
Quality Traits and Nutritional Value of Pork and Poultry Meat from Animals Fed with Seaweeds

 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Seaweed Production—A Brief Overview
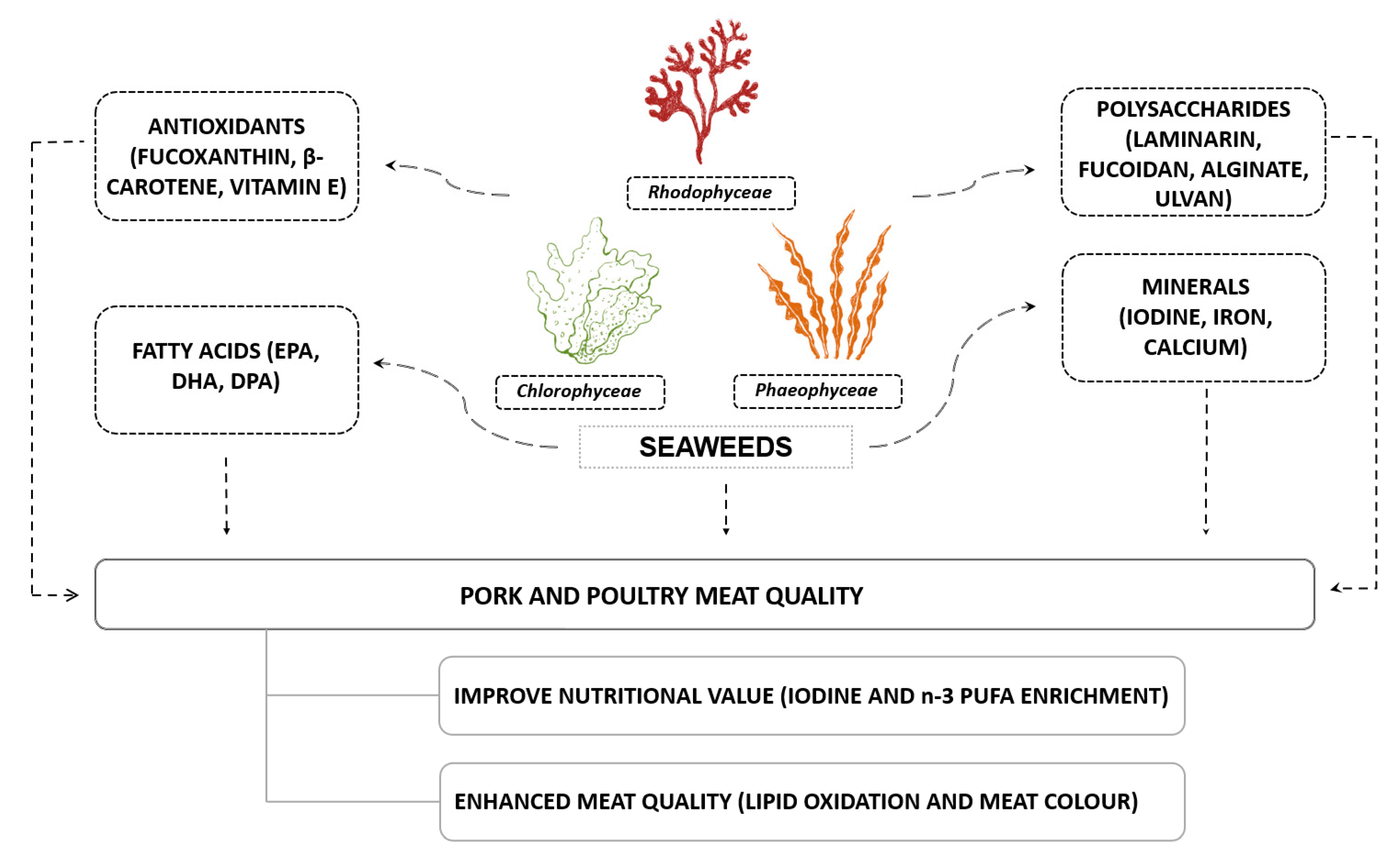
3. Nutritional Properties of Seaweeds
4. Effect of Dietary Seaweeds on Pork Meat Quality
5. Effect of Dietary Seaweeds on Poultry Meat Quality
6. Future Challenges
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Jaggard, K.W.; Qi, A.; Ober, E. Possible changes to arable crop yields by 2050. Philos. Trans. R. Soc. Biol. Sci. 2010, 365, 2835–2851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, M.; Cardoso, C.; Afonso, C.; Bandarra, N.M.; Prates, J.A.M. Current knowledge and future perspectives of the use of seaweeds for livestock production and meat quality: A systematic review. J. Anim. Physiol. Anim. Nutr. 2021, 105, 1075–1102. [Google Scholar] [CrossRef]
- The National Pork Board. Pork Checkoff. Available online: https://porkcheckoff.org/pork-branding/facts-statistics/ (accessed on 15 November 2021).
