
Broadening the Scope of Polyoxometalates as Artificial Proteases in Surfactant Solutions: Hydrolysis of Ovalbumin by Zr(IV)-Substituted Keggin Complex


Abstract
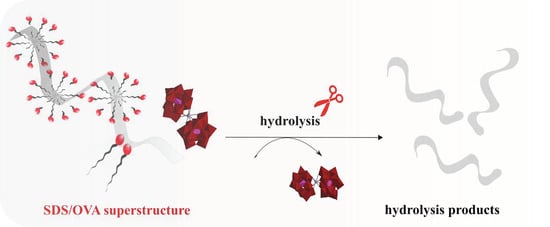
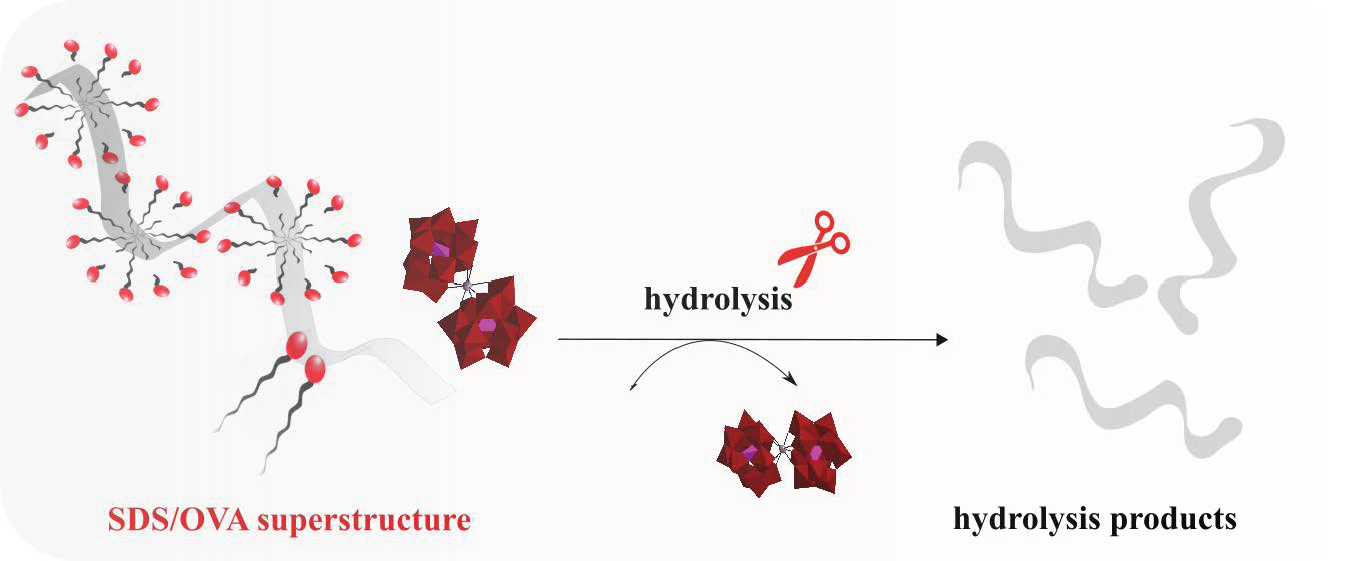
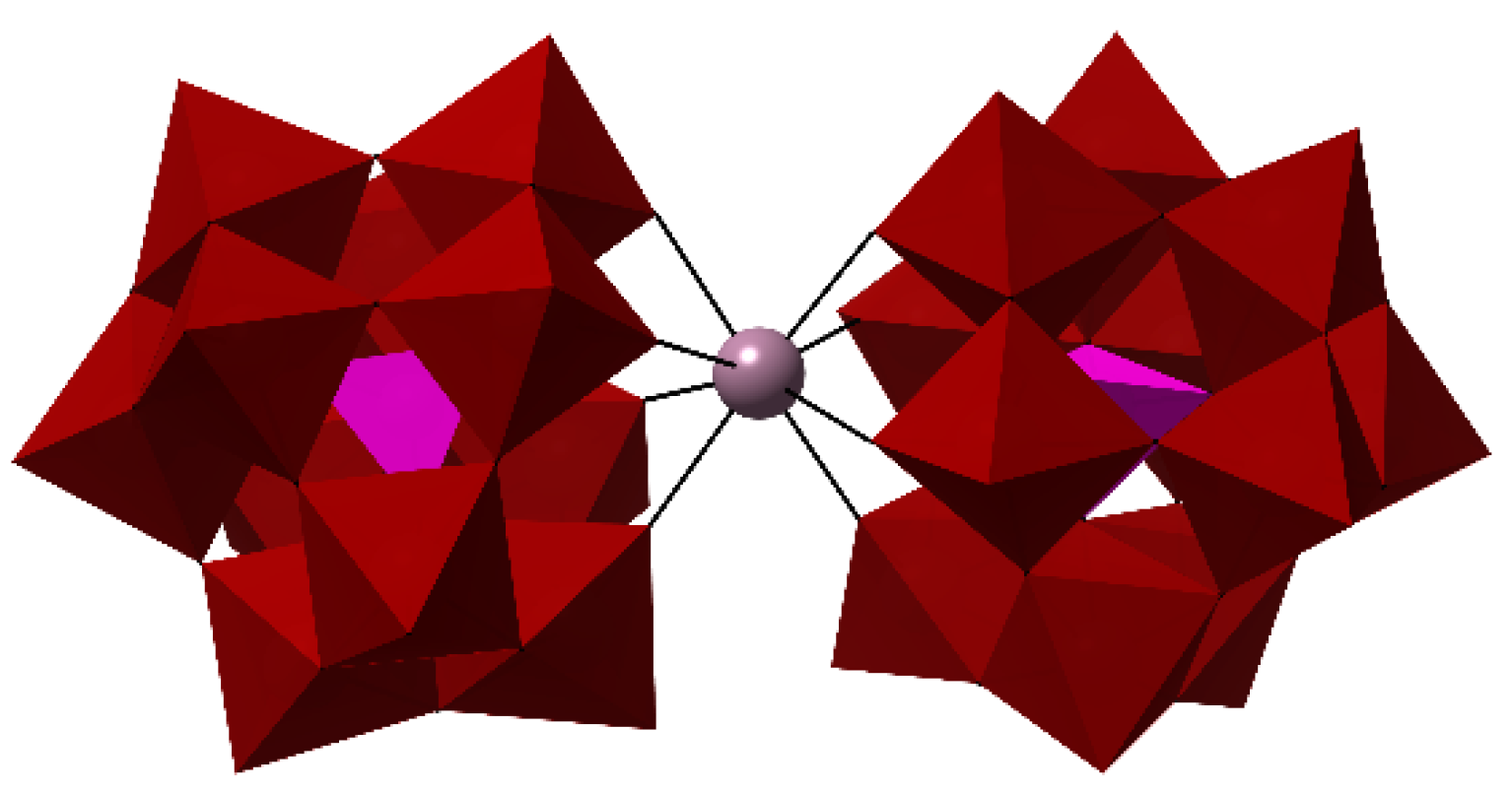
:
1. Introduction
2. Results and Discussion
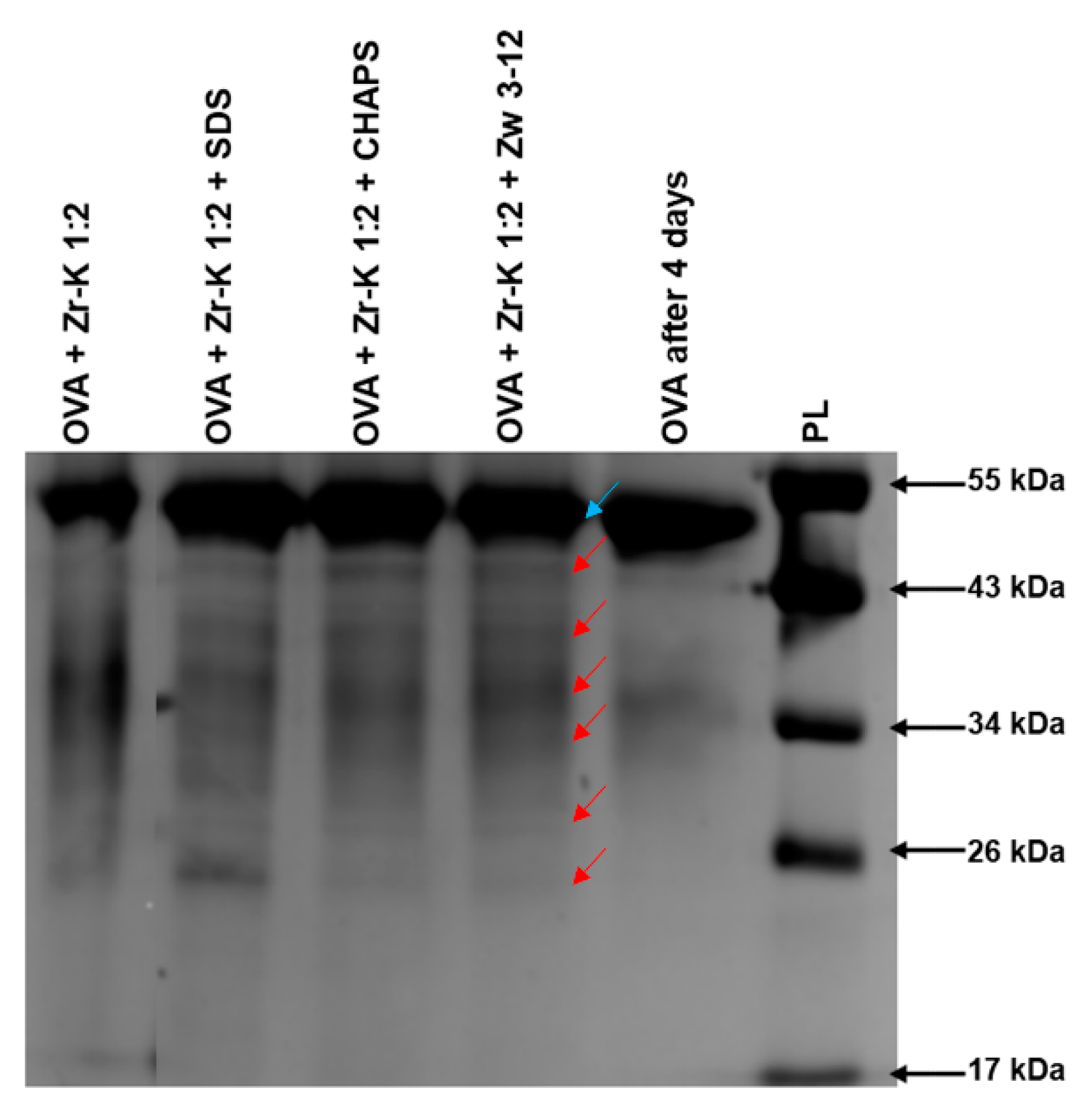
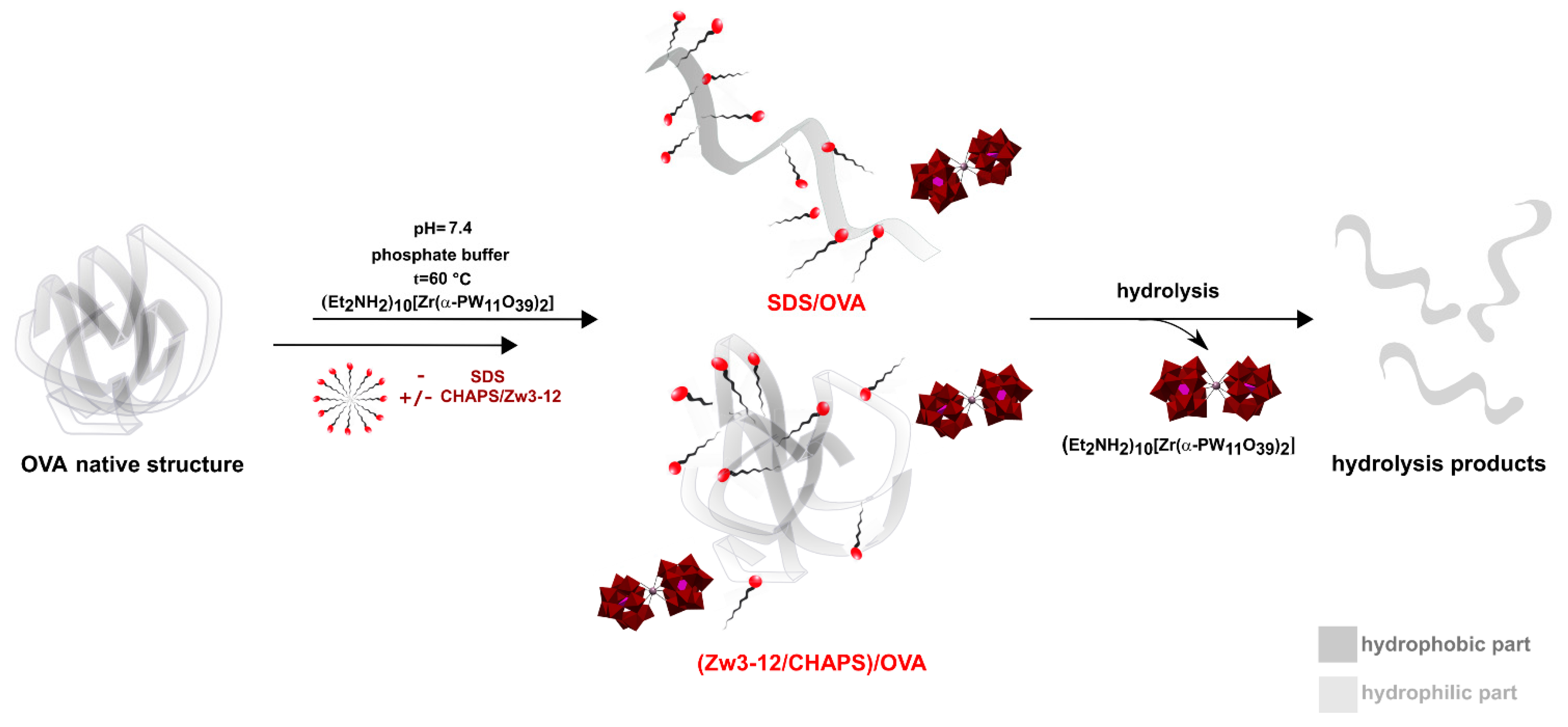
2.1. Hydrolysis of OVA by Zr-K 1:2 in the Absence or Presence of Surfactants
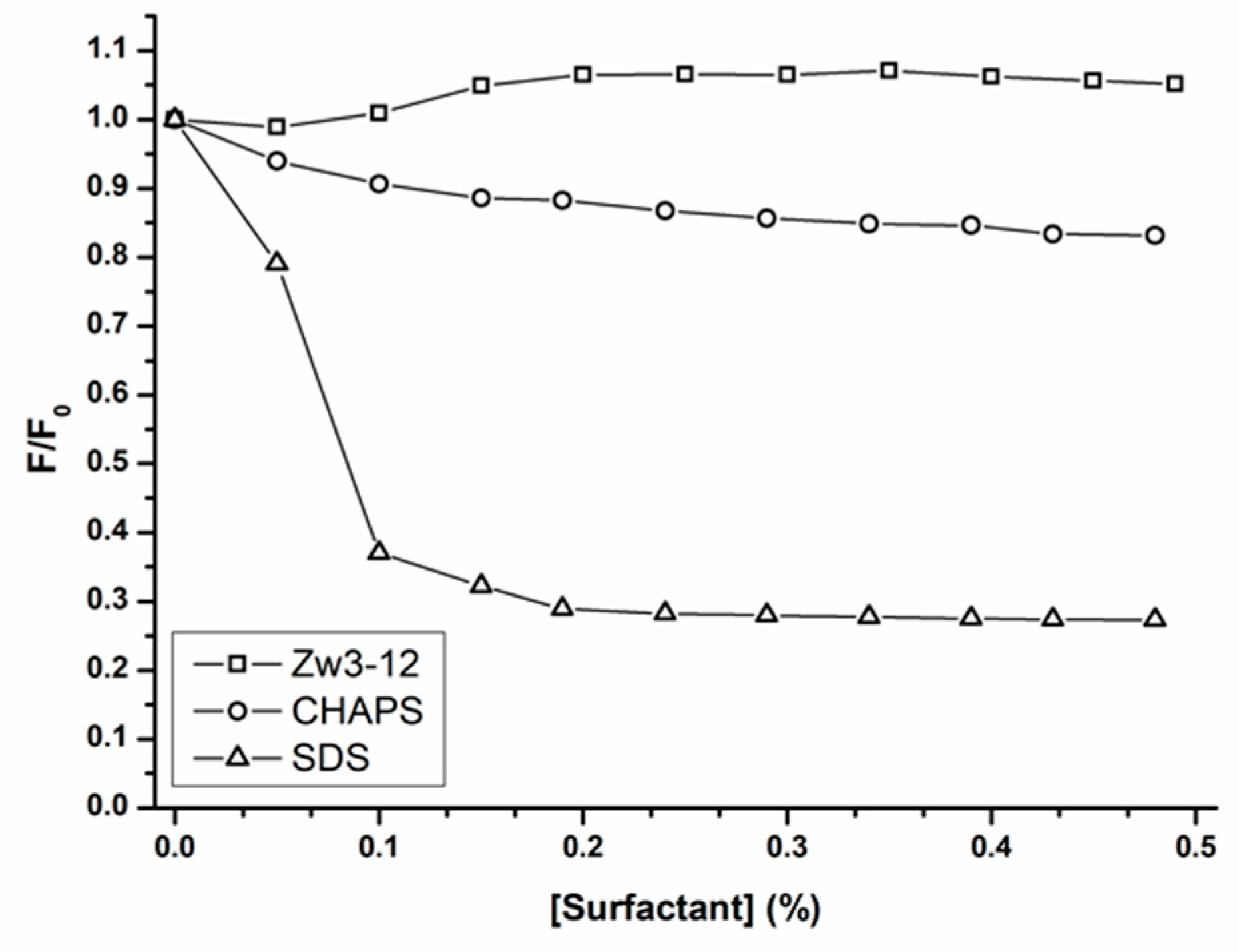
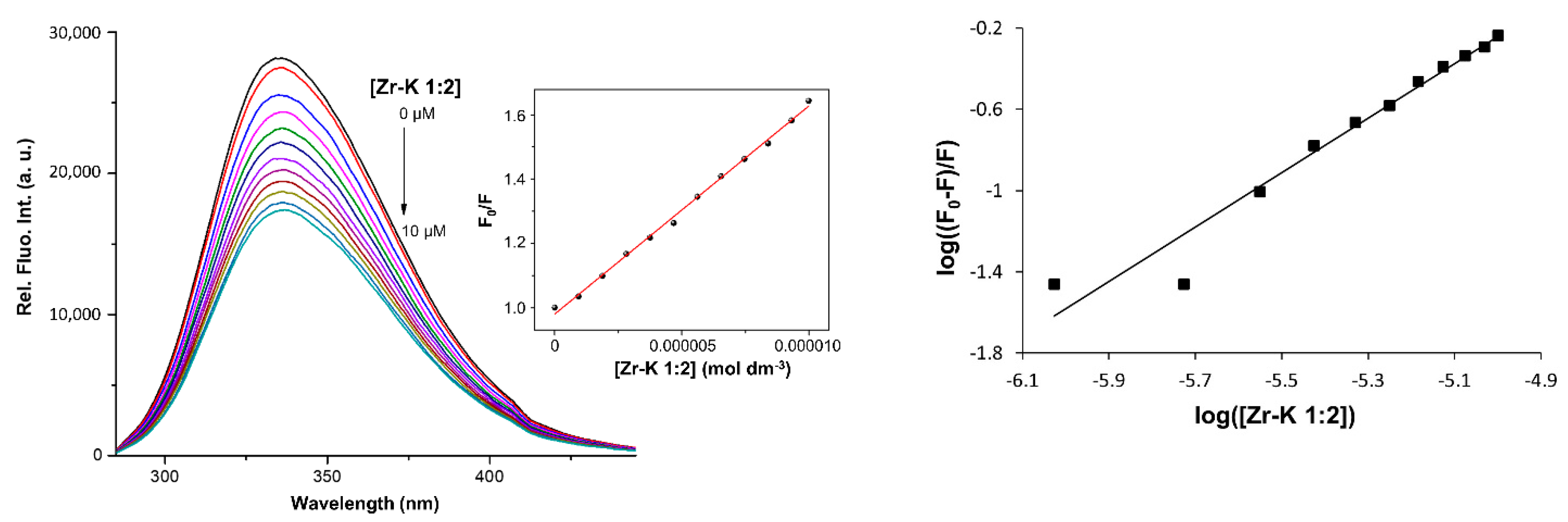
2.2. Interaction between OVA and Zr-K 1:2 in the Absence or Presence of Surfactants
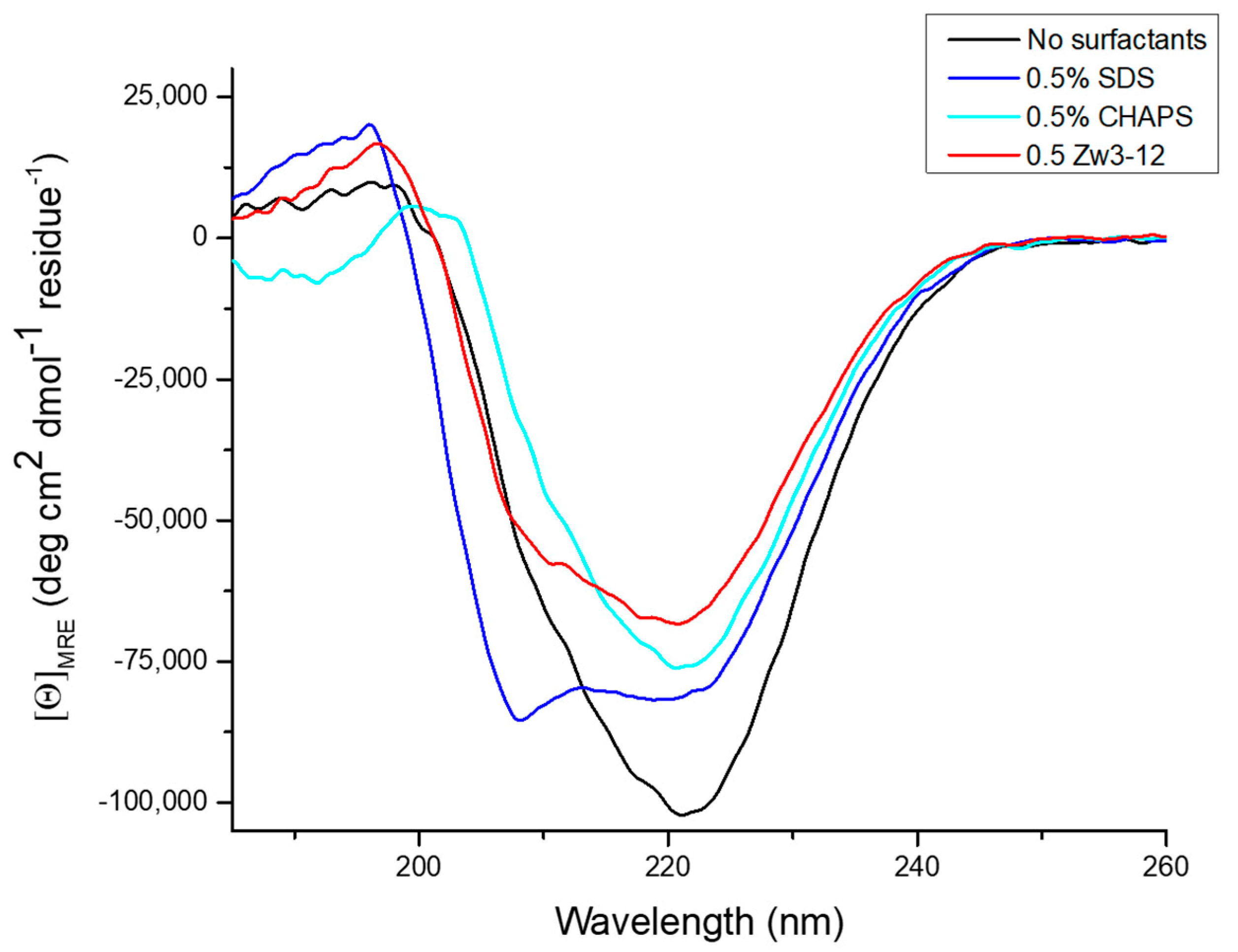
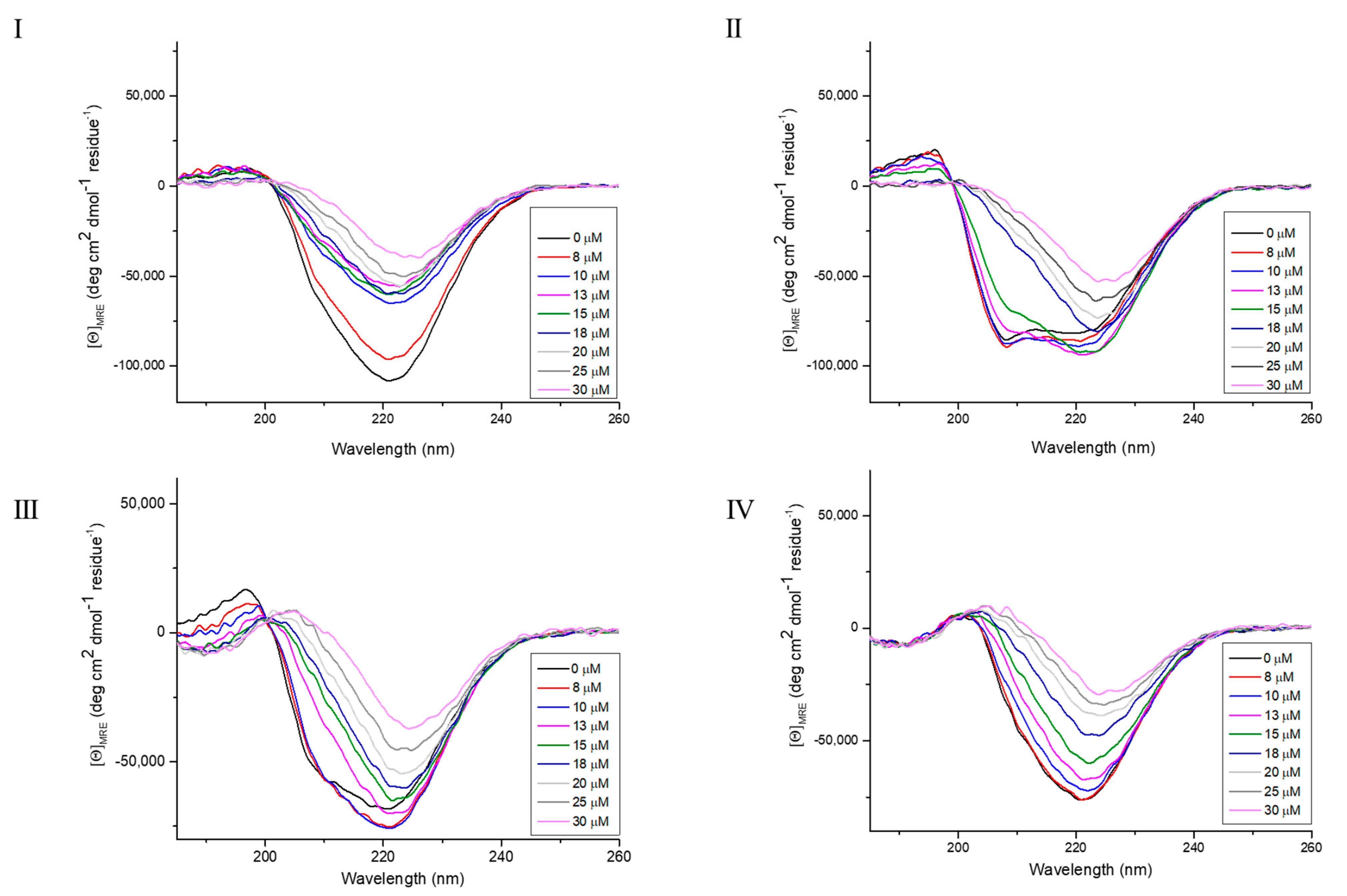
2.3. Effect of Zr-K 1:2 and Surfactants on the Secondary Structure of OVA
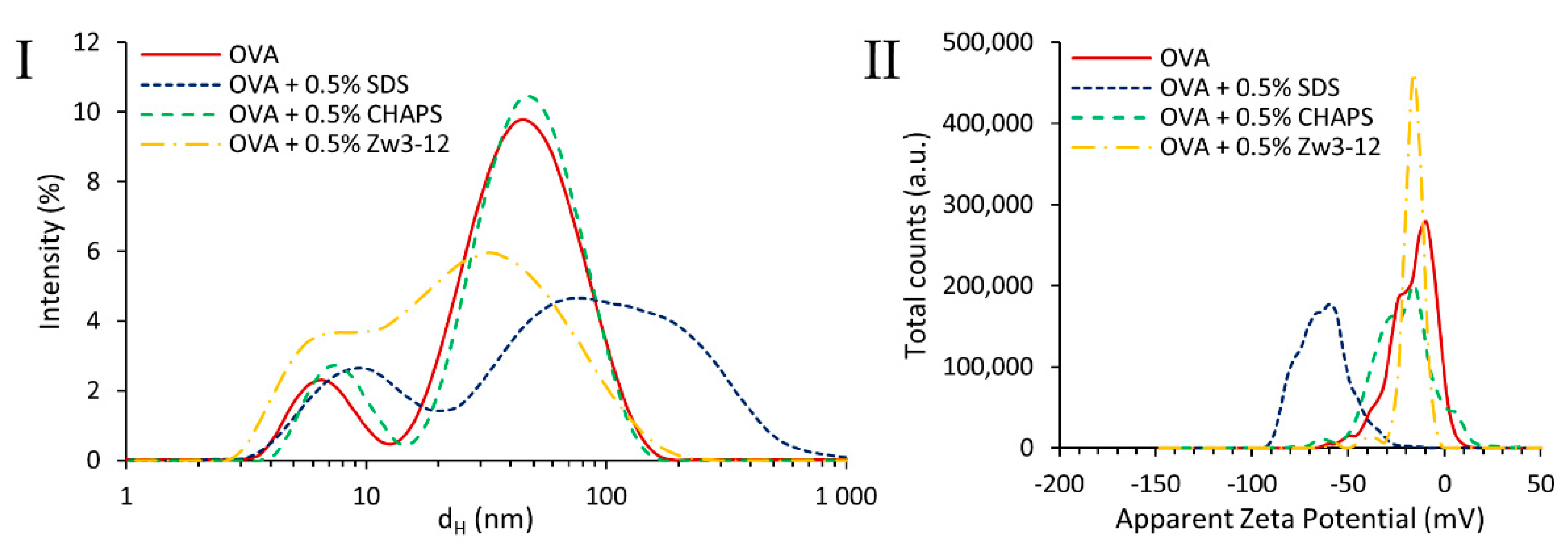
2.4. Particle Size Distribution and Zeta Potential
2.5. 31P NMR Stability Study
3. Materials and Methods
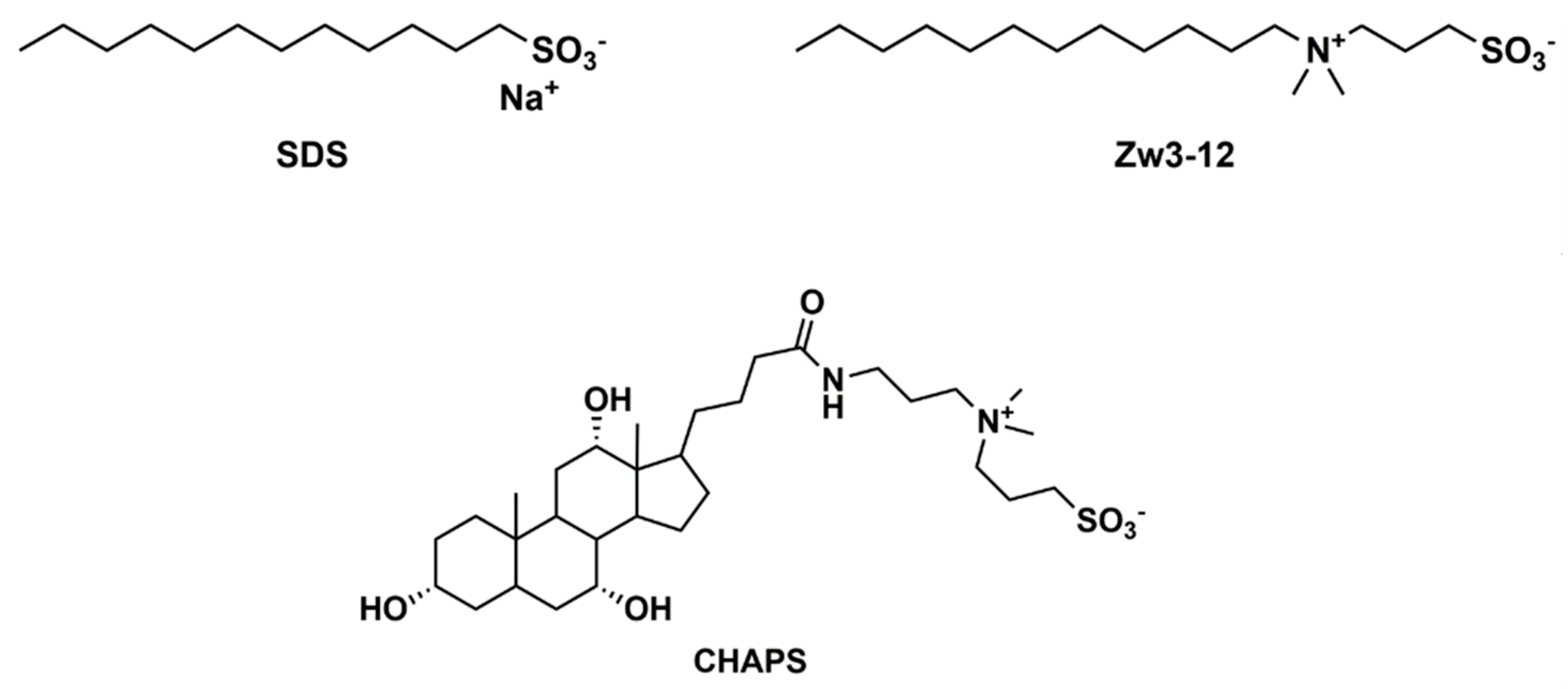
3.1. Materials
3.2. Methods
3.2.1. Hydrolysis Experiments
3.2.2. Fluorescence Spectroscopy
3.2.3. Circular Dichroism Spectroscopy
3.2.4. Particle Size Distribution and Zeta Potential
3.2.5. 31P NMR Spectroscopy
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scott, K.A.; Bond, P.J.; Ivetac, A.; Chetwynd, A.P.; Khalid, S.; Sansom, M.S.P. Coarse-Grained MD simulations of membrane protein-bilayer self-assembly. Structure 2008, 16, 621–630. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; Knowles, T.J.; Postis, V.L.G.; Jamshad, M.; Parslow, R.A.; Lin, Y.-P.; Goldman, A.; Sridhar, P.; Overduin, M.; Muench, S.P.; et al. A method for detergent-free isolation of membrane proteins in their local lipid environment. Nat. Protoc. 2016, 11, 1149–1162. [Google Scholar] [CrossRef]
- Shah, T.R.; Misra, A. Challenges in Delivery of Therapeutic Genomics and Proteomics; Elsevier: London, UK, 2011; pp. 387–427. [Google Scholar]
