Dietary Whey Protein Supplementation Increases Immunoglobulin G Production by Affecting Helper T Cell Populations after Antigen Exposure


Abstract
:1. Introduction
2. Materials and Methods
2.1. Dietary Treatment
2.2. Animal Experiment
2.3. Hematological Analysis
2.4. Statistical Analysis
3. Results
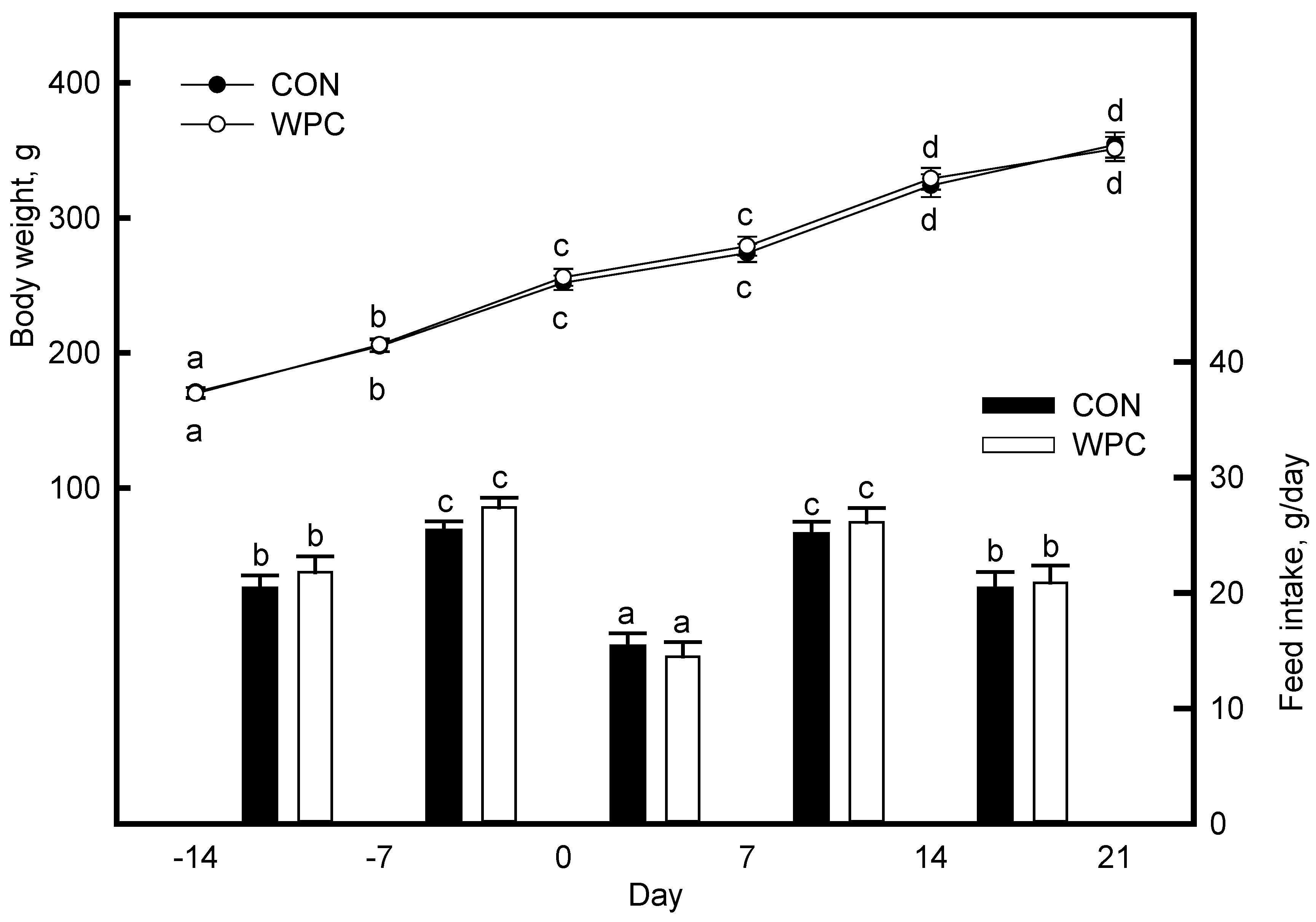
3.1. Dietary Treatments Did Not Affect the Body Weight (BW) and Feed Intake (FI)
3.2. Dietary WPC Changed the White Blood Cells (Neutrophil and Lymphocyte) Population after Antigen Injection
3.3. Dietary WPC Increased B Cell and Helper T Cell Populations after Antigen Injection
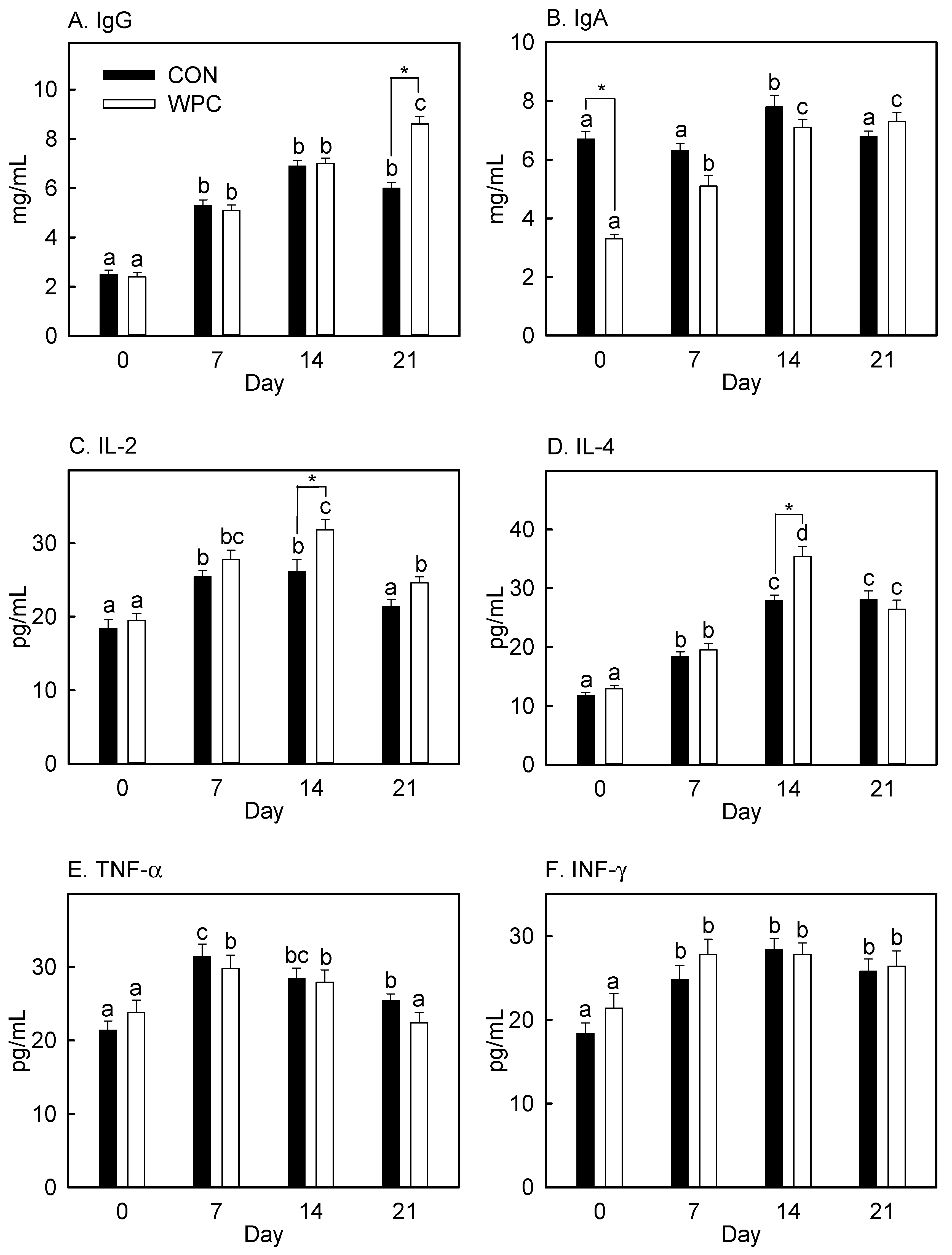
3.4. Dietary WPC Increased Blood IgG and Cytokines (IL-2 and IL-4) after Antigen Injection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Murphy, K.P.; Murphy, K.M.; Travers, P.; Walport, M.; Janeway, C. Janeway’s Immunobiology; Garland Science: New York, NY, USA, 2016. [Google Scholar]
- Parker, D.C. T cell-dependent B cell activation. Annu. Rev. Immunol. 1993, 11, 331–360. [Google Scholar] [CrossRef] [PubMed]
- Woof, J.M.; Burton, D.R. Human antibody-Fc receptor interactions illuminated by crystal structures. Nat. Rev. Immunol. 2004, 4, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Gill, H.S.; Darragh, A.J.; Cross, M.L. Optimizing immunity and gut function in the elderly. J. Nutr. Health Aging 2001, 5, 80–91. [Google Scholar] [PubMed]
- Childs, C.E.; Calder, P.C.; Miles, E.A. Diet and Immune Function. Nutrients 2019, 11, 1933. [Google Scholar] [CrossRef] [Green Version]
- Pae, M.; Meydani, S.N.; Wu, D. The role of nutrition in enhancing immunity in aging. Aging Dis. 2012, 3, 91–129. [Google Scholar] [PubMed]
- Gill, H.S.; Rutherfurd, K.J.; Cross, M.L. Dietary probiotic supplementation enhances natural killer cell activity in the elderly: An investigation of age-related immunological changes. J. Clin. Immunol. 2001, 21, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Chandra, R.K. Effect of vitamin and trace-element supplementation on immune responses and infection in elderly subjects. Lancet 1992, 340, 1124–1127. [Google Scholar] [CrossRef]
