Three-Dimensional Mapping of Retrograde Multi-Labeled Motor Neuron Columns in the Spinal Cord

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Fluorescent Tracers
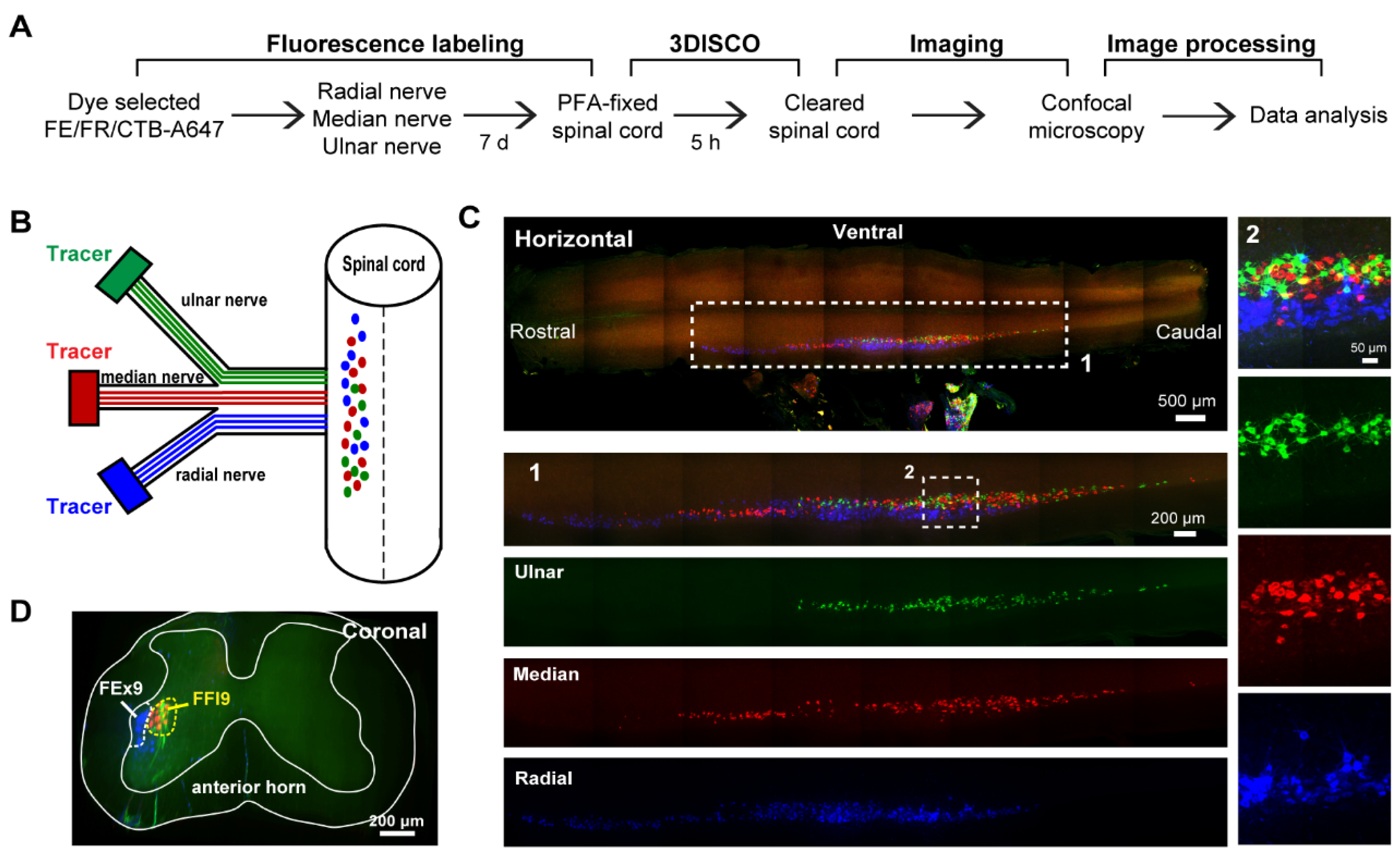
2.3. Retrograde Tracing
2.4. Tissue Harvesting
2.5. Clearing Procedure
2.6. Confocal Fluorescence Microscopy
2.7. Image Processing and Analysis
2.8. Statistical Analysis
3. Results
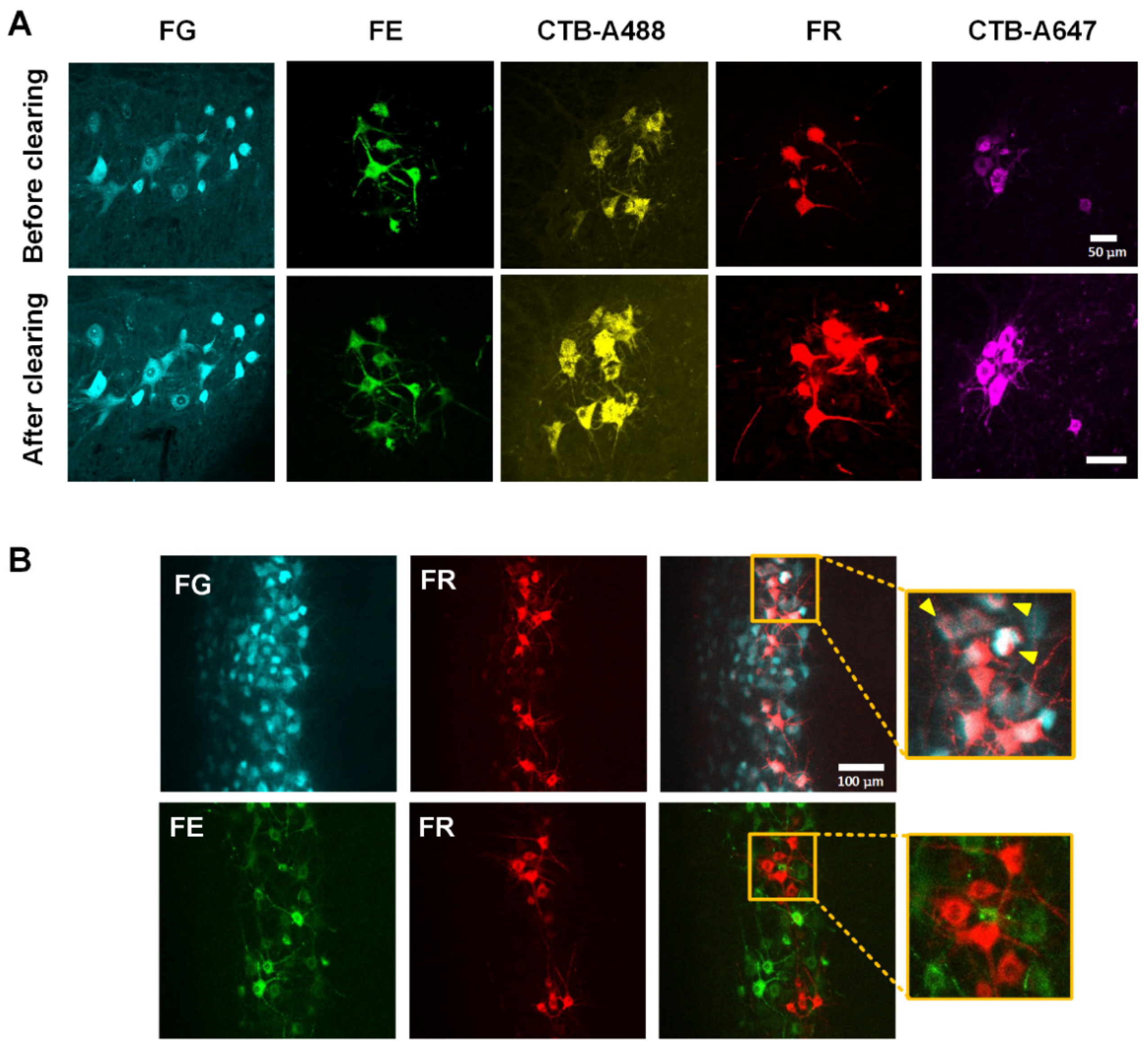
3.1. Screening of Fluorescent Dyes for Multi-Labeling of Motor Neurons
3.2. The Motor Neurons of Brachial Plexus in the Spinal Cord
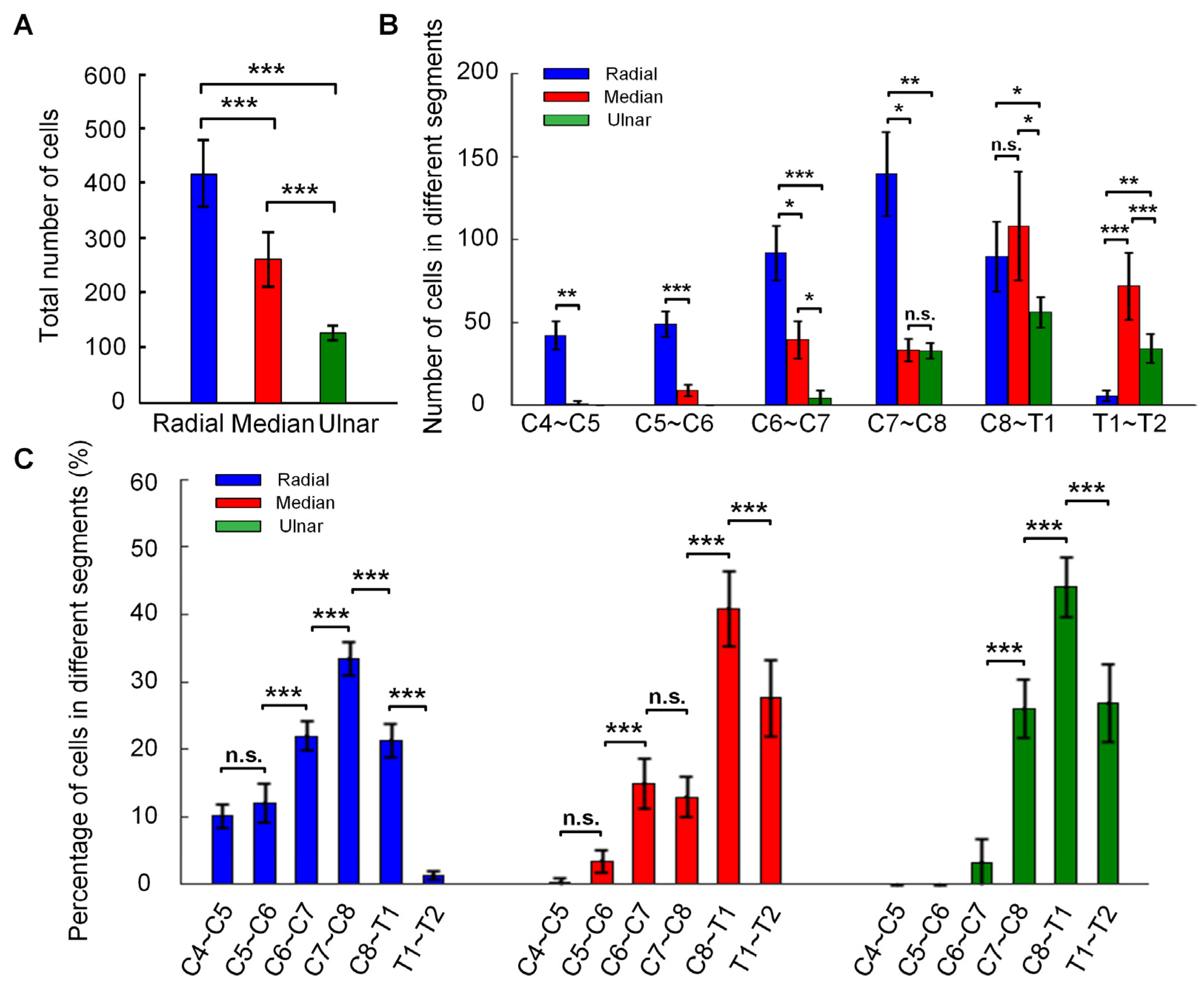
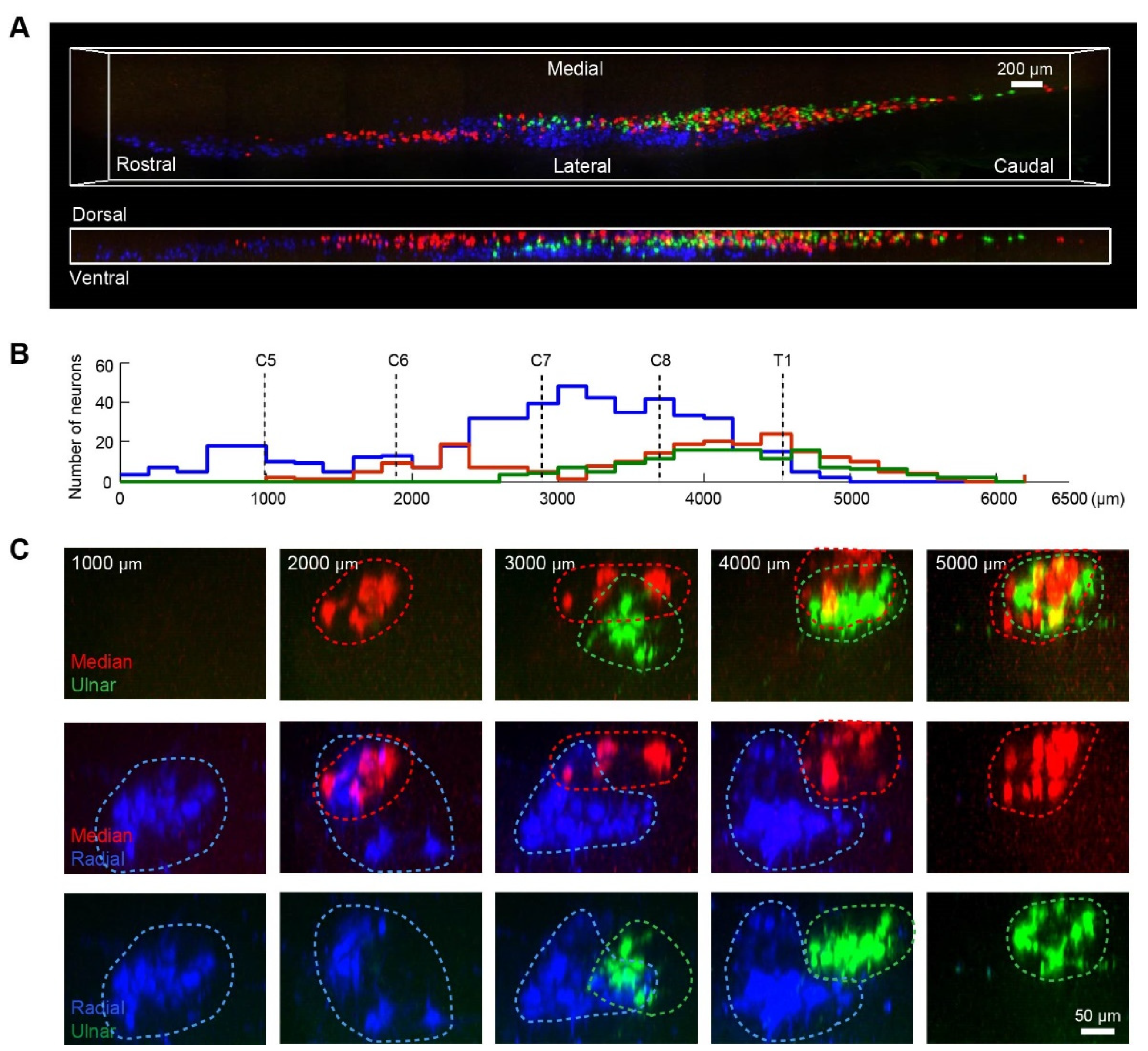
3.3. The Distribution Characteristics of Motor Neurons of Ulnar, Median, and Radial Nerves
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taylor, C.A.; Braza, D.; Rice, J.B.; Dillingham, T. The Incidence of Peripheral Nerve Injury in Extremity Trauma. Am. J. Phys. Med. Rehabil. 2008, 87, 381–385. [Google Scholar] [CrossRef]
- Li, R.; Liu, Z.; Pan, Y.; Chen, L.; Zhang, Z.; Lu, L. Peripheral Nerve Injuries Treatment: A Systematic Review. Cell Biochem. Biophys. 2014, 68, 449. [Google Scholar] [CrossRef]
- Yang, X.X.; Huang, Z.Q.; Li, Z.H.; Ren, D.F.; Tang, J.G. Risk factors and the surgery affection of respiratory complication and its mortality after acute traumatic cervical spinal cord injury. Medicine 2017, 96, e7887. [Google Scholar] [CrossRef]
- Lykissas, M.G. Current concepts in end-to-side neurorrhaphy. World J. Orthop. 2011, 2, 102–106. [Google Scholar] [CrossRef]
- Aboutaleb Kadkhodaeian, H. Comparison of nerve repair with end to end, end to side with window and end to side without window methods in lower extremity of rat. Med. J. Islamic Repub. Iran 2011, 25, 40–46. [Google Scholar]
- Nicaise, C.; Frank, D.M.; Hala, T.J.; Authelet, M.; Pochet, R.; Adriaens, D.; Brion, J.P.; Wright, M.C.; Lepore, A.C. Early Phrenic Motor Neuron Loss and Transient Respiratory Abnormalities after Unilateral Cervical Spinal Cord Contusion. J. Neurotraum. 2013, 30, 1092–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yingtao, Y.; Jianyi, X.; Tingting, Y.; Zhilong, C.; Zhiyong, X.; Jiedong, W.; Yiqiang, H.; Yongchao, W.; Dan, Z. Flufenamic acid inhibits secondary hemorrhage and BSCB disruption after spinal cord injury. Theranostics 2018, 8, 4181–4198. [Google Scholar] [CrossRef]