- Guzman, M.L.; Marques, M.R.; Me, M.E.O.; Stippler, E.S. Enzymatic activity in the presence of surfactants commonly used in dissolution media, Part 1: Pepsin. Results Pharma Sci. 2016, 6, 15–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ly, H.G.T.; Absilis, G.; Janssens, R.; Proost, P.; Parac-Vogt, T.N. Highly amino acid selective hydrolysis of myoglobin at aspartate residues as promoted by Zirconium(IV)-substituted polyoxometalates. Angew. Chem. Int. Ed. 2015, 54, 7391–7394. [Google Scholar] [CrossRef] [PubMed]
- Stroobants, K.; Absillis, G.; Moelants, E.; Proost, P.; Parac-Vogt, T.N. Regioselective hydrolysis of human serum albumin by ZrIV-substituted polyoxotungstates at the interface of positively charged protein surface patches and negatively charged amino acid residues. Chem. Eur. J. 2014, 20, 3894–3897. [Google Scholar] [CrossRef]
- Sap, A.; Absillis, G.; Parac-Vogt, T.N. Selective hydrolysis of oxidized insulin chain B by a Zr(IV)-substituted Wells-Dawson polyoxometalate. Dalton Trans. 2015, 44, 1539–1548. [Google Scholar] [CrossRef] [Green Version]
- Sap, A.; van Tichelen, L.; Mortier, A.; Proost, P.; Parac-Vogt, T.N. Tuning the selectivity and reactivity of metal-substituted polyoxometalates as artificial proteases by varying the nature of the embedded Lewis acid metal ion. Eur. J. Inorg. Chem. 2016, 32, 5098–5105. [Google Scholar] [CrossRef]
- Anyushin, A.V.; Sap, A.; Quanten, T.; Proost, P.; Parac-Vogt, T.N. Selective hydrolysis of Ovalbumin promoted by Hf(IV)-substituted Wells-Dawson-type polyoxometalate. Front. Chem. 2018, 6, 614. [Google Scholar] [CrossRef]
- Stroobants, K.; Moelants, E.; Ly, H.G.T.; Proost, P.; Bartik, K.; Parac-Vogt, T.N. Polyoxometalates as a novel class of artificial proteases: Selective hydrolysis of Lysozyme under physiological pH and temperature promoted by a Cerium(IV) Keggin-type polyoxometalate. Chem. Eur. J. 2013, 19, 2848–2858. [Google Scholar] [CrossRef] [PubMed]
