
Diverse Krill Lipid Fractions Differentially Reduce LPS-Induced Inflammatory Markers in RAW264.7 Macrophages In Vitro


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Preparation and Analysis of Three Krill Lipid Fractions with Different Compositions
2.3. Cell Culture
2.4. Cell Viability Assay
2.5. Determination of NO Production
2.6. Cytokine Determinations
2.7. Quantitative Real-Time Fluorescent PCR (RT-PCR) Analysis
2.7.1. General
2.7.2. RNA Extraction and cDNA Synthesis
2.7.3. Quantitative RT-PCR
2.8. Statistics
3. Results and Discussion
3.1. Compositions of Three Krill Lipid Fractions
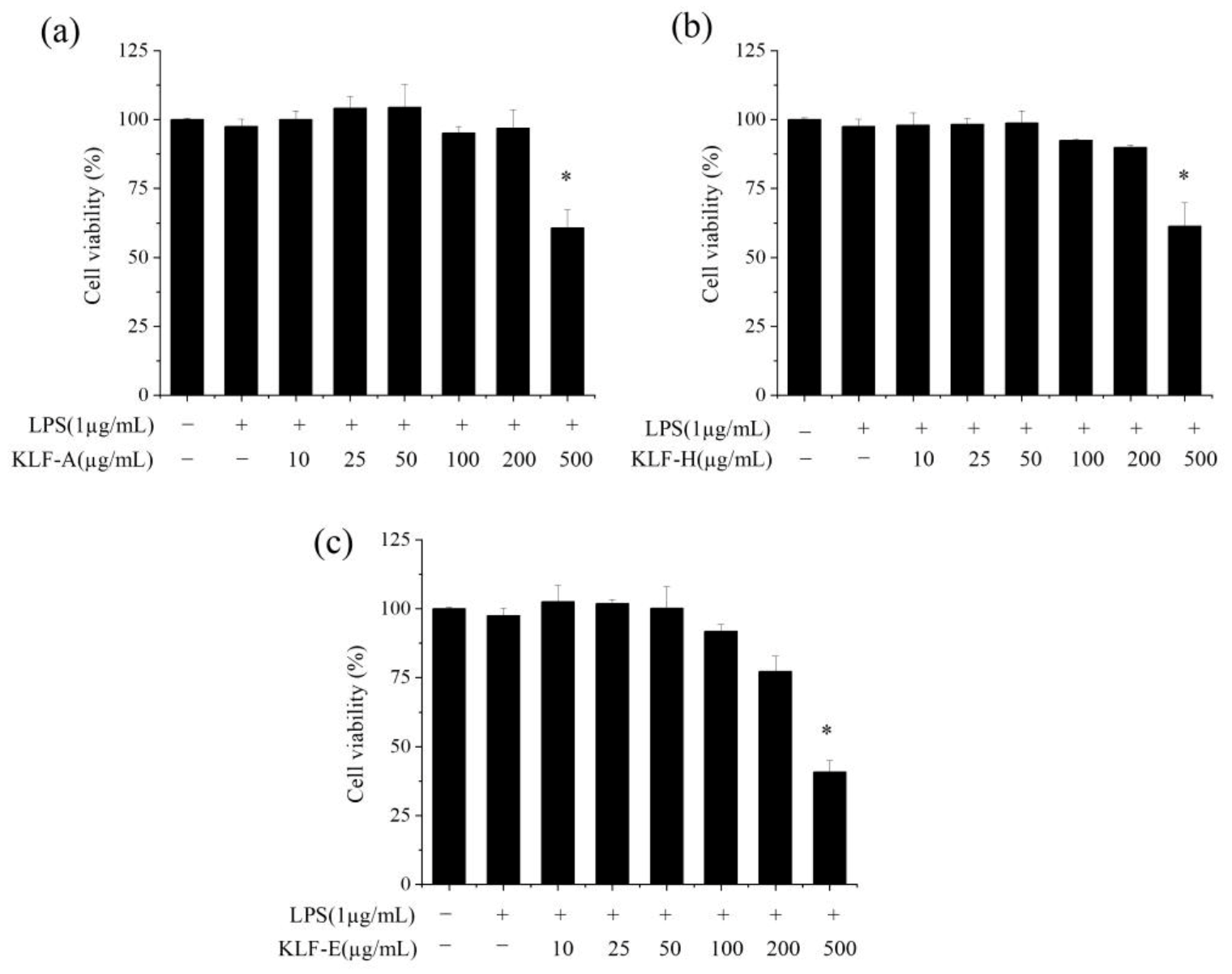
3.2. Effect of Three Krill Lipid Fractions on the Viability of RAW264.7 Cells with LPS Stimulation
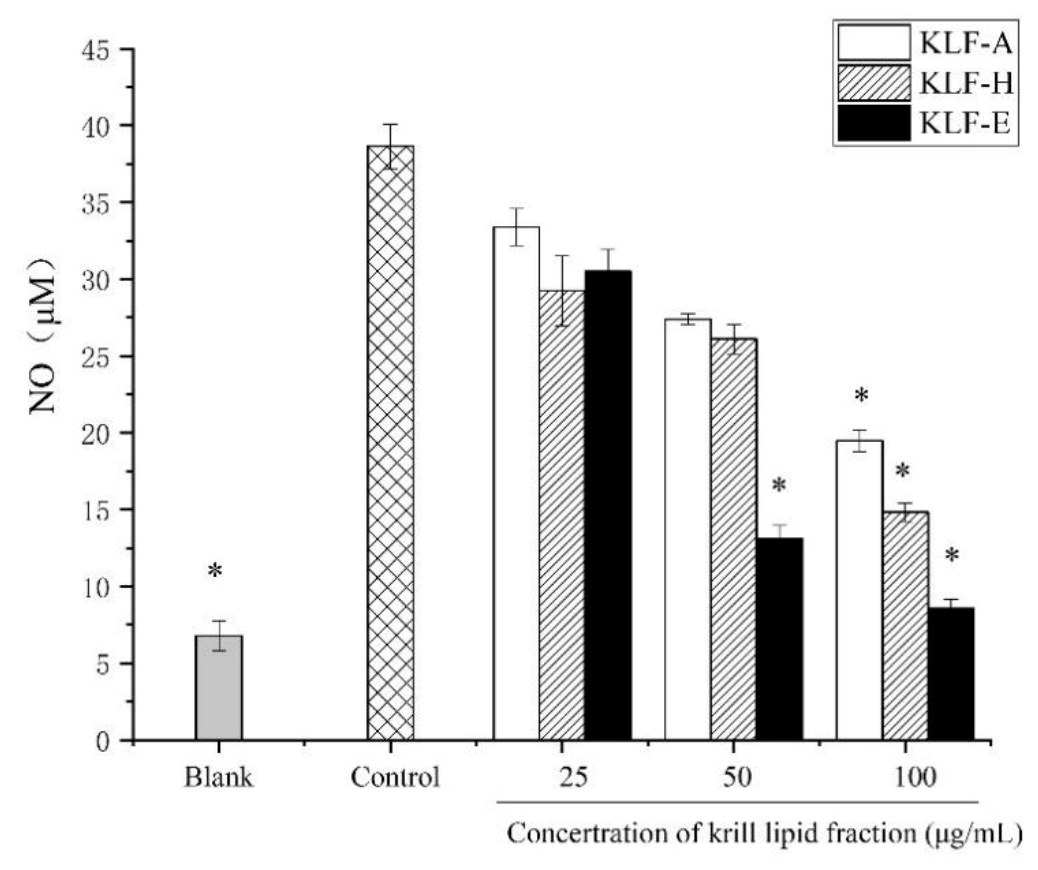
3.3. Effect of Three Krill Lipid Fractions on NO Production in RAW264.7 Cells with LPS Stimulation
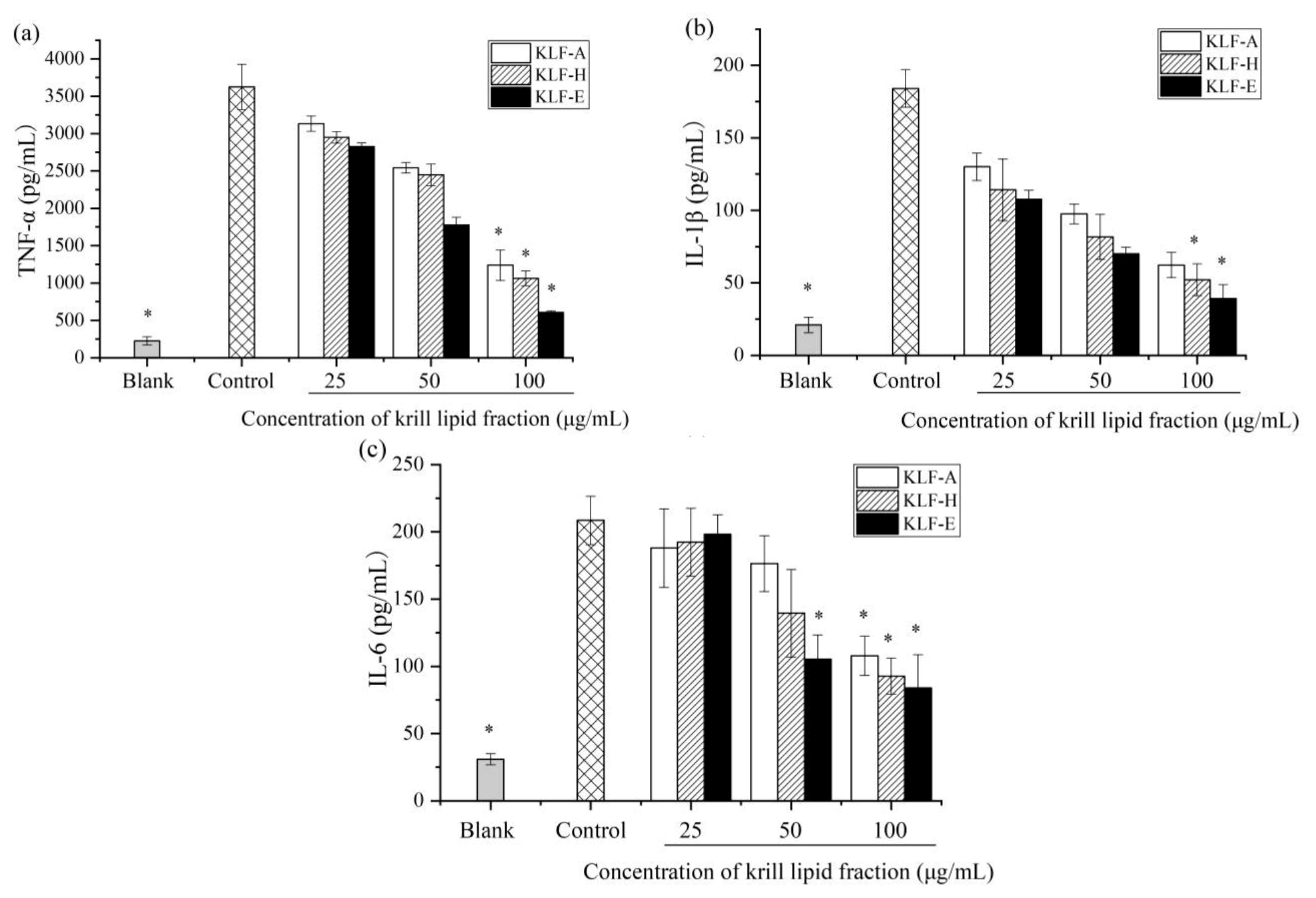
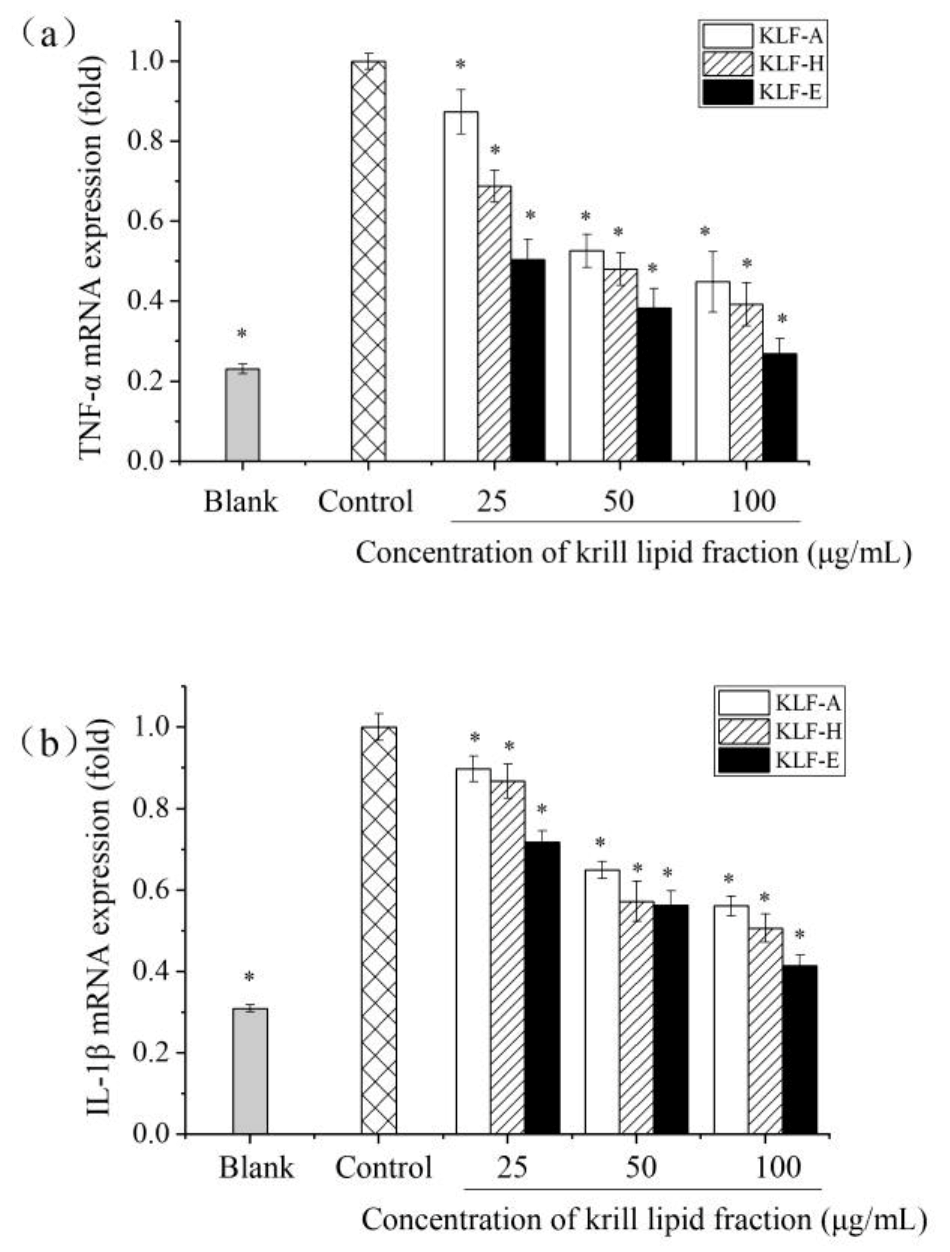
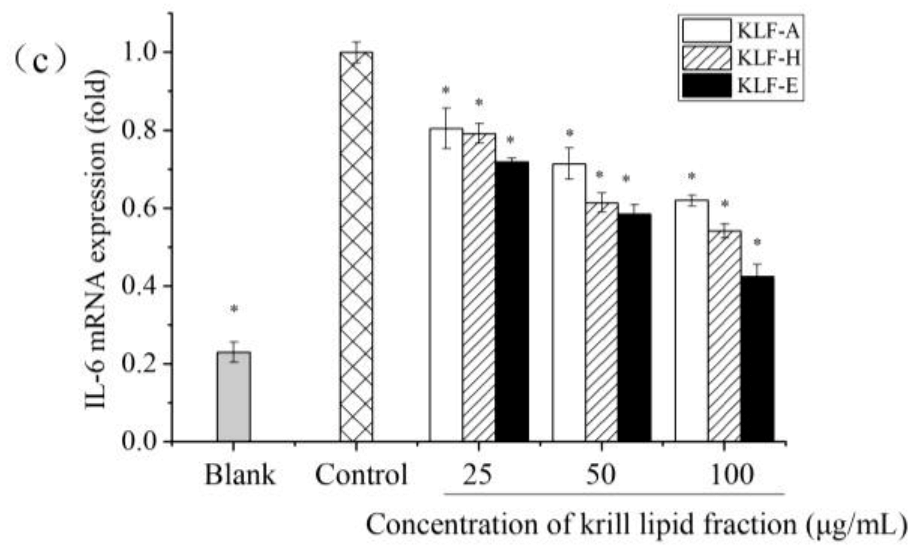
3.4. Effect of Three Krill Lipid Fractions on Cytokine Release and Related Gene Expression in RAW264.7 Cells with LPS Stimulation
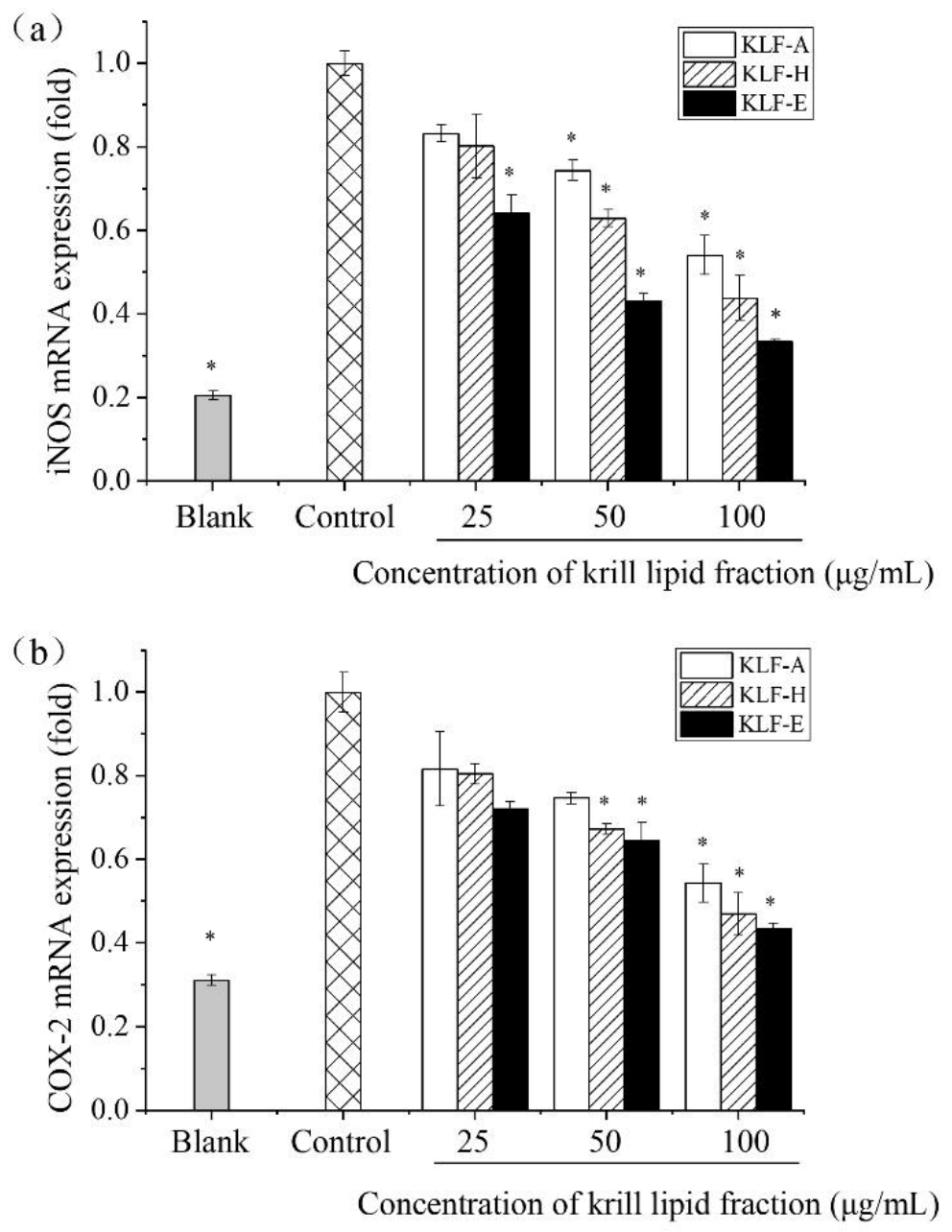
3.5. Effect of Three Krill Lipid Fractions on Genes Expression of iNOS and COX-2 in RAW264.7 Cells
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Barnes, P.J.; Karin, M. Nuclear factor-κB—A pivotal transcription factor in chronic inflammatory diseases. N. Engl. J. Med. 1997, 336, 1066–1071. [Google Scholar] [CrossRef]
- Monteiro, R.; Azevedo, I. Chronic inflammation in obesity and the metabolic syndrome. Mediat. Inflamm. 2010, 289645. [Google Scholar] [CrossRef]
- Nathan, C. Points of control in inflammation. Nature 2002, 420, 846–852. [Google Scholar] [CrossRef]
- Gautam, R.; Jachak, S.M. Recent developments in anti-inflammatory natural products. Med. Res. Rev. 2009, 29, 767–820. [Google Scholar] [CrossRef]