- Schilling, J.; Vranjes, N.; Fierz, W.; Joller, H.; Gyurech, D.; Ludwig, E.; Marathias, K.; Geroulanos, S. Clinical outcome and immunology of postoperative arginine, ω-3 fatty acids, and nucleotide-enriched enteral feeding: A randomized prospective comparison with standard enteral and low calorie/low fat IV solutions. Nutrition 1996, 12, 423–429. [Google Scholar] [CrossRef]
- Lupton, J.R.; Brooks, J.; Butte, N.; Caballero, B.; Flatt, J.; Fried, S. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids; National Academy Press: Washington, DC, USA, 2002; Volume 5, pp. 589–768. [Google Scholar]
- Jahan-Mihan, A.; Luhovyy, B.L.; El Khoury, D.; Anderson, G.H. Dietary proteins as determinants of metabolic and physiologic functions of the gastrointestinal tract. Nutrients 2011, 3, 574–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, W.W.; Johnson, C.A.; McCabe, G.P.; Carnell, N.S. Dietary protein requirements of younger and older adults. Am. J. Clin. Nutr. 2008, 88, 1322–1329. [Google Scholar] [CrossRef]
- Wu, G. Dietary protein intake and human health. Food Funct. 2016, 7, 1251–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katona, P.; Katona-Apte, J. The Interaction between Nutrition and Infection. Clin. Infect. Dis. 2008, 46, 1582–1588. [Google Scholar] [CrossRef] [PubMed]
- Obanewa, O.; Newell, M.-L. Maternal nutritional status during pregnancy and infant immune response to routine childhood vaccinations. Future Virol. 2017, 12, 525–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebhardt, B.M.; Newberne, P.M. Nutrition and immunological responsiveness. T-cell function in the offspring of lipotrope and protein-deficient rats. Immunology 1974, 26, 489–495. [Google Scholar] [PubMed]
- Castaneda, C.; Charnley, J.M.; Evans, W.J.; Crim, M.C. Elderly women accommodate to a low-protein diet with losses of body cell mass, muscle function, and immune response. Am. J. Clin. Nutr. 1995, 62, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yin, Y.-L.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef] [Green Version]
- Guadagni, M.; Biolo, G. Effects of inflammation and/or inactivity on the need for dietary protein. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Bounous, G.; Kongshavn, P.A. Influence of dietary proteins on the immune system of mice. J. Nutr. 1982, 112, 1747–1755. [Google Scholar] [CrossRef] [PubMed]
