
Bitter Gourd Honey Ameliorates Hepatic and Renal Diabetic Complications on Type 2 Diabetes Rat Models by Antioxidant, Anti-Inflammatory, and Anti-Apoptotic Mechanisms

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Kits
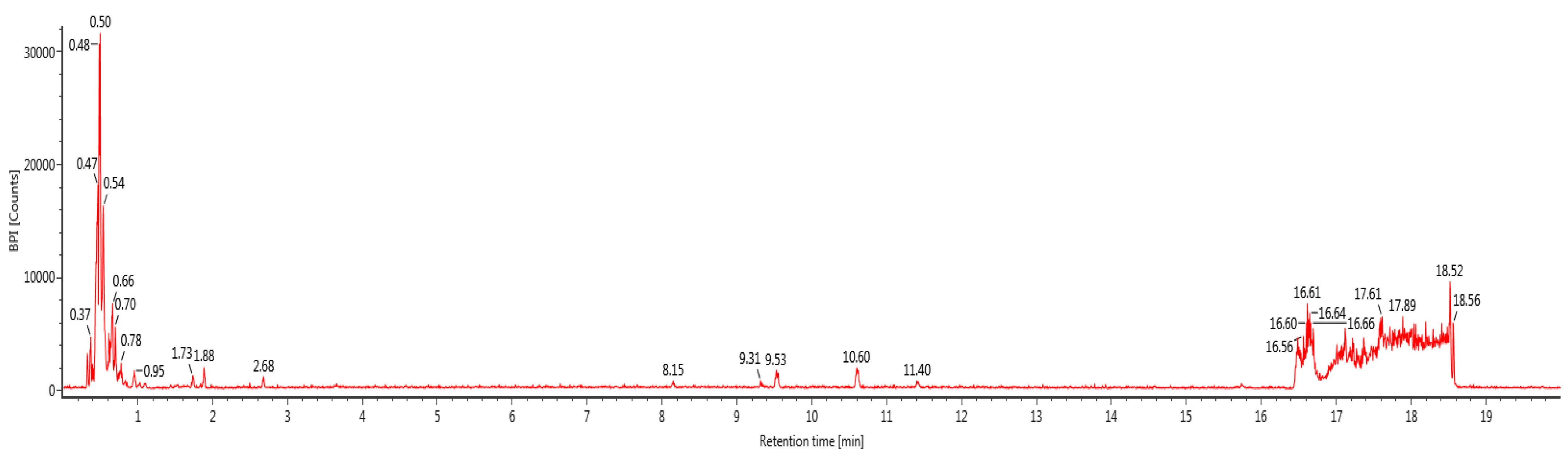
2.2. Liquid Chromatography-Mass Spectroscopy (LC-MS)
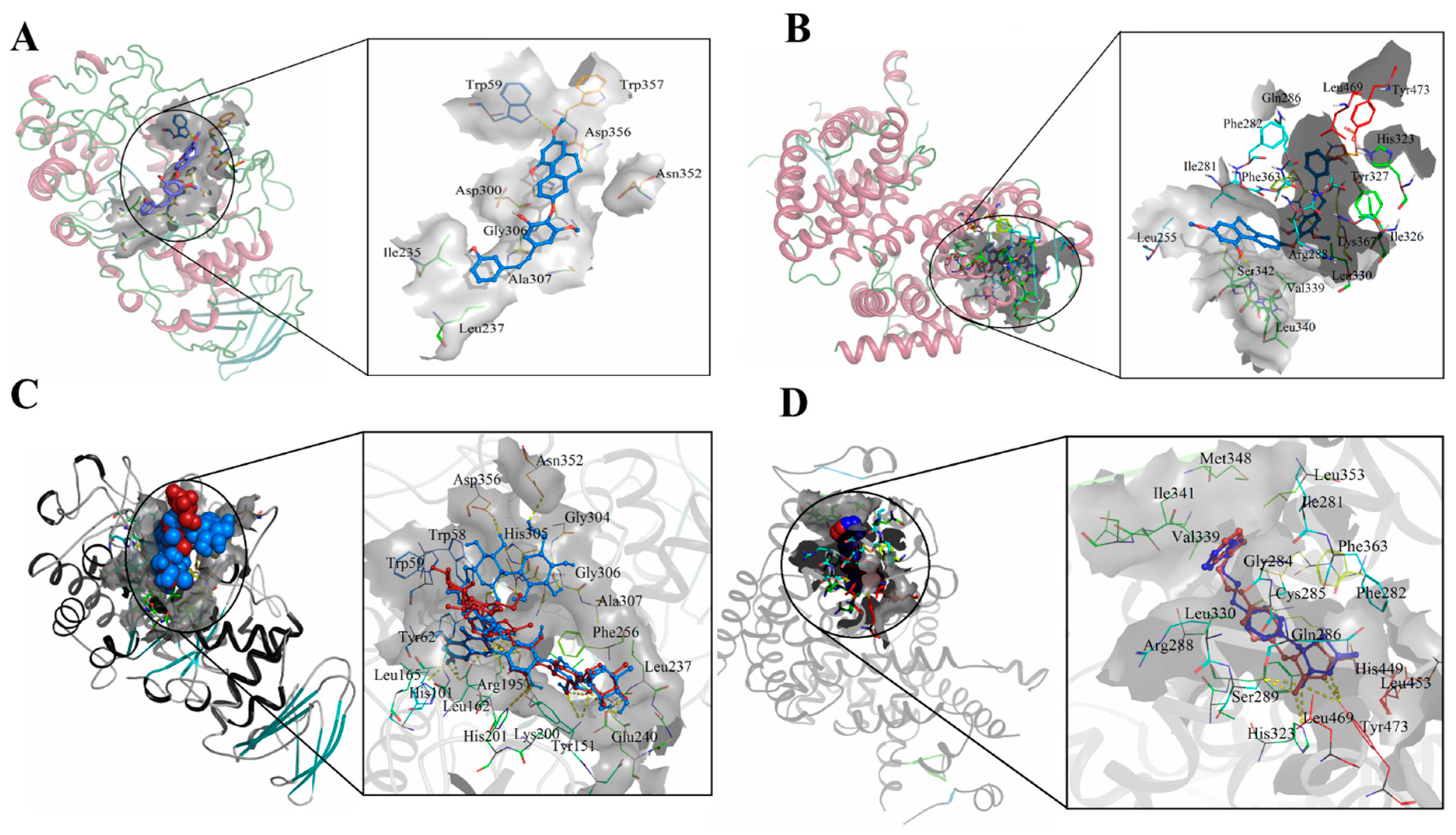
2.3. Molecular Docking
2.4. Animal Studies
2.5. Induction of Diabetes in Animal Model
2.6. Experimental Design
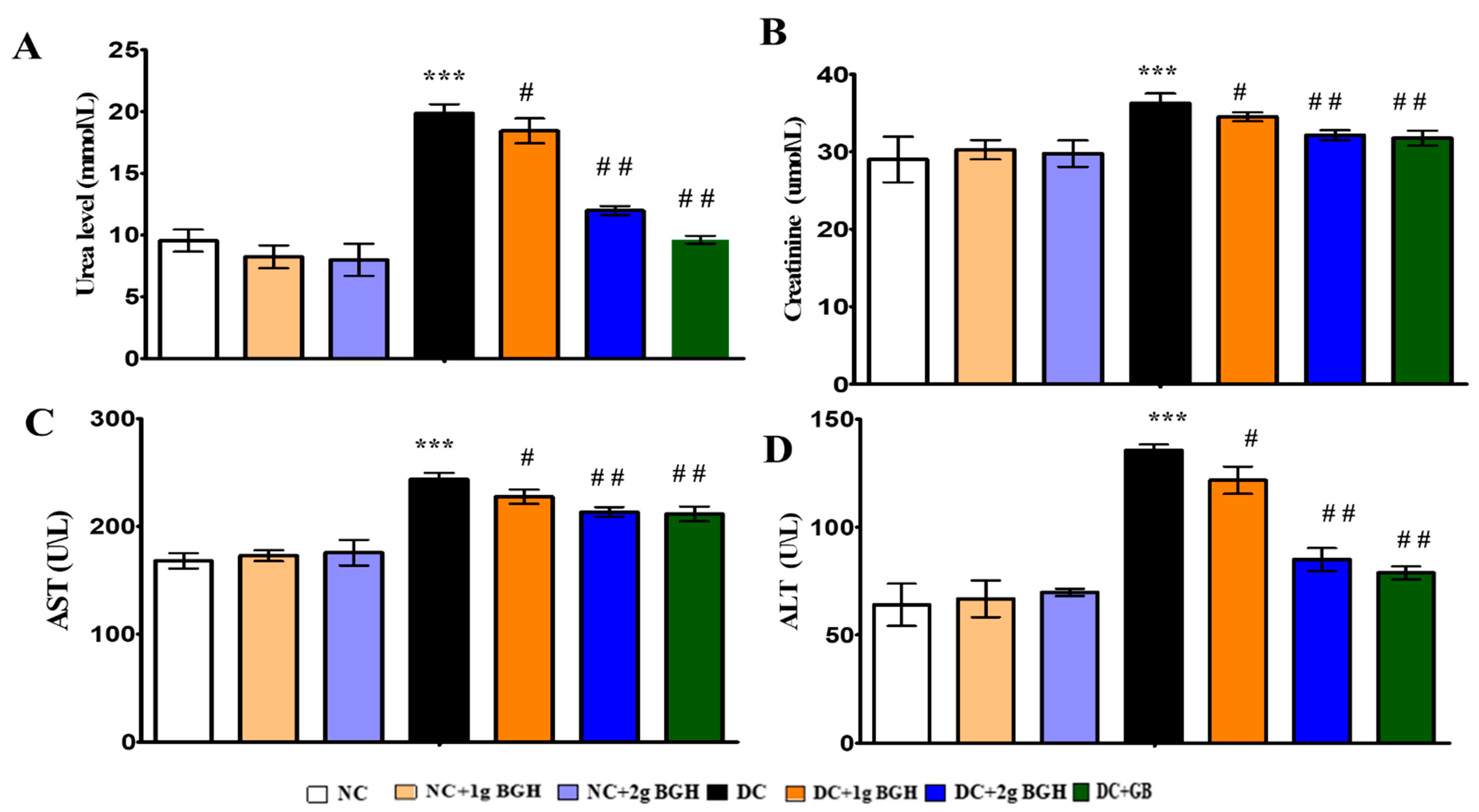
2.7. Determination of Serum Hepatic and Renal Markers
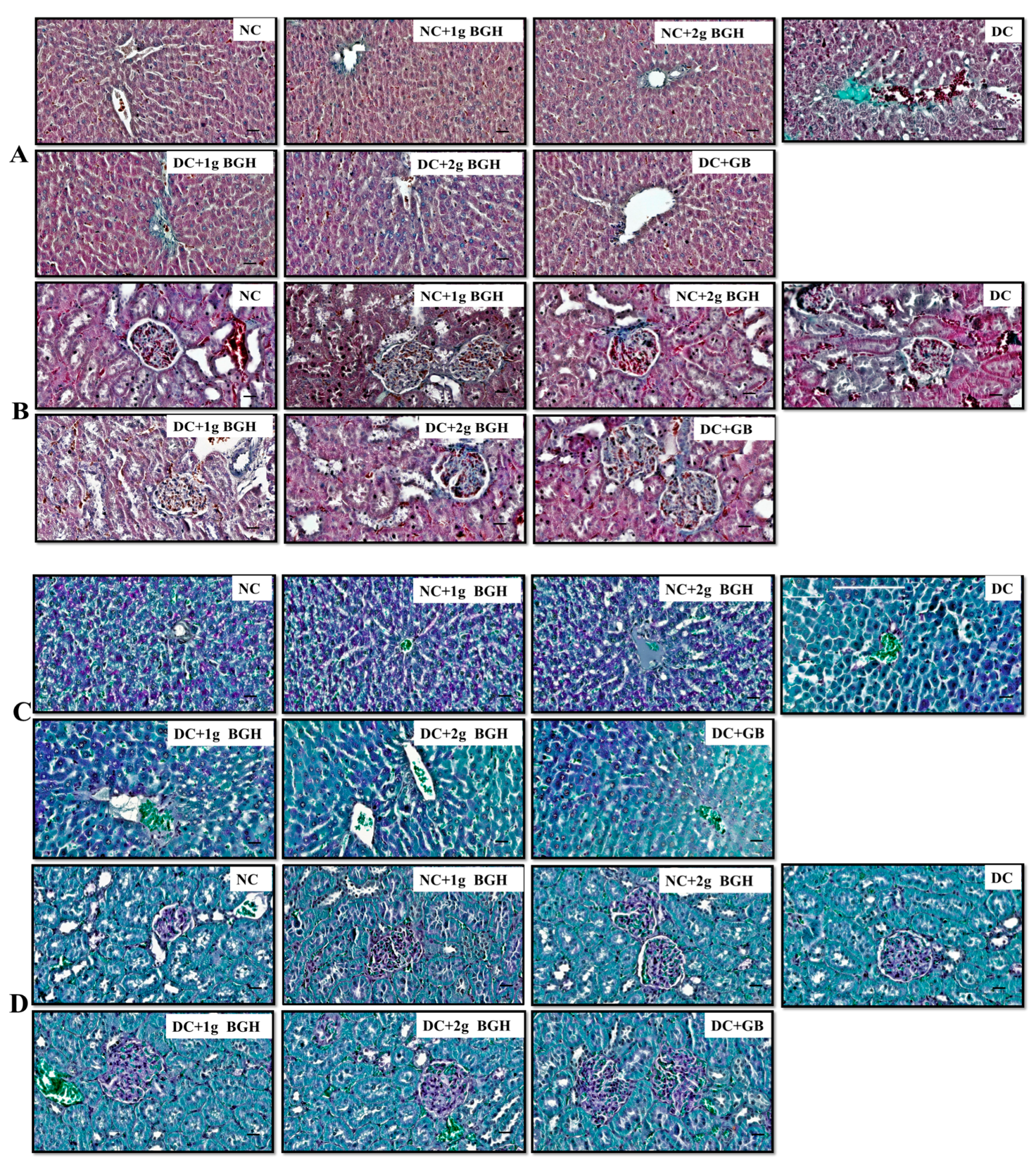
2.8. Masson’s Trichome and Periodic acid Schiff (PAS) Staining of Liver and Kidney Tissue
2.9. Determining the Levels of Antioxidant Enzyme and Oxidative Stress Marker in Liver and Kidney Homogenates
2.10. Immunohistochemistry and Immunofluorescence
2.11. Statistical Analysis
3. Results
3.1. Identification of Polyphenols in BGH by LC-ESI-MS/MS
3.2. Molecular Docking against Detected Compounds in LCMS
3.3. Effect of BGH on Liver, Kidney Functional Markers
3.4. Effect of BGH on Histopathological Changes in Diabetic Rats
3.5. Effect of BGH on Antioxidant Enzyme in Liver and Kidney Homogenates
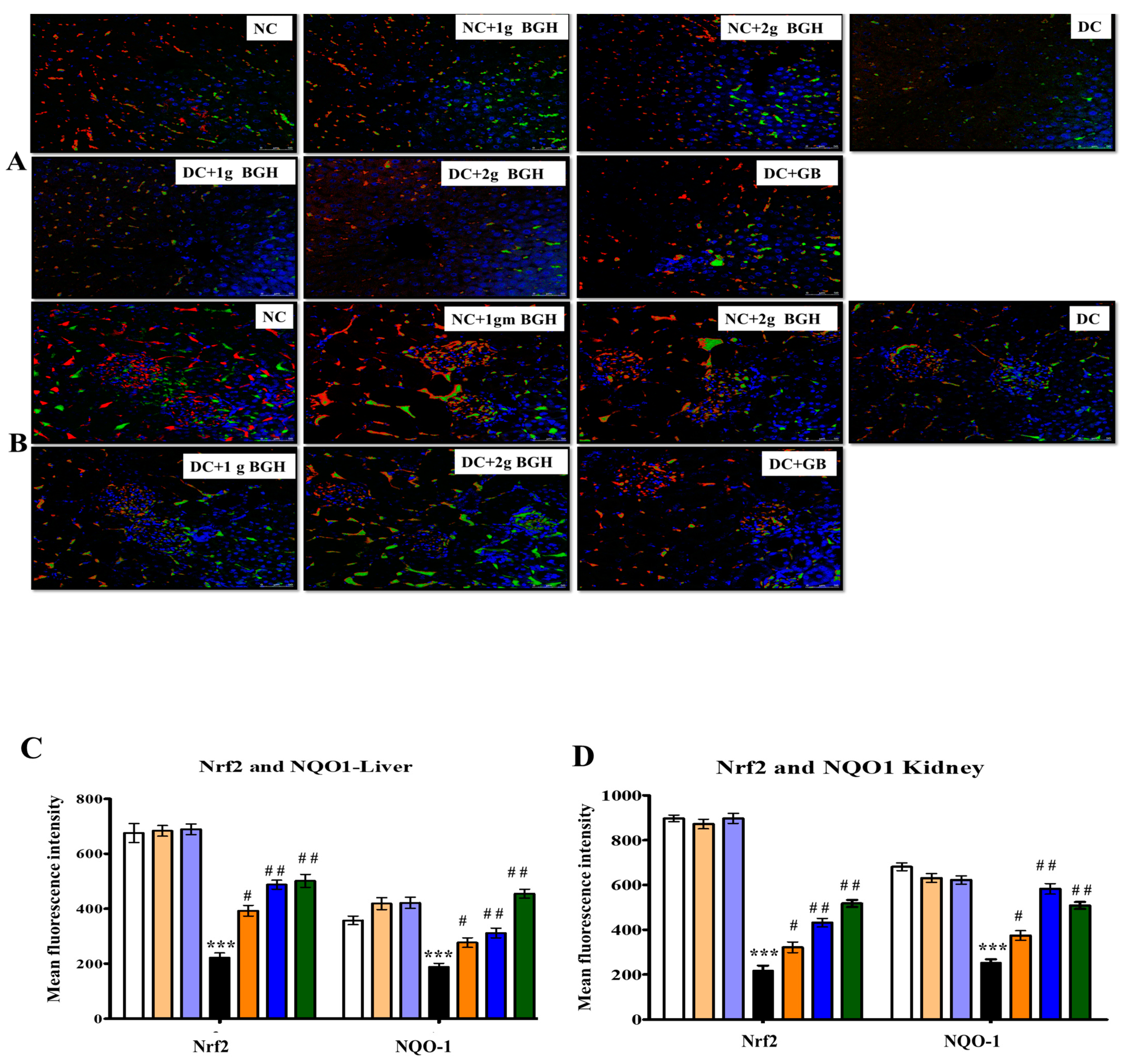
3.6. Effect of BGH on the Expression of Transcription Factor Nrf2 and NOQ1 in the Hepatocyte and Renal Tissues of Diabetic Rats
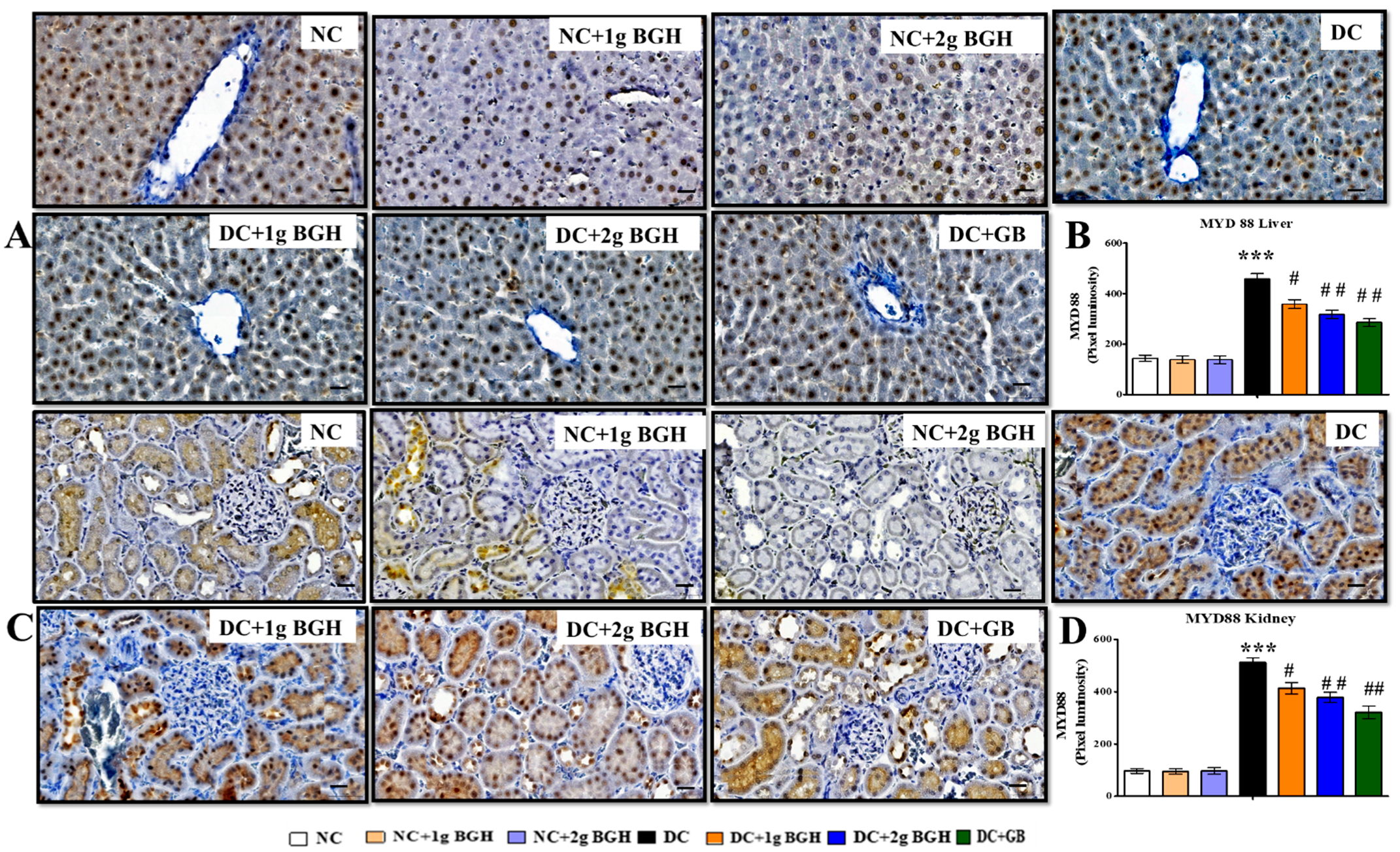
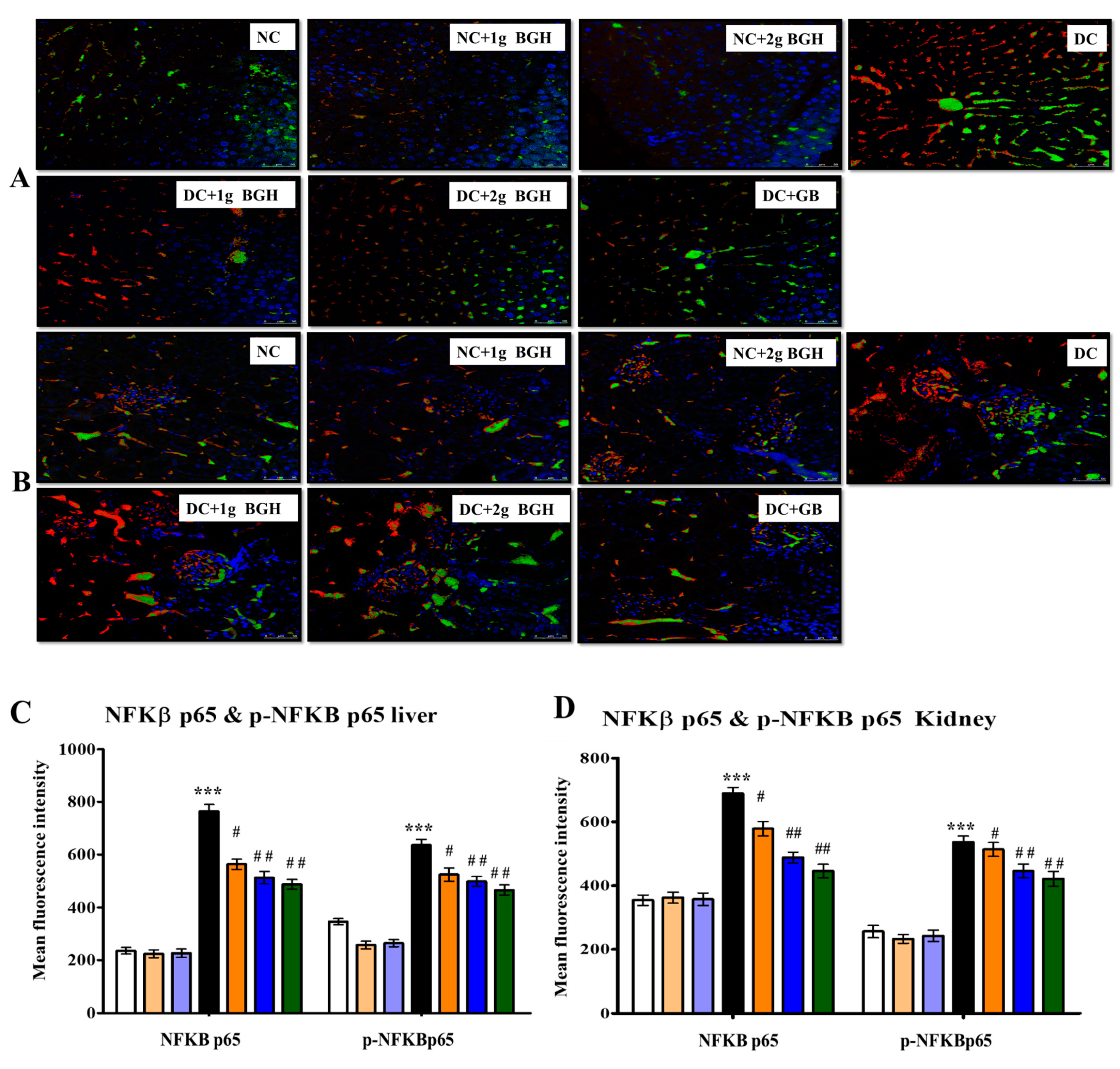
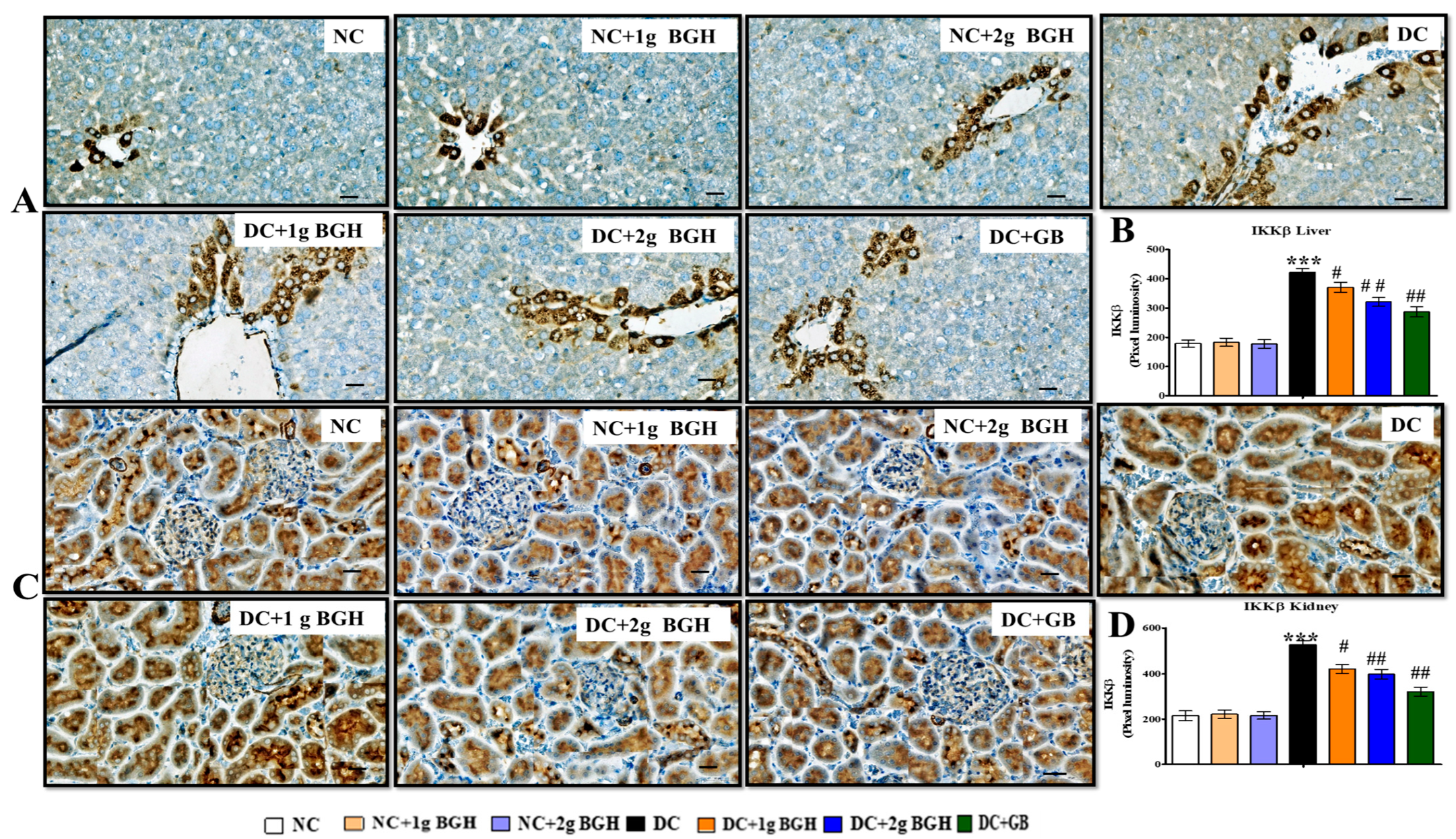
3.7. Effect of BGH on the Expression of Inflammation Markers, MyD88–IKKB–NF-κB in the Hepatocyte and Renal Tissues of Diabetic Rats
3.8. Effect of BGH on the Expression of Apoptotic Marker in the Renal and Hepatocyte Tissues of Diabetic Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Boutten, A.; Goven, D.; Artaud-Macari, E.; Boczkowski, J.; Bonay, M. NRF2 targeting: A promising therapeutic strategy in chronic obstructive pulmonary disease. Trends Mol. Med. 2011, 17, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.-Y.; Kwak, M.-K.; Pi, J. Nrf2 in Host Defense: Over the Rainbow. Oxidative Med. Cell. Longev. 2013, 2013, 975839. [Google Scholar] [CrossRef] [Green Version]
- Blachier, M.; Leleu, H.; Peck-Radosavljevic, M.; Valla, D.C.; Roudot-Thoraval, F. The burden of liver disease in Europe: A review of available epidemiological data. J. Hepatol. 2013, 58, 593–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caldwell, S.H.; Oelsner, D.H.; Iezzoni, J.C.; Hespenheide, E.E.; Battle, E.H.; Driscoll, C.J. Cryptogenic cirrhosis: Clinical characterization and risk factors for underlying disease. Hepatology 1999, 29, 664–669. [Google Scholar] [CrossRef]
- Huang, E.S.; Liu, J.Y.; Moffet, H.H.; John, P.M.; Karter, A.J. Glycemic Control, Complications, and Death in Older Diabetic Patients: The Diabetes and Aging Study. Diabetes Care 2011, 34, 1329–1336. [Google Scholar] [CrossRef] [Green Version]
- Grandfils, N.; Detournay, B.; Attali, C.; Joly, D.; Simon, D.; Vergès, B.; Toussi, M.; Briand, Y.; Delaitre, O. Glucose Lowering Therapeutic Strategies for Type 2 Diabetic Patients with Chronic Kidney Disease in Primary Care Setting in France: A Cross-Sectional Study. Int. J. Endocrinol. 2013, 2013, 640632. [Google Scholar] [CrossRef] [Green Version]