- Haddad, P.S.; Azar, G.A.; Groom, S.; Boivin, M. Natural health products, modulation of immune function and prevention of chronic diseases. Evid. Based Complement Alternat. Med. 2005, 2, 513–520. [Google Scholar] [CrossRef]
- Xie, D.; Gong, M.; Wei, W.; Jin, J.; Wang, X.; Wang, X.; Jin, Q. Antarctic krill (Euphausia superba) oil: A comprehensive review of chemical composition, extraction technologies, health benefits, and current applications. Compr. Rev. Food Sci. F 2019, 18, 514–534. [Google Scholar] [CrossRef] [Green Version]
- Deutsch, L. Evaluation of the effect of neptune krill oil on chronic inflammation and arthritic symptoms. J. Am. Coll. Nutr. 2007, 26, 39–48. [Google Scholar] [CrossRef]
- Batetta, B.; Griinari, M.; Carta, G.; Murru, E.; Ligresti, A.; Cordeddu, L.; Giordano, E.; Sanna, F.; Bisogno, T.; Uda, S.; et al. Endocannabinoids may mediate the ability of (n-3) fatty acids to reduce ectopic fat and inflammatory mediators in obese zucker rats. J. Nutr. 2009, 139, 1495–1501. [Google Scholar] [CrossRef] [Green Version]
- Bonaterra, A.G.; Driscoll, D.; Schwarzbach, H.; Kinscherf, R. Krill oil-in-water emulsion protects against lipopolysaccharide-induced proinflammatory activation of macrophages in vitro. Mar. Drugs 2017, 15, 74. [Google Scholar] [CrossRef] [Green Version]
- Costanzo, M.; Cesi, V.; Prete, E.; Negroni, A.; Palone, F.; Cucchiara, S.; Oliva, S.; Leter, B.; Stronati, L. Krill oil reduces intestinal inflammation by improving epithelial integrity and impairing adherent-invasive Escherichia coli pathogenicity. Diges. Liver Dis. 2016, 48, 34–42. [Google Scholar] [CrossRef]
- Ierna, M.; Kerr, A.; Scales, H.; Berge, K.; Griinari, M. Supplementation of diet with krill oil protects against experimental rheumatoid arthritis. BMC Musculoskel Dis. 2010, 11, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimstad, T.; Bjørndal, B.; Cacabelos, D.; Aasprong, O.G.; Janssen, E.A.M.; Omdal, R.; Svardal, A.; Hausken, T.; Bohov, P.; Portero-Otin, M.; et al. Dietary supplementation of krill oil attenuates inflammation and oxidative stress in experimental ulcerative colitis in rats. Scand J. Gastroenterolo. 