- Bounous, G.; Shenouda, N.; Kongshavn, P.A.; Osmond, D.G. Mechanism of altered B-cell response induced by changes in dietary protein type in mice. J. Nutr. 1985, 115, 1409–1417. [Google Scholar] [CrossRef] [Green Version]
- Madureira, A.R.; Tavares, T.; Gomes, A.M.; Pintado, M.E.; Malcata, F.X. Invited review: Physiological properties of bioactive peptides obtained from whey proteins. J. Dairy Sci. 2010, 93, 437–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, J.R.; Falvo, M.J. Protein—Which is Best? J. Sports Sci. Med. 2004, 3, 118–130. [Google Scholar] [PubMed]
- Nabil, S.; Gauthier, S.F.; Drouin, R.; Poubelle, P.E.; Pouliot, Y. In Vitro Digestion of Proteins and Growth Factors in a Bovine Whey Protein Extract as Determined Using a Computer-Controlled Dynamic Gastrointestinal System (TIM-1). Food Dig. 2011, 2, 13–22. [Google Scholar] [CrossRef]
- Mann, B.; Athira, S.; Sharma, R.; Kumar, R.; Sarkar, P. Bioactive Peptides from Whey Proteins. In Whey Proteins; Deeth, H.C., Bansal, N., Eds.; Academic Press: New York, NY, USA, 2019; Chapter 14; pp. 519–547. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Oh, D.H.; Lee, B.H. Bioactive Peptides. Foods 2017, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Amouheydari, M.; Ehsani, M.R.; Javadi, I. Effect of a dietary supplement composed of hydrolyzed milk proteins and vanillin on the reduction of infection and oxidative stress induced by chemotherapy. J. Food Biochem. 2020, 44. [Google Scholar] [CrossRef]
- Garg, G.; Singh, S.; Singh, A.K.; Rizvi, S.I. Whey protein concentrate supplementation protects rat brain against aging-induced oxidative stress and neurodegeneration. Appl. Physiol. Nutr. Metab. 2018, 43, 437–444. [Google Scholar] [CrossRef]
- Miralles, B.; Hernández-Ledesma, B.; Fernández-Tomé, S.; Amigo, L.; Recio, I. Health-related functional value of dairy proteins and peptides. In Proteins in Food Processing, 2nd ed.; Woodhead Publishing Ltd.: Cambridge, UK, 2018; pp. 523–568. [Google Scholar] [CrossRef]
- Rusu, D.; Drouin, R.; Pouliot, Y.; Gauthier, S.; Poubelle, P.E. A bovine whey protein extract stimulates human neutrophils to generate bioactive IL-1Ra through a NF-κB- and MAPK-dependent mechanism. J. Nutr. 2010, 140, 382–391. [Google Scholar] [CrossRef] [Green Version]