- Re, D.B.; Le, V.V.; Yu, C.; Amoroso, M.W.; Politi, K.A.; Phani, S.; Ikiz, B.; Hoffmann, L.; Koolen, M.; Nagata, T. Necroptosis Drives Motor Neuron Death in Models of Both Sporadic and Familial ALS. Neuron 2014, 81, 1001–1008. [Google Scholar] [CrossRef] [Green Version]
- Frakes, A.E.; Ferraiuolo, L.; Haidet-Phillips, A.M.; Schmelzer, L.; Braun, L.; Miranda, C.J.; Ladner, K.J.; Bevan, A.K.; Foust, K.D.; Godbout, J.P. Microglia Induce Motor Neuron Death via the Classical NF-κB Pathway in Amyotrophic Lateral Sclerosis. Neuron 2014, 81, 1009–1023. [Google Scholar] [CrossRef] [Green Version]
- Santos, D.; Giudetti, G.; Micera, S.; Navarro, X.; del Valle, J. Focal release of neurotrophic factors by biodegradable microspheres enhance motor and sensory axonal regeneration in vitro and in vivo. Brain Res. 2016, 1636, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Novikova, L.; Novikov, L.; Kellerth, J.O. Persistent neuronal labeling by retrograde fluorescent tracers: A comparison between Fast Blue, Fluoro-Gold and various dextran conjugates. J. Neurosci. Methods 1997, 74, 9–15. [Google Scholar] [CrossRef]
- Kobbert, C.; Apps, R.; Bechmann, I.; Lanciego, J.L.; Mey, J.; Thanos, S. Current concepts in neuroanatomical tracing. Prog. Neurobiol. 2000, 62, 327–351. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, A.; Moradzadeh, A.; Hunter, D.A.; Kawamura, D.H.; Puppala, V.K.; Tung, T.H.; Mackinnon, S.E.; Myckatyn, T.M. Retrograde Labeling in Peripheral Nerve Research: It Is Not All Black and White. J. Reconstr. Microsurg. 2007, 23, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Zele, T.; Sketelj, J.; Bajrovic, F.F. Efficacy of fluorescent tracers in retrograde labeling of cutaneous afferent neurons in the rat. J. Neurosci. Methods 2010, 191, 208–214. [Google Scholar] [CrossRef]
- Lai, B.Q.; Qiu, X.C.; Ke, Z.; Zhang, R.Y.; Hui, J.; Ge, L.; Shen, H.Y.; Wu, J.L.; Ling, E.A.; Zeng, Y.S. Cholera Toxin B Subunit Shows Transneuronal Tracing after Injection in an Injured Sciatic Nerve. PLoS ONE 2015, 10, e0144030. [Google Scholar] [CrossRef]
- Vandeweerd, J.; Hontoir, F.; De Knoop, A.; De Swert, K.; Nicaise, C. Retrograde Neuroanatomical Tracing of Phrenic Motor Neurons in Mice. J. Vis. Exp. 2018, e56758. [Google Scholar] [CrossRef]