- Fehlenberg, V.; Baumann, M.; Gasparri, N.I.; Piquer-Rodríguez, M.; Gavier-Pizarro, G.; Kuemmerle, T. The role of soybean production as an underlying driver of deforestation in the South American Chaco. Glob. Environ. Chang. 2017, 45, 24–34. [Google Scholar] [CrossRef]
- Martins, C.; Cullere, M.; Zotte, A.D.; Cardoso, C.; Alves, S.P.; De Bessa, R.J.B.; Freire, J.P.B.; Falcaõ-E-Cunha, L. Incorporation of two levels of black soldier fly (Hermetia illucens L.) larvae fat or extruded linseed in diets of growing rabbits: Effects on growth performance and diet digestibility. Czech J. Anim. Sci. 2018, 63, 356–362. [Google Scholar]
- Kierończyk, B.; Rawski, M.; Jozefiak, A.; Mazurkiewicz, J.; Swiatkiewicz, S.; Siwek, M.; Bednarczyk, M.; Szumacher-Strabel, M.; Cieslak, A.; Benzertiha, A.; et al. Effects of replacing soybean oil with selected insect fats on broilers. Anim. Feed. Sci. Technol. 2018, 240, 170–183. [Google Scholar] [CrossRef]
- Vastolo, A.; Calabró, S.; Liotta, L.; Musco, N.; Di Rosa, A.R.; Cutrignelli, M.I.; Chiofalo, B. In Vitro Fermentation and Chemical Characteristics of Mediterranean By-Products for Swine Nutrition. Animals 2019, 9, 556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correia, C.S.; Alfaia, C.M.; Madeira, M.S.; Lopes, P.A.; Matos, T.J.S.; Cunha, L.F.; Prates, J.A.M.; Freire, J.P.B. Dietary inclusion of tomato pomace improves meat oxidative stability of young pigs. J. Anim. Physiol. Anim. Nutr. 2017, 101, 1215–1226. [Google Scholar] [CrossRef]
- Martins, C.F.; Pestana Assuncao, J.; Ribeiro Santos, D.M.; Madeira, M.S.M.D.S.; Alfaia, C.M.R.P.M.; Lopes, P.A.A.B.; Freire, J.P.B.; Coelho, D.F.M.; Lemos, J.P.C.; de Almeida, A.M.; et al. Effect of dietary inclusion of Spirulina on production performance, nutrient digestibility and meat quality traits in post-weaning piglets. J. Anim. Physiol. Anim. Nutr. 2021, 105, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.; Pestana, J.; Alfaia, C.; Costa, M.; Ribeiro, D.; Coelho, D.; Lopes, P.; Almeida, A.; Freire, J.; Prates, J. Effects of Chlorella vulgaris as a Feed Ingredient on the Quality and Nutritional Value of Weaned Piglets’ Meat. Foods 2021, 10, 1155. [Google Scholar] [CrossRef]
- Samarasinghe, M.; van der Heide, M.; Weisbjerg, M.; Sehested, J.; Sloth, J.; Bruhn, A.; Vestergaard, M.; Nørgaard, J.; Hernández-Castellano, L. A descriptive chemical analysis of seaweeds, Ulva sp., Saccharina latissima and Ascophyllum nodosum harvested from Danish and Icelandic waters. Anim. Feed. Sci. Technol. 2021, 278, 115005. [Google Scholar] [CrossRef]
- Samarasinghe, M.B.; Sehested, J.; Weisbjerg, M.R.; van der Heide, M.E.; Nørgaard, J.V.; Vestergaard, M.; Hernández-Castellano, L.E. Feeding milk supplemented with Ulva sp., Ascophyllum nodosum, or Saccharina latissima to preweaning dairy calves: Effects on growth, gut microbiota, gut histomorphology, and short-chain fatty acids in digesta. J. Dairy Sci. 2021, 104, 12117–12126. [Google Scholar] [CrossRef]
- Pereira, L.; Cotas, J. Historical Use of Seaweed as an Agricultural Fertilizer in the European Atlantic Area. In Seaweeds as Plant Fertilizer, Agricultural Biostimulants and Animal Fodder; CRC Press: Boca Raton, FL, USA, 2019; pp. 1–22. [Google Scholar]
- The Orkney Sheep. The Orkney Sheep Foundation. Available online: https://www.theorkneysheepfoundation.org.uk/the-sheep/ (accessed on 15 November 2021).
