
MALDI-TOF MS Based Typing for Rapid Screening of Multiple Antibiotic Resistance E. coli and Virulent Non-O157 Shiga Toxin-Producing E. coli Isolated from the Slaughterhouse Settings and Beef Carcasses

 ,
,  , and
, and
Abstract
:1. Introduction
Literature Review
2. Materials and Methods
2.1. Sampling and Sample Collection
2.2. Isolation and Identification of Bacterial Strains
2.3. Antimicrobial Resistance
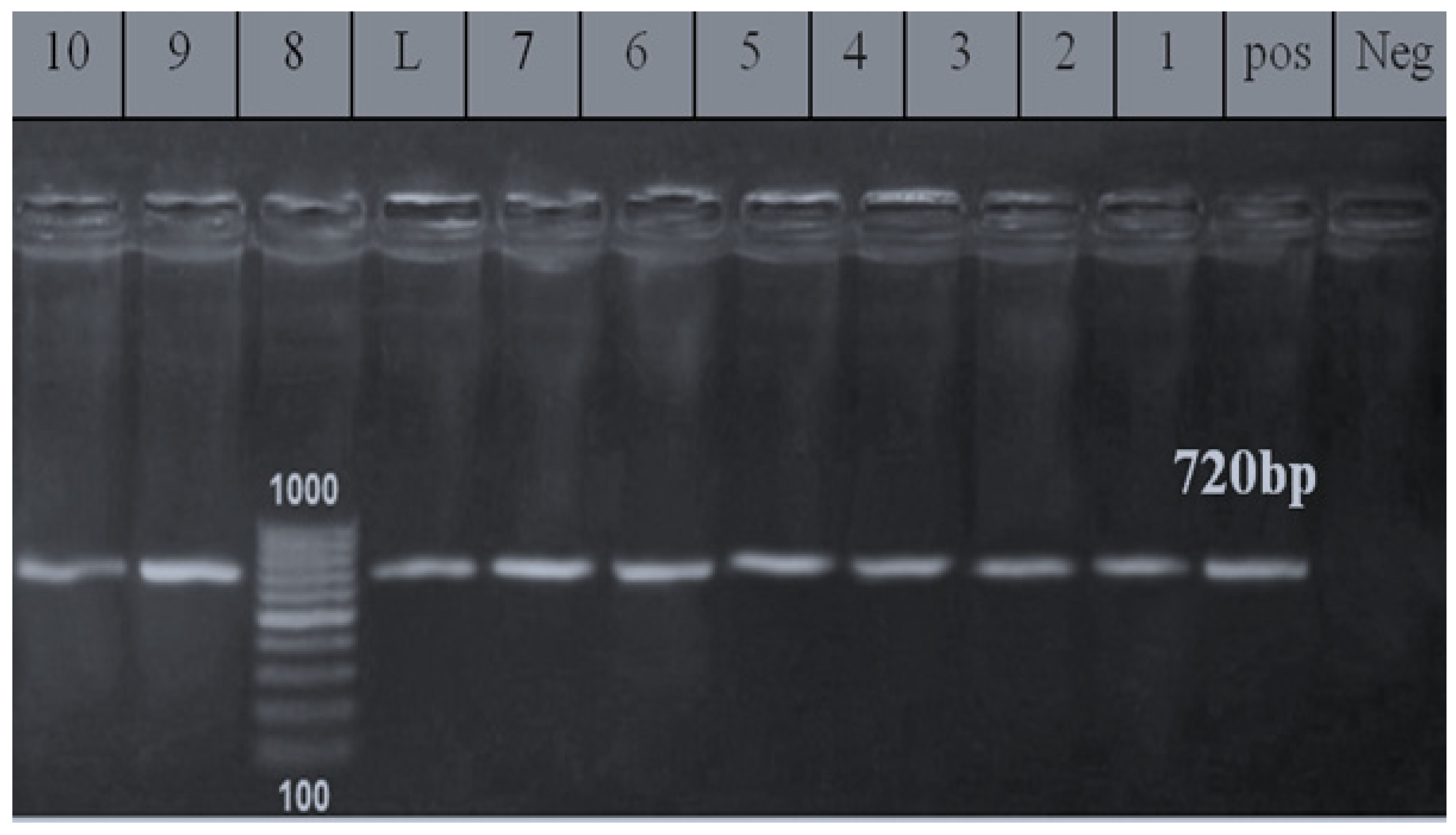
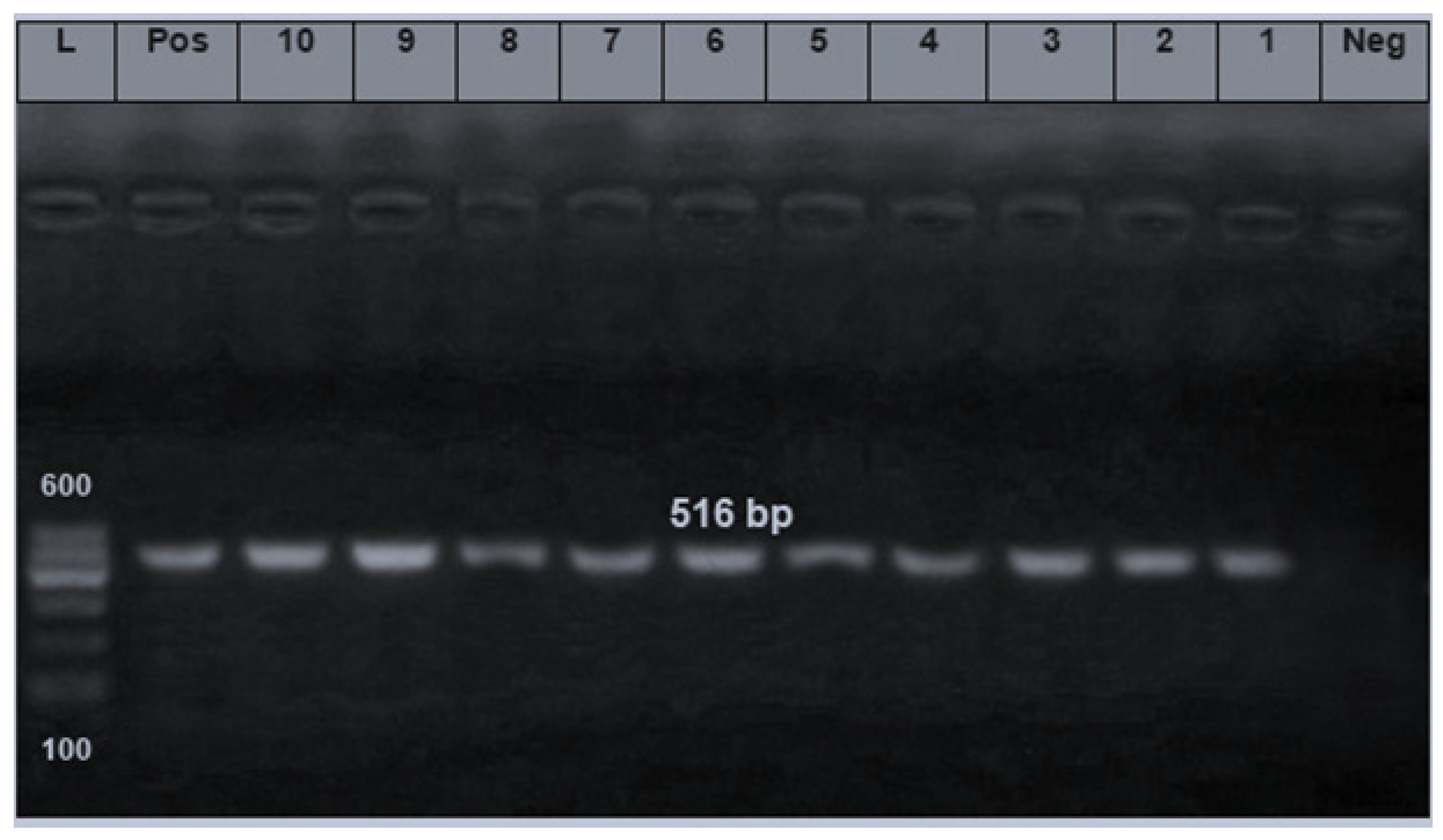
2.4. PCR Assay
2.5. Statistical Analysis