2012, 47, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.; Rosticci, M.; Morbini, M.; Cagnati, M.; Grandi, E.; Parini, A.; Borghi, C. Lipid-lowering and anti-inflammatory effects of omega 3 ethyl esters and krill oil: A randomized, cross-over, clinical trial. Arch. Med. Sci. 2016, 12, 507–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skarpańska-Stejnborn, A.; Pilaczyńska-Szcześniak, Ł.; Basta, P.; Foriasz, J.; Arlet, J. Effects of supplementation with neptune krill oil (euphasia superba) on selected redox parameters and pro-Inflammatory markers in athletes during exhaustive exercise. J. Hum. Kinet. 2010, 25, 49. [Google Scholar] [CrossRef]
- Cabre, E.; Manosa, M.; Gassull, M.A. Omega-3 fatty acids and inflammatory bowel diseases—A systematic review. Br. J. Nutr. 2012, 107 (Suppl. 2), S240–S252. [Google Scholar] [CrossRef] [Green Version]
- Chapkin, R.S.; Kim, W.; Lupton, J.R.; McMurray, D.N. Dietary docosahexaenoic and eicosapentaenoic acid: Emerging mediators of inflammation. Prostag. Leukot. Ess. 2009, 81, 187–191. [Google Scholar] [CrossRef] [Green Version]
- Lordan, R.; Redfern, S.; Tsoupras, A.; Zabetakis, I. Inflammation and cardiovascular disease: Are marine phospholipids the answer? Food Func. 2020, 11, 2861–2885. [Google Scholar] [CrossRef]
- Ambati, R.R.; Phang, S.M.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications—A review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef]
- Grammas, P.; Hamdheydari, L.; Benaksas, E.J.; Mou, S.; Pye, Q.N.; Wechter, W.J.; Floyd, R.A.; Stewart, C.; Hensley, K. Anti-inflammatory effects of tocopherol metabolites. Biochem. Biophys. Res. Commun. 2004, 319, 1047–1052. [Google Scholar] [CrossRef]
- Kimble, L.; Mathison, B.; Chew, B.P. Astaxanthin mediates inflammatory biomarkers associated with arthritis in human chondrosarcoma cells induced with interleukin. FASEB J. 2013, 27, 37–51. [Google Scholar] [CrossRef]