- Ly, H.G.T.; Fu, G.; Kondinski, A.; Bueken, B.; de Vos, D.; Parac-Vogt, T.N. Superactivity of MOF-808 toward peptide bond hydrolysis. J. Am. Chem. Soc. 2018, 140, 6325–6335. [Google Scholar] [CrossRef]
- Moons, J.; de Azambuja, F.; Mihailovic, J.; Kozma, K.; Smiljanic, K.; Amiri, M.; Velickovic, T.C.; Nyman, M.; Parac-Vogt, T.N. Discrete Hf18 metal-oxo cluster as a heterogeneous nanozyme for site-specific proteolysis. Angew. Chem. Int. Ed. 2020, 59, 9094–9101. [Google Scholar] [CrossRef]
- Loosen, A.; de Azambuja, F.; Smolders, S.; Moons, J.; Simms, C.; de Vos, D.; Parac-Vogt, T.N. Interplay between structural parameters and reactivity of Zr6-based MOFs as artificial proteases. Chem. Sci. 2020, 11, 6662–6669. [Google Scholar] [CrossRef]
- Ly, H.G.T.; Fu, G.; de Azambuja, F.; de Vos, D.; Parac-Vogt, T.N. Nanozymatic Activity of UiO-66 metal-organic frameworks: Tuning the nanopore environment enhances hydrolytic activity toward peptide bonds. ACS Appl. Nano Mater. 2020, 3, 8931–8938. [Google Scholar] [CrossRef]
- Nisar, A.; Wang, X. Surfactant-encapsulated polyoxometalate building blocks: Controlled assembly and their catalytic properties. Dalton Trans. 2012, 41, 9832–9845. [Google Scholar] [CrossRef]
- Pope, M.T.; Müller, A. Polyoxometalate chemistry: An old field with new dimensions in several disciplines. Angew. Chem. Int. Ed. 1991, 30, 34–48. [Google Scholar] [CrossRef]
- Proust, A.; Matt, B.; Villanneau, R.; Guillemot, G.; Gouzerh, P.; Izzet, G. Functionalization and post-functionalization: A step towards polyoxometalate-based materials. Chem. Soc. Rev. 2012, 41, 7605–7622. [Google Scholar] [CrossRef] [PubMed]
- Quanten, T.; Shestakova, P.; van Bulck, D.D.; Kirschhock, C.; Parac-Vogt, T.N. Interaction study and reactivity of ZrIV-substituted Wells-Dawson polyoxometalate towards hydrolysis of peptide bonds in surfactant solutions. Chem. Eur. J. 2016, 22, 3775–3784. [Google Scholar] [CrossRef] [PubMed]