- Chapman, V.J.; Chapman, D.J. Seaweed as Animal Fodder, Manure and for Energy. In Seaweeds and Their Uses; Springer: Singapore, 1980; pp. 30–61. [Google Scholar]
- Makkar, H.P.; Tran, G.; Heuzé, V.; Giger-Reverdin, S.; Lessire, M.; Lebas, F.; Ankers, P. Seaweeds for livestock diets: A review. Anim. Feed Sci. Technol. 2016, 212, 1–17. [Google Scholar] [CrossRef]
- Øverland, M.; Mydland, L.T.; Skrede, A. Marine macroalgae as sources of protein and bioactive compounds in feed for monogastric animals. J. Sci. Food Agric. 2019, 99, 13–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, M.; Pio, L.; Bule, P.; Cardoso, V.; Alfaia, C.M.; Coelho, D.; Brás, J.; Fontes, C.M.G.A.; Prates, J.A.M. An individual alginate lyase is effective in the disruption of Laminaria digitata recalcitrant cell wall. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Alves, C.; Silva, J.; Pinteus, S.; Gaspar, H.; Alpoim, M.C.M.D.C.; Botana, L.M.; Pedrosa, R. From Marine Origin to Therapeutics: The Antitumor Potential of Marine Algae-Derived Compounds. Front. Pharmacol. 2018, 9, 777. [Google Scholar] [CrossRef] [Green Version]
- Kalasariya, H.S.; Yadav, V.K.; Yadav, K.K.; Tirth, V.; Algahtani, A.; Islam, S.; Gupta, N.; Jeon, B.-H. Seaweed-Based Molecules and Their Potential Biological Activities: An Eco-Sustainable Cosmetics. Molecules 2021, 26, 5313. [Google Scholar] [CrossRef]
- Corino, C.; Di Giancamillo, A.; Modina, S.; Rossi, R. Prebiotic Effects of Seaweed Polysaccharides in Pigs. Animals 2021, 11, 1573. [Google Scholar] [CrossRef]
- Karre, L.; Lopez, K.; Getty, K.J. Natural antioxidants in meat and poultry products. Meat Sci. 2013, 94, 220–227. [Google Scholar] [CrossRef]
- Madeira, M.S.; Alfaia, C.M.; Lopes, P.A.; Pestana, J.; Coelho, D.; Fontes, C.M.G.A.; Prates, J.A.M. Ameliorating Pork Marbling and Quality with Novel Feeding Approaches. In Advances in Animal Health, Medicine and Production; Springer International Publishing: New York, NY, USA, 2020; pp. 161–177. [Google Scholar]
- Nollet, L.M.; Toldra, F. (Eds.) Handbook of Muscle Foods Analysis; CRC Press: Boca Raton, FL, USA, 2008; ISBN 9780429148255. [Google Scholar]
- Nabrdalik, K.; Krzyżak, K.; Hajzler, W.; Drożdż, K.; Kwiendacz, H.; Gumprecht, J.; Lip, G. Fat, Sugar or Gut Microbiota in Reducing Cardiometabolic Risk: Does Diet Type Really Matter? Nutrients 2021, 13, 639. [Google Scholar] [CrossRef]
- Czyż, K.; Sokoła-Wysoczańska, E.; Wyrostek, A.; Cholewińska, P. An attempt to enrich pig meat with omega-3 fatty acids using linseed oil ethyl ester diet supplement. Agriculture 2021, 11, 365. [Google Scholar] [CrossRef]
- Petracci, M.; Cavani, C. Muscle Growth and Poultry Meat Quality Issues. Nutrients 2011, 4, 1–12. [Google Scholar] [CrossRef]
- Bogucka, J.; Ribeiro, D.; Costa, R.; Bednarczyk, M. Effect of synbiotic dietary supplementation on histological and histopathological parameters of Pectoralis major muscle of broiler chickens. Czech J. Anim. Sci. 2018, 63, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Alfaia, C.; Pestana, J.; Rodrigues, M.; Coelho, D.; Aires, M.; Ribeiro, D.; Major, V.; Martins, C.; Santos, H.; Lopes, P.; et al. Influence of dietary Chlorella vulgaris and carbohydrate-active enzymes on growth performance, meat quality and lipid composition of broiler chickens. Poult. Sci. 2021, 100, 926–937. [Google Scholar] [CrossRef] [PubMed]
- Pestana, J.; Puerta, B.; Santos, H.; Madeira, M.; Alfaia, C.; Lopes, P.; Pinto, R.; Lemos, J.; Fontes, C.; Lordelo, M.; et al. Impact of dietary incorporation of Spirulina (Arthrospira platensis) and exogenous enzymes on broiler performance, carcass traits, and meat quality. Poult. Sci. 2020, 99, 2519–2532. [Google Scholar] [CrossRef] [PubMed]
- Sellier, P.; Monin, G. genetics of pig meat quality: A review. J. Muscle Foods 1994, 5, 187–219. [Google Scholar] [CrossRef]