- Akeson, A.; Woods, C.; Mosher, L.; Thomas, C.; Jackson, R. Inhibition of IL-1β expression in THP-1 cells by probucol and tocopherol. Atherosclerosis 1991, 86, 261–270. [Google Scholar] [CrossRef]
- Fox, E.S.; Brower, J.S.; Bellezzo, J.M.; Leingang, K.A. N-Acetylcysteine and α-tocopherol reverse the inflammatory response in activated rat Kupffer cells. J. Immunol. 1997, 158, 5418–5423. [Google Scholar]
- Wu, S.J.; Liu, P.L.; Ng, L.T. Tocotrienol-rich fraction of palm oil exhibits anti-inflammatory property by suppressing the expression of inflammatory mediators in human monocytic cells. Mol. Nutr. Food Res. 2008, 52, 921–929. [Google Scholar] [CrossRef]
- Sun, D.; Cao, C.; Li, B.; Chen, H.; Li, J.; Cao, P.; Liu, Y. Antarctic krill lipid extracted by subcritical n-butane and comparison with supercritical CO2 and conventional solvent extraction. LWT-Food Sci. Technol. 2018, 94, 1–7. [Google Scholar] [CrossRef]
- Xie, D.; Jin, J.; Sun, J.; Liang, L.; Wang, X.; Zhang, W.; Wang, X.; Jin, Q. Comparison of solvents for extraction of krill oil from krill meal: Lipid yield, phospholipids content, fatty acids composition and minor components. Food Chem. 2017, 33, 434–441. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Murakami, M.; Nakano, H.; Konosu, S.; Kokura, T.; Yamamoto, H.; Kosaka, M.; Hata, K. Supercritical carbon dioxide extraction of oils from Antarctic krill. J. Agric. Food Chem. 1986, 34, 904–907. [Google Scholar] [CrossRef]
- Xie, D.; Mu, H.; Tang, T.; Wang, X.; Wei, W.; Jin, J.; Wang, X.; Jin, Q. Production of three types of krill oils from krill meal by a three-step solvent extraction procedure. Food Chem. 2018, 248, 279–286. [Google Scholar] [CrossRef]
- Li, D.M.; Zhou, D.Y.; Zhu, B.W.; Chi, Y.L.; Sun, L.M.; Dong, X.P.; Qin, L.; Qiao, W.Z.; Murata, Y. Effects of krill oil intake on plasma cholesterol and glucose levels in rats fed a high-cholesterol diet. J. Sci. Food Agric. 2013, 93, 2669–2675. [Google Scholar] [CrossRef]