- Kayama, Y.; Raaz, U.; Jagger, A.; Adam, M.; Schellinger, I.N.; Sakamoto, M.; Suzuki, H.; Toyama, K.; Spin, J.M.; Tsao, P.S. Diabetic Cardiovascular Disease Induced by Oxidative Stress. Int. J. Mol. Sci. 2015, 16, 25234–25263. [Google Scholar] [CrossRef]
- Kwak, M.-K.; Egner, P.A.; Dolan, P.M.; Ramos-Gomez, M.; Groopman, J.D.; Itoh, K.; Yamamoto, M.; Kensler, T.W. Role of phase 2 enzyme induction in chemoprotection by dithiolethiones. Mutat. Res. Mol. Mech. Mutagen. 2001, 480-481, 305–315. [Google Scholar] [CrossRef]
- Itoh, K.; Chiba, T.; Takahashi, S.; Ishii, T.; Igarashi, K.; Katoh, Y.; Oyake, T.; Hayashi, N.; Satoh, K.; Hatayama, I.; et al. An Nrf2/Small Maf Heterodimer Mediates the Induction of Phase II Detoxifying Enzyme Genes through Antioxidant Response Elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar] [CrossRef]
- Chu, Y.-F.; Sun, J.; Wu, X.; Liu, R.H. Antioxidant and Antiproliferative Activities of Common Vegetables. J. Agric. Food Chem. 2002, 50, 6910–6916. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Khalil, I.; Sulaiman, S.A.; Gan, S.H. Physicochemical and antioxidant properties of Malaysian honeys produced by Apis cerana, Apis dorsata and Apis mellifera. BMC Complement. Altern. Med. 2013, 13, 43. [Google Scholar] [CrossRef] [Green Version]
- Bagul, P.K.; Middela, H.; Mattapally, S.; Padiya, R.; Bastia, T.; Madhusudana, K.; Reddy, B.R.; Chakravarty, S.; Banerjee, S.K. Attenuation of insulin resistance, metabolic syndrome and hepatic oxidative stress by resveratrol in fructose-fed rats. Pharmacol. Res. 2012, 66, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Tanigawa, S.; Fujii, M.; Hou, D.-X. Action of Nrf2 and Keap1 in ARE-mediated NQO1 expression by quercetin. Free Radic. Biol. Med. 2007, 42, 1690–1703. [Google Scholar] [CrossRef] [PubMed]
- Erejuwa, O.O.; Sulaiman, S.A.; Ab Wahab, M.S. Honey: A Novel Antioxidant. Molecules 2012, 17, 4400–4423. [Google Scholar] [CrossRef] [PubMed]
- Rajabzadeh, A.; Saki, G.; Hemadi, M.; Khodadadi, A.; Sarkaki, A. Evaluation of effect of honey and vitamin E on apoptosis in testes of rat exposed to noise stress. Jentashapir J. Cell. Mol. Biol. 2013, 3, 523–532. [Google Scholar]
- Erejuwa, O.; Sulaiman, S.; Ab Wahab, M.; Sirajudeen, K.; Salzihan, M. Effects of Malaysian Tualang honey supplementation on glycemia, free radical scavenging enzymes and markers of oxidative stress in kidneys of normal and streptozotocin-induced diabetic rats. Int. J. Cardiol. 2009, 137, S45. [Google Scholar] [CrossRef]
- Owoyele, B.V.; Adenekan, O.T.; Soladoye, A.O. Effects of honey on inflammation and nitric oxide production in Wistar rats. J. Chin. Integr. Med. 2011, 9, 447–452. [Google Scholar] [CrossRef]
- Tonks, A.; Cooper, R.; Jones, K.; Blair, S.; Parton, J. Honey stimulates inflammatory cytokine production from monocytes. Cytokine 2003, 21, 242–247. [Google Scholar] [CrossRef]
- Bashkaran, K.; Zunaina, E.; Bakiah, S.; Sulaiman, S.A.; Sirajudeen, K.; Naik, V. Anti-inflammatory and antioxidant effects of Tualang honey in alkali injury on the eyes of rabbits: Experimental animal study. BMC Complement. Altern. Med. 2011, 11, 90. [Google Scholar] [CrossRef] [Green Version]
