The Maturation Pathway of Nickel Urease

School of Life Sciences, Centre for Protein Science and Crystallography, State Key Laboratory of Agrobiotechnology, The Chinese University of Hong Kong, Hong Kong 999077, China
*
Author to whom correspondence should be addressed.
Inorganics 2019, 7(7), 85; https://0-doi-org.brum.beds.ac.uk/10.3390/inorganics7070085
Submission received: 7 June 2019
/
Revised: 28 June 2019
/
Accepted: 4 July 2019
/
Published: 6 July 2019
(This article belongs to the Special Issue Bioinorganic Chemistry of Nickel)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Maturation of urease involves post-translational insertion of nickel ions to form an active site with a carbamylated lysine ligand and is assisted by urease accessory proteins UreD, UreE, UreF and UreG. Here, we review our current understandings on how these urease accessory proteins facilitate the urease maturation. The urease maturation pathway involves the transfer of Ni2+ from UreE → UreG → UreF/UreD → urease. To avoid the release of the toxic metal to the cytoplasm, Ni2+ is transferred from one urease accessory protein to another through specific protein–protein interactions. One central theme depicts the role of guanosine triphosphate (GTP) binding/hydrolysis in regulating the binding/release of nickel ions and the formation of the protein complexes. The urease and [NiFe]-hydrogenase maturation pathways cross-talk with each other as UreE receives Ni2+ from hydrogenase maturation factor HypA. Finally, the druggability of the urease maturation pathway is reviewed.
1. Introduction
Urease catalyzes the hydrolysis of urea into carbon dioxide and carbamate, which spontaneously decomposes into ammonia and another carbon dioxide. The enzyme is involved in nitrogen metabolism that is found in bacteria, archaea, fungi, plants, and some invertebrates [1,2,3]. While the Jack bean (Canavanlia ensiformis) urease was the first nickel enzyme identified [4] and the first enzyme to be crystallized [5], the mechanism of urease maturation is the most well studied in bacteria, in particular Klebsiella aerogenes and Helicobacter pylori. Urease has been implicated in the pathogenesis of bacterial infections. For example, H. pylori can colonize in the acidic stomach due to the ureolytic activity of urease [6]. H. pylori infection increases the risk of peptic ulcer and gastric cancer [7,8]. Most of the ureases discovered so far are nickel enzymes. One notable exception is an iron urease found in Helicobacter mustelae in ferret stomach [9].
The concentration of free nickel ions is tightly regulated in cells because Ni2+ can inactivate enzymes by displacing weaker ions such as Mg2+ in the active sites [10,11]. To avoid cytotoxicity, cells have to evolve a mechanism to deliver nickel from one protein to another without releasing the toxic metal to the cytoplasm. In the urease maturation pathway, there are four urease accessory proteins, UreD, UreE, UreF and UreG, involved in the nickel delivery. In this article, we review how these metallochaperones interact with each other and with urease to facilitate the transfer of nickel ions in the urease maturation pathway. Correct metalation is ensured by specific protein–protein interactions that are allosterically regulated by binding/hydrolysis of guanosine triphosphate (GTP). We also review the cross-talking between the maturation pathways of ureases and [NiFe]-hydrogenases, urease maturation in plants and the potential of the urease maturation pathway in antibacterial drug discovery. Finally, some unanswered questions on the urease maturation pathway are discussed.
2. Structures of Urease
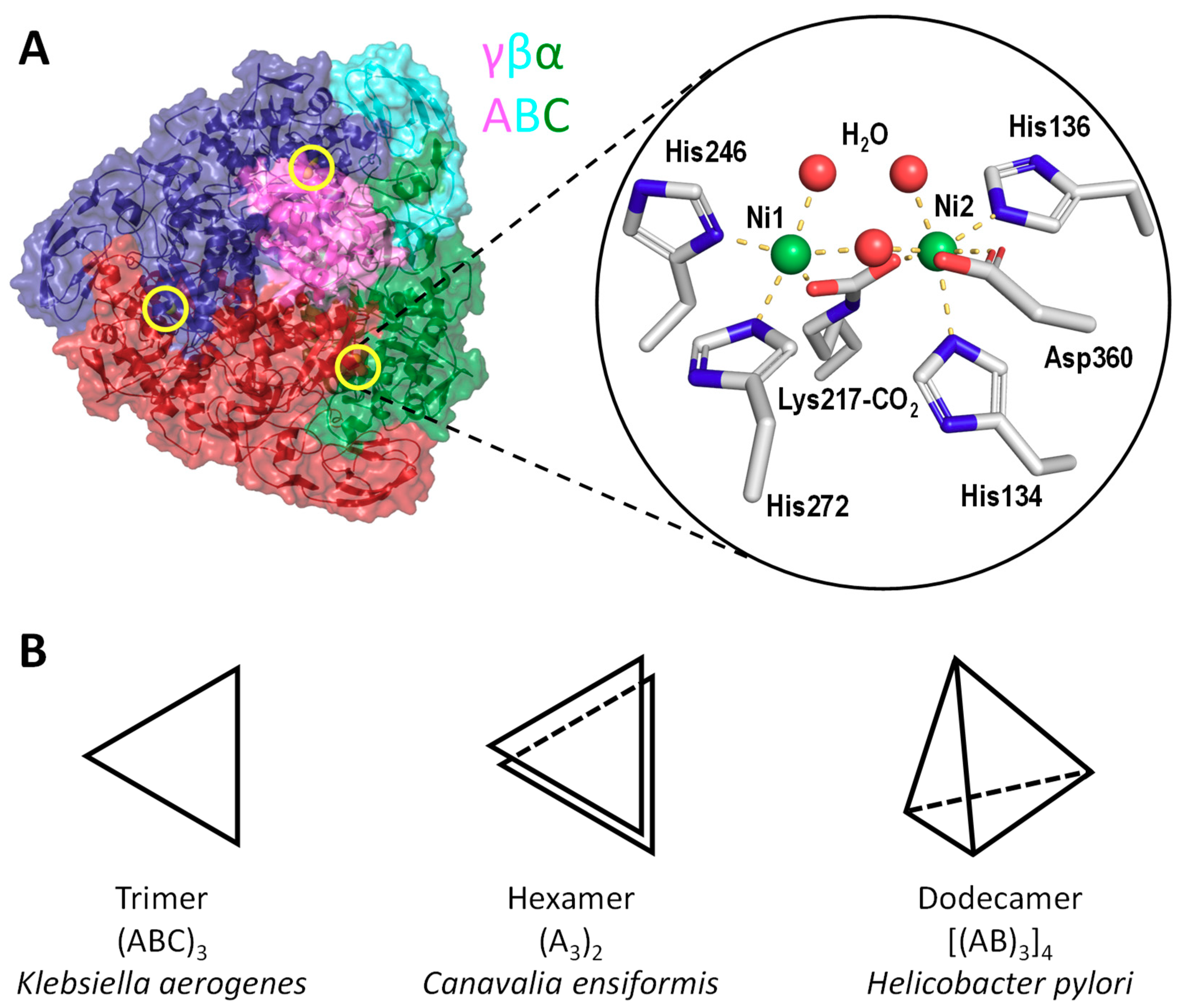
The urease sequence is highly conserved. Most of the bacterial ureases are comprised of three subunits [12]. For example, K. aerogenes urease has three subunits: α-subunit (UreC), β-subunit (UreB) and γ-subunit (UreA) (Figure 1A) [13]. In H. pylori, the two smaller β and γ subunits are fused to form the UreA, and the large α-subunit is renamed as UreB [14]. In plants, all three subunits are fused into one polypeptide chain as UreA [15]. In K. aerogenes [16,17] and Sporosarcina pasteurii [18], the urease contains three catalytic sites constituted by the α, β and γ subunits. Three copies of the subunits UreABC form a trimeric disc-like structure (Figure 1A). In plant urease, the disc-like structure dimerizes to form a hexameric urease (UreA3)2 (Figure 1B) [19]. H. pylori UreA contains a 27-residue C-terminal extension that is responsible for the formation of a dodecameric quaternary structure (UreAB3)4 [20].
The urease active site is located in the α-subunit and is highly conserved. It contains two Ni2+ ions [16,18,19,20] bridged by the carboxyl group of a carbamylated lysine [17,20,21], and chelated by four histidine residues and one aspartate residue (Figure 1A). Purified K. aerogenes urease can be activated in vitro by incubation with high concentration of bicarbonate and Ni2+ [22]. The carbamylation is likely to come from carbon dioxide, instead of bicarbonate as suggested by the result of pH jump experiment [22]. Mutagenesis studies suggest that the carbamylated lysine residues are essential to urease maturation [23]. Urease activity can be inhibited by Zn2+, Cu2+, Co2+ and Mn2+ [24].
3. Genetic Studies Showed the Importance of Urease Accessory Proteins
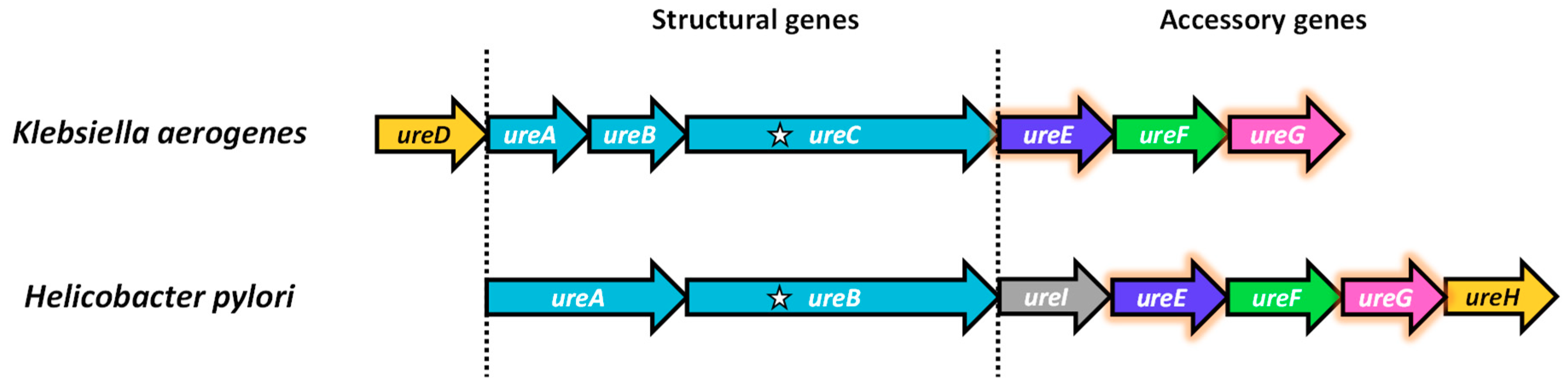
In K. aerogenes, the urease operon is arranged as ureDABCEFG [25,26] (Figure 2). ureA, ureB and ureC respectively encode the γ, β and α subunits of the urease structural genes. ureD, ureE, ureF and ureG encode for the urease accessory proteins essential for the maturation of urease. In H. pylori, the operon is arranged as ureABIEFGH [27,28]. There is a unique ureI encoding for an acid-gated urea channel for colonization in the acidic stomach [29,30]. ureH is an orthologue of ureD. For simplicity, UreD is used in this article to denote the protein product of ureH in H. pylori or ureD in other species.
Bacterial cells harboring the urease operon showed nickel-dependent activation of urease [26,31,32]. Urease purified from cells containing the intact urease operon was active, whereas the urease purified from cells containing only the urease structural genes was inactive [33,34]. This observation suggests that the urease maturation is assisted by the urease accessory proteins. Deletion [26], knockout [35] and transposon mutagenesis [28] of individual accessory genes, ureG, ureF and ureD, abolished urease activity. The activity could be partially [26] or fully [32] recovered by gene complementation. Deletion of ureE either lowered [26] or abolished the urease activity [32,35,36], which was partially regenerated by adding nickel [32]. These results suggested that UreG, UreF and UreD are absolutely required for urease maturation, while UreE facilitates the process.
4. The Formation of UreGFD Complex
Urease apoprotein was shown to interact with UreD [37,38], UreF/UreD [39,40], or UreG/UreF/UreD [38,40,41,42]. It has been known that UreD and UreF can form a UreFD complex, which can then recruit UreG to form a UreGFD complex [43]. The UreGFD complex can form an activation complex [43,44] with urease apoprotein and activate urease in a GTP-dependent manner [43,44]. Substitution of the P-loop residues of UreG abolished its ability to activate urease [45,46].