- Han, S.; Li, D.D.; Kou, Y.H.; Fu, Z.G.; Yin, X.F. Multiple retrograde tracing methods compatible with 3DISCO clearing. Artif. Cells Nanomed. Biotechnol. 2019, 47, 4240–4247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolopoulos-Stournaras, S.; Iles, J.F. Motor neuron columns in the lumbar spinal cord of the rat. J. Comp. Neurol. 1983, 217, 75–85. [Google Scholar] [CrossRef]
- Oka, Y.; Ohtani, R.; Satou, M.; Ueda, K. Location of forelimb motoneurons in the Japanese toad (Bufo japonicus): A horseradish peroxidase study. J. Comp. Neurol. 1989, 286, 376–383. [Google Scholar] [CrossRef]
- Tosolini, A.P.; Morris, R. Spatial characterization of the motor neuron columns supplying the rat forelimb. Neuroscience 2012, 200, 19–30. [Google Scholar] [CrossRef]
- Bacskai, T.; Fu, Y.; Sengul, G.; Rusznak, Z.; Paxinos, G.; Watson, C. Musculotopic organization of the motor neurons supplying forelimb and shoulder girdle muscles in the mouse. Brain Struct. Funct. 2013, 218, 221–238. [Google Scholar] [CrossRef] [PubMed]
- Tosolini, A.P.; Mohan, R.; Morris, R. Targeting the Full Length of the Motor End Plate Regions in the Mouse Forelimb Increases the Uptake of Fluoro-Gold into Corresponding Spinal Cord Motor Neurons. Front. Neurol. 2013, 4, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacskai, T.; Rusznak, Z.; Paxinos, G.; Watson, C. Musculotopic organization of the motor neurons supplying the mouse hindlimb muscles: A quantitative study using Fluoro-Gold retrograde tracing. Brain Struct. Funct. 2014, 219, 303–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohan, R.; Tosolini, A.P.; Morris, R. Targeting the motor end plates in the mouse hindlimb gives access to a greater number of spinal cord motor neurons: An approach to maximize retrograde transport. Neuroscience 2014, 274, 318–330. [Google Scholar] [CrossRef] [PubMed]
- Mohan, R.; Tosolini, A.P.; Morris, R. Segmental distribution of the motor neuron columns that supply the rat hindlimb: A muscle/motor neuron tract-tracing analysis targeting the motor end plates. Neuroscience 2015, 307, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Pfister, B.J.; Gordon, T.; Loverde, J.R.; Kochar, A.S.; Mackinnon, S.E.; Cullen, D.K. Biomedical engineering strategies for peripheral nerve repair: Surgical applications, state of the art, and future challenges. Crit. Rev. Biomed. Eng. 2011, 39, 81–124. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Gu, Y.; Xu, J.; Lin, S.; Chen, L.; Lu, J. Contralateral C7 transfer via the prespinal and retropharyngeal route to repair brachial plexus root avulsion: A preliminary report. Neurosurgery 2008, 63, 553–558. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, L.; Lao, J.; Zhang, X. Total brachial plexus injury: Contralateral C7 root transfer to the lower trunk versus the median nerve. Neural Regen. Res. 2018, 13, 1968–1973. [Google Scholar] [CrossRef]
- Richardson, D.S.; Lichtman, J.W. Clarifying Tissue Clearing. Cell 2015, 162, 246–257. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Larin, K.V.; Luo, Q.; Tuchin, V.V. Recent progress in tissue optical clearing. Laser Photonics Rev. 2013, 7, 732–757. [Google Scholar] [CrossRef] [Green Version]
- Becker, K.; Jährling, N.; Saghafi, S.; Weiler, R.; Dodt, H.-U. Chemical clearing and dehydration of GFP expressing mouse brains. PLoS ONE 2012, 7, e33916. [Google Scholar] [CrossRef]
- Ertürk, A.; Becker, K.; Jährling, N.; Mauch, C.; Hojer, C.; Egen, J.; Hellal, F.; Bradke, F.; Sheng, M.; Dodt, H.-U. Three-dimensional imaging of solvent-cleared organs using 3DISCO. Nat. Protoc. 2012, 7, 1983–1995. [Google Scholar] [CrossRef]
- Pan, C.; Cai, R.; Quacquarelli, F.P.; Ghasemigharagoz, A.; Lourbopoulos, A.; Matryba, P.; Plesnila, N.; Dichgans, M.; Hellal, F.; Ertürk, A. Shrinkage-mediated imaging of entire organs and organisms using uDISCO. Nat. Methods 2016, 13, 859–867. [Google Scholar] [CrossRef]
- Hama, H.; Hioki, H.; Namiki, K.; Hoshida, T.; Kurokawa, H.; Ishidate, F.; Kaneko, T.; Akagi, T.; Saito, T.; Saido, T. ScaleS: An optical clearing palette for biological imaging. Nat. Neurosci. 2015, 18, 1518–1529. [Google Scholar] [CrossRef] [PubMed]
- Susaki, E.A.; Tainaka, K.; Perrin, D.; Kishino, F.; Tawara, T.; Watanabe, T.M.; Yokoyama, C.; Onoe, H.; Eguchi, M.; Yamaguchi, S.; et al. Whole-brain imaging with single-cell resolution using chemical cocktails and computational analysis. Cell 2014, 157, 726–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, K.; Wallace, J.; Kim, S.Y.; Kalyanasundaram, S.; Andalman, A.S.; Davidson, T.J.; Mirzabekov, J.J.; Zalocusky, K.A.; Mattis, J.; Denisin, A.K.; et al. Structural and molecular interrogation of intact biological systems. Nature 2013, 497, 332–337. [Google Scholar] [CrossRef]