- Hussein, S.Z.; Mohd Yusoff, K.; Makpol, S.; Mohd Yusof, Y.A. Gelam honey inhibits the production of proinflammatory, mediators NO, PGE2, TNF-α, and IL-6 in carra-geenan-induced acute paw edema in rats. Evid.-Based Complementary Altern. Med. 2012, 2012, 109636. [Google Scholar] [CrossRef] [Green Version]
- Karim, K.; Giribabu, N.; Salleh, N. Marantodes pumilum Var Alata (Kacip Fatimah) ameliorates derangement in RANK/RANKL/OPG pathway and reduces inflammation and oxidative stress in the bone of estrogen-deficient female rats with type-2 diabetes. Phytomedicine 2021, 91, 153677. [Google Scholar] [CrossRef]
- Dallakyan, S.; Olson, A.J. Small-molecule library screening by docking with PyRx. In Chemical Biology; Humana Press: New York, NY, USA, 2015; pp. 243–250. [Google Scholar]
- Williams, L.K.; Zhang, X.; Caner, S.; Tysoe, C.; Nguyen, N.T.; Wicki, J.; Williams, D.E.; Coleman, J.; McNeill, J.H.; Yuen, V. The amylase inhibitor montbretin A reveals a new glycosidase inhibition motif. Nature Chem. Biol. 2015, 11, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Liberato, M.V.; Nascimento, A.S.; Ayers, S.D.; Lin, J.Z.; Cvoro, A.; Silveira, R.L.; Martínez, L.; Souza, P.C.; Saidemberg, D.; Deng, T. Medium chain fatty acids are selective peroxisome proliferator activated receptor (PPAR) γ activators and pan-PPAR partial agonists. PLoS ONE 2012, 7, e36297. [Google Scholar]
- Hemmati, M.; Karamian, M.; Malekaneh, M. Anti-atherogenic Potential of Natural Honey: Anti-diabetic and Antioxidant Approaches. J. Pharm. Pharmacol. 2015, 3, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Aziz, M.S.A.; Giribabu, N.; Rao, P.V.; Salleh, N. Pancreatoprotective effects of Geniotrigona thoracica stingless bee honey in streptozotocin-nicotinamide-induced male diabetic rats. Biomed. Pharmacother. 2017, 89, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Sokolovska, J.; Isajevs, S.; Sugoka, O.; Sharipova, J.; Paramonova, N.; Isajeva, D.; Rostoka, E.; Sjakste, T.; Sjakste, N.; Kalvinsh, I. Comparison of the Effects of Glibenclamide on Metabolic Parameters, GLUT1 Expression, and Liver Injury in Rats With Severe and Mild Streptozotocin-Induced Diabetes Mellitus. Medicine 2012, 48, 78. [Google Scholar] [CrossRef] [Green Version]
- Esterbauer, H.; Cheeseman, K.H. Determination of aldehydic lipid peroxidation products: Malonaldehyde and 4-hydroxynonenal. Methods Enzymol. 1990, 186, 407–421. [Google Scholar] [PubMed]
- Misra, H.P.; Fridovich, I. The Role of Superoxide Anion in the Autoxidation of Epinephrine and a Simple Assay for Superoxide Dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A. [136] Assay of catalases and peroxidases. Methods Enzym. 1955, 2, 764–775. [Google Scholar]
- Rotruck, J.T.; Pope, A.L.; Ganther, H.E.; Swanson, A.B.; Hafeman, D.G.; Hoekstra, W.G. Selenium: Biochemical Role as a Component of Glutathione Peroxidase. Science 1973, 179, 588–590. [Google Scholar] [CrossRef]