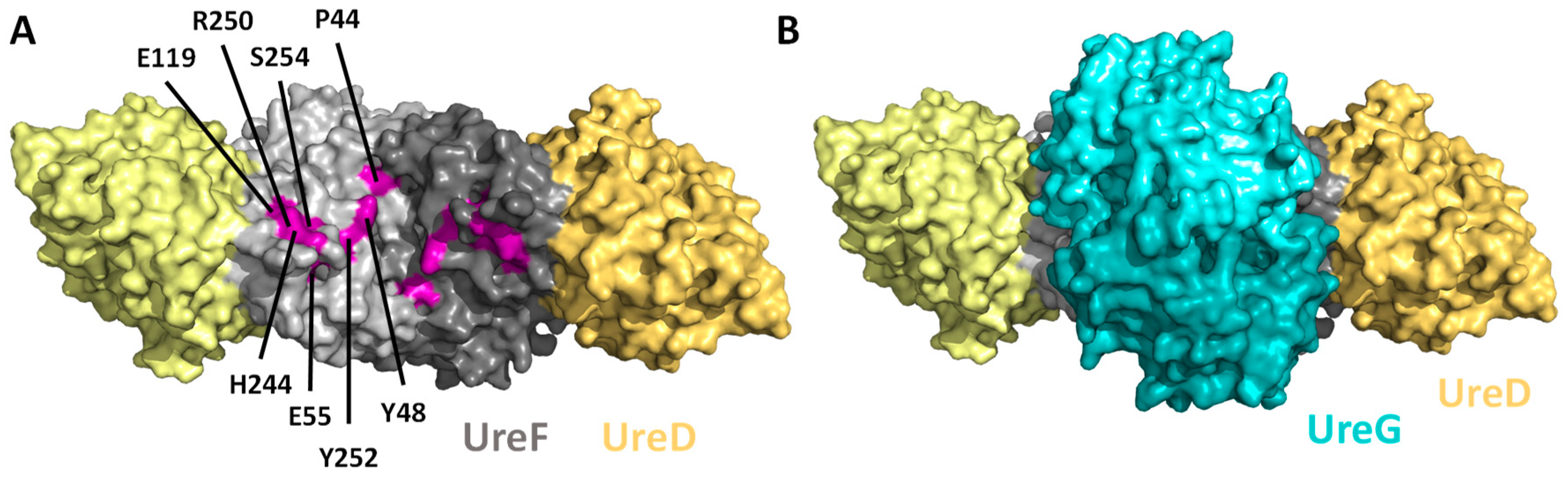
The structural studies provided insights into how UreD, UreF and UreG interact with each other to form the protein complexes required for urease maturation. UreD forms a complex with UreF [35,38,40,47]. Crystal structure of the H. pylori UreFD complex revealed that it is a 2:2 heterodimer with UreF at the middle providing the dimerization interface (Figure 3A) [48]. UreD bound to both ends of the UreF dimer, forming a rod-shape head-to-head dimer of heterodimer UreFD (Figure 3A). UreF contains highly conserved residues at its C-terminal tail, which is unstructured and susceptible to proteolytic cleavage when expressed alone [48,49]. The C-terminal tail of UreF was shown to be essential for the interaction with UreD [40,48], the assembly of an activation complex [40], and urease activation [40,48]. Upon binding to UreD, these C-terminal residues become structured and form an extra helix-10 and a loop structure stabilized by hydrogen bonds involving a conserved Arg-250 residue [48]. These conformational changes were shown to be important for recruiting UreG to form the UreGFD complex by mutagenesis studies. For example, the R250A variant abolished the formation of UreGFD complex and urease activation [43].
The crystal structure of the H. pylori UreGFD complex (PDB: 4HI0) revealed that it is a dimer of the heterotrimers of UreGFD [43] (Figure 3B). The complex contains a guanosine diphosphate (GDP)-bound UreG dimer sitting on the UreF wherein the dimerization axes of UreG and UreFD are almost perpendicular to each other. UreF-mediated dimerization is required to provide a complete UreG binding site. Substitutions that broke the dimerization of UreF abolished the recruitment of UreG to the UreGFD complex and urease maturation [40,43]. UreG binds to a saddle-like structure of UreF consisting of clusters of conserved residues [43]––some of them have been identified to be important for recruiting UreG to the activation complex and for activating urease [50] (Figure 3A).
5. UreG Dissociates from the UreGFD Complex and Forms a Dimer in the Presence of Ni/GTP
That UreG undergoes Ni/GTP-dependent dimerization was identified when Ni2+ and GTP were added to the GDP-bound UreGFD complex, which was then dissociated into a UreG dimer and the UreFD complex [43]. UreG is a nickel chaperone [43,51,52] and a SIMIBI (after Signal recognition particle, MinD and BioD) class GTPase [53]. UreG remains as a monomer in the absence of GTP and it binds Ni2+ with lower affinity [54,55]. UreG only dimerizes when both GTP and Ni2+ were present, and the UreG dimer binds one Ni2+ per dimer with a Kd of 0.36 µM [43,52]. Zn2+ can also induce dimerization of UreG [54,56,57], but the Zn/UreG dimer was not stable and dissociated to monomer when the excess Zn2+ was removed by gel filtration [43]. Moreover, this Zn/UreG dimer is inactive in GTP hydrolysis [52]. Dimerization is required for GTP hydrolysis. The cysteine and histidine in the conserved CPH motif (Cys-Pro-His) are important for UreG dimerization and nickel binding [43,54], as well as urease activation [43,46,58]. After GTP hydrolysis, the UreG dimer dissociates back to monomer and releases one Ni2+, providing a plausible mechanism for coupling GTP hydrolysis to nickel delivery [43]. GTP-dependent dimerization [59,60,61,62,63] and conformational changes [64] were also observed in HypB, another SIMIBI GTPase involved in the [NiFe]-hydrogenase maturation pathway.
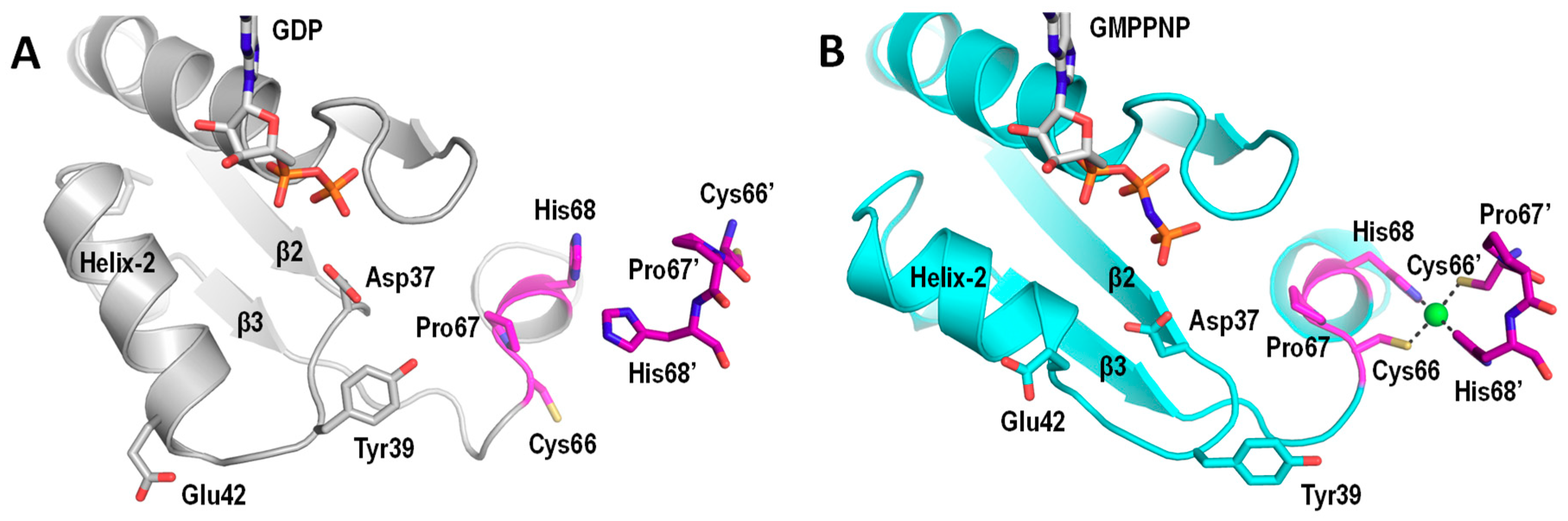
Structural insights into how Ni2+ and GTP induce dimerization of UreG were provided by the crystal structure of the UreG dimer in complex with Ni2+ and GMPPNP, a nonhydrolyzable analogue of GTP [51]. The GTP binding pocket is sandwiched between two UreG chains and a Ni2+ ion is coordinated by the conserved CPH motif from each chain [51]. The structure of Ni/GMPPNP-bound UreG dimer is compared to that of the GDP-bound UreG in the UreGFD complex (Figure 4). Upon GTP binding, the γ-phosphate of GTP introduces a charge–charge repulsion on Asp37 in the G2 switch, initiating a swinging motion of helix-2, and Glu42 forms a hydrogen bond with Arg130 of the opposite chain. Consequently, the zip-up motion of β2 and β3 propagates the conformational changes to the CPH motif, where Cys66 and His68 reorientate towards the dimeric interface to chelate a Ni2+ ion in a square-planar geometry (Figure 4B) [51]. The structural changes observed also explain why UreG dissociates from the UreGFD complex upon GTP binding as residue Tyr39 in the G2 region swings outward and makes steric clashes with UreF [48].
6. UreG–UreE Interaction Is GTP-Dependent
UreG interacts with UreE in 2:2 ratio to form a UreE2G2 complex in the presence of GTP and Mg2+. The formation of UreE2G2 is independent of Zn2+ and Ni2+ [52]. In the presence of GDP, the complex dissociates into the UreE2G complex and a UreG monomer [52]. Interestingly, addition of Ni2+ and GTPγS to the UreGFD complex and UreE promotes UreG to switch its protein binding partner from UreFD to UreE, resulting in the formation of the UreE2G2 and UreFD complexes [51]. The D37A/E42A variant of UreG failed to dissociate from the UreGFD complex and formed the Ni/GTP-bound UreG dimer, presumably disrupting the conformational changes induced by GTP binding [51]. Interestingly, the D37A/E42A variant also failed to form the UreE2G2 complex in the presence of Ni/GTP, further supporting that the conformational changes in UreG are important for the UreE–UreG interaction [51]. The formation of the UreE2G2 complex facilitates the transfer of Ni2+ from UreE to UreG. By monitoring the thiolate-to-Ni2+ transition at 337 nm, it has been shown that Ni2+ was transferred from UreE to UreG within the UreE2G2 complex in its GTP-bound state, but not in the UreE2G complex in the presence of GDP [52].
How UreE interacts with UreG is not known. Crystal structures of UreE from S. pasteurii [65], K. aerogenes [66] and H. pylori [67,68] have been solved, and share high structural homology. UreE exists as a dimer in solution [68,69,70,71]. High protein concentration and the presence of Zn2+, Cu2+ or Ni2+ can induce UreE tetramerization [65,72,73,74], but mutagenesis studies showed that the formation of the tetramer was not essential to urease activation [68]. The C-terminal domain of two UreE chains interact with each other to form a dimer. At the dimeric interface, UreE has one conserved metal-binding site formed by His102 (numbered according to the H. pylori sequence) in a GNXH motif from each of the UreE chain [65,66,67,68]. This central histidine is essential to urease maturation [68,75,76]. The variable C-terminus histidine-rich tail of UreE [76] has been shown to bind Ni2+ (residue 143–157 in K. aerogenes [70,72,77]; residue 137–147 in S. pasteurii [73]). In the crystal structure of H. pylori UreE, an extra histidine (His152) of this C-terminal tail was shown to bind a Ni2+ or Zn2+ at the dimeric interface (Figure 5B) [67,68]. Neither the H102A substitution nor truncation of the variable C-terminal tail (residue 158–170 of H. pylori UreE) affect the formation of the UreE2G2 complex [52]. Truncation of the C-terminal tail of K. aerogenes UreE showed a 25–60% decrease in urease activation [77,78]. The functional role of the C-terminal tail is unclear. However, it has been proposed that the C-terminal tail may play a role in regulating the binding and release of Ni2+ [65,67]. Based on charge and shape complementarity, models of how UreE interacts with UreG have been proposed [71,79,80]. The models predict that the nickel binding sites of UreE and UreG should point towards each other, which is supported by the mutagenesis studies that show R101A UreE or C66A UreG destabilized the UreE2G2 complex [52].