- Sap, A.; Vandebroek, L.; Goovaerts, V.; Martens, E.; Proost, P.; Parac-Vogt, T.N. Highly selective and tunable protein hydrolysis by a polyoxometalate complex in surfactant Solutions: A step toward the development of artificial metalloproteases for membrane proteins. ACS Omega 2017, 2, 2026–2033. [Google Scholar] [CrossRef] [Green Version]
- Quanten, T.; de Mayaer, T.; Shestakova, P.; Parac-Vogt, T.N. Selectivity and reactivity of ZrIV and CeIV substituted Keggin type polyoxometalates toward Cytochrome c in surfactant solutions. Front. Chem. 2018, 6, 372. [Google Scholar] [CrossRef] [PubMed]
- Quanten, T.; Savić, N.D.; Parac-Vogt, T.N. Hydrolysis of peptide bonds in protein micelles promoted by a Zirconium(IV)-substituted polyoxometalate as an artificial protease. Chem. Eur. J. 2020, 26, 11170–11179. [Google Scholar] [CrossRef] [PubMed]
- Tofani, L.; Feis, A.; Snoke, R.E.; Berti, D.; Baglioni, P.; Smulevich, G. Spectroscopic and interfacial properties of myoglobin/surfactant complexes. Biophys. J. 2004, 87, 1186–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, S.; Das, S.; Ghosh, S. Interaction of myoglobin with cationic Gemini surfactants in phosphate buffer at pH 7.4. J. Surfact. Deterg. 2015, 18, 471–476. [Google Scholar] [CrossRef]
- Abeyrathne, E.D.N.S.; Lee, H.Y.; Jo, C.; Nam, K.C.; Ahn, D.U. Enzymatic hydrolysis of ovalbumin and the functional properties of the hydrolysates. Poult. Sci. 2014, 93, 2678–2686. [Google Scholar] [CrossRef]
- Huntington, J.A.; Stein, P.E. Structure and Properties of Ovalbumin. J. Chromatogr. B Biomed. Sci. Appl. 2001, 25, 189–198. [Google Scholar] [CrossRef]