- Cai, L.; Kang, Y.J. Oxidative stress and diabetic cardiomyopathy: A brief review. Cardiovasc. Toxicol. 2001, 1, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Inoguchi, T.; Li, P.; Umeda, F.; Yu, H.Y.; Kakimoto, M.; Imamura, M.; Aoki, T.; Etoh, T.; Hashimoto, T.; Naruse, M.; et al. High glucose level and free fatty acid stimulate reactive oxygen species production through protein kinase C--dependent activation of NAD(P)H oxidase in cultured vascular cells. Diabetes 2000, 49, 1939–1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishikawa, T.; Edelstein, D.; Du, X.L.; Yamagishi, S.-I.; Matsumura, T.; Kaneda, Y.; Yorek, M.A.; Beebe, D.J.; Oates, P.J.; Hammes, H.-P.; et al. Normalizing mitochondrial superoxide production blocks three pathways of hyperglycaemic damage. Nature 2000, 404, 787–790. [Google Scholar] [CrossRef]
- Rask-Madsen, C.; King, G.L. Mechanisms of Disease: Endothelial dysfunction in insulin resistance and diabetes. Nat. Clin. Pract. Endocrinol. Metab. 2007, 3, 46–56. [Google Scholar] [CrossRef]
- Qinna, N.; Badwan, A. Impact of streptozotocin on altering normal glucose homeostasis during insulin testing in diabetic rats compared to normoglycemic rats. Drug Des. Dev. Ther. 2015, 9, 2515–2525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandhi, G.R.; Vasconcelos, A.B.S.; Wu, D.-T.; Li, H.-B.; Antony, P.J.; Li, H.; Geng, F.; Gurgel, R.Q.; Narain, N.; Gan, R.-Y. Citrus Flavonoids as Promising Phytochemicals Targeting Diabetes and Related Complications: A Systematic Review of In Vitro and In Vivo Studies. Nutrients 2020, 12, 2907. [Google Scholar] [CrossRef]
- Weston, R.J. The contribution of catalase and other natural products to the antibacterial activity of honey: A review. Food Chem. 2000, 71, 235–239. [Google Scholar] [CrossRef]
- Erejuwa, O.; Sulaiman, S.; Wahab, M.; Sirajudeen, K.; Salleh, M.M.; Gurtu, S. Antioxidant protection of Malaysian tualang honey in pancreas of normal and streptozotocin-induced diabetic rats. Ann. D’endocrinologie 2010, 71, 291–296. [Google Scholar] [CrossRef]
- Plumb, G.W.; de Pascual-Teresa, S.; Santos-Buelga, C.; Rivas-Gonzalo, J.-C.; Williamson, G. Antioxidant properties of gallocatechin and prodelphinidins from pomegranate peel. Redox Rep. 2002, 7, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Ying, Z.-H.; Li, H.-M.; Yu, W.-Y.; Yu, C.-H. Iridin Prevented Against Lipopolysaccharide-Induced Inflammatory Responses of Macrophages via Inactivation of PKM2-Mediated Glycolytic Pathways. J. Inflamm. Res. 2021, 14, 341–354. [Google Scholar] [CrossRef]
- Cheung, Y.; Meenu, M.; Yu, X.; Xu, B. Phenolic acids and flavonoids profiles of commercial honey from different floral sources and geographic sources. Int. J. Food Prop. 2019, 22, 290–308. [Google Scholar] [CrossRef]
- Date, K.; Satoh, A.; Iida, K.; Ogawa, H. Pancreatic α-amylase controls glucose assimilation by duodenal retrieval through N-glycan-specific binding, en-docytosis, and degradation. J. Biol. Chem. 2015, 290, 17439–17450. [Google Scholar] [CrossRef] [Green Version]
- Derosa, G.; Maffioli, P. Peroxisome proliferator-activated receptor-γ (PPAR-γ) agonists on glycemic control, lipid profile and cardiovascular risk. Curr. Mol. Pharmacol. 2012, 5, 272–281. [Google Scholar] [CrossRef]
- Khalil, M.; Alam, N.; Moniruzzaman, M.; Sulaiman, S.A.; Gan, S. Phenolic Acid Composition and Antioxidant Properties of Malaysian Honeys. J. Food Sci. 2011, 76, C921–C928. [Google Scholar] [CrossRef]