- Le Bihan-Duval, E.; Debut, M.; Berri, C.M.; Sellier, N.; Santé-Lhoutellier, V.; Jégo, Y.; Beaumont, C. Chicken meat quality: Genetic variability and relationship with growth and muscle characteristics. BMC Genet. 2008, 9, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valente, L.M.P.; Cabrita, A.R.J.; Maia, M.R.G.; Valente, I.M.; Engrola, S.; Fonseca, A.J.M.; Ribeiro, D.M.; Lordelo, M.; Martins, C.F.; Falcão-E-Cunha, L.; et al. Microalgae as feed ingredients for livestock production and aquaculture. In Microalgae-Cultivation, Recovery of Compounds and Applications; Galanakis, C.M., Ed.; Academic Press, Inc.: London, UK, 2020; pp. 239–302. [Google Scholar]
- de Tonnac, A.; Mourot, J. Effect of dietary sources of n-3 fatty acids on pig performance and technological, nutritional and sensory qualities of pork. Animal 2018, 12, 1527–1535. [Google Scholar] [CrossRef] [PubMed]
- Corino, C.; Modina, S.C.; Di Giancamillo, A.; Chiapparini, S.; Rossi, R. Seaweeds in Pig Nutrition. Animals 2019, 9, 1126. [Google Scholar] [CrossRef] [Green Version]
- Morais, T.; Inácio, A.; Coutinho, T.; Ministro, M.; Cotas, J.; Pereira, L.; Bahcevandziev, K. Seaweed Potential in the Animal Feed: A Review. J. Mar. Sci. Eng. 2020, 8, 559. [Google Scholar] [CrossRef]
- FAO. FAO Statistics. Available online: https://www.fao.org/figis/servlet/TabSelector (accessed on 15 November 2021).
- Buschmann, A.H.; Camus, C.; Infante, J.; Neori, A.; Israel, Á.; Hernández-González, M.C.; Pereda, S.V.; Gomez-Pinchetti, J.L.; Golberg, A.; Tadmor-Shalev, N.; et al. Seaweed production: Overview of the global state of exploitation, farming and emerging research activity. Eur. J. Phycol. 2017, 52, 391–406. [Google Scholar] [CrossRef]
- Holden, J.J.; Dudas, S.E.; Juanes, F. Is commercial harvesting of beach-cast seaweed ecologically sustainable? Integr. Environ. Assess. Manag. 2016, 12, 825–827. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Lazaro, C.; Sanchez-Jerez, P. Regional Integrated Multi-Trophic Aquaculture (RIMTA): Spatially separated, ecologically linked. J. Environ. Manag. 2020, 271, 110921. [Google Scholar] [CrossRef] [PubMed]
- García-Poza, S.; Leandro, A.; Cotas, C.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. The Evolution Road of Seaweed Aquaculture: Cultivation Technologies and the Industry 4.0. Int. J. Environ. Res. Public Health 2020, 17, 6528. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, M.F.; Filbee-Dexter, K.; Norderhaug, K.M.; Fredriksen, S.; Frisk, N.L.; Fagerli, C.W.; Wernberg, T. Detrital carbon production and export in high latitude kelp forests. Oecologia 2020, 192, 227–239. [Google Scholar] [CrossRef]
- Guerra-García, J.M.; Hachero-Cruzado, I.; González-Romero, P.; Jiménez-Prada, P.; Cassell, C.; Ros, M. Towards Integrated Multi-Trophic Aquaculture: Lessons from Caprellids (Crustacea: Amphipoda). PLoS ONE 2016, 11, e0154776. [Google Scholar] [CrossRef] [Green Version]
- Machado, M.; Machado, S.; Pimentel, F.B.; Freitas, V.; Alves, R.C.; Oliveira, M.B.P.P. Amino Acid Profile and Protein Quality Assessment of Macroalgae Produced in an Integrated Multi-Trophic Aquaculture System. Foods 2020, 9, 1382. [Google Scholar] [CrossRef]
- Zhang, J.; Kitazawa, D. Assessing the bio-mitigation effect of integrated multi-trophic aquaculture on marine environment by a numerical approach. Mar. Pollut. Bull. 2016, 110, 484–492. [Google Scholar] [CrossRef]
- Chary, K.; Aubin, J.; Sadoul, B.; Fiandrino, A.; Covès, D.; Callier, M.D. Integrated multi-trophic aquaculture of red drum (Sciaenops ocellatus) and sea cucumber (Holothuria scabra): Assessing bioremediation and life-cycle impacts. Aquaculture 2020, 516, 734621. [Google Scholar] [CrossRef]