- Jing, D.; Zhang, S.W.; Luo, W.J.; Gao, X.F.; Men, Y.; Ma, C.; Liu, X.H.; Yi, Y.T.; Bugde, A.; Zhou, B.O.; et al. Tissue clearing of both hard and soft tissue organs with the PEGASOS method (vol 28, pg 803, 2018). Cell Res. 2019, 29, 506. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.; Pan, C.; Ghasemigharagoz, A.; Todorov, M.I.; Forstera, B.; Zhao, S.; Bhatia, H.S.; Parradamas, A.; Mrowka, L.; Theodorou, D. Panoptic Imaging of Transparent Mice Reveals Whole-Body Neuronal Projections and Skull-Meninges Connections. Nat. Neurosci. 2019, 22, 317–327. [Google Scholar] [CrossRef]
- Zhu, J.T.; Yu, T.T.; Li, Y.S.; Xu, J.Y.; Qi, Y.S.; Yao, Y.T.; Ma, Y.L.; Wan, P.; Chen, Z.L.; Li, X.N.; et al. MACS: Rapid Aqueous Clearing System for 3D Mapping of Intact Organs. Adv. Sci. 2020, 7, 1903185. [Google Scholar] [CrossRef] [Green Version]
- Wan, P.; Zhu, J.; Xu, J.; Li, Y.; Yu, T.; Zhu, D. Evaluation of seven optical clearing methods in mouse brain. Neurophotonics 2018, 5, 035007. [Google Scholar] [CrossRef] [Green Version]
- Ertürk, A.; Mauch, C.P.; Hellal, F.; Förstner, F.; Keck, T.; Becker, K.; Jährling, N.; Steffens, H.; Richter, M.; Hübener, M.; et al. Three-dimensional imaging of the unsectioned adult spinal cord to assess axon regeneration and glial responses after injury. Nat. Med. 2012, 18, 166–171. [Google Scholar] [CrossRef]
- Launay, P.-S.; Godefroy, D.; Khabou, H.; Rostene, W.; Sahel, J.-A.; Baudouin, C.; Melik Parsadaniantz, S.; Reaux-Le Goazigo, A. Combined 3DISCO clearing method, retrograde tracer and ultramicroscopy to map corneal neurons in a whole adult mouse trigeminal ganglion. Exp. Eye Res. 2015, 139, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Belle, M.; Godefroy, D.; Dominici, C.; Heitz-Marchaland, C.; Zelina, P.; Hellal, F.; Bradke, F.; Chédotal, A. A Simple Method for 3D Analysis of Immunolabeled Axonal Tracts in a Transparent Nervous System. Cell Rep. 2014, 9, 1191–1201. [Google Scholar] [CrossRef]