- Badr, G.; Mohany, M.; Metwalli, A. Effects of undenatured whey protein supplementation on CXCL12- and CCL21-mediated B and T cell chemotaxis in diabetic mice. Lipids Health Dis. 2011, 10, 203. [Google Scholar] [CrossRef] [Green Version]
- Badr, G.; Ebaid, H.; Mohany, M.; Abuelsaad, A.S. Modulation of immune cell proliferation and chemotaxis towards CC chemokine ligand (CCL)-21 and CXC chemokine ligand (CXCL)-12 in undenatured whey protein-treated mice. J. Nutr. Biochem. 2012, 23, 1640–1646. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, J.; Dupont, C.; Lemieux, P. Anti-inflammatory potential of a malleable matrix composed of fermented whey proteins and lactic acid bacteria in an atopic dermatitis model. J. Inflamm. 2007, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.F.; Middleton, N.; Montgomery, M.; Dey, M.; Carr, R.I. Immunostimulation of murine spleen cells by materials associated with bovine milk protein fractions. J. Dairy Sci. 1998, 81, 1825–1832. [Google Scholar] [CrossRef]
- Ebert, E.C.; Roberts, A.I. Lamina propria lymphocytes produce interferon-γ and develop suppressor activity in response to lactoglobulin. Dig. Dis. Sci. 2001, 46, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Tai, C.S.; Chen, Y.Y.; Chen, W.L. β-Lactoglobulin Influences Human Immunity and Promotes Cell Proliferation. BioMed Res. Int. 2016, 2016, 7123587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, C.W.; Seow, H.F.; Husband, A.J.; Regester, G.O.; Watson, D.L. Effects of purified bovine whey factors on cellular immune functions in ruminants. Vet. Immunol. Immunopathol. 1997, 56, 85–96. [Google Scholar] [CrossRef]
- Akal, C. Benefits of whey proteins on human health. In Dairy in Human Health and Disease Across the Lifespan; Academic Press: New York, NY, USA, 2017; pp. 363–372. [Google Scholar] [CrossRef]
- Vasconcelos, Q.; Bachur, T.P.R.; Aragão, G.F. Whey protein supplementation and its potentially adverse effects on health: A systematic review. Appl. Physiol. Nutr. Metab. 2021, 46, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Laiosa, M. Steroid Hormones and their Effect on the Immune System. In Encyclopedic Reference of Immunotoxicology; Vohr, H.-W., Ed.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 603–608. [Google Scholar] [CrossRef]