- Hasselström, L.; Thomas, J.B.; Nordström, J.; Cervin, G.; Nylund, G.M.; Pavia, H.; Gröndahl, F. Socioeconomic prospects of a seaweed bioeconomy in Sweden. Sci. Rep. 2020, 10, 1–7. [Google Scholar]
- Xiao, X.; Agusti, S.; Lin, F.; Li, K.; Pan, Y.; Yu, Y.; Zheng, Y.; Wu, J.; Duarte, C.M. Nutrient removal from Chinese coastal waters by large-scale seaweed aquaculture. Sci. Rep. 2017, 7, 46613. [Google Scholar] [CrossRef] [Green Version]
- Geo, L.O.; Halim; Ariani, W.O.R. Farming production analysis of seaweed and farmer’s perception towards climate change effect in Southeast Sulawesi, Indonesia. Pak. J. Biol. Sci. 2020, 23, 1004–1009. [Google Scholar]
- Kadam, S.U.; Álvarez, C.; Tiwari, B.K.; O’Donnell, C.P. Processing of Seaweeds; Elsevier: Amsterdam, The Netherlands, 2015; pp. 61–78. [Google Scholar]
- Chan, J.C.-C.; Cheung, P.C.-K.; Ang, P.O. Comparative Studies on the Effect of Three Drying Methods on the Nutritional Composition of Seaweed Sargassum hemiphyllum (Turn.) C. Ag. J. Agric. Food Chem. 1997, 45, 3056–3059. [Google Scholar] [CrossRef]
- Neoh, Y.Y.; Matanjun, P.; Lee, J.S. Comparative study of drying methods on chemical constituents of Malaysian red seaweed. Dry. Technol. 2016, 34, 1745–1751. [Google Scholar] [CrossRef]
- Brugger, D.; Bolduan, C.; Becker, C.; Buffler, M.; Zhao, J.; Windisch, W.M. Effects of whole plant brown algae (Laminaria japonica) on zootechnical performance, apparent total tract digestibility, faecal characteristics and blood plasma urea in weaned piglets. Arch. Anim. Nutr. 2019, 74, 19–38. [Google Scholar] [CrossRef]
- Balasubramanian, B.; Shanmugam, S.; Park, S.; Recharla, N.; Koo, J.; Andretta, I.; Kim, I. Supplemental Impact of Marine Red Seaweed (Halymenia palmata) on the Growth Performance, Total Tract Nutrient Digestibility, Blood Profiles, Intestine Histomorphology, Meat Quality, Fecal Gas Emission, and Microbial Counts in Broilers. Animals 2021, 11, 1244. [Google Scholar] [CrossRef] [PubMed]
- Krogdahl, A.; Jaramillo-Torres, A.; Ahlstrøm, Ø.; Chikwati, E.; Aasen, I.-M.; Kortner, T.M. Protein value and health aspects of the seaweeds Saccharina latissima and Palmaria palmata evaluated with mink as model for monogastric animals. Anim. Feed. Sci. Technol. 2021, 276, 114902. [Google Scholar] [CrossRef]
- Graiff, A.; Ruth, W.; Kragl, U.; Karsten, U. Chemical characterization and quantification of the brown algal storage compound laminarin—A new methodological approach. Environ. Boil. Fishes 2015, 28, 533–543. [Google Scholar] [CrossRef]
- O’Doherty, J.; McDonnell, P.; Figat, S. The effect of dietary laminarin and fucoidan in the diet of the weanling piglet on performance and selected faecal microbial populations. Livest. Sci. 2010, 134, 208–210. [Google Scholar] [CrossRef]
- Ford, L.; Curry, C.; Campbell, M.; Theodoridou, K.; Sheldrake, G.; Dick, J.; Stella, L.; Walsh, P.J. Effect of Phlorotannins from Brown Seaweeds on the In Vitro Digestibility of Pig Feed. Animals 2020, 10, 2193. [Google Scholar] [CrossRef]
- Jerez-Timaure, N.; Sánchez-Hidalgo, M.; Pulido, R.; Mendoza, J. Effect of Dietary Brown Seaweed (Macrocystis pyrifera) Additive on Meat Quality and Nutrient Composition of Fattening Pigs. Foods 2021, 10, 1720. [Google Scholar] [CrossRef]
- Dierick, N.; Ovyn, A.; De Smet, S. Effect of feeding intact brown seaweed Ascophyllum nodosum on some digestive parameters and on iodine content in edible tissues in pigs. J. Sci. Food Agric. 2009, 89, 584–594. [Google Scholar] [CrossRef]
- Moroney, N.; O’Grady, M.; O’Doherty, J.; Kerry, J. Addition of seaweed (Laminaria digitata) extracts containing laminarin and fucoidan to porcine diets: Influence on the quality and shelf-life of fresh pork. Meat Sci. 2012, 92, 423–429. [Google Scholar] [CrossRef]