- Jung, H.A.; Jin, S.E.; Ahn, B.R.; Lee, C.M.; Choi, J.S. Anti-inflammatory activity of edible brown alga eisenia bicyclis and its constituents fucosterol and phlorotannins in LPS-stimulated RAW264.7 macrophages. Food Chem. Toxicol. 2013, 59, 199–206. [Google Scholar] [CrossRef]
- Moro, C.; Palacios, I.; Lozano, M.; D’Arrigo, M.; Guillamón, E.; Villares, A.; Martínez, J.A.; García-Lafuente, A. Anti-inflammatory activity of methanolic extracts from edible mushrooms in LPS activated RAW 264.7 macrophages. Food Chem. 2012, 130, 350–355. [Google Scholar] [CrossRef]
- Fricke, H.; Gercken, G.; Schreiber, W.; Oehlenschläger, J. Lipid, sterol and fatty acid composition of antarctic krill (Euphausia superba Dana). Lipids 1984, 19, 821–827. [Google Scholar] [CrossRef]
- Kim, H.D.; Lee, S.B.; Ko, S.C.; Jung, W.K.; Kim, Y.M.; Kim, S.B. Anti-inflammatory effect of ozonated krill (Euphausia superba) oil in lipopolysaccharide-stimulated RAW 264.7 macrophages. Fish. Aquat. Sci. 2018, 21, 15. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Frede, K.; Henze, A.; Khalil, M.; Baldermann, S.; Schweigert, F.J.; Rawel, H. Stability and cellular uptake of lutein-loaded emulsions. J. Funct. Foods 2014, 8, 118–127. [Google Scholar] [CrossRef]
- Sessa, M.; Balestrieri, M.L.; Ferrari, G.; Servillo, L.; Castaldo, D.; D’Onofrio, N.; Donsì, F.; Tsao, R. Bioavailability of encapsulated resveratrol into nanoemulsion-based delivery systems. Food Chem. 2014, 147, 42–50. [Google Scholar] [CrossRef]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2008, 15, 252–259. [Google Scholar] [CrossRef]
- Bian, K.; Ke, Y.; Kamisaki, Y.; Murad, F. Proteomic modification by nitric oxide. J. Pharmacol. Sci. 2006, 101, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Guzik, T.J.; Korbut, R.; Adamek-Guzik, T. Nitric oxide and superoxide in inflammation and immune regulation. J. Physiol. Pharmacol. 2003, 54, 469–487. [Google Scholar] [PubMed]
- Lawrence, T.; Willoughby, D.A.; Gilroy, D.W. Anti-inflammatory lipid mediators and insights into the resolution of inflammation. Nat. Rev. Immunol. 2002, 2, 787–795. [Google Scholar] [CrossRef]