- Gettins, P.G.W. Serpin structure, mechanism, and function. Chem. Rev. 2002, 102, 4751–4804. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Cai, J.; Guo, X. Effect of surfactant structure on reverse micellar extraction of Ovalbumin. Process Biochem. 2015, 50, 272–278. [Google Scholar] [CrossRef]
- Ting, B.P.C.P.; Pouliot, Y.; Gauthier, S.F.; Mine, Y. Separation, fractionation of egg proteins and peptides for nutraceutical applications. In Extraction and Concentration Processes in the Food Beverage and Nutraceutical Industries; Woodhead Publishing: Cambridge, UK, 2013; pp. 595–618. [Google Scholar]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Van Rompuy, L.; Savić, N.D.; Rodriguez, A.; Parac-Vogt, T.N. Selective hydrolysis of transferrin promoted by Zr-substituted polyoxometalates. Molecules 2020, 25, 3472. [Google Scholar] [CrossRef]
- Hellmann, N.; Schneider, D. Hands on: Using tryptophan fluorescence spectroscopy to study protein structure. In Protein Supersecondary Structures, 2nd ed.; Kister, A.E., Ed.; Springer Science: New York, NY, USA, 2019; pp. 379–401. [Google Scholar]
- Favicchio, R.; Dragan, A.I.; Kneale, G.G.; Read, C.M. Fluorescence spectroscopy and anisotropy in the analysis of DNA-protein interactions. In DNA-Protein Interactions, 3rd ed.; Leblanc, B., Moss, T., Eds.; Springer: New York, NY, USA, 2009; pp. 589–611. [Google Scholar]
- Ly, H.G.T.; Parac-Vogt, T.N. Spectroscopic study of the interaction between horse heart myoglobin and Zirconium(IV)-substituted polyoxometalates as artificial proteases. Chemphyschem 2017, 18, 2451–2458. [Google Scholar] [CrossRef]
- Goovaerts, V.; Stroobants, K.; Absillis, G.; Parac-Vogt, T.N. Molecular interactions between serum albumin proteins and Keggin type polyoxometalates studied using luminescence spectroscopy. Phys. Chem. Chem. Phys. 2013, 15, 18378–18387. [Google Scholar] [CrossRef]