- Moroney, N.; O’Grady, M.; Robertson, R.C.; Stanton, C.; O’Doherty, J.; Kerry, J. Influence of level and duration of feeding polysaccharide (laminarin and fucoidan) extracts from brown seaweed (Laminaria digitata) on quality indices of fresh pork. Meat Sci. 2015, 99, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Michalak, I.; Chojnacka, K.; Korniewicz, D. Effect of Marine Macroalga Enteromorpha sp. Enriched with Zn(II) and Cu(II) ions on the Digestibility, Meat Quality and Carcass Characteristics of Growing Pigs. J. Mar. Sci. Eng. 2020, 8, 347. [Google Scholar] [CrossRef]
- Liu, Q.; Long, Y.; Zhang, Y.; Zhang, Z.; Yang, B.; Chen, C.; Huang, L.; Su, Y. Phenotypic and genetic correlations of pork myoglobin content with meat colour and other traits in an eight breed-crossed heterogeneous population. Animal 2021, 15, 100364. [Google Scholar] [CrossRef]
- Ordway, G.A.; Garry, D.J. Myoglobin: An essential hemoprotein in striated muscle. J. Exp. Biol. 2004, 207, 3441–3446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoa, V.-B.; Cho, S.-H.; Seong, P.-N.; Kang, S.-M.; Kim, Y.-S.; Moon, S.-S.; Choi, Y.-M.; Kim, J.-H.; Seol, K.-H. The significant influences of pH, temperature and fatty acids on meat myoglobin oxidation: A model study. J. Food Sci. Technol. 2020, 58, 3972–3980. [Google Scholar] [CrossRef]
- Kim, G.-D.; Jeong, J.-Y.; Hur, S.J.; Yang, H.-S.; Jeon, J.-T.; Joo, S.-T. The Relationship between Meat Color (CIE L* and a*), Myoglobin Content, and Their Influence on Muscle Fiber Characteristics and Pork Quality. Food Sci. Anim. Resour. 2010, 30, 626–633. [Google Scholar] [CrossRef] [Green Version]
- Moroney, N.C.; O’Grady, M.N.; Lordan, S.; Stanton, C.; Kerry, J.P. Seaweed Polysaccharides (Laminarin and Fucoidan) as Functional Ingredients in Pork Meat: An Evaluation of Anti-Oxidative Potential, Thermal Stability and Bioaccessibility. Mar. Drugs 2015, 13, 2447–2464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigors, S.; O’Doherty, J.; Rattigan, R.; Sweeney, T. Effect of Supplementing Seaweed Extracts to Pigs until d35 Post-Weaning on Performance and Aspects of Intestinal Health. Mar. Drugs 2021, 19, 183. [Google Scholar] [CrossRef]
- Sweeney, T.; O’Doherty, J. Marine macroalgal extracts to maintain gut homeostasis in the weaning piglet. Domest. Anim. Endocrinol. 2016, 56, S84–S89. [Google Scholar] [CrossRef]
- Moroney, N.; O’Grady, M.; O’Doherty, J.; Kerry, J. Effect of a brown seaweed (Laminaria digitata) extract containing laminarin and fucoidan on the quality and shelf-life of fresh and cooked minced pork patties. Meat Sci. 2013, 94, 304–311. [Google Scholar] [CrossRef]
- Rajauria, G.; Draper, J.; McDonnell, M.; O’Doherty, J. Effect of dietary seaweed extracts, galactooligosaccharide and vitamin E supplementation on meat quality parameters in finisher pigs. Innov. Food Sci. Emerg. Technol. 2016, 37, 269–275. [Google Scholar] [CrossRef]
- Bocanegra, A.; Bastida, S.; Benedi, J.; Ródenas, S.; Sánchez-Muniz, F.J. Characteristics and Nutritional and Cardiovascular-Health Properties of Seaweeds. J. Med. Food 2009, 12, 236–258. [Google Scholar] [CrossRef] [PubMed]
- Vigors, S.; O’Doherty, J.V.; Rattigan, R.; McDonnell, M.J.; Rajauria, G.; Sweeney, T. Effect of a Laminarin Rich Macroalgal Extract on the Caecal and Colonic Microbiota in the Post-Weaned Pig. Mar. Drugs 2020, 18, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulshreshtha, G.; Hincke, M.T.; Prithiviraj, B.; Critchley, A. A Review of the Varied Uses of Macroalgae as Dietary Supplements in Selected Poultry with Special Reference to Laying Hen and Broiler Chickens. J. Mar. Sci. Eng. 2020, 8, 536. [Google Scholar] [CrossRef]
- Ahmed, S.T.; Mun, H.-S.; Islam, M.M.; Yang, C.-J. Effects of fermentedcorni fructusand fermented kelp on growth performance, meat quality, and emission of ammonia and hydrogen sulphide from broiler chicken droppings. Br. Poult. Sci. 2014, 55, 745–751. [Google Scholar] [CrossRef]