- Casoni, F.; Malone, S.A.; Belle, M.; Luzzati, F.; Collier, F.; Allet, C.; Hrabovszky, E.; Rasika, S.; Prevot, V.; Chédotal, A.; et al. Development of the neurons controlling fertility in humans: New insights from 3D imaging and transparent fetal brains. Development 2016, 143, 3969–3981. [Google Scholar] [CrossRef] [Green Version]
- Žygelytė, E.; Bernard, M.E.; Tomlinson, J.E.; Martin, M.J.; Terhorst, A.; Bradford, H.E.; Lundquist, S.A.; Sledziona, M.; Cheetham, J. RetroDISCO: Clearing technique to improve quantification of retrograde labeled motor neurons of intact mouse spinal cords. J. Neurosci. Methods 2016, 271, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Robinson, G.A.; Madison, R.D. Preferential motor reinnervation in the mouse: Comparison of femoral nerve repair using a fibrin sealant or suture. Muscle Nerve 2003, 28, 227–231. [Google Scholar] [CrossRef]
- Richmond, F.J.R.; Gladdy, R.; Creasy, J.L.; Kitamura, S.; Smits, E.; Thomson, D.B. Efficacy of seven retrograde tracers, compared in multiple-labelling studies of feline motoneurones. J. Neurosci. Methods 1994, 53, 35–46. [Google Scholar] [CrossRef]
- Choi, D.; Li, D.; Raisman, G. Fluorescent retrograde neuronal tracers that label the rat facial nucleus: A comparison of Fast Blue, Fluoro-ruby, Fluoro-emerald, Fluoro-Gold and DiI. J. Neurosci. Methods 2002, 117, 167–172. [Google Scholar] [CrossRef]
- Hirakawa, M.; Mccabe, J.T.; Kawata, M. Time-related changes in the labeling pattern of motor and sensory neurons innervating the gastrocnemius muscle, as revealed by the retrograde transport of the cholera toxin B subunit. Cell Tissue Res. 1992, 267, 419–427. [Google Scholar] [CrossRef]
- Pollin, M.M.; McHanwell, S.; Slater, C.R. The effect of age on motor neurone death following axotomy in the mouse. Development 1991, 112, 83–89. [Google Scholar] [CrossRef]
- Welin, D.; Novikova, L.N.; Wiberg, M.; Kellerth, J.O.; Novikov, L.N. Survival and regeneration of cutaneous and muscular afferent neurons after peripheral nerve injury in adult rats. Exp. Brain Res. 2008, 186, 315–323. [Google Scholar] [CrossRef]
- Scarisbrick, I.A.; Haase, P.; Hrycyshyn, A.W. The arrangement of forearm motoneurons in young and adult rats and the possibility of naturally occurring motoneuron death. J. Anat. 1990, 171, 57–67. [Google Scholar] [CrossRef]