7. How Urease Accessory Proteins Facilitate Urease Maturation
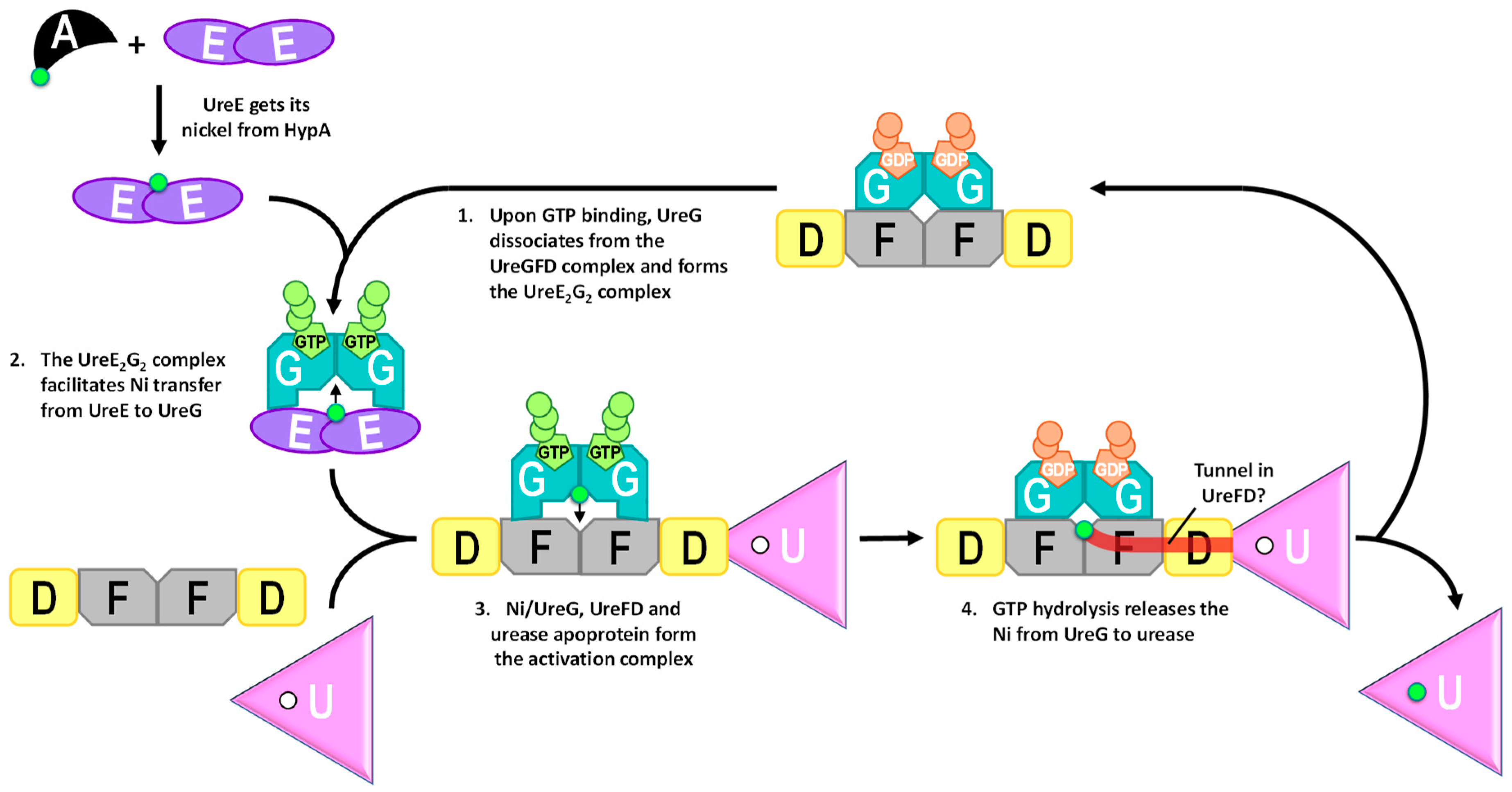
GTP-dependent conformational changes of UreG provide a mechanism where GTP binding/hydrolysis facilitates the delivery of nickel along the urease maturation pathway (Figure 6). It has been shown that the Ni/UreE dimer, providing the sole nickel source, can activate urease in the presence of UreGFD and GTP [51]. Binding of GTP induces UreG to dissociate from the UreGFD complex and bind with UreE to form the UreE2G2 complex. The UreE2G2 complex, which can also activate urease in the presence of UreFD complex [51], facilitates the transfer of nickel from UreE to UreG [52]. Direct protein–protein interactions among the urease accessory proteins are required as separating Ni/UreE and UreGFD/urease by a dialysis membrane abolished the urease activation in vitro [51]. After UreG gets its Ni2+, it can interact with UreFD and urease apoprotein to form the activation complex [43]. Mutagenesis studies suggested that UreG binds to UreFD in the activation complex [46,50]. That Ni/GTP-bound UreG, which dissociates from the UreFD complex, can interact with UreFD in the activation complex suggests that UreFD may undergo conformational changes upon binding of urease apoprotein to accommodate Ni/GTP-bound UreG in the activation complex. Whether UreE is involved in this activation complex is unclear. It has been shown that UreG can pull down UreE, UreD, UreF and urease [46,82]. Given that UreG can also interact with UreE, this observation did not conclusively demonstrate the interaction of UreE to the activation complex. Furthermore, it has been shown that urease can be activated in the presence of Ni/UreG, UreFD and urease apoprotein without UreE [51], suggesting that UreE is not essential in the formation of the activation complex.
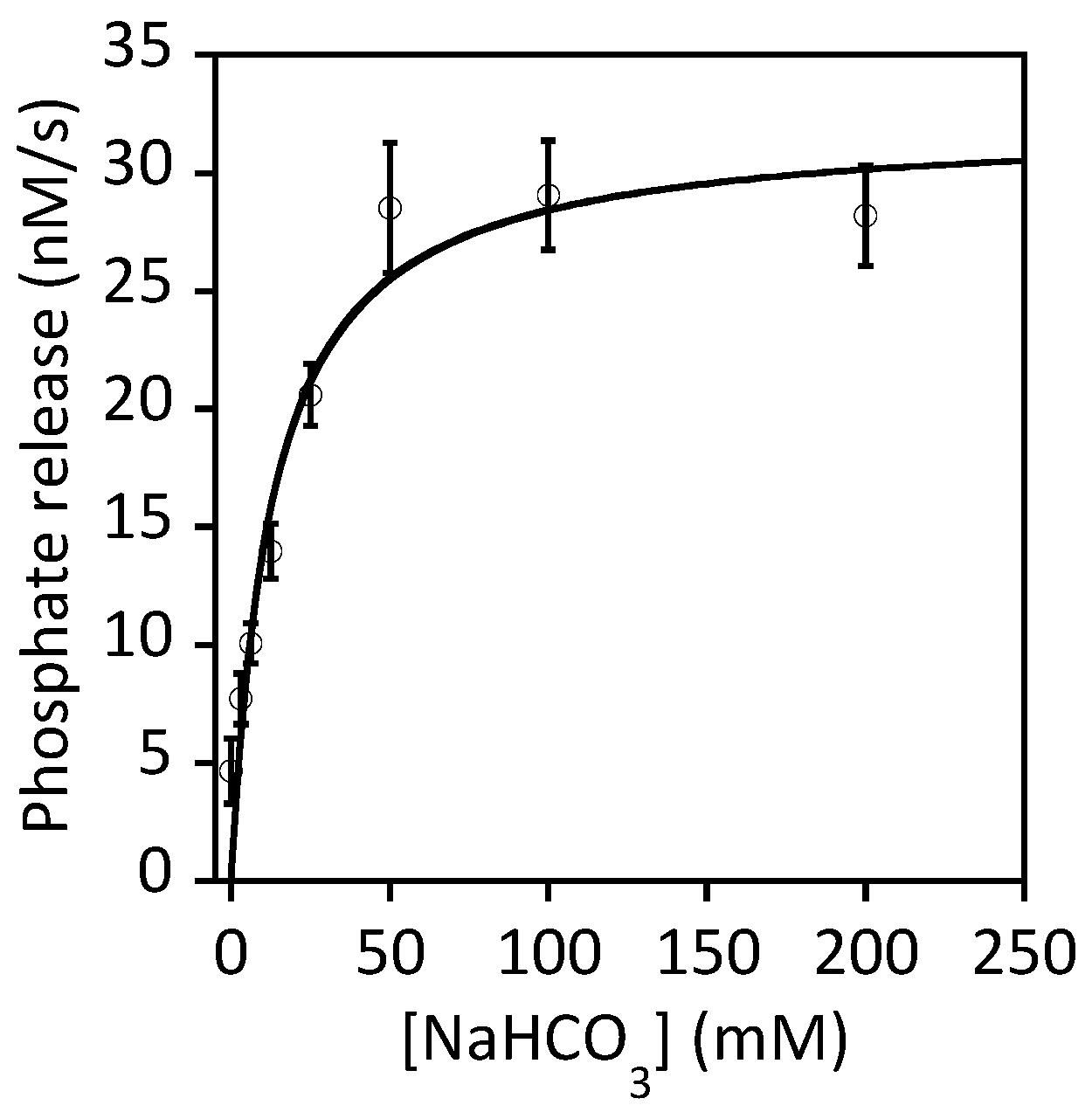
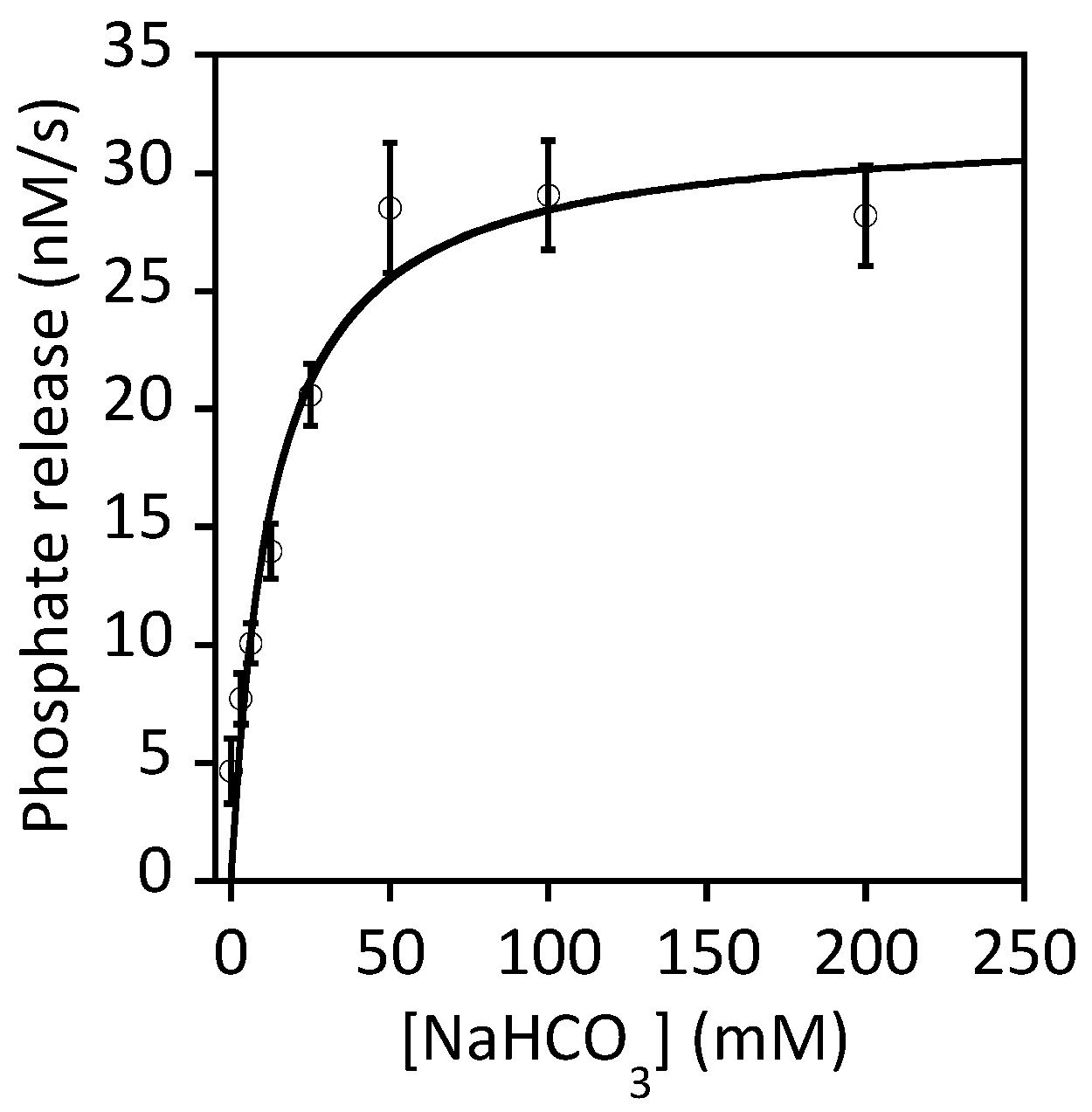
It is well established that GTP hydrolysis is essential to urease maturation [41,83]. Substituting the conserved Lys-20 and Thr-21 residues to alanine in the P-loop region of K. aerogenes UreG abolished its ability to bind ATP-linked agarose, the formation of the activation complex and urease maturation [45]. As the Ni/UreG dimer will dissociate into monomers and release the bound Ni2+ upon GTP hydrolysis [43], the GTPase activity of UreG must be tightly regulated to prevent nonproductive release of Ni2+ outside the activation complex. It has been shown that the GTPase activity is only detectable by addition of bicarbonate [43]. One hypothesis is that bicarbonate can serve as one of the substrates of UreG that results in the formation of an intermediate of a carboxy-phosphate [41]. The effect of bicarbonate on the UreG activity exhibited a classical Michaelis–Menten kinetics ([52], Figure 7). Using a saturated concentration of GTP, we estimated the Km value of bicarbonate was 13 ± 4 mM. In addition to bicarbonate, GTPase activity of UreG can also be increased by ~2–3 fold by addition of UreE, NH4+ or K+ [52]. Even under the most favorable conditions, the activity of UreG is still very low. In our hands, the turnover number (kcat) of H. pylori UreG is ~0.006 s−1 at 37 °C in the presence of K+ and saturated concentration of GTP (Figure 7). The low intrinsic activity of UreG should be of advantage as it prevents premature hydrolysis that releases the bound Ni2+ to the cytoplasm. It is unclear how and when the GTPase activity of UreG is activated. One possibility is that the GTPase activity of UreG is stimulated by the conformational changes induced during the formation of the activation complex. Conformational changes in the formation of the UreFD/urease complex have been suggested by cross-linking experiments [84].