- Islam, M.M.; Ahmed, S.T.; Mun, H.S.; Kim, Y.J.; Yang, C.J. Effect of Sea Tangle (Laminaria japonica) and Charcoal Supplementation as Alternatives to Antibiotics on Growth Performance and Meat Quality of Ducks. Asian-Australas. J. Anim. Sci. 2014, 27, 217–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Ghany, A.M. effects of kelp seaweed (ascophyllum nodosum) on growth performance during the growing period of an imported heavy-weight-line of broad breasted bronze turkeys. Egypt. Poult. Sci. J. 2020, 40, 47–64. [Google Scholar] [CrossRef] [Green Version]
- Bonos, E.; Kargopoulos, A.; Nikolakakis, I.; Paneri, P.F.; Christaki, E. The Seaweed Ascophyllum nodosum as a Potential Functional Ingredient in Chicken Nutrition. J. Oceanogr. Mar. Res. 2017, 4, 140. [Google Scholar] [CrossRef]
- Paul, S.S.; Venkata, H.G.R.V.; Raju, M.; Rao, S.V.R.; Nori, S.S.; Suryanarayan, S.; Kumar, V.; Perveen, Z.; Prasad, C.S. Dietary supplementation of extracts of red sea weed (Kappaphycus alvarezii) improves growth, intestinal morphology, expression of intestinal genes and immune responses in broiler chickens. J. Sci. Food Agric. 2021, 101, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Abudabos, A.M.; Okab, A.B.; Aljumaah, R.; Samara, E.; Abdoun, K.; Al-Haidary, A.A. Nutritional Value of Green Seaweed (Ulva Lactuca) for Broiler Chickens. Ital. J. Anim. Sci. 2013, 12, e28. [Google Scholar] [CrossRef]
- Matshogo, T.; Mlambo, V.; Mnisi, C.; Manyeula, F. Effect of pre-treating dietary green seaweed with fibrolytic enzymes on growth performance, blood indices, and meat quality parameters of Cobb 500 broiler chickens. Livest. Sci. 2021, 251, 104652. [Google Scholar] [CrossRef]
- Zhu, W.; Li, D.; Wang, J.; Wu, H.; Xia, X.; Bi, W.; Guan, H.; Zhang, L. Effects of polymannuronate on performance, antioxidant capacity, immune status, cecal microflora, and volatile fatty acids in broiler chickens. Poult. Sci. 2015, 94, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.N.; Albrecht, D.; Scholz, A.; Gutierrez-Buey, G.; Lazarus, J.H.; Dayan, C.M.; Okosieme, O.E. Global epidemiology of hyperthyroidism and hypothyroidism. Nat. Rev. Endocrinol. 2018, 14, 301–316. [Google Scholar] [CrossRef]
- Chittimoju, S.B.; Pearce, E.N. Iodine Deficiency and Supplementation in Pregnancy. Clin. Obstet. Gynecol. 2019, 62, 330–338. [Google Scholar] [CrossRef]


{kind=link}
| Seaweed | Dry Matter | Ash | Crude Protein | Crude Fat | Crude Fibre | NDF | ADF | ADL | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Phaeophyceae (brown) | |||||||||
| Laminaria japonica | 97.5 | 14.9 | 20.5 | 3.0 | 13.3 | 35.6 | 28.8 | N/A | [53] |
| Ascophylum nodosum | 93.2 | 29.5 | 11.4 | 3.0 | N/A | 34.5 | 18.9 | 12.9 | [11] |
| Sacharina latissimi | 94.0 | 39.9 | 15.2 | 1.5 | N/A | 21.7 | 8.0 | 2.8 | [11] |
| Chlorophyceae (green) | |||||||||
| Ulva sp. | 93.6 | 51.3 | 14.6 | 1.15 | N/A | 21.0 | 7.45 | 3.2 | [11] |
| Rhodophyceae (red) | |||||||||
| Halymenia palmata | 90.6 | 19.0 | 18.5 | 1.69 | 1.83 | N/A | N/A | N/A | [54] |
| Palmaria palmata | 93.6 | 21.0 | 26.8 | 8.0 | N/A | N/A | N/A | N/A | [55] |
| Seaweed | Incorporation Rate/Type of Product | Animal and Initial Live Weight | Main Findings | Reference |
|---|---|---|---|---|
| Macrocystis pyrifera (brown) | 0%, 2%, and 4% dietary seaweed meal | Castrated male and female pigs 52.5 ± 2.8 kg | Meat from pigs fed with 4% seaweed had less red intensity (a*) compared to control and 2% group Meat ash content was increased in 4% group compared to the other groups, albeit having significantly less Mn, Fe, and Cu | [59] |
| Ascophylum nodosum (brown) | 0% and 2% dietary seaweed meal | Male and female piglets (Seghers hybrid × Pietrain) 6.88 ± 1.21 kg | Significant accumulation of iodine in several tissues, including muscle | [60] |