- Vaisman, N.; Niv, E.; Izkhakov, Y. Catalytic amounts of fructose may improve glucose tolerance in subjects with uncontrolled non-insulin-dependent diabetes. Clin. Nutr. 2006, 25, 617–621. [Google Scholar] [CrossRef]
- Thibault, L. Dietary Carbohydrates: Effects on Self-selection, Plasma Glucose and Insulin, and Brain Indoleaminergic Systems in Rat. Appetite 1994, 23, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Erejuwa, O.O.; Sulaiman, S.A.; Ab Wahab, M.S.; Sirajudeen, K.N.S.; Salleh, M.S.M.; Gurtu, S. Glibenclamide or metformin combined with honey improves glycemic control in streptozotocin-induced dia-betic rats. Int. J. Biol. Sci. 2011, 7, 244. [Google Scholar] [CrossRef] [Green Version]
- Samat, S.; Nor, N.A.M.; Hussein, F.N.; Ismail, W.I.W. Effects of Gelam and Acacia honey acute administration on some biochemical parameters of Sprague Dawley rats. BMC Complement. Altern. Med. 2014, 14, 146. [Google Scholar] [CrossRef] [Green Version]
- Robertson, R.P.; Harmon, J.; Tran, P.O.; Tanaka, Y.; Takahashi, H. Glucose toxicity in β-cells: Type 2 diabetes, good radicals gone bad, and the glutathione connection. Diabetes 2003, 52, 581–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erejuwa, O.O.; Sulaiman, S.A.; Wahab, M.S.; Sirajudeen, K.N.S.; Salleh, M.S.; Gurtu, S. Hepatoprotective effect of tualang honey supplementation in streptozotocin-induced diabetic rats. Int. J. Appl. Res. Nat. Prod. 2012, 4, 37–41. [Google Scholar]
- Mogensen, C.E.; Steffes, M.W.; Deckert, T.; Christiansen, J.S. Functional and morphological renal manifestations in diabetes mellitus. Diabetologia 1981, 21, 89–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imtara, H.; Al-Waili, N.; Bakour, M.; Al-Waili, W.; Lyoussi, B. Evaluation of antioxidant, diuretic, and wound healing effect of Tulkarm honey and its effect on kidney function in rats. Veter-World 2018, 11, 1491–1499. [Google Scholar] [CrossRef] [Green Version]
- Bhalchandra, W.; Alqadhi, Y.A. Administration of Honey and Royal Jelly Ameliorate Cisplatin Induced Changes in Liver and Kidney Function in Rat. Biomed. Pharmacol. J. 2018, 11, 2191–2199. [Google Scholar]
- Simmons, R.A. Developmental origins of diabetes: The role of oxidative stress. Free Radic. Biol. Med. 2006, 40, 917–922. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, T.; Imai, H. Hydrogen peroxide produced by superoxide dismutase SOD-2 activates sperm in Caenorhabditis elegans. J. Biol. Chem. 2017, 292, 14804–14813. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, J.H.M.; Talyuli, O.A.C.; Goncalves, R.L.S.; Paiva-Silva, G.O.; Sorgine, M.H.F.; Alvarenga, P.H.; Oliveira, P. Catalase protects Aedes aegypti from oxidative stress and increases midgut infection prevalence of Dengue but not Zika. PLoS Neglected Trop. Dis. 2017, 11, e0005525. [Google Scholar] [CrossRef] [Green Version]