- Tayal, V.; Kalra, B.S. Cytokines and anti-cytokines as therapeutics—An update. Eur. J. Pharmacol. 2008, 579, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.C.; Jeon, Y.J. Anti-inflammatory effect of enzymatic hydrolysates from Styela clava flesh tissue in lipopolysaccharide-stimulated RAW 264.7 macrophages and in vivo zebrafish model. Nutr. Res. Prac. 2015, 9, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feghali, C.A.; Wright, T.M. Cytokines in acute a- chronic inflammation. Front. Biosci. 1997, 2, d12–d26. [Google Scholar] [PubMed] [Green Version]
- Jaffer, U.; Wade, R.G.; Gourlay, T. Cytokines in the systemic inflammatory response syndrome: A review. HSR Proc. Intensive Care Cardiovasc. Anesth. 2010, 2, 161–175. [Google Scholar] [PubMed]
- McCarty, M.F. Interleukin-6 as a central mediator of cardiovascular risk associated with chronic inflammation, smoking, diabetes, and visceral obesity: Down-regulation with essential fatty acids, ethanol and pentoxifylline. Med. Hypotheses 1999, 52, 465–477. [Google Scholar] [CrossRef]
- Costanzo, M.; Cesi, V.; Palone, F.; Pierdomenico, M.; Colantoni, E.; Leter, B.; Vitali, R.; Negroni, A.; Cucchiara, S.; Stronati, L. Krill oil, vitamin D and Lactobacillus reuteri cooperate to reduce gut inflammation. Benef. Microbes 2018, 9, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Park, S.J.; Yun, K.J.; Cho, Y.W.; Park, H.J.; Lee, K.T. Isoliquiritigenin isolated from the roots of Glycyrrhiza uralensis inhibits LPS-induced iNOS and COX-2 expression via the attenuation of NF-kappaB in RAW 264.7 macrophages. Eur. J. Pharmacol. 2008, 584, 175–184. [Google Scholar] [CrossRef]
- Chang, M.; Qiu, F.; Lan, N.; Zhang, T.; Guo, X.; Jin, Q.; Liu, R.; Wang, X. Analysis of phytochemical composition of camellia oleifera oil and evaluation of its anti-Inflammatory effect in lipopolysaccharide-stimulated RAW 264.7 macrophages. Lipids 2020, 55, 353–363. [Google Scholar] [CrossRef]