- Byers, C.T.; Fan, R.; Messina, A.; Morrison, W.A.; Galea, M.P. Comparing the efficacy of two fluorescent retrograde tracers in labeling the motor and sensory neuron populations of the rat sciatic nerve. J. Neurosci. Methods 2002, 114, 159–164. [Google Scholar] [CrossRef]
- Preston, D.C.; Shapiro, B.E. Median Neuropathy at the Wrist. In Electromyography and Neuromuss, 3rd ed.; Preston, D.C., Shapiro, B.E., Eds.; W.B. Saunders: London, UK, 2013; pp. 267–288. [Google Scholar] [CrossRef]
- Oh, C.-S.; Won, H.-S.; Lee, K.-S.; Chung, I.-H.; Kim, S.M. Anatomic variation of the innervation of the flexor digitorum profundus muscle and its clinical implications. Muscle Nerve 2009, 39, 498–502. [Google Scholar] [CrossRef]
- Glover, J.C.; Petursdottir, G.; Jansen, J. Fluorescent dextran-amines used as axonal tracers in the nervous system of the chicken embryo. J. Neurosci. Methods 1986, 18, 243–254. [Google Scholar] [CrossRef]
- Dubový, P.; Raška, O.; Klusáková, I.; Stejskal, L.; Čelakovský, P.; Haninec, P. Ciliary neurotrophic factor promotes motor reinnervation of the musculocutaneous nerve in an experimental model of end-to-side neurorrhaphy. BMC Neurosci. 2011, 12, 58. [Google Scholar] [CrossRef] [Green Version]
- Brushart, T.M. Motor axons preferentially reinnervate motor pathways. J. Neurosci. 1993, 13, 2730–2738. [Google Scholar] [CrossRef]
- Brushart, T.M. Preferential reinnervation of motor nerves by regenerating motor axons. J. Neurosci. 1988, 8, 1026–1031. [Google Scholar] [CrossRef] [PubMed]
- Schofield, B.R. Retrograde axonal tracing with fluorescent markers. Curr. Protoc. Neurosci. 2008, 43, 1–17. [Google Scholar] [CrossRef]
- Yao, F.; Zhang, E.; Gao, Z.; Ji, H.; Marmouri, M.; Xia, X. Did you choose appropriate tracer for retrograde tracing of retinal ganglion cells? The differences between cholera toxin subunit B and Fluorogold. PLoS ONE 2018, 13, e0205133. [Google Scholar] [CrossRef] [Green Version]
- Schmued, L.C.; Fallon, J.H. Fluoro-gold: A new fluorescent retrograde axonal tracer with numerous unique properties. Brain Res. 1986, 377, 147–154. [Google Scholar] [CrossRef]
- Colin, W.; Donoff, R.B.; Foote, W.E. Fluorescent latex microspheres as a retrograde tracer in the peripheral nervous system. Brain Res. 1989, 486, 334–339. [Google Scholar] [CrossRef]
- Katz, L.C.; Iarovici, D.M. Green fluorescent latex microspheres: A new retrograde tracer. Neuroscience 1990, 34, 511–520. [Google Scholar] [CrossRef]