- Omotayo, E.O.; Gurtu, S.; Sulaiman, S.A.; Ab Wahab, M.S.; Sirajudeen, K.N.S.; Salleh, S.M. Hypoglycemic and Antioxidant Effects of Honey Supplementation in Streptozotocin-induced Diabetic Rats. Int. J. Vitam. Nutr. Res. 2010, 80, 74–82. [Google Scholar] [CrossRef]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Cuadrado, A. Transcription Factor Nrf2: A novel target to modulate inflammatory and neuroprotective responses in Parkinson’s disease. SpringerPlus 2015, 4, 1–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aly, M.S.; Galaly, S.R.; Moustafa, N.; Mohammed, H.M.; Khadrawy, S.M.; Mahmoud, A.M. Hesperidin protects against diethylnitrosamine/carbon tetrachloride-induced renal repercussions via up-regulation of Nrf2/HO-1 signaling and attenuation of oxidative stress. J. Appl. Pharm. Sci. 2017, 7, 7–14. [Google Scholar]
- Mahmoud, A.M.; Hozayen, W.G.; Ramadan, S.M. Berberine ameliorates methotrexate-induced liver injury by activating Nrf2/HO-1 pathway and PPARγ, and suppressing oxidative stress and apoptosis in rats. Biomed. Pharmacother. 2017, 94, 280–291. [Google Scholar] [CrossRef]
- Kamel, E.M.; Mahmoud, A.M.; Ahmed, S.A.; Lamsabhi, A.M. A phytochemical and computational study on flavonoids isolated from Trifolium resupinatum L. and their novel hepatoprotective activity. Food Funct. 2016, 7, 2094–2106. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Wilkinson, F.L.; McCarthy, E.M.; Moreno-Martinez, D.M.; Langford-Smith, A.; Romero, M.; Duarte, J.; Alexander, L.-S. Endothelial microparticles prevent lipid-induced endothelial damage via Akt/eNOS signaling and reduced oxidative stress. FASEB J. 2017, 31, 4636–4648. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, A.M.; Germoush, M.O.; Al-Anazi, K.M.; Mahmoud, A.H.; Farah, M.A.; Allam, A.A. Commiphora molmol protects against methotrexate-induced nephrotoxicity by up-regulating Nrf2/ARE/HO-1 signaling. Biomed. Pharmacother. 2018, 106, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Erejuwa, O.O.; Sulaiman, S.A.; Ab Wahab, M.S.; Sirajudeen, K.N.S.; Salleh, S.M.; Gurtu, S. Honey Supplementation in Spontaneously Hypertensive Rats Elicits Antihypertensive Effect via Amelioration of Renal Oxidative Stress. Oxidative Med. Cell. Longev. 2012, 2012, 374037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranneh, Y.; Akim, A.M.; Ab Hamid, H.; Khazaai, H.; Fadel, A.; Mahmoud, A.M. Stingless bee honey protects against lipopolysaccharide induced-chronic subclinical systemic inflammation and oxidative stress by modulating Nrf2, NF-κB and p38 MAPK. Nutr. Metab. 2019, 16, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Kim, S.; Chung, H.-T.; Pae, H.-O. Reactive Oxygen Species in the Activation of MAP Kinases. Methods Enzymol. 2013, 528, 27–48. [Google Scholar] [PubMed]
- Bachstetter, D.A.; van Eldik, L.J. The p38 MAP kinase family as regulators of proinflammatory cytokine production in de-generative diseases of the CNS. Aging Dis. 2010, 1, 199. [Google Scholar]
- Kassim, M.; Yusoff, K.M.; Ong, G.; Sekaran, S.; Yusof, M.Y.B.M.; Mansor, M. Gelam honey inhibits lipopolysaccharide-induced endotoxemia in rats through the induction of heme oxygenase-1 and the inhibition of cytokines, nitric oxide, and high-mobility group protein B1. Fitoterapia 2012, 83, 1054–1059. [Google Scholar] [CrossRef] [PubMed]