- Krzych, U.; Strausser, H.R.; Bressler, J.P.; Goldstein, A.L. Quantitative Differences in Immune Responses during the Various Stages of the Estrous Cycle in Female BALB/c Mice. J. Immunol. 1978, 121, 1603–1605. [Google Scholar] [CrossRef]
- Kimmel, S.R. Vaccine adverse events: Separating myth from reality. Am. Fam. Physician 2002, 66, 2113. [Google Scholar] [PubMed]
- Kobayashi, S.D.; Malachowa, N.; DeLeo, F.R. Neutrophils and Bacterial Immune Evasion. J. Innate Immun. 2018, 10, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Takiishi, T.; Fenero, C.I.M.; Câmara, N.O.S. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue Barriers 2017, 5, e1373208. [Google Scholar] [CrossRef]
- Arbizu, S.; Chew, B.; Mertens-Talcott, S.U.; Noratto, G. Commercial whey products promote intestinal barrier function with glycomacropeptide enhanced activity in downregulating bacterial endotoxin lipopolysaccharides (LPS)-induced inflammation: In vitro. Food Funct. 2020, 11, 5842–5852. [Google Scholar] [CrossRef]
- Kotler, B.M.; Kerstetter, J.E.; Insogna, K.L. Claudins, dietary milk proteins, and intestinal barrier regulation. Nutr. Rev. 2013, 71, 60–65. [Google Scholar] [CrossRef]
- Hering, N.A.; Andres, S.; Fromm, A.; van Tol, E.A.; Amasheh, M.; Mankertz, J.; Fromm, M.; Schulzke, J.D. Transforming growth factor-β, a whey protein component, strengthens the intestinal barrier by upregulating claudin-4 in HT-29/B6 cells. J. Nutr. 2011, 141, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Xiao, K.; Jiao, L.; Cao, S.; Song, Z.; Hu, C.; Han, X. Whey protein concentrate enhances intestinal integrity and influences transforming growth factor-β1 and mitogen-activated protein kinase signalling pathways in piglets after lipopolysaccharide challenge. Br. J. Nutr. 2016, 115, 984–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macpherson, A.J.; Slack, E. The functional interactions of commensal bacteria with intestinal secretory IgA. Curr. Opin. Gastroenterol. 2007, 23, 673–678. [Google Scholar] [CrossRef]
- Azrad, M.; Hamo, Z.; Tkhawkho, L.; Peretz, A. Elevated serum immunoglobulin A levels in patients with Clostridium difficile infection are associated with mortality. Pathog. Dis. 2018, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Sette, A.; Weiskopf, D. Cytotoxic CD4 T Cells: Differentiation, Function, and Application to Dengue Virus Infection. Front. Immunol. 2016, 7, 531. [Google Scholar] [CrossRef]
