45 | 3.72 | ||||||
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||||||
| Items | Day | H_D | H_L | I_H | I_T | S_T | S_S | SEM | p-Value | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | T | A × T | ||||||||||
| Lactic acid bacteria | 0 | 8.36 Bb | 8.50 Bb | 6.80 Cd | 9.21 Aa | 7.41 Cc | 6.33 Be | 0.076 | <0.001 | 0.047 | <0.001 | 0.012 |
| 5 | 8.97 Ab | 8.88 Ab | 8.31 Bc | 8.95 Bb | 9.35 Aa | 9.43 Aa | 0.085 | <0.001 | ||||
| 14 | 8.77 Ab | 8.71 Ab | 8.65 Ab | 7.86 Cc | 8.55 Bb | 9.31 Aa | 0.075 | <0.001 | ||||
| 45 | 6.49 Cb | 7.45 Ca | 6.65 Cb | 6.59 Db | 6.10 Db | 6.61 Bb | 0.146 | <0.001 | ||||
| 90 | 5.35 Db | 6.49 Da | 4.75 Dc | 5.16 Eb | 4.05 Ed | 4.13 Cd | 0.071 | <0.001 | ||||
| SEM | 0.087 | 0.061 | 0.072 | 0.063 | 0.125 | 0.133 | ||||||
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||||||
| Enterobacteria | 0 | 6.53 Ad | 8.44 Aa | 7.48 Ac | 7.99 Ab | 7.30 Ac | 6.06 Ae | 0.096 | <0.001 | 0.048 | <0.001 | 0.004 |
| 5 | ND | ND | ND | ND | ND | ND | - | - | ||||
| 14 | ND | ND | ND | ND | ND | ND | - | - | ||||
| 45 | ND | ND | ND | ND | ND | ND | - | - | ||||
| 90 | ND | ND | ND | ND | ND | ND | - | - | ||||
| SEM | 0.035 | 0.036 | 0.063 | 0.026 | 0.004 | 0.062 | ||||||
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||||||
| Total aerobic bacteria | 0 | 7.81 Bc | 9.12 Ab | 9.13 Ab | 9.50 Aa | 7.64 Cc | 6.91 Cd | 0.091 | <0.001 | 0.009 | <0.001 | <0.001 |
| 5 | 8.73 Aa | 8.79 Ba | 8.29 Bb | 8.99 Ba | 9.08 Aa | 9.12 Ba | 0.120 | 0.004 | ||||
| 14 | 8.75 Ab | 8.54 Cc | 8.50 Bc | 7.83 Cd | 8.49 Bc | 9.44 Aa | 0.060 | <0.001 | ||||
| 45 | 6.19 Cc | 8.53 Ca | 4.68 Ce | 6.67 Db | 6.20 Dc | 5.65 Ed | 0.043 | <0.001 | ||||
| 90 | 6.22 Cb | 5.61 Dc | 4.78 Cd | 5.42 Ec | 4.39 Ee | 6.69 Da | 0.070 | <0.001 | ||||
| SEM | 0.054 | 0.655 | 0.081 | 0.092 | 0.115 | 0.066 | ||||||
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||||||
| Yeasts | 0 | 5.59 Cd | 5.26 Dd | 7.72 Bb | 8.19 Aa | 6.19 Cc | 6.60 Cc | 0.134 | <0.001 | 0.648 | <0.001 | <0.001 |
| 5 | 8.09 Ac | 7.25 Bd | 9.22 Aa | 8.19 Ac | 8.73 Ab | 8.46 Bbc | 0.132 | <0.001 | ||||
| 14 | 7.74 Aa | 7.54 Ab | 7.72 Ba | 7.59 Bab | 7.50 Bb | 7.43 Ab | 0.044 | 0.002 | ||||
| 45 | 6.15 Bab | 6.55 Ca | 3.77 Ce | 5.46 Cc | 5.97 Cb | 4.77 Dd | 0.133 | <0.001 | ||||
| 90 | 6.65 Ba | 4.83 Eb | 3.64 Cc | 5.23 Db | 4.73 Db | 5.11 Db | 0.203 | <0.001 | ||||
| SEM | 0.170 | 0.071 | 0.083 | 0.070 | 0.113 | 0.239 | ||||||
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||||||
| Items | Day | H_D | H_L | I_H | I_T | S_T | S_S | SEM | p-Value | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | T | A × T | ||||||||||
| Raw reads | 0 | 72672B5 | 77394 | 57874 C | 61165 | 76717 A | 46439 C | 6798 | 0.048 | <0.001 | <0.001 | 0.002 |
| 5 | 82678 A | 84881 | 82158 AB | 82724 | 82624 A | 84795 A | 1217 | 0.420 | ||||
| 14 | 83060 A | 85767 | 86449 A | 85417 | 84053 A | 83291 A | 1454 | 0.656 | ||||
| 45 | 83672 Aa6 | 80723 a | 68678 BCb | 82555 a | 64388 Bbc | 59492 Bc | 1995 | <0.001 | ||||
| 90 | 80527 Aa | 69953 ab | 63394 Cbc | 74775 ab | 56173 Bc | 53520 BCc | 3522 | 0.001 | ||||
| SEM | 1768 | 3461 | 4412 | 6015 | 3256 | 2531 | ||||||
| p-value | 0.007 | 0.051 | 0.004 | 0.074 | <0.001 | <0.001 | ||||||
| Clean reads | 0 | 59974 B | 70842 | 43763 B | 48132 B | 62964 AB | 40468 B | 6401 | 0.040 | <0.001 | <0.001 | <0.001 |
| 5 | 67089 ABab | 71377 ab | 63326 Ab | 75744 Aa | 69921 Aab | 75001 Aa | 1968 | 0.009 | ||||
| 14 | 67377 AB | 69169 | 69527 A | 75848 A | 68202 A | 72667 A | 2016 | 0.146 | ||||