GTP hydrolysis changes the conformation of the CPH motif in such a way that the square-planer coordination by Cys66 and His68 is disrupted (Figure 4), promoting the release of Ni2+ in the activation complex. Interestingly, it has been shown that interaction between UreG and UreF is essential to the urease maturation [46,48,50]. Mutations that broke the UreG–UreF interaction also abolished urease maturation, suggesting that UreG is likely bound to the activation complex via the UreFD complex.
As the nickel binding site of UreG is far away from the active site of urease, it is not fully understood how the Ni2+ is transferred from UreG to urease after GTP hydrolysis. One intriguing hypothesis suggested that the Ni2+ reaches urease through a tunnel within the core of the UreFD complex [82,85,86]. Large cavities have been identified in the structures of H. pylori UreFD complex [82,85,86]. The cavities form a tunnel, which is wide enough for Ni2+ to pass through, connecting the nickel binding site of UreG to an exit at UreD. The tunnel hypothesis is supported by: (1) Both UreF and UreD were able to bind Ni2+ [38,86]; (2) Tunnel-disrupting variants of UreD were shown to greatly reduce the urease maturation without affecting the UreD–urease interaction or the formation of the UreGFD complex [82,86]. As UreD is responsible for binding urease in the activation complex, the tunnel within the UreFD complex should provide a mechanism for the transfer of Ni2+ from UreG to the urease within the activation complex.
8. UreE Gets Its Nickel from Cross-Talking to the Hydrogenase Maturation Pathway
Hydrogenase maturation factors HypA and HypB, which are responsible for the delivery of nickel ions to the active site of [NiFe]-hydrogenase [88,89,90,91,92,93,94], are also essential for urease maturation [31,90]. Genes of hydrogenase maturation factors were first identified in Escherichia coli [94,95], where hypA and hypB are encoded in the hyp operon. In H. pylori, the two genes are located in different locations in the chromosome [96]. Urease activity was greatly reduced in hypA or hypB knockout strains of H. pylori [31,90]. The urease activity could be restored by addition of Ni2+ in the growth medium. These observations suggest that the hydrogenase maturation factors HypA and HypB are required for the urease maturation. Structures of HypA [97,98] revealed a mixed α/β topology containing a nickel binding domain with micromolar affinity [99,100,101,102], and a zinc binding domain [98,100,102,103,104,105] with nanomolar affinity [102]. Residues coordinating the Zn2+ [100,105] and Ni2+ [36,99,101] are essential for urease maturation. HypA interacts with the C-terminal domain of UreE [52,106,107,108,109] to form a 1:2 complex [52,101,107,108]. The complex formation creates a unique nanomolar affinity nickel-binding site, which is not found in any of the individual proteins [107]. Ni2+ can be transferred from HypA to UreE in the complex [52,108]. It has been shown that HypA and UreG compete with each other for UreE [109]. In the presence of Mg2+ and GTP, UreE dissociates from the HypA/UreE2 complex and interacts with UreG to form the UreE2G2 complex, suggesting the switching of the protein binding partners is GTP-dependent [52]. It has been demonstrated that Ni2+ can be transferred from HypA to UreE to UreG [52]. HypA, on the other hand, obtains its Ni2+ from HypB [60,110] with the help of SlyD [106,111,112,113,114] of the hydrogenase maturation pathway, which has been reviewed in [115,116].
9. Urease Maturation Pathway in Plants
In plants, urease and its accessory genes contain introns and are scattered on different chromosomes. The urease structural genes and homologues of UreD, UreF and UreG have been identified in plants [117,118]. Similar to the maturation of bacterial ureases, the formation of the UreGFD/urease activation complexes was shown to be essential to the maturation of ureases in rice (Oryza sativa) [119,120] and Arabidopsis [120,121]. Plant UreG, previously named as p32, is encoded by Eu3 [122]. It has a GTPase domain homologous to bacterial UreG and an histidine-rich N-terminal extension and a plant-specific region containing two HXH motifs [120]. The two HXH motifs, but not the histidine-rich extension, were found to be essential for urease activation [120]. Interestingly, no UreE homologue was found in plants. It was postulated that the plant UreG, with the plant specific HXH motifs, may combine the function of bacterial UreE and UreG into one protein [56,120,121,122].
10. Urease Maturation Pathway Is Druggable
Urease is a microbial virulence factor for gastric infection by H. pylori, urinary tract infection by Proteus mirabilis and Klebsiella strains, and lung infection by fungus Cryptococcus neoformans [123]. The first line treatment of H. pylori infection is combining proton-pump inhibitors with antibiotics in the triple therapy [124]. However, the emerging antibiotic resistance make the development of new drugs pressing. Recent research findings suggest that the urease maturation pathway could become a new druggable target [65,123]. Bismuth is a heavy metal that is effective for H. pylori eradication [124]. Recent study showed that colloidal bismuth subcitrate inhibits the biosynthesis of active H. pylori urease by displacing Ni2+ in UreG [125]. Virtual screening has identified two compounds that inhibit the GTPase activity of UreG, reduce urease activity with higher potency than clinically using acetohydroxamic acid, and suppress H. pylori infection in a mammalian cell model [125]. These results demonstrate that the urease maturation pathway is a novel druggable target for treatment of H. pylori infection.
11. Future Perspectives
While major progress has been made on how urease accessory proteins interplay to deliver Ni2+ to ureases, some interesting questions remain unanswered. (1) The structure of the activation complex is not known despite several structural models that have been proposed previously [126,127]. A high-resolution structure of the activation complex should provide novel structural insights into how GTPase activity of UreG is stimulated and the mechanism of nickel transfer within the complex. (2) The mechanism of carbamylation of the active site lysine is relatively understudied. Does it take place before or after the insertion of Ni2+? Is the carbamylation reaction catalyzed by other factors? (3) Urease requires two Ni2+ to function. Since only one Ni2+ ion is carried per UreG dimer, it takes two round trips of UreG to activate urease. MBP-UreD and MBP-UreFD were shown to dissociate from urease after activation [37,39]. It is unclear what triggers the dissociation of the activation complex after urease activation. (4) What could be the driving force for the transfer of Ni2+ from UreG to the urease through the tunnel? How is the free energy change of GTP hydrolysis coupled to the nickel transfer? (5) Unlike the release of Ni2+ from UreG that can be explained by GTP hydrolysis, it is not known why nickel is transferred unidirectionally from HypA to UreE and from UreE to UreG. Structure determination of the HypA/UreE2 and UreE2G2 complexes should help to provide structural insights to this question.
12. Conclusions
The urease maturation pathway represents the most well-studied paradigm on how nickel ions are delivered from one protein to another through specific protein–protein interactions. One central theme depicts the binding/hydrolysis of GTP allosterically regulate the switching of protein-binding partners and the transfer of nickel ions from UreE → UreG → UreF/UreD → urease. This mechanism ensures that the nickel ions are always protein-bound to avoid leaking of the toxic metal to the cytoplasm. UreE also receives its nickel ions from interacting with HypA of the hydrogenase maturation pathway. A better understanding of the urease maturation pathway allow us to develop drugs against the H. pylori infection.
Funding
This work was funded by the grants from the Research Grants Council of Hong Kong (14117314, AoE/M-05/12, and AoE/M-403/16) and from the Research Committee of The Chinese University of Hong Kong (3132814 and 3132815).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Krajewska, B. Ureases I. Functional, Catalytic and Kinetic Properties: A Review. J. Mol. Catal. B Enzym. 2009, 59, 9–21. [Google Scholar] [CrossRef]
- Hanlon, D.P. The Distribution of Arginase and Urease in Marine Invertebrates. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1975, 52, 261–264. [Google Scholar] [CrossRef]
- Alonso-Saez, L.; Waller, A.S.; Mende, D.R.; Bakker, K.; Farnelid, H.; Yager, P.L.; Lovejoy, C.; Tremblay, J.-E.; Potvin, M.; Heinrich, F.; et al. Role for Urea in Nitrification by Polar Marine Archaea. Proc. Natl. Acad. Sci. USA 2012, 109, 17989–17994. [Google Scholar] [CrossRef] [PubMed]
- Dixon, N.E.; Gazzola, C.; Watters, J.J.; Blakeley, R.L.; Zerner, B. Jack Bean Urease (EC 3.5.1.5). A Metalloenzyme. A Simple Biological Role for Nickel? J. Am. Chem. Soc. 1975, 97, 4131–4133. [Google Scholar] [CrossRef] [PubMed]
- Sumner, J.B. The Isolation and Crystallization of the Enzyme Urease. J. Biol. Chem. 1926, 69, 435–442. [Google Scholar]
- Kao, C.-Y.; Sheu, B.-S.; Wu, J.-J. Helicobacter pylori infection: An overview of bacterial virulence facztors and pathogenesis. Biomed. J. 2016, 39, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Ishaq, S.; Nunn, L. Helicobacter pylori and Gastric Cancer: A State of the Art Review. Gastroenterol. Hepatol. Bed Bench 2015, 8, S6–S14. [Google Scholar] [PubMed]
- Narayanan, M.; Reddy, K.M.; Marsicano, E. Peptic Ulcer Disease and Helicobacter pylori Infection. Mo. Med. 2018, 115, 219–224. [Google Scholar] [PubMed]
- Carter, E.L.; Tronrud, D.E.; Taber, S.R.; Karplus, P.A.; Hausinger, R.P. Iron-Containing Urease in a Pathogenic Bacterium. Proc. Natl. Acad. Sci. USA 2011, 108, 13095–13099. [Google Scholar] [CrossRef]