- Cooper, M.D. The early history of B cells. Nat. Rev. Immunol. 2015, 15, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Crotty, S. A brief history of T cell help to B cells. Nat. Rev. Immunol. 2015, 15, 185–189. [Google Scholar] [CrossRef]
- Coldsmith, M.A.; Green, W.C. The Cytokine Handbook, 2nd ed.; Thomson, A., Ed.; Academic Press: London, UK, 1994; pp. 55–80. [Google Scholar]
- Hatakeyama, M.; Taniguchi, T. Peptide Growth Factors and Their Receptors I; Sporn, M.S., Roberts, A.B., Eds.; Springer: New York, NY, USA, 1990; p. 523. [Google Scholar]
- Liao, W.; Lin, J.X.; Leonard, W.J. IL-2 family cytokines: New insights into the complex roles of IL-2 as a broad regulator of T helper cell differentiation. Curr. Opin. Immunol. 2011, 23, 598–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, M.A.; Hural, J. Functions of IL-4 and control of its expression. Crit. Rev. Immunol. 1997, 17, 1–32. [Google Scholar] [CrossRef]
- Shtrichman, R.; Samuel, C.E. The role of gamma interferon in antimicrobial immunity. Curr. Opin. Microbiol. 2001, 4, 251–259. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Ingredient | CON | WPC |
|---|---|---|
| Amino acids mixture 1 | 10.0 | - |
| Whey protein 2 | - | 10.0 |
| Ground whole wheat | 341.7 | 341.7 |
| Ground whole yellow corn | 200.0 | 200.0 |
| Ground whole oats | 120.0 | 120.0 |
| Wheat middlings | 100.0 | 100.0 |
| Fish meal | 90.0 | 90.0 |
| Soybean meal | 50.0 | 50.0 |
| Soybean oil | 25.0 | 25.0 |
| Alfalfa meal | 20.0 | 20.0 |
| Corn gluten meal | 20.0 | 20.0 |
| Dicalcium phosphate | 15.0 | 15.0 |
| Brewer’s dried yeast | 10.0 | 10.0 |
| Ground limestone | 5.0 | 5.0 |
| Salt | 5.0 | 5.0 |
| Vitamin premixture 3 | 2.5 | 2.5 |
| Mineral premixture 4 | 2.5 | 2.5 |
| Choline chloride | 1.3 | 1.3 |
| L-Lysine | 1.0 | 1.0 |
| DL-Methionine | 1.0 | 1.0 |
| Variable | Day 0 | Day 7 | Day 14 | Day 21 | ||||
|---|---|---|---|---|---|---|---|---|
| CON | WPC | CON | WPC | CON | WPC | CON | WPC | |
| Red blood cells, 106 cells/µL | 8.8 ± 0.6 | 8.7 ± 0.5 | 8.9 ± 0.4 | 8.8 ± 0.4 | 8.2 ± 0.5 | 8.5 ± 0.5 | 8.3 ± 0.4 | 8.3 ± 0.5 |