- Búfalo, M.C.; Ferreira, I.; Costa, G.; Francisco, V.; Liberal, J.; Cruz, M.T.; Lopes, M.C.; Batista, M.T.; Sforcin, J.M. Propolis and its constituent caffeic acid suppress LPS-stimulated pro-inflammatory response by blocking NF-κB and MAPK activation in macrophages. J. Ethnopharmacol. 2013, 149, 84–92. [Google Scholar] [CrossRef]
- Hadagali, M.D.; Chua, L.S. The anti-inflammatory and wound healing properties of honey. Eur. Food Res. Technol. 2014, 239, 1003–1014. [Google Scholar] [CrossRef]
- Ghazali, W.S.W.; Romli, A.C.; Mohamed, M. Effects of honey supplementation on inflammatory markers among chronic smokers: A randomized controlled trial. BMC Complement. Altern. Med. 2017, 17, 175. [Google Scholar] [CrossRef] [PubMed]
- Tahir, A.A.; Sani, N.F.A.; Murad, N.A.; Makpol, S.; Ngah, W.Z.W.; Yusof, Y.A.M. Combined ginger extract & Gelam honey modulate Ras/ERK and PI3K/AKT pathway genes in colon cancer HT29 cells. Nutr. J. 2015, 14, 31. [Google Scholar] [PubMed] [Green Version]
- Anarkooli, J.I.; Ganji, H.B.; Pourheidar, M. The Protective Effects of Insulin and Natural Honey against Hip-pocampal Cell Death in Streptozotocin-Induced Diabetic Rats. J. Diabetes Res. 2014, 2014, 491571. [Google Scholar]
- Sairazi, N.S.M.; Sirajudeen, K.N.S.; Mustapha, M.; Mummedy, S.; Asari, M.A.; Sulaiman, S.A. Tualang Honey Reduced Neuroinflammation and Caspase-3 Activity in Rat Brain after Kainic Acid-Induced Status Epilepticus. Evid.-Based Complement. Altern. Med. 2018, 2018, 7287820. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No | Component Name | Formula | Observed m/z | Observed RT (min) |
|---|---|---|---|---|
| 1 | Shancilin | C30H28O6 | 483.1835 | 0.4 |
| 2 | 5-Hydroxymethyl furoic acid | C6H6O4 | 141.0194 | 0.46 |
| 3 | Isomaltose | C12H22O11 | 341.1093 | 0.49 |
| 4 | Sodium ferulate | C10H9NaO4 | 215.0318 | 0.5 |
| 5 | 6″-O-p-Hydroxybenzoyliridin | C31H30O15 | 641.1532 | 0.52 |
| 6 | Raffinose | C18H32O16 | 503.1622 | 0.53 |
| 7 | 2-Hydroxy-succinic acid | C4H6O5 | 133.0145 | 0.55 |
| 8 | 5R-5-Hydroxymethyl-2(5H)-furanone | C5H6O3 | 113.0247 | 0.6 |
| 9 | Fumaric acid | C4H4O4 | 115.0039 | 0.61 |
| 10 | 5,3′,5′-Trihydroxy-6,7,4′-trimethoxy flavone | C18H16O8 | 359.0756 | 0.64 |
| 11 | Iridin | C24H26O13 | 521.1293 | 0.64 |
| 12 | Stachyose | C24H42O21 | 665.2146 | 0.66 |
| 13 | Vanillic acid β-D-glucopyranosyl ester | C14H18O9 | 329.0886 | 0.72 |
| 14 | Succinic acid | C4H6O4 | 117.0196 | 1.05 |
| 15 | Asperuloside | C18H22O11 | 413.1093 | 1.52 |
| 16 | Courmaric acid | C9H8O2 | 147.0453 | 2.4 |
| 17 | Lamiophlomiol A | C11H14O6 | 241.0719 | 3.26 |
| 18 | Geniposidic acid | C16H22O10 | 373.1141 | 3.65 |
| 19 | Leucocyanidin | C15H14O7 | 305.0666 | 7.62 |
| 20 | Gallocatechin | C15H14O7 | 305.0666 | 7.62 |
| Compound | Binding Energies | |
|---|---|---|
| Amylase | PPAR-Gama | |
| Shancilin | −8.3 | −9.7 |
| Iridin | −7.3 | −5.5 |
| Stachyose | −6.5 | −6.5 |
| Asperuloside | −6.7 | −7.4 |
| Gallocatechin | −6.8 | −6.3 |
| Lamiophlomiol_A | −6.9 | −7.5 |
| Geniposidic_acid | −6.1 | −6.2 |
| 5,3′,5′-Trihydroxy-6,7,4′-trimethoxy flavones | −6.5 | −6.3 |
| Leucocyanidin | −6.5 | −6.4 |
| Vanillic_acid_β_D_glucopyranosyl_ester | −6.2 | −6.8 |
| Coumaric_acid | −6.7 | −7.4 |
| Raffinose | −6.7 | −6.2 |
| Isomaltose | −6.4 | −5.9 |
| Sodium_ferulate | −5.8 | −5.9 |
| Hydroxymethylfuroicacid | −5.2 | −5.4 |