- Hoover, J.E.; Durkovic, R.G. Retrograde Labeling of Lumbosacral Interneurons Following Injections of Red and Green Fluorescent Microspheres into Hindlimb Motor Nuclei of the Cat. Somatosens. Mot. Res. 1992, 9, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Tainaka, K.; Kuno, A.; Kubota, S.I.; Murakami, T.; Ueda, H.R. Chemical Principles in Tissue Clearing and Staining Protocols for Whole-Body Cell Profiling. Annu. Rev. Cell Dev. Biol. 2016, 32, 713–741. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Qi, Y.; Gong, H.; Luo, Q.; Zhu, D. Optical clearing for multiscale biological tissues. J. Biophotonics 2018, 11, e201700187. [Google Scholar] [CrossRef]
- Ertürk, A.; Bradke, F. High-resolution imaging of entire organs by 3-dimensional imaging of solvent cleared organs (3DISCO). Exp. Neurol. 2013, 242, 57–64. [Google Scholar] [CrossRef]
- Ertürk, A.; Lafkas, D.; Chalouni, C. Imaging Cleared Intact Biological Systems at a Cellular Level by 3DISCO. JOVE J. Vis. Exp. 2014, e51382. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fluorescent Dyes | FG * | GB | CTB-A488 | FE | RB | FR | CTB-A647 |
|---|---|---|---|---|---|---|---|
| Excitation (nm) Emission (nm) | 361 | 460 | 490 | 494 | 530 | 555 | 650 |
| 536 | 505 | 520 | 521 | 590 | 580 | 668 | |
| Labeling effectiveness | + | − | + | + | − | + | + |
| Compatibility with 3DISCO | + | / | + | + | / | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Yin, X.; Qi, Y.; Chen, B.; Li, Y.; Wan, P.; Yao, Y.; Zhu, D.; Jiang, B.; Yu, T. Three-Dimensional Mapping of Retrograde Multi-Labeled Motor Neuron Columns in the Spinal Cord. Photonics 2021, 8, 145. https://0-doi-org.brum.beds.ac.uk/10.3390/photonics8050145
Xu J, Yin X, Qi Y, Chen B, Li Y, Wan P, Yao Y, Zhu D, Jiang B, Yu T. Three-Dimensional Mapping of Retrograde Multi-Labeled Motor Neuron Columns in the Spinal Cord. Photonics. 2021; 8(5):145. https://0-doi-org.brum.beds.ac.uk/10.3390/photonics8050145
Chicago/Turabian StyleXu, Jianyi, Xiaofeng Yin, Yisong Qi, Bo Chen, Yusha Li, Peng Wan, Yingtao Yao, Dan Zhu, Baoguo Jiang, and Tingting Yu. 2021. "Three-Dimensional Mapping of Retrograde Multi-Labeled Motor Neuron Columns in the Spinal Cord" Photonics 8, no. 5: 145. https://0-doi-org.brum.beds.ac.uk/10.3390/photonics8050145