| Platelets, 106 cells/µL | 1.1 ± 0.2 | 1.1 ± 0.3 | 1.3 ± 0.2 | 1.2 ± 0.2 | 1.2 ± 0.3 | 1.1 ± 0.2 | 1.2 ± 0.1 | 1.4 ± 0.2 |
| White blood cells, 103 cells/µL | 8.9 ± 0.8 | 7.3 ± 0.9 | 11.8 ± 1.1 | 12.4 ± 1.0 | 9.8 ± 0.7 | 12.4 ± 0.8 | 13.1 ± 1.1 | 12.8 ± 1.0 |
| Neutrophils, % | 21.0 ± 1.6 a | 21.0 ± 2.2 a | 21.7 ± 1.8 a | 17.9 ± 1.1 *,b | 17.4 ± 1.1 b | 12.0 ± 0.8 *,c | 17.8 ± 1.4 b | 10.1 ± 0.7 *,c |
| Lymphocytes, % | 76.9 ± 2.4 | 76.6 ± 2.8 a | 76.5 ± 2.1 | 80.0 ± 2.2 a | 80.2 ± 2.1 | 85.2 ± 2.1 b | 79.3 ± 1.8 | 88.2 ± 2.2 *,b |
| etc. 1, % | 2.1 ± 0.4 | 2.4 ± 0.5 | 1.8 ± 0.2 | 2.1 ± 0.4 | 2.4 ± 0.5 | 2.8 ± 0.4 | 2.9 ± 0.4 | 1.8 ± 0.3 |
| CON | WPC | |
|---|---|---|
| B cells (CD45R+), % | ||
| Day 0 | 20.6 ± 1.8 | 22.3 ± 1.2 a |
| Day 7 | 21.4 ± 1.1 | 20.8 ± 1.8 a |
| Day 14 | 23.0 ± 1.8 | 26.0 ± 1.9 b |
| Day 21 | 24.0 ± 2.1 | 28.1 ± 3.1 *,b |
| T cells (CD3+), % | ||
| Day 0 | 56.3 ± 1.2 | 54.3 ± 1.3 a |
| Day 7 | 55.1 ± 1.8 | 59.2 ± 2.1 b |
| Day 14 | 57.2 ± 1.8 | 59.2 ± 2.4 b |
| Day 21 | 55.3 ± 2.1 | 60.1 ± 1.8 *,b |
| Cytotoxic T cells (CD3+/CD8+), % | ||
| Day 0 | 25.3 ± 0.8 | 22.4 ± 0.6 * |
| Day 7 | 22.8 ± 1.8 | 22.8 ± 1.4 |
| Day 14 | 20.4 ± 1.4 | 20.9 ± 1.7 |
| Day 21 | 20.8 ± 1.5 | 20.7 ± 1.4 |
| Helper T cells (CD3+/CD4+), % | ||
| Day 0 | 31.0 ± 1.8 | 31.9 ± 1.2 a |
| Day 7 | 32.3 ± 1.1 | 36.4 ± 1.9 *,b |
| Day 14 | 36.8 ± 2.1 | 38.3 ± 1.8 b |
| Day 21 | 34.5 ± 1.3 | 39.4 ± 1.5 *,b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ha, D.J.; Kim, J.; Kim, S.; Go, G.-W.; Whang, K.-Y. Dietary Whey Protein Supplementation Increases Immunoglobulin G Production by Affecting Helper T Cell Populations after Antigen Exposure. Foods 2021, 10, 194. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10010194
Ha DJ, Kim J, Kim S, Go G-W, Whang K-Y. Dietary Whey Protein Supplementation Increases Immunoglobulin G Production by Affecting Helper T Cell Populations after Antigen Exposure. Foods. 2021; 10(1):194. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10010194
Chicago/Turabian StyleHa, Dong Jin, Jonggun Kim, Saehun Kim, Gwang-Woong Go, and Kwang-Youn Whang. 2021. "Dietary Whey Protein Supplementation Increases Immunoglobulin G Production by Affecting Helper T Cell Populations after Antigen Exposure" Foods 10, no. 1: 194. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10010194