- Ibrahim, A.; Mbodji, K.; Hassan, A. Anti-inflammatory and anti-angiogenic effect of long chain n-3 polyunsaturated fatty acids in intestinal microvascular endothelium. Clin. Nutr. 2011, 30, 678–687. [Google Scholar] [CrossRef]
- Yang, B.; Li, R.; Greenlief, C.M.; Fritsche, K.L.; Gu, Z.; Cui, J.; Lee, J.C.; Beversdorf, D.Q.; Sun, G.Y. Unveiling anti-oxidative and anti-inflammatory effects of docosahexaenoic acid and its lipid peroxidation production lipopolysaccharide-stimulated BV-2 microglial cells. J. Neuroinflamm. 2018, 9, 202. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Khan, N.A.; McMurray, D.N.; Prior, I.A.; Wang, N.; Chapkin, R.S. Regulatory activity of polyunsaturated fatty acids in T-cell signaling. Prog. Lipid Res. 2010, 49, 250–261. [Google Scholar] [CrossRef] [Green Version]
- Lapointe, L.; Harvey, S.; Aziz, H.; Jordan, R.; Hegele, A.; Lemieux, P. A single-dose, comparative bioavailability study of a formulation containing OM3 as phospholipid and free fatty acid to an ethyl ester formulation in the fasting and fed states. Clin. Ther. 2019, 41, 426–444. [Google Scholar] [CrossRef] [Green Version]
- Awada, M.; Meynier, A.; Soulage, C.O. n-3 PUFA added to high-fat diets affect differently adiposity and inflammation when carried by phospholipids or triacylglycerols in mice. Nutr. Metab. 2013, 10, 23. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Zhang, T.; Jiang, Z.; Guo, Y.; Qiu, F.; Liu, R.; Zhang, L.; Chang, M.; Liu, R.; Jin, Q.; et al. Physical properties and cellular antioxidant activity of vegetable oil emulsions with different chain lengths and saturation of triglycerides. LWT-Food Sci. Technol. 2020, 121, 108948. [Google Scholar] [CrossRef]
- Saw, C.; Huang, Y.; Kong, A.N. Synergistic anti-inflammatory effects of low doses of curcumin in combination with polyunsaturated fatty acids: Docosahexaenoic acid or eicosapentaenoic acid. Biochem. Pharmacol. 2010, 3, 421–430. [Google Scholar] [CrossRef]