- Capdevila, D.A.; Edmonds, K.A.; Giedroc, D.P. Metallochaperones and Metalloregulation in Bacteria. Essays Biochem. 2017, 61, 177–200. [Google Scholar] [CrossRef]
- Waldron, K.J.; Robinson, N.J. How Do Bacterial Cells Ensure That Metalloproteins Get the Correct Metal? Nat. Rev. Microbiol. 2009, 7, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Kappaun, K.; Piovesan, A.R.; Carlini, C.R.; Ligabue-Braun, R. Ureases: Historical Aspects, Catalytic, and Non-Catalytic Properties—A Review. J. Adv. Res. 2018, 13, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Todd, M.J.; Hausinger, R.P. Purification and Characterization of the Nickel-Containing Multicomponent Urease from Klebsiella aerogenes. J. Biol. Chem. 1987, 262, 5963–5967. [Google Scholar] [PubMed]
- Hu, L.T.; Mobley, H.L. Purification and N-Terminal Analysis of Urease from Helicobacter pylori. Infect. Immun. 1990, 58, 992–998. [Google Scholar] [PubMed]
- Blakeley, R.L.; Zerner, B. Jack Bean Urease: The First Nickel Enzyme. J. Mol. Catal. 1984, 23, 263–292. [Google Scholar] [CrossRef]
- Pearson, M.A.; Michel, L.O.; Hausinger, R.P.; Karplus, P.A. Structures of Cys319 Variants and Acetohydroxamate-Inhibited Klebsiella aerogenes Urease. Biochemistry 1997, 36, 8164–8172. [Google Scholar] [CrossRef] [PubMed]
- Jabri, E.; Carr, M.B.; Hausinger, R.P.; Karplus, P.A. The Crystal Structure of Urease from Klebsiella aerogenes. Science 1995, 268, 998–1004. [Google Scholar] [CrossRef]
- Benini, S.; Rypniewski, W.R.; Wilson, K.S.; Miletti, S.; Ciurli, S.; Mangani, S. A New Proposal for Urease Mechanism Based on the Crystal Structures of the Native and Inhibited Enzyme from Bacillus pasteurii: Why Urea Hydrolysis Costs Two Nickels. Structure 1999, 7, 205–216. [Google Scholar] [CrossRef]
- Balasubramanian, A.; Ponnuraj, K. Crystal Structure of the First Plant Urease from Jack Bean: 83 Years of Journey from Its First Crystal to Molecular Structure. J. Mol. Biol. 2010, 400, 274–283. [Google Scholar] [CrossRef]
- Ha, N.; Oh, S.; Sung, J.Y.; Cha, K.A.; Lee, M.H.; Oh, B. Supramolecular Assembly and Acid Resistance of Helicobacter pylori Urease. Nat. Struct. Mol. Biol. 2001, 8, 505–509. [Google Scholar] [CrossRef]
- Ciurli, S.; Benini, S.; Rypniewski, W.R.; Wilson, K.S.; Miletti, S.; Mangani, S. Structural Properties of the Nickel Ions in Urease: Novel Insights into the Catalytic and Inhibition Mechanisms. Coord. Chem. Rev. 1999, 190–192, 331–355. [Google Scholar] [CrossRef]
- Park, I.S.; Hausinger, R.P. Requirement of Carbon Dioxide for in vitro Assembly of the Urease Nickel Metallocenter. Science 1995, 267, 1156–1158. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.A.; Schaller, R.A.; Michel, L.O.; Karplus, P.A.; Hausinger, R.P. Chemical Rescue of Klebsiella aerogenes Urease Variants Lacking the Carbamylated-Lysine Nickel Ligand. Biochemistry 1998, 37, 6214–6220. [Google Scholar] [CrossRef] [PubMed]
- Park, I.S.; Hausinger, R.P. Metal Ion Interactions with Urease and UreD-Urease Apoproteins. Biochemistry 1996, 35, 5345–5352. [Google Scholar] [CrossRef] [PubMed]
- Mulrooney, S.B.; Hausinger, R.P. Sequence of the Klebsiella aerogenes Urease Genes and Evidence for Accessory Proteins Facilitating Nickel Incorporation. J. Bacteriol. 1990, 172, 5837–5843. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Mulrooney, S.B.; Renner, M.J.; Markowicz, Y.; Hausinger, R.P. Klebsiella aerogenes Urease Gene Cluster: Sequence of UreD and Demonstration That Four Accessory Genes (UreD, UreE, UreF, and UreG) Are Involved in Nickel Metallocenter Biosynthesis. J. Bacteriol. 1992, 174, 4324–4330. [Google Scholar] [CrossRef] [PubMed]
- Akada, J.K.; Shirai, M.; Takeuchi, H.; Tsuda, M.; Nakazawa, T. Identification of the Urease Operon in Helicobacter pylori and Its Control by mRNA Decay in Response to pH. Mol. Microbiol. 2000, 36, 1071–1084. [Google Scholar] [CrossRef]
- Cussac, V.; Ferrero, R.L.; Labigne, A. Expression of Helicobacter pylori Urease Genes in Escherichia coli Grown under Nitrogen-Limiting Conditions. J. Bacteriol. 1992, 174, 2466–2473. [Google Scholar] [CrossRef]
- Weeks, D.L.; Eskandari, S.; Scott, D.R.; Sachs, G. A H+-Gated Urea Channel: The Link between Helicobacter pylori Urease and Gastric Colonization. Science 2000, 287, 482–485. [Google Scholar] [CrossRef]
- Rektorschek, M.; Buhmann, A.; Weeks, D.; Schwan, D.; Bensch, K.W.; Eskandari, S.; Scott, D.; Sachs, G.; Melchers, K. Acid Resistance of Helicobacter pylori Depends on the UreI Membrane Protein and an Inner Membrane Proton Barrier. Mol. Microbiol. 2000, 36, 141–152. [Google Scholar] [CrossRef]
- Benoit, S.L.; Zbell, A.L.; Maier, R.J. Nickel Enzyme Maturation in Helicobacter hepaticus: Roles of Accessory Proteins in Hydrogenase and Urease Activities. Microbiology 2007, 153, 3748–3756. [Google Scholar] [CrossRef] [PubMed]
- Sriwanthana, B.; Island, M.D.; Maneval, D.; Mobley, H.L.T. Single-Step Purification of Proteus mirabilis Urease Accessory Protein UreE, a Protein with a Naturally Occurring Histidine Tail, by Nickel Chelate Affinity Chromatography. J. Bacteriol. 1994, 176, 6836–6841. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Foxall, P.A.; Russell, R.; Mobley, H.L.T. Purification of Recombinant Helicobacter pylori Urease Apoenzyme Encoded by UreA and UreB. Infect. Immun. 1992, 60, 2657–2667. [Google Scholar] [PubMed]
- Lee, M.H.; Mulrooney, S.B.; Hausinger, R.P. Purification, Characterization, and in vivo Reconstitution of Klebsiella aerogenes Urease Apoenzyme. J. Bacteriol. 1990, 172, 4427–4431. [Google Scholar] [CrossRef] [PubMed]
- Voland, P.; Weeks, D.L.; Marcus, E.A.; Prinz, C.; Sachs, G.; Scott, D. Interactions among the Seven Helicobacter pylori Proteins Encoded by the Urease Gene Cluster. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 284, G96–G106. [Google Scholar] [CrossRef] [PubMed]
- Benoit, S.L.; Mehta, N.; Weinberg, M.V.; Maier, C.; Maier, R.J. Interaction between the Helicobacter pylori Accessory Proteins HypA and UreE Is Needed for Urease Maturation. Microbiology 2007, 153, 1474–1482. [Google Scholar] [CrossRef] [PubMed]
- Park, I.S.; Carr, M.B.; Hausinger, R.P. In vitro Activation of Urease Apoprotein and Role of UreD as a Chaperone Required for Nickel Metallocenter Assembly. Proc. Natl. Acad. Sci. USA 1994, 91, 3233–3237. [Google Scholar] [CrossRef] [PubMed]
- Carter, E.L.; Hausinger, R.P. Characterization of the Klebsiella aerogenes Urease Accessory Protein UreD in Fusion with the Maltose Binding Protein. J. Bacteriol. 2010, 192, 2294–2304. [Google Scholar] [CrossRef]
- Moncrief, M.B.C.; Hausinger, R.P. Purification and Activation Properties of UreD-UreF-Urease Apoprotein Complexes. J. Bacteriol. 1996, 178, 5417–5421. [Google Scholar] [CrossRef]
- Kim, J.K.; Mulrooney, S.B.; Hausinger, R.P. The UreEF Fusion Protein Provides a Soluble and Functional Form of the UreF Urease Accessory Protein. J. Bacteriol. 2006, 188, 8413–8420. [Google Scholar] [CrossRef] [Green Version]
- Soriano, A.; Hausinger, R.P. GTP-Dependent Activation of Urease Apoprotein in Complex with the UreD, UreF, and UreG Accessory Proteins. Proc. Natl. Acad. Sci. USA 1999, 96, 11140–11144. [Google Scholar] [CrossRef] [PubMed]
- Park, I.; Hausinger, R.P. Evidence for the Presence of Urease Apoprotein Complexes Containing UreD, UreF, and UreG in Cells That Are Competent for in vivo Enzyme Activation. J. Bacteriol. 1995, 177, 1947–1951. [Google Scholar] [CrossRef] [PubMed]
- Fong, Y.H.; Wong, H.C.; Yuen, M.H.; Lau, P.H.; Chen, Y.W.; Wong, K.-B. Structure of UreG/UreF/UreH Complex Reveals How Urease Accessory Proteins Facilitate Maturation of Helicobacter pylori Urease. PLoS Biol. 2013, 11, e1001678. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, M.A.; Han, L.; Zhong, Y.; Boer, J.L.; Ruotolo, B.T.; Hausinger, R.P. Analysis of a Soluble (UreD:UreF:UreG)2 Accessory Protein Complex and Its Interactions with Klebsiella aerogenes Urease by Mass Spectrometry. J. Am. Soc. Mass Spectrom. 2013, 24, 1328–1337. [Google Scholar] [CrossRef] [PubMed]
- Moncrief, M.B.C.; Hausinger, R.P. Characterization of UreG, Identification of a UreD-UreF-UreG Complex, and Evidence Suggesting That a Nucleotide-Binding Site in UreG Is Required for in vivo Metallocenter Assembly of Klebsiella aerogenes Urease. J. Bacteriol. 1997, 179, 4081–4086. [Google Scholar] [CrossRef]
- Boer, J.L.; Quiroz-Valenzuela, S.; Anderson, K.L.; Hausinger, R.P. Mutagenesis of Klebsiella aerogenes UreG to Probe Nickel Binding and Interactions with Other Urease-Related Proteins. Biochemistry 2010, 49, 5859–5869. [Google Scholar] [CrossRef]
- Rain, J.C.; Selig, L.; De Reuse, H.; Battaglia, V.; Reverdy, C.; Simon, S.; Lenzen, G.; Petel, F.; Wojcik, J.; Schächter, V.; et al. The Protein-Protein Interaction Map of Helicobacter pylori. Nature 2001, 409, 211–215. [Google Scholar] [CrossRef]
- Fong, Y.H.; Wong, H.C.; Chuck, C.P.; Chen, Y.W.; Sun, H.; Wong, K.-B. Assembly of Preactivation Complex for Urease Maturation in Helicobacter pylori: Crystal Structure of UreF-UreH Protein Complex. J. Biol. Chem. 2011, 286, 43241–43249. [Google Scholar] [CrossRef]
- Lam, R.; Romanov, V.; Johns, K.; Battaile, K.P.; Wu-Brown, J.; Guthrie, J.L.; Hausinger, R.P.; Pai, E.F.; Chirgadze, N.Y. Crystal Structure of a Truncated Urease Accessory Protein UreF from Helicobacter pylori. Proteins 2010, 78, 2839–2848. [Google Scholar] [CrossRef]
- Boer, J.L.; Hausinger, R.P. Klebsiella aerogenes UreF: Identification of the UreG Binding Site and Role in Enhancing the Fidelity of Urease Activation. Biochemistry 2012, 51, 2298–2308. [Google Scholar] [CrossRef]
- Yuen, M.H.; Fong, Y.H.; Nim, Y.S.; Lau, P.H.; Wong, K.-B. Structural Insights into how GTP-Dependent Conformational Changes in a Metallochaperone UreG Facilitate Urease Maturation. Proc. Natl. Acad. Sci. USA 2017, 114, E10890–E10898. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, H.; Lai, T.-P.; Sun, H. UreE-UreG Complex Facilitates Nickel Transfer and Preactivates GTPase of UreG in Helicobacter pylori. J. Biol. Chem. 2015, 290, 12474–12485. [Google Scholar] [CrossRef] [PubMed]