| 2_Hydroxy_succinicacid | −4.5 | −4.9 |
| 5_Hydroxymethyl_furanone | −4.7 | −4.7 |
| Succinic_acid | −4.3 | −4.5 |
| Fumaric_acid | −4.7 | −4.7 |
| Parameters | Normal Rats | Diabetic Control | Diabetic Rats | ||||
|---|---|---|---|---|---|---|---|
| Control | 1 g/kg/ Day BGH | 2 g/kg/ Day BGH | 1 g/kg/ Day BGH | 2 g/kg/ Day BGH | 2 mg/kg/ Day Glibenclamide | ||
| Lipid peroxide levels (μ moles of MDA formed/gram wet weight of tissues) | 6.27 ± 0.48 | 5.48 ± 0.59 | 6.48 ± 0.69 | 17.38 ± 0.87 * | 13.84 ± 0.82 # | 12.17 ± 0.97 ## | 10.14 ± 0.88 ## |
| Superoxide dismutase activity (Units/mg protein/min) | 3.56 ± 0.09 | 2.85 ± 0.07 | 3.25 ± 0.07 | 0.86 ± 0.08 * | 1.26 ± 0.08 # | 1.84 ± 0.09 ## | 2.06 ± 0.09 ## |
| Catalyse activity (μ moles of H2O2metabolized/mg protein/min) | 2.56 ± 0.06 | 3.04 ± 0.06 | 2.89 ± 0.09 | 0.84 ± 0.05 * | 1.20 ± 0.07 # | 1.44 ± 0.05 ## | 1.65 ± 0.06 ## |
| Glutathione peroxidase activity (μ moles of GSH consume/mg protein/min) | 2.49 ± 0.08 | 2.32 ± 0.06 | 2.42 ± 0.06 | 0.74 ± 0.07 * | 1.28 ± 0.06 # | 1.42 ± 0.07 ## | 1.62 ± 0.09 ## |
| Parameters | Normal Rats | Diabetic | Diabetic Rats | ||||
|---|---|---|---|---|---|---|---|
| 1 g/kg/ Day BGH | 2 g/kg/ Day BGH | 1 g/kg/ Day BGH | 2 g/kg /Day BGH | 2 mg/kg/ Day Glibenclamide | |||
| Lipid peroxide levels (μ moles of MDA formed/gram wet weight of tissues) | 6.97 ± 0.72 | 6.28 ± 0.87 | 5.87 ± 0.68 | 19.48 ± 0.57 * | 15.84 ± 0.89 # | 12.47 ± 0.97 ## | 11.42 ± 0.78 ## |
| Superoxide dismutase activity (units/mg protein/min) | 3.56 ± 0.08 | 3.65 ± 0.04 | 3.75 ± 0.09 | 0.87 ± 0.02 * | 1.32 ± 0.12 # | 1.74±0.09 ## | 1.96 ± 0.13 ## |
| Catalyse activity (μ moles of H2O2 metabolized/mg protein/min) | 2.16 ± 0.07 | 2.95 ± 0.06 | 2.49 ± 0.08 | 0.84 ± 0.02 * | 1.08 ± 0.07 # | 1.34±0.14 ## | 1.65 ± 0.07## |
| Glutathione peroxidase activity (μ moles of GSH consume/mg protein/min) | 2.19 ± 0.11 | 1.82 ± 0.08 | 1.97 ± 0.02 | 0.55 ± 0.04 * | 0.87 ± 0.09 # | 1.45±0.09 ## | 1.62 ± 0.08 ## |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arigela, C.S.; Nelli, G.; Gan, S.H.; Sirajudeen, K.N.S.; Krishnan, K.; Abdul Rahman, N.; Pasupuleti, V.R. Bitter Gourd Honey Ameliorates Hepatic and Renal Diabetic Complications on Type 2 Diabetes Rat Models by Antioxidant, Anti-Inflammatory, and Anti-Apoptotic Mechanisms. Foods 2021, 10, 2872. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10112872
Arigela CS, Nelli G, Gan SH, Sirajudeen KNS, Krishnan K, Abdul Rahman N, Pasupuleti VR. Bitter Gourd Honey Ameliorates Hepatic and Renal Diabetic Complications on Type 2 Diabetes Rat Models by Antioxidant, Anti-Inflammatory, and Anti-Apoptotic Mechanisms. Foods. 2021; 10(11):2872. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10112872
Chicago/Turabian StyleArigela, Chandra Sekhar, Giribabu Nelli, Siew Hua Gan, Kuttulebbai Nainamohamed Salam Sirajudeen, Kumarathevan Krishnan, Nurhanan Abdul Rahman, and Visweswara Rao Pasupuleti. 2021. "Bitter Gourd Honey Ameliorates Hepatic and Renal Diabetic Complications on Type 2 Diabetes Rat Models by Antioxidant, Anti-Inflammatory, and Anti-Apoptotic Mechanisms" Foods 10, no. 11: 2872. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10112872