- Stroobants, K.; Goovaerts, V.; Absillis, G.; Bruylants, G.; Moelants, E.; Proost, P.; Parac-Vogt, T.N. Molecular origin of the hydrolytic activity and fixed regioselectivity of a ZrIV-substituted polyoxotungstate as artificial protease. Chem. Eur. J. 2014, 20, 9567–9577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goovaerts, V.; Stroobants, K.; Absillis, G.; Parac-Vogt, T.N. Understanding the regioselective hydrolysis of human serum albumin by Zr(IV)-substituted polyoxotungstates using tryptophan fluorescence spectroscopy. Inorganics 2015, 3, 230–245. [Google Scholar] [CrossRef] [Green Version]
- Vivian, J.T.; Callis, P.R. Mechanisms of tryptophan fluorescence shifts in proteins. Biophys. J. 2001, 80, 2093–2109. [Google Scholar] [CrossRef] [Green Version]
- Vallée-Bélisle, A.; Michnick, S.W. Visualizing transient protein-folding intermediates by tryptophan-scanning mutagenesis. Nat. Struct. Mol. Biol. 2012, 19, 731–736. [Google Scholar] [CrossRef]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 3rd ed.; Springer: New York, NY, USA, 2006. [Google Scholar]
- Kelly, S.; Price, N. The use of circular dichroism in the investigation of protein structure and function. Curr. Protein Pept. Sci. 2000, 1, 349–384. [Google Scholar] [CrossRef] [Green Version]
- Sreerama, N.; Woody, R.W. Estimation of protein secondary structure from circular dichroism spectra: Comparison of CONTIN, SELCON, and CDSSTR methods with an expanded reference set. Anal. Biochem. 2000, 287, 252–260. [Google Scholar] [CrossRef]
- Whitmore, L.; Wallace, B.A. DICHROWEB, an online server for protein secondary structure analyses from circular dichroism spectroscopic data. Nucleic Acids Res. 2004, 32, W668–W673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitmore, L.; Wallace, B.A. Protein secondary structure analyses from circular dichroism spectroscopy: Methods and reference databases. Biopolymers 2008, 89, 392–400. [Google Scholar] [CrossRef]