- Leipe, D.D.; Wolf, Y.I.; Koonin, E.V.; Aravind, L. Classification and Evolution of P-Loop GTPases and Related ATPases. J. Mol. Biol. 2002, 317, 41–72. [Google Scholar] [CrossRef] [PubMed]
- Zambelli, B.; Turano, P.; Musiani, F.; Neyroz, P.; Ciurli, S. Zn2+-Linked Dimerization of UreG from Helicobacter pylori, a Chaperone Involved in Nickel Trafficking and Urease Activation. Proteins Struct. Funct. Bioinform. 2009, 74, 222–239. [Google Scholar] [CrossRef] [PubMed]
- Zambelli, B.; Stola, M.; Musiani, F.; De Vriendt, K.; Samyn, B.; Devreese, B.; Van Beeumen, J.; Turano, P.; Dikiy, A.; Bryant, D.A.; et al. UreG, a Chaperone in the Urease Process, Is an Intrinsically Unstructured Protein Binding a Single Zn2+ Ion. J. Biol. Chem. 2005, 280, 4684–4695. [Google Scholar] [CrossRef] [PubMed]
- Real-Guerra, R.; Staniscuaski, F.; Zambelli, B.; Musiani, F.; Ciurli, S.; Carlini, C.R. Biochemical and Structural Studies on Native and Recombinant Glycine max UreG: A Detailed Characterization of a Plant Urease Accessory Protein. Plant Mol. Biol. 2012, 78, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Miraula, M.; Ciurli, S.; Zambelli, B. Intrinsic Disorder and Metal Binding in UreG Proteins from Archae Hyperthermophiles: GTPase Enzymes Involved in the Activation of Ni(II) Dependent Urease. J. Biol. Inorg. Chem. 2015, 20, 739–755. [Google Scholar] [CrossRef] [PubMed]
- Martin-Diaconescu, V.; Bellucci, M.; Musiani, F.; Ciurli, S.; Maroney, M.J. Unraveling the Helicobacter pylori UreG Zinc Binding Site Using X-Ray Absorption Spectroscopy (XAS) and Structural Modeling. J. Biol. Inorg. Chem. 2012, 17, 353–361. [Google Scholar] [CrossRef]
- Xia, W.; Li, H.; Yang, X.; Wong, K.B.; Sun, H. Metallo-GTPase HypB from Helicobacter pylori and Its Interaction with Nickel Chaperone Protein HypA. J. Biol. Chem. 2012, 287, 6753–6763. [Google Scholar] [CrossRef] [PubMed]
- Lacasse, M.J.; Douglas, C.D.; Zamble, D.B. Mechanism of Selective Nickel Transfer from HypB to HypA, Escherichia coli [NiFe]-Hydrogenase Accessory Proteins. Biochemistry 2016, 55, 6821–6831. [Google Scholar] [CrossRef] [PubMed]
- Gasper, R.; Scrima, A.; Wittinghofer, A. Structural Insights into HypB, a GTP-Binding Protein That Regulates Metal Binding. J. Biol. Chem. 2006, 281, 27492–27502. [Google Scholar] [CrossRef] [PubMed]
- Cai, F.; Ngu, T.T.; Kaluarachchi, H.; Zamble, D.B. Relationship between the GTPase, Metal-Binding, and Dimerization Activities of E. Coli HypB. J. Inorg. Biochem. 2011, 16, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.H.; Lee, K.M.; Wong, K.B. Interaction between Hydrogenase Maturation Factors HypA and HypB Is Required for [NiFe]-Hydrogenase Maturation. PLoS ONE 2012, 7, e32592. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.H.; Li, T.; Wong, C.O.; Wong, K.B. Structural Basis for GTP-Dependent Dimerization of Hydrogenase Maturation Factor HypB. PLoS ONE 2012, 7, e30547. [Google Scholar] [CrossRef] [PubMed]
- Remaut, H.; Safarov, N.; Ciurli, S.; Van Beeumen, J. Structural Basis for Ni2+ Transport and Assembly of the Urease Active Site by the Metallochaperone UreE from Bacillus pasteurii. J. Biol. Chem. 2001, 276, 49365–49370. [Google Scholar] [CrossRef]
- Song, H.K.; Mulrooney, S.B.; Huber, R.; Hausinger, R.P. Crystal Structure of Klebsiella aerogenes UreE, a Nickel-Binding Metallochaperone for Urease Activation. J. Biol. Chem. 2001, 276, 49359–49364. [Google Scholar] [CrossRef]
- Banaszak, K.; Martin-Diaconescu, V.; Bellucci, M.; Zambelli, B.; Rypniewski, W.; Maroney, M.J.; Ciurli, S. Crystallographic and X-Ray Absorption Spectroscopic Characterization of Helicobacter pylori UreE Bound to Ni2+ and Zn2+ Reveals a Role for the Disordered C-Terminal Arm in Metal Trafficking. Biochem. J. 2012, 441, 1017–1026. [Google Scholar] [CrossRef]
- Shi, R.; Munger, C.; Asinas, A.; Benoit, S.L.; Miller, E.; Matte, A.; Maier, R.J.; Cygler, M. Crystal Structures of apo and Metal-Bound Forms of the UreE Protein from Helicobacter pylori: Role of Multiple Metal Binding Sites. Biochemistry 2010, 49, 7080–7088. [Google Scholar] [CrossRef]
- Stola, M.; Musiani, F.; Mangani, S.; Turano, P.; Safarov, N.; Zambelli, B.; Ciurli, S. The Nickel Site of Bacillus pasteurii UreE, a Urease Metallo-Chaperone, as Revealed by Metal-Binding Studies and X-Ray Absorption Spectroscopy. Biochemistry 2006, 45, 6495–6509. [Google Scholar] [CrossRef]
- Lee, M.H.; Pankratz, H.S.; Wang, S.; Scott, R.A.; Finnegan, M.G.; Johnson, M.K.; Ippolito, J.A.; Christianson, D.W.; Hausinger, R.P. Purification and Characterization of Klebsiella aerogenes UreE Protein: A Nickel-binding Protein That Functions in Urease Metallocenter Assembly. Protein Sci. 1993, 2, 1042–1052. [Google Scholar] [CrossRef]
- Bellucci, M.; Zambelli, B.; Musiani, F.; Turano, P.; Ciurli, S. Helicobacter pylori UreE, a Urease Accessory Protein: Specific Ni2+- and Zn2+-Binding Properties and Interaction with Its Cognate UreG. Biochem. J. 2009, 422, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Grossoehme, N.E.; Mulrooney, S.B.; Hausinger, R.P.; Wilcox, D.E. Thermodynamics of Ni2+, Cu2+, and Zn2+ Binding to the Urease Metallochaperone. Biochemistry 2007, 46, 10506–10516. [Google Scholar] [CrossRef] [PubMed]
- Won, H.; Lee, Y.; Kim, J.; Shin, I.S.; Lee, M.H.; Lee, B. Structural Characterization of the Nickel-Binding Properties of Bacillus pasteurii Urease Accessory Protein (Ure)E in Solution. J. Biol. Chem. 2004, 279, 17466–17472. [Google Scholar] [CrossRef] [PubMed]
- Ciurli, S.; Safarov, N.; Miletti, S.; Dikiy, A.; Christensen, S.K.; Kornetzky, K.; Bryant, D.A.; Vandenberghe, I.; Devreese, B.; Samyn, B.; et al. Molecular Characterization of Bacillus pasteurii UreE, a Metal-Binding Chaperone for the Assembly of the Urease Active Site. J. Biol. Inorg. Chem. 2002, 7, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Colpas, G.J.; Hausinger, R.P. In vivo and in citro Kinetics of Metal Transfer by the Klebsiella aerogenes Urease Nickel Metallochaperone, UreE. J. Biol. Chem. 2000, 275, 10731–10737. [Google Scholar] [CrossRef] [PubMed]
- Colpas, G.J.; Brayman, T.G.; Ming, L.J.; Hausinger, R.P. Identification of Metal-Binding Residues in the Klebsiella aerogenes Urease Nickel Metallochaperone, UreE. Biochemistry 1999, 38, 4078–4088. [Google Scholar] [CrossRef] [PubMed]
- Brayman, T.G.; Hausinger, R.P. Purification, Characterization, and Functional Analysis of a Truncated Klebsiella aerogenes UreE Urease Accessory Protein Lacking the Histidine-Rich Carboxyl Terminus. J. Bacteriol. 1996, 178, 5410–5416. [Google Scholar] [CrossRef] [PubMed]
- Mulrooney, S.B.; Ward, S.K.; Hausinger, R.P. Purification and Properties of the Klebsiella aerogenes UreE Metal-Binding Domain, a Functional Metallochaperone of Urease. J. Bacteriol. 2005, 187, 3581–3585. [Google Scholar] [CrossRef]
- Musiani, F.; Zambelli, B.; Stola, M.; Ciurli, S. Nickel Trafficking: Insights into the Fold and Function of UreE, a Urease Metallochaperone. J. Inorg. Biochem. 2004, 98, 803–813. [Google Scholar] [CrossRef]
- Merloni, A.; Dobrovolska, O.; Zambelli, B.; Agostini, F.; Bazzani, M.; Musiani, F.; Ciurli, S. Molecular Landscape of the Interaction between the Urease Accessory Proteins UreE and UreG. Biochim. Biophys. Acta Proteins Proteom. 2014, 1844, 1662–1674. [Google Scholar] [CrossRef]
- Jurrus, E.; Engel, D.; Star, K.; Monson, K.; Brandi, J.; Felberg, L.E.; Brookes, D.H.; Wilson, L.; Chen, J.; Liles, K.; et al. Improvements to the APBS Biomolecular Solvation Software Suite. Protein Sci. 2018, 27, 112–128. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, M.A.; Wang, B.; Feig, M.; Hausinger, R.P. Mutational and Computational Evidence That a Nickel-Transfer Tunnel in UreD Is Used for Activation of Klebsiella aerogenes Urease. Biochemistry 2015, 54, 6392–6401. [Google Scholar] [CrossRef] [PubMed]
- Soriano, A.; Colpas, G.J.; Hausinger, R.P. UreE Stimulation of GTP-Dependent Urease Activation in the UreD-UreF-UreG-Urease Apoprotein Complex. Biochemistry 2000, 39, 12435–12440. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Kuchar, J.; Hausinger, R.P. Chemical Cross-Linking and Mass Spectrometric Identification of Sites of Interaction for UreD, UreF, and Urease. J. Biol. Chem. 2004, 279, 15305–15313. [Google Scholar] [CrossRef]
- Musiani, F.; Gioia, D.; Masetti, M.; Falchi, F.; Cavalli, A.; Recanatini, M.; Ciurli, S. Protein Tunnels: The Case of Urease Accessory Proteins. J. Chem. Theory Comput. 2017, 13, 2322–2331. [Google Scholar] [CrossRef]
- Zambelli, B.; Berardi, A.; Martin-Diaconescu, V.; Mazzei, L.; Musiani, F.; Maroney, M.J.; Ciurli, S. Nickel Binding Properties of Helicobacter pylori UreF, an Accessory Protein in the Nickel-Based Activation of Urease. J. Biol. Inorg. Chem. 2014, 19, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Baykov, A.A.; Evtushenko, O.A.; Avaeva, S.M. A Malachite Green Procedure for Orthophosphate Determination and Its Use in Alkaline Phosphatase-Based Enzyme Immunoassay. Anal. Biochem. 1988, 171, 266–270. [Google Scholar] [CrossRef]
- Maier, T.; Lottspeich, F.; Böck, A. GTP Hydrolysis by HypB Is Essential for Nickel Insertion into Hydrogenases of Escherichia coli. Eur. J. Biochem. 1995, 230, 133–138. [Google Scholar] [CrossRef]
- Olson, J.W.; Fu, C.; Maier, R.J. The HypB Protein from Bradyrhizobium japonicum can Store Nickel and Is Required for the Nickel-dependent Transcriptional Regulation of Hydrogenase. Mol. Microbiol. 1997, 24, 119–128. [Google Scholar] [CrossRef]
- Olson, J.W.; Mehta, N.S.; Maier, R.J. Requirement of Nickel Metabolism Proteins HypA and HypB for Full Activity of Both Hydrogenase and Urease in Helicobacter pylori. Mol. Microbiol. 2001, 39, 176–182. [Google Scholar] [CrossRef]
- Hube, M.; Blokesch, M.; Böck, A. Network of Hydrogenase Maturation in Escherichia coli: Role of Accessory Proteins HypA and HybF. J. Bacteriol. 2002, 184, 3879–3885. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, D.; Gutekunst, K.; Klissenbauer, M.; Schulz-Friedrich, R.; Appel, J. Mutagenesis of Hydrogenase Accessory Genes of Synechocystis sp. PCC 6803: Additional Homologues of hypA and hypB Are Not Active in Hydrogenase Maturation. FEBS J. 2006, 273, 4516–4527. [Google Scholar] [CrossRef] [PubMed]