- Ohgami, K.; Shiratori, K.; Kotake, S.; Nishida, T.; Mizuki, N.; Yazawa, K.; Ohno, S. Effects of astaxanthin on lipopolysaccharide-Induced inflammation in vitro and in vivo. Invest. Opth. Vis. Sci. 2003, 44, 2694. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| GAPDH | ATG TAC GTA GCC ATC CAG GC | AGG AAG GAA GGC TGG AAG AG |
| IL-1β | CTG TCC TGC GTG TTG AAA | TTC TGC TTG AGA GGT GCT GA |
| IL-6 | AGG AGA CTT GCC TGG TGA AA | CAG GGG TGG TTA TTG CAT CT |
| TNF-α | AGG CCT TGT GTT GTG TTT CCA | TGG GGG ACA GCT TCC TTC TT |
| iNOS | CAC CTT GGA GTT CAC CCA GT | ACC ACT CGT ACT TGG GAT GC |
| COX-2 | TGA AAC CCA CTC CAA ACA CA | GAG AAG GCT TCC CAG CTT TT |
| Analytical Determination | KLF-A | KLF-H | KLF-E |
|---|---|---|---|
| Lipid yield (% d.b.) | 5.23 ± 0.25 b | 5.01 ± 0.36 b | 8.97 ± 0.40 a |
| Lipid extraction efficiency (% total lipid) | 25.37 ± 1.21 b | 24.21 ± 1.72 b | 45.94 ± 2.03 a |
| Phospholipids (g/100 g) | 2.39 ± 0.11 c | 35.02 ± 2.06 b | 62.79 ± 2.45 a |
| Astaxanthin (mg/kg) | 519.80 ± 23.56 a | 30.03 ± 0.68 b | 9.50 ± 0.06 b |
| Tocopherols (mg/100 g) | 29.65 ± 0.52 a | 11.57 ± 0.45 b | 3.73 ± 0.35 c |
| EPA (mg/g) | 74.24 ± 4.31 c | 132.57 ± 8.97 b | 224.01 ± 9.97 a |
| DHA(mg/g) | 25.51 ± 3.17 c | 94.79 ± 7.24 b | 134.04 ± 7.34 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, D.; He, F.; Wang, X.; Wang, X.; Jin, Q.; Jin, J. Diverse Krill Lipid Fractions Differentially Reduce LPS-Induced Inflammatory Markers in RAW264.7 Macrophages In Vitro. Foods 2021, 10, 2887. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10112887
Xie D, He F, Wang X, Wang X, Jin Q, Jin J. Diverse Krill Lipid Fractions Differentially Reduce LPS-Induced Inflammatory Markers in RAW264.7 Macrophages In Vitro. Foods. 2021; 10(11):2887. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10112887
Chicago/Turabian StyleXie, Dan, Fangyuan He, Xiaosan Wang, Xingguo Wang, Qingzhe Jin, and Jun Jin. 2021. "Diverse Krill Lipid Fractions Differentially Reduce LPS-Induced Inflammatory Markers in RAW264.7 Macrophages In Vitro" Foods 10, no. 11: 2887. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10112887