- Zemser, M.; Friedman, M.; Katzhendler, J.; Greene, L.L.; Minsky, A.; Gorinstein, S. Relationship between functional properties and structure of Ovalbumin. J. Protein Chem. 1994, 13, 261–274. [Google Scholar] [CrossRef]
- Dong, A.; Meyer, J.D.; Brown, J.L.; Manning, M.C.; Carpenter, J.F. Comparative fourier transform infrared and circular dichroism spectroscopic analysis of α1-Proteinase inhibitor and Ovalbumin in aqueous solution. Arch. Biochem. Biophys. 2000, 383, 148–155. [Google Scholar] [CrossRef]
- Bruch, M.; Weiss, V.; Engel, J. Plasma serine proteinase inhibitors (serpins) exhibit major conformational changes and a large increase in conformational stability upon cleavage at their reactive sites. J. Biol. Chem. 1988, 263, 16626–16630. [Google Scholar] [CrossRef]
- González-Pérez, A.; Ruso, J.M.; Prieto, G.; Sarmiento, F. Physicochemical study of Ovalbumin in the presence of sodium dodecyl sulphate in aqueous media. Colloid Polym. Sci. 2004, 282, 351–356. [Google Scholar] [CrossRef]
- Meyer, M.L.; Kauzmann, W. The effects of detergents and urea on the rotatory dispersion of Ovalbumin. Arch. Biochem. Biophys. 1962, 99, 348–349. [Google Scholar] [CrossRef]
- Mattice, W.L.; Riser, J.M.; Clark, D.S. Conformational properties of the complexes formed by proteins and sodium dodecyl sulfate. Biochemistry 1976, 15, 4264–4272. [Google Scholar] [CrossRef]
- Visentini, F.F.; Perez, A.A.; Santiago, L.G. Self-assembled nanoparticles from heat treated ovalbumin as nanocarriers for polyunsaturated fatty acids. Food Hydrocoll. 2019, 93, 242–252. [Google Scholar] [CrossRef]
- Sap, A.; de Zitter, E.; van Meervelt, L.; Parac-Vogt, T.N. Structural characterization of the complex between hen Egg-White Lysozyme and ZrIV-substituted Keggin polyoxometalate as artificial protease. Chem. Eur. J. 2015, 21, 11692–11695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokolov, M.N.; Chubarova, E.V.; Peresypkina, E.V.; Virovets, A.V.; Fedin, V.P. Complexes of ZrIV and HfIV with monolacunary Keggin-and Dawson-type anions. Russ. Chem. Bull. 2007, 56, 220–224. [Google Scholar] [CrossRef]