- Maier, T.; Jacobi, A.; Sauter, M.; Böck, A. The Product of the HypB Gene, Which Is Required for Nickel Incorporation into Hydrogenases, Is a Novel Guanine Nucleotide-Binding Protein. J. Bacteriol. 1993, 175, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Jacobi, A.; Rossmann, R.; Böck, A. The hyp Operon Gene Products Are Required for the Maturation of Catalytically Active Hydrogenase Isoenzymes in Escherichia coli. Arch. Microbiol. 1992, 158, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Lutz, S.; Jacobi, A.; Schlensog, V.; Böhm, R.; Sawers, G.; Böck, A. Molecular Characterization of an Operon (hyp) Necessary for the Activity of the Three Hydrogenase Isoenzymes in Escherichia coli. Mol. Microbiol. 1991, 5, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Tomb, J.F.; White, O.; Kerlavage, A.R.; Clayton, R.A.; Sutton, G.G.; Fleischmann, R.D.; Ketchum, K.A.; Klenk, H.P.; Gill, S.; Brian, A.D. The Complete Genome Sequence of the Gastric Pathogen Helicobacter pylori. Nature 1997, 388, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Spronk, C.A.E.M.; Żerko, S.; Górka, M.; Koźmiński, W.; Bardiaux, B.; Zambelli, B.; Musiani, F.; Piccioli, M.; Basak, P.; Blum, F.C.; et al. Structure and Dynamics of Helicobacter pylori Nickel-chaperone HypA: An Integrated Approach Using NMR Spectroscopy, Functional Assays and Computational Tools. JBIC J. Biol. Inorg. Chem. 2018, 23, 1309–1330. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Li, H.; Sze, K.H.; Sun, H. Structure of a Nickel Chaperone, HypA, from Helicobacter pylori Reveals Two Distinct Metal Binding Sites. J. Am. Chem. Soc. 2009, 131, 10031–10040. [Google Scholar] [CrossRef]
- Mehta, N.; Olson, J.W.; Maier, R.J. Characterization of Helicobacter pylori Nickel Metabolism Accessory Proteins Needed for Maturation of Both Urease and Hydrogenase. J. Bacteriol. 2003, 185, 726–734. [Google Scholar] [CrossRef]
- Herbst, R.W.; Perovic, I.; Martin-diaconescu, V.; Brien, K.O.; Chivers, P.T.; Pochapsky, S.S.; Pochapsky, T.C.; Maroney, M.J. Communication between the Zinc and Nickel Sites in Dimeric HypA: Metal Recognition and pH Sensing. J. Am. Chem. Soc. 2010, 132, 10338–10351. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.Q.; Johnson, R.C.; Merrell, D.S.; Maroney, M.J. Nickel Ligation of the N-Terminal Amine of HypA Is Required for Urease Maturation in Helicobacter pylori. Biochemistry 2017, 56, 1105–1116. [Google Scholar] [CrossRef] [PubMed]
- Atanassova, A.; Zamble, D.B. Escherichia coli HypA Is a Zinc Metalloprotein with a Weak Affinity for Nickel. J. Bacteriol. 2005, 187, 4689–4697. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Arai, T.; Matsumi, R.; Atomi, H.; Imanaka, T.; Miki, K. Crystal Structure of HypA, a Nickel-Binding Metallochaperone for [NiFe] Hydrogenase Maturation. J. Mol. Biol. 2009, 394, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.C.; Herbst, R.W.; Iwig, J.S.; Chivers, P.T.; Maroney, M.J. A Dynamic Zn Site in Helicobacter pylori HypA: A Potential Mechanism for Metal-Specific Protein Activity. J. Am. Chem. Soc. 2007, 129, 16–17. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.C.; Hu, H.Q.; Merrell, D.S.; Maroney, M.J. Dynamic HypA Zinc Site Is Essential for Acid Viability and Proper Urease Maturation in Helicobacter pylori. Metallomics 2015, 7, 674–682. [Google Scholar] [CrossRef]
- Stingl, K.; Schauer, K.; Ecobichon, C.; Labigne, A.; Lenormand, P.; Rousselle, J.-C.; Namane, A.; de Reuse, H. In vivo Interactome of Helicobacter pylori Urease Revealed by Tandem Affinity Purification. Mol. Cell. Proteom. 2008, 7, 2429–2441. [Google Scholar] [CrossRef]
- Hu, H.Q.; Huang, H.T.; Maroney, M.J. The Helicobacter pylori HypA·UreE2 Complex Contains a Novel High-Affinity Ni(II)-Binding Site. Biochemistry 2018, 57, 2932–2942. [Google Scholar] [CrossRef]
- Yang, X.; Li, H.; Cheng, T.; Xia, W.; Lai, Y.-T.; Sun, H. Nickel Translocation between Metallochaperones HypA and UreE in Helicobacter pylori. Metallomics 2014, 6, 40–42. [Google Scholar] [CrossRef]
- Benoit, S.L.; McMurry, J.L.; Hill, S.A.; Maier, R.J. Helicobacter pylori Hydrogenase Accessory Protein HypA and Urease Accessory Protein UreG Compete with Each Other for UreE Recognition. Biochim. Biophys. Acta-Gen. Subj. 2012, 1820, 1519–1525. [Google Scholar] [CrossRef]
- Douglas, C.D.; Ngu, T.T.; Kaluarachchi, H.; Zamble, D.B. Metal Transfer within the Escherichia coli HypB−HypA Complex of Hydrogenase Accessory Proteins. Biochemistry 2013, 52, 6030–6039. [Google Scholar] [CrossRef]
- Kaluarachchi, H.; Zhang, J.W.; Zamble, D.B. Escherichia coli SlyD, More Than a Ni(II) Reservoir. Biochemistry 2011, 50, 10761–10763. [Google Scholar] [CrossRef]
- Cheng, T.; Li, H.; Yang, X.; Xia, W.; Sun, H. Interaction of SlyD with HypB of Helicobacter pylori Facilitates Nickel Trafficking. Metallomics 2013, 5, 804–807. [Google Scholar] [CrossRef]
- Zhang, J.W.; Butland, G.; Greenblatt, J.F.; Emili, A.; Zamble, D.B. A Role for SlyD in the Escherichia coli Hydrogenase Biosynthetic Pathway. J. Biol. Chem. 2005, 280, 4360–4366. [Google Scholar] [CrossRef]
- Leach, M.R.; Jie, W.Z.; Zamble, D.B. The Role of Complex Formation between the Escherichia coli Hydrogenase Accessory Factors HypB and SlyD. J. Biol. Chem. 2007, 282, 16177–16186. [Google Scholar] [CrossRef]
- Lacasse, M.J.; Zamble, D.B. [NiFe]-Hydrogenase Maturation. Biochemistry 2016, 55, 1689–1701. [Google Scholar] [CrossRef]
- Zeer-wanklyn, C.J.; Zamble, D.B. Microbial Nickel: Cellular Uptake and Delivery to Enzyme Centers. Curr. Opin. Chem. Biol. 2017, 37, 80–88. [Google Scholar] [CrossRef]
- Witte, C.-P. Urea Metabolism in Plants. Plant Sci. 2011, 180, 431–438. [Google Scholar] [CrossRef]
- Polacco, J.C.; Mazzafera, P.; Tezotto, T. Opinion—Nickel and Urease in Plants: Still Many Knowledge Gaps. Plant Sci. 2013, 199–200, 79–90. [Google Scholar] [CrossRef]
- Cao, F.-Q.; Werner, A.K.; Dahncke, K.; Romeis, T.; Liu, L.-H.; Witte, C.-P. Identification and Characterization of Proteins Involved in Rice Urea and Arginine Catabolism. Plant Physiol. 2010, 154, 98–108. [Google Scholar] [CrossRef] [Green Version]
- Myrach, T.; Zhu, A.; Witte, C.-P. The Assembly of the Plant Urease Activation Complex and the Essential Role of the Urease Accessory Protein G (UreG) in Delivery of Nickel to Urease. J. Biol. Chem. 2017, 292, 14556–14565. [Google Scholar] [CrossRef]
- Witte, C.-P.; Rosso, M.G.; Romeis, T. Identification of Three Urease Accessory Proteins that are Required for Urease Activation in Arabidopsis. Plant Physiol. 2005, 139, 1155–1162. [Google Scholar] [CrossRef]
- Freyermuth, S.K.; Bacanamwo, M.; Polacco, J.C. The Soybean Eu3 Gene Encodes an Ni-Binding Protein Necessary for Urease Activity. Plant J. 2000, 21, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, J.C. The Emerging Role of Urease as a General Microbial Virulence Factor. PLoS Pathog. 2014, 10, e1004062. [Google Scholar] [CrossRef] [PubMed]
- Safavi, M.; Sabourian, R.; Foroumadi, A. Treatment of Helicobacter pylori Infection: Current and Future Insights. World J. Clin. Cases 2016, 4, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Koohi-moghadam, M.; Wang, R.; Chang, Y.; Woo, P.C.Y.; Wang, J.; Li, H.; Sun, H. Metallochaperone UreG Serves as a New Target for Design of Urease Inhibitor: A Novel Strategy for Development of Antimicrobials. PLoS Biol. 2018, 16, e2003887. [Google Scholar] [CrossRef] [PubMed]
- Eschweiler, J.D.; Farrugia, M.A.; Dixit, S.M.; Hausinger, R.P.; Ruotolo, B.T. A Structural Model of the Urease Activation Complex Derived from Ion Mobility-Mass Spectrometry and Integrative Modeling. Structure 2018, 26, 599–606.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ligabue-Braun, R.; Real-Guerra, R.; Carlini, C.R.; Verli, H. Evidence-Based Docking of the Urease Activation Complex. J. Biomol. Struct. Dyn. 2013, 31, 854–861. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structures of ureases. (A) Crystal structure of K. aerogenes urease (PDB: 1FWJ [16]). The basic catalytic unit of ureases consists of α, β and γ subunits encoding by ureC, ureB and ureA, respectively. In K. aerogenes and S. pasteurii, three copies of UreABC form a trimeric disk-like structure. The positions of three actives sites are circled in yellow and zoomed in at the right. The two nickel ions are coordinated by a carbamylated lysine residue, four histidine, one aspartate, and three water molecules. (B) The trimeric urease, schematically represented as triangles, forms the basic repeating unit of ureases to form more complex quaternary structures. In plant ureases, such as Canavalia ensiformis (PDB: 3LA4 [19]), two urease trimers are stacked together in a C2 symmetry to form a hexameric quaternary structure. In H. pylori, four trimers assemble in a tetrahedral symmetry to form a dodecameric urease (PDB: 1E9Z [20]).
Figure 1.
Structures of ureases. (A) Crystal structure of K. aerogenes urease (PDB: 1FWJ [16]). The basic catalytic unit of ureases consists of α, β and γ subunits encoding by ureC, ureB and ureA, respectively. In K. aerogenes and S. pasteurii, three copies of UreABC form a trimeric disk-like structure. The positions of three actives sites are circled in yellow and zoomed in at the right. The two nickel ions are coordinated by a carbamylated lysine residue, four histidine, one aspartate, and three water molecules. (B) The trimeric urease, schematically represented as triangles, forms the basic repeating unit of ureases to form more complex quaternary structures. In plant ureases, such as Canavalia ensiformis (PDB: 3LA4 [19]), two urease trimers are stacked together in a C2 symmetry to form a hexameric quaternary structure. In H. pylori, four trimers assemble in a tetrahedral symmetry to form a dodecameric urease (PDB: 1E9Z [20]).
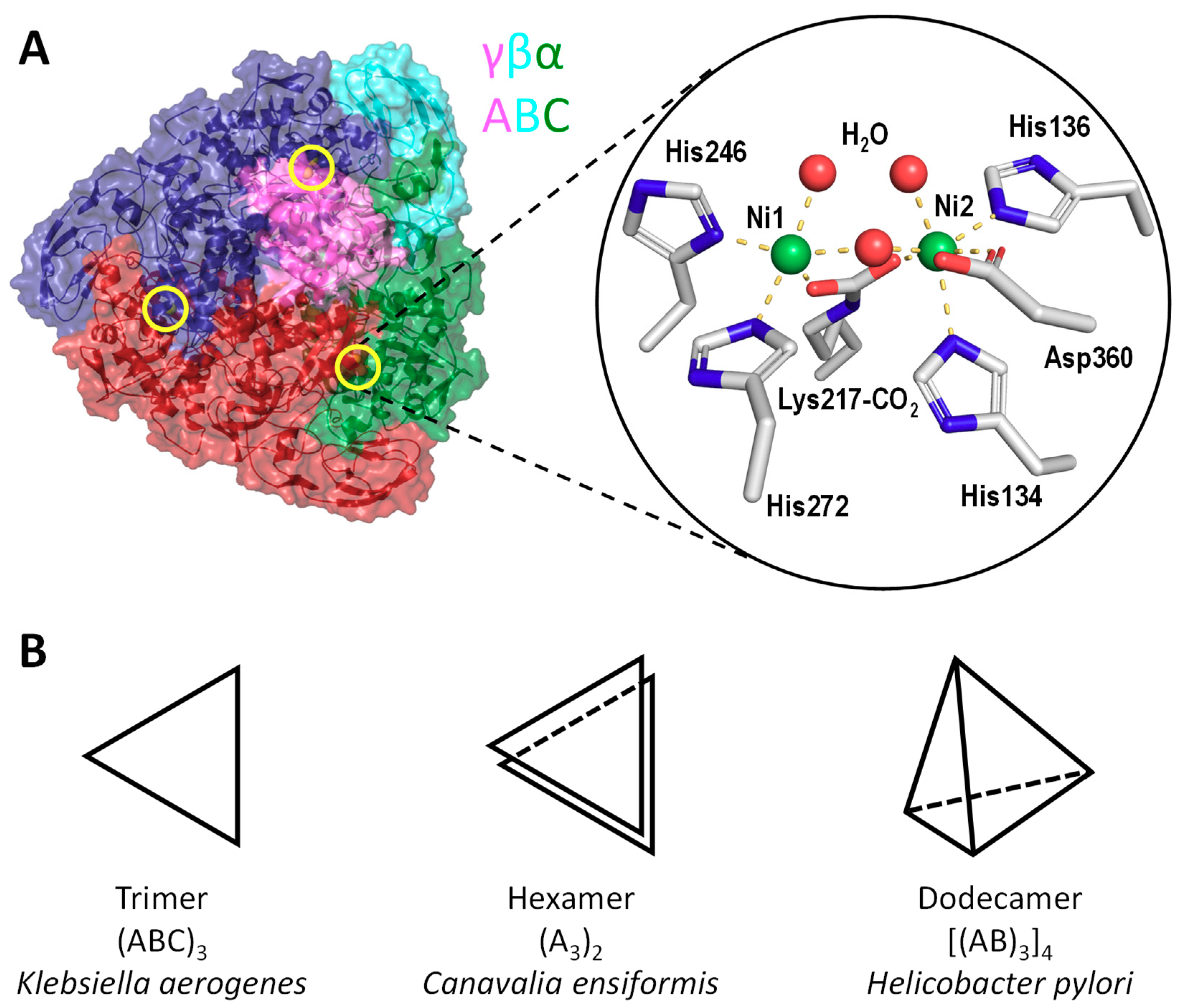
Figure 2.