| Laminaria digitata (brown) | 500 and 420 mg/kg of feed of laminarin and fucoidan, respectively | Male and female pigs (Large White × Landrace) 14.51 kg | Decreased lipid oxidation (TBARS) in the muscle of supplemented pigs | [61] |
| Laminaria digitata (brown) | 450 and 900 mg/kg of feed of laminarin and fucoidan | Male and female pigs (Large White × Landrace) 82 kg | Lipid oxidation was lower in the longissimus dorsi of pigs when supplemented three weeks before slaughter When pigs were supplemented for 6 weeks before slaughter, the total saturated fatty acid content of meat was lower | [62] |
| Enteromorpha sp. (green) | 4% (starter), 3% (grower), and 2.5% (finisher) (premix containing enriched biomass with Cu and Zn) | Male and female pigs (Polish Landrace × Polish Large White × Hampshire/Pietrain) 40 kg | No effects on meat or carcass quality parameters | [63] |
| Seaweed | Incorporation Rate/Type of Product | Animal and Initial Live Weight | Main Findings | Reference |
|---|---|---|---|---|
| Laminaria japonica (brown) | 0% and 0.5% fermented seaweed meal | Ross broiler chickens | Reduced lipid oxidation of breast/thigh meat mixture | [76] |
| Laminaria japonica (brown) | 0%, 0.1%, 0.5%, and 1% of seaweed meal–charcoal (1:1) mixture | Ducks (Cherry berry, SUPER M3 F1) | Cholesterol was significantly reduced in meat of 1% supplemented ducks Supplementation reduced meat lipid oxidation The group of 1% had higher content of C20:5 n-3, C22:6 n-3, and overall n-3 PUFA in meat | [77] |
| Ascophylum nodosum (brown) | 0%, 1%, and 2% seaweed meal | Broad Breasted Bronze turkeys 2567 ± 39.1 g | Supplementation increased eviscerated weight Redness of thigh and breast meat was increased in supplemented groups | [78] |
| Ascophylum nodosum (brown) | 0%, 0.5%, 1%, and 2% seaweed meal | Broiler chickens | No effect on lipid oxidation Supplementation increased C18:3 n-6 in breast meat and decreased C20:1 n-9 in thigh meat | [79] |
| Halymenia palmata (red) | 0%, 0.05%, 0.1%, 0,15%, 0.25% seaweed meal | Ross 308 broiler chickens 45 ± 0.5 g | Linear decrease in cooking loss and drip loss (day 7) with increasing seaweed Linear increase in breast yield and gizzard weight with increasing seaweed | [54] |
| Kappaphycus alvarezii (red) | 0%, 0.05%, 0.15%, and 0.5% dried alkaline extract and aqueous extract | Vencobb 400 broiler chickens | No effect on meat or carcass characteristics | [80] |
| Ulva lactuca (green) | 0%, 1%, and 3% seaweed meal, replacing maize | Ross chickens | Supplementation increased breast yield and reduced abdominal fat, 3% dietary inclusion increased dressing yield There was no effect on breast meat colour | [81] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, D.M.; Martins, C.F.; Costa, M.; Coelho, D.; Pestana, J.; Alfaia, C.; Lordelo, M.; de Almeida, A.M.; Freire, J.P.B.; Prates, J.A.M. Quality Traits and Nutritional Value of Pork and Poultry Meat from Animals Fed with Seaweeds. Foods 2021, 10, 2961. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10122961
Ribeiro DM, Martins CF, Costa M, Coelho D, Pestana J, Alfaia C, Lordelo M, de Almeida AM, Freire JPB, Prates JAM. Quality Traits and Nutritional Value of Pork and Poultry Meat from Animals Fed with Seaweeds. Foods. 2021; 10(12):2961. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10122961
Chicago/Turabian StyleRibeiro, David Miguel, Cátia Falcão Martins, Mónica Costa, Diogo Coelho, José Pestana, Cristina Alfaia, Madalena Lordelo, André Martinho de Almeida, João Pedro Bengala Freire, and José António Mestre Prates. 2021. "Quality Traits and Nutritional Value of Pork and Poultry Meat from Animals Fed with Seaweeds" Foods 10, no. 12: 2961. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10122961