| 45 | 73966 Aa | 68423 a | 60153 Ab | 73618 Aa | 53947 ABbc | 48486 Bc | 2315 | <0.001 | ||||
| 90 | 71015 Aa | 62192 abc | 57832 Aabc | 66824 Aab | 50882 Bbc | 47072 Bc | 3807 | 0.006 | ||||
| SEM | 2024 | 4567 | 4166 | 4066 | 3753 | 2236 | ||||||
| p-value | 0.006 | 0.641 | 0.015 | 0.011 | 0.017 | <0.001 | ||||||
| Observed species | 0 | 632 Aa | 236 b | 121 Bb | 102 Cb | 135 Cb | 232 b | 40.4 | <0.001 | <0.001 | <0.001 | 0.003 |
| 5 | 307 C | 184 | 143 B | 89.3 C | 138 C | 255 | 36.8 | 0.054 | ||||
| 14 | 300 C | 253 | 176 B | 189 B | 212 BC | 257 | 22.1 | 0.077 | ||||
| 45 | 326 C | 321 | 244 A | 315 A | 252 AB | 310 | 25.8 | 0.151 | ||||
| 90 | 411 B | 292 | 291 A | 251 AB | 309 A | 340 | 34.8 | 0.244 | ||||
| SEM | 22.2 | 40.8 | 1544 | 23.3 | 21.2 | 28.5 | ||||||
| p-value | <0.0001 | 0.2324 | <0.0001 | <0.0001 | 0.0007 | 0.1154 | ||||||
| Shannon | 0 | 6.99 Aa | 2.72 b | 3.27 Bb | 2.62 Bb | 3.07 Bb | 4.46 ABb | 0.767 | 0.017 | <0.001 | <0.001 | 0.839 |
| 5 | 4.74 Ba | 3.19 ab | 3.60 ABab | 1.36 Cb | 2.85 Bab | 3.56 Bab | 0.521 | 0.096 | ||||
| 14 | 4.97 B | 4.31 | 3.70 AB | 2.68 B | 3.83 AB | 3.81 B | 0.334 | 0.057 | ||||
| 45 | 4.90 B | 4.78 | 4.35 A | 3.76 A | 3.90 AB | 5.16 AB | 0.352 | 0.197 | ||||
| 90 | 5.39 B | 4.52 | 4.52 A | 3.56 A | 4.82 A | 5.69 A | 0.384 | 0.076 | ||||
| SEM | 0.199 | 0.724 | 0.217 | 0.271 | 0.268 | 0.405 | ||||||
| p-value | <0.001 | 0.263 | 0.011 | <0.001 | 0.003 | 0.019 | ||||||
| Simpson | 0 | 0.976 A | 0.483 | 0.718 | 0.655 A | 0.750 BC | 0.871 AB | 0.116 | 0.140 | 0.009 | 0.005 | 0.956 |
| 5 | 0.897 Ba | 0.665 ab | 0.841 a | 0.407 Bb | 0.691 Cab | 0.748 Ba | 0.075 | 0.051 | ||||
| 14 | 0.920 Ba | 0.829 a | 0.842 a | 0.630 Ab | 0.814 ABa | 0.805 ABa | 0.037 | 0.013 | ||||
| 45 | 0.893 B | 0.881 | 0.864 | 0.715 A | 0.835 AB | 0.916 A | 0.042 | 0.268 | ||||
| 90 | 0.920 B | 0.856 | 0.860 | 0.725 A | 0.901 A | 0.933 A | 36.9 | 0.228 | ||||
| SEM | 0.012 | 0.110 | 0.045 | 0.055 | 0.030 | 0.031 | ||||||
| p-value | 0.005 | 0.125 | 0.205 | 0.005 | 0.005 | 0.011 | ||||||
| Chao1 | 0 | 648 Aa | 250 b | 124 Bb | 107 Cb | 139 Cb | 234 b | 40.6 | <0.001 | <0.001 | <0.001 | 0.002 |
| 5 | 319 C | 193 | 149 B | 98.0 C | 145 C | 271 | 38.1 | 0.050 | ||||
| 14 | 313 C | 262 | 186 B | 208 B | 219 BC | 271 | 23.6 | 0.117 | ||||
| 45 | 340 C | 337 | 250 A | 336 A | 261 AB | 317 | 25.6 | 0.086 | ||||
| 90 | 433 B | 301 | 297 A | 260 B | 316 A | 349 | 0.221 | |||||
| SEM | 23.4 | 40.6 | 17.1 | 26.2 | 22.0 | 29.5 | ||||||
| p-value | <0.001 | 0.205 | <0.001 | <0.001 | <0.001 | 0.128 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Han, H.; Sun, L.; Na, N.; Xu, H.; Chang, S.; Jiang, Y.; Xue, Y. Bacterial Succession Pattern during the Fermentation Process in Whole-Plant Corn Silage Processed in Different Geographical Areas of Northern China. Processes 2021, 9, 900. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9050900
Wang C, Han H, Sun L, Na N, Xu H, Chang S, Jiang Y, Xue Y. Bacterial Succession Pattern during the Fermentation Process in Whole-Plant Corn Silage Processed in Different Geographical Areas of Northern China. Processes. 2021; 9(5):900. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9050900
Chicago/Turabian StyleWang, Chao, Hongyan Han, Lin Sun, Na Na, Haiwen Xu, Shujuan Chang, Yun Jiang, and Yanlin Xue. 2021. "Bacterial Succession Pattern during the Fermentation Process in Whole-Plant Corn Silage Processed in Different Geographical Areas of Northern China" Processes 9, no. 5: 900. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9050900