Comparison of K. aerogenes and H. pylori urease operons. Orthologous genes are indicated using the same color. The locations of the active-site lysine residue involved in carbamylation and nickel binding are indicated as stars. H. pylori contains an extra ureI gene encoding for an acid-gated urea channel.
Figure 2.
Comparison of K. aerogenes and H. pylori urease operons. Orthologous genes are indicated using the same color. The locations of the active-site lysine residue involved in carbamylation and nickel binding are indicated as stars. H. pylori contains an extra ureI gene encoding for an acid-gated urea channel.

Figure 3.
Dimerization of UreF creates a saddle-like structure for recruiting UreG to the UreGFD complex. (A) Crystal structure of the UreFD complex (PDB: 3SF5 [48]). UreF (grey) interacts with UreD (yellow) to form a 2:2 heterodimer in a rod-shape topological arrangement of D:F:F:D. A saddle-like structure is formed at the UreF dimer surface. Invariant residues of UreF that have been shown to be essential to UreG binding and urease activation are highlighted in purple and are numbered according to the H. pylori sequence [48,50]. (B) Crystal structure of the UreGFD complex showing that UreG dimer (cyan) binds to the saddle-like structure of UreF (PDB: 4HI0 [43]).
Figure 3.
Dimerization of UreF creates a saddle-like structure for recruiting UreG to the UreGFD complex. (A) Crystal structure of the UreFD complex (PDB: 3SF5 [48]). UreF (grey) interacts with UreD (yellow) to form a 2:2 heterodimer in a rod-shape topological arrangement of D:F:F:D. A saddle-like structure is formed at the UreF dimer surface. Invariant residues of UreF that have been shown to be essential to UreG binding and urease activation are highlighted in purple and are numbered according to the H. pylori sequence [48,50]. (B) Crystal structure of the UreGFD complex showing that UreG dimer (cyan) binds to the saddle-like structure of UreF (PDB: 4HI0 [43]).
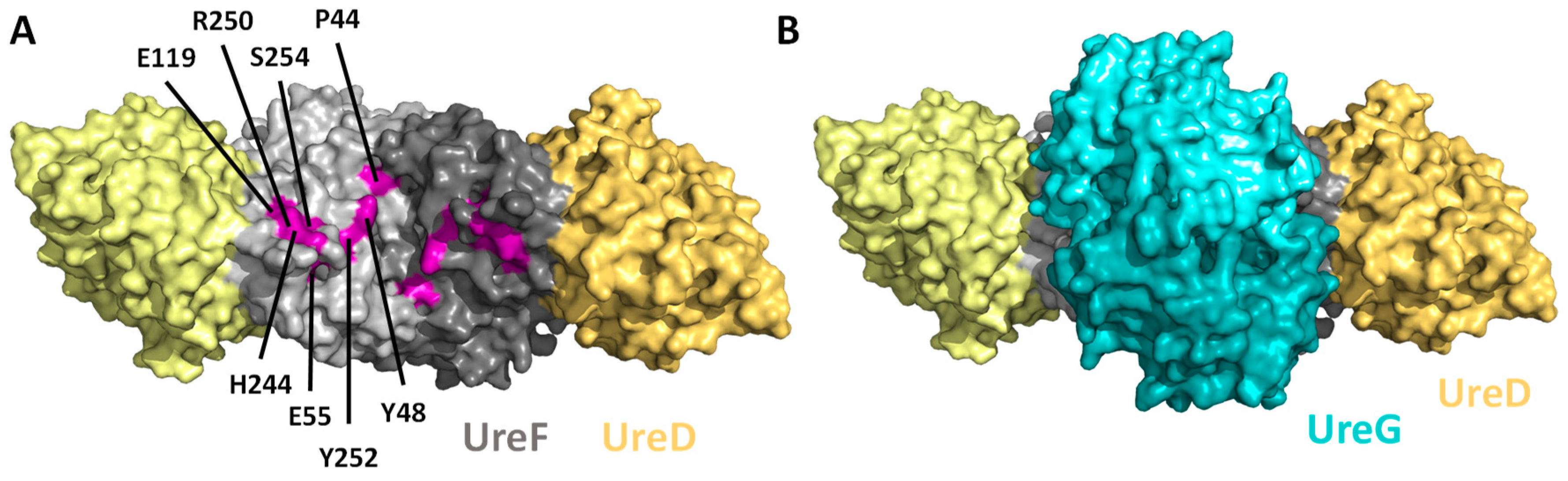
Figure 4.
Guanosine triphosphate (GTP)-dependent conformational changes of UreG. (A) The structures of GDP-bound H. pylori UreG (PDB: 4HI0 [43]) is compared to the (B) Ni/GMPPNP-bound K. pneumoniae UreG (PDB: 5XKT [51]). Invariant residues involved in the conformational changes and nickel binding are indicated and numbered according to the H. pylori sequence. Residues in the opposite chain are numbered with apostrophes. Noteworthy, Cys66 and His68 of the CPH motif (magenta) are pointing away from each other in the GDP-bound UreG. Upon binding of GMPPNP, they reorient to form a square-planar nickel binding site at the dimeric interface. Moreover, the swinging motion of Tyr39 creates steric clashes that induce dissociation of UreG from the UreGFD complex.
Figure 4.
Guanosine triphosphate (GTP)-dependent conformational changes of UreG. (A) The structures of GDP-bound H. pylori UreG (PDB: 4HI0 [43]) is compared to the (B) Ni/GMPPNP-bound K. pneumoniae UreG (PDB: 5XKT [51]). Invariant residues involved in the conformational changes and nickel binding are indicated and numbered according to the H. pylori sequence. Residues in the opposite chain are numbered with apostrophes. Noteworthy, Cys66 and His68 of the CPH motif (magenta) are pointing away from each other in the GDP-bound UreG. Upon binding of GMPPNP, they reorient to form a square-planar nickel binding site at the dimeric interface. Moreover, the swinging motion of Tyr39 creates steric clashes that induce dissociation of UreG from the UreGFD complex.
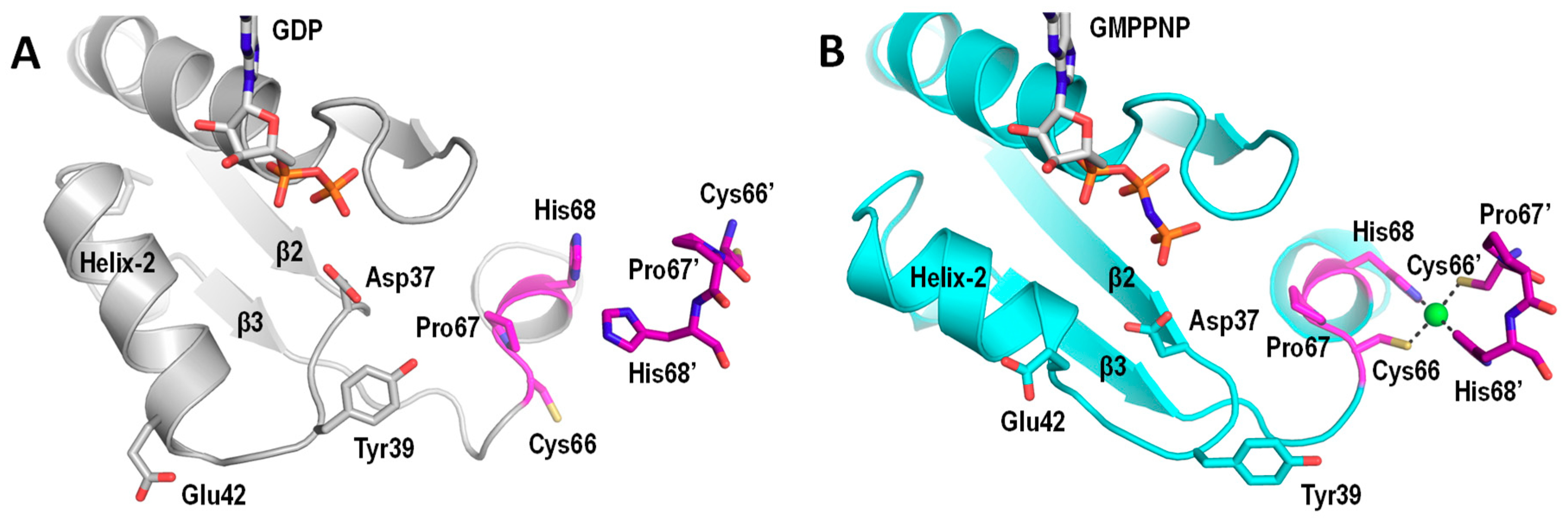
Figure 5.
Structures of UreE and UreG. Cartoon representation of K. pneumoniae Ni/GMPPNP-bound UreG (A, PDB: 5XKT [51]) and H. pylori Ni-bound UreE (B, PDB: 3TJ8 [67]). Both UreG and UreE bind a nickel ion at the dimeric interface. The surface electrostatic potentials of (C) UreG and (D) UreE were calculated using the APBS program [81] and are color coded (red, −5 kT/e; blue, +5 kT/e). (E) It has been suggested that UreG and UreE are complementary in charge and shape and are likely to form a UreE2G2 complex with their nickel binding sites pointing towards each other [71,80].
Figure 5.
Structures of UreE and UreG. Cartoon representation of K. pneumoniae Ni/GMPPNP-bound UreG (A, PDB: 5XKT [51]) and H. pylori Ni-bound UreE (B, PDB: 3TJ8 [67]). Both UreG and UreE bind a nickel ion at the dimeric interface. The surface electrostatic potentials of (C) UreG and (D) UreE were calculated using the APBS program [81] and are color coded (red, −5 kT/e; blue, +5 kT/e). (E) It has been suggested that UreG and UreE are complementary in charge and shape and are likely to form a UreE2G2 complex with their nickel binding sites pointing towards each other [71,80].

Figure 6.
The urease maturation pathway.

Figure 7.
Dependency of GTPase activity of UreG on bicarbonate concentration. GTPase activity of H. pylori UreG (5 µM) was measured at 37 °C in 0–200 mM NaHCO3 at saturated concentration of GTP (2 mM) in 200 mM KCl, 2.5 µM NiSO4, 2 mM MgSO4, 1 mM TCEP, 20 mM HEPES pH 7.5 by following the concentration of phosphate released as described previously [87]. Fitting the data to the Michaelis–Mention equation yielded a Km value of 13 ± 4 mM and a kcat value of 6.4 ± 0.6 × 10−3 s−1.
Figure 7.
Dependency of GTPase activity of UreG on bicarbonate concentration. GTPase activity of H. pylori UreG (5 µM) was measured at 37 °C in 0–200 mM NaHCO3 at saturated concentration of GTP (2 mM) in 200 mM KCl, 2.5 µM NiSO4, 2 mM MgSO4, 1 mM TCEP, 20 mM HEPES pH 7.5 by following the concentration of phosphate released as described previously [87]. Fitting the data to the Michaelis–Mention equation yielded a Km value of 13 ± 4 mM and a kcat value of 6.4 ± 0.6 × 10−3 s−1.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nim, Y.S.; Wong, K.-B. The Maturation Pathway of Nickel Urease. Inorganics 2019, 7, 85. https://0-doi-org.brum.beds.ac.uk/10.3390/inorganics7070085
AMA Style
Nim YS, Wong K-B. The Maturation Pathway of Nickel Urease. Inorganics. 2019; 7(7):85. https://0-doi-org.brum.beds.ac.uk/10.3390/inorganics7070085
Chicago/Turabian StyleNim, Yap Shing, and Kam-Bo Wong. 2019. "The Maturation Pathway of Nickel Urease" Inorganics 7, no. 7: 85. https://0-doi-org.brum.beds.ac.uk/10.3390/inorganics7070085
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.