- Wallace, B.A.; Janes, R.W. Modern Techniques for Circular Dichroism and Synchrotron Radtion Circular Dichroism Spectroscopy; IOS Press: Amsterdam, The Netherlands, 2009; pp. 1–18. [Google Scholar]
- Hunter, R.J. Foundations of Colloid Science, 2nd ed.; Oxford Clarendon Press: New York, NY, USA, 2001. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Surfactant | Hydrolysis Efficiency (%) |
|---|---|
| No surfactant | 57 |
| 0.5 wt% (17.33 mM) SDS | 20 |
| 0.5 wt% (8.13 mM) CHAPS | 35 |
| 0.5 wt% (14.90 mM) Zw3-12 | 35 |
| Surfactants | n | Ka (M−1) | R2 | Hydrolysis (%) after 4 Days |
|---|---|---|---|---|
| / | 1.34 | 2.86·106 | 0.9543 | 57 |
| SDS | 0.73 | 2.47·103 | 0.9858 | 20 |
| Zw3-12 | 0.97 | 4.37·104 | 0.9953 | 35 |
| CHAPS | 0.84 | 9.72·103 | 0.9724 | 35 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savić, N.D.; Salazar Marcano, D.E.; Quanten, T.; Parac-Vogt, T.N. Broadening the Scope of Polyoxometalates as Artificial Proteases in Surfactant Solutions: Hydrolysis of Ovalbumin by Zr(IV)-Substituted Keggin Complex. Inorganics 2021, 9, 22. https://0-doi-org.brum.beds.ac.uk/10.3390/inorganics9040022
Savić ND, Salazar Marcano DE, Quanten T, Parac-Vogt TN. Broadening the Scope of Polyoxometalates as Artificial Proteases in Surfactant Solutions: Hydrolysis of Ovalbumin by Zr(IV)-Substituted Keggin Complex. Inorganics. 2021; 9(4):22. https://0-doi-org.brum.beds.ac.uk/10.3390/inorganics9040022
Chicago/Turabian StyleSavić, Nada D., David E. Salazar Marcano, Thomas Quanten, and Tatjana N. Parac-Vogt. 2021. "Broadening the Scope of Polyoxometalates as Artificial Proteases in Surfactant Solutions: Hydrolysis of Ovalbumin by Zr(IV)-Substituted Keggin Complex" Inorganics 9, no. 4: 22. https://0-doi-org.brum.beds.ac.uk/10.3390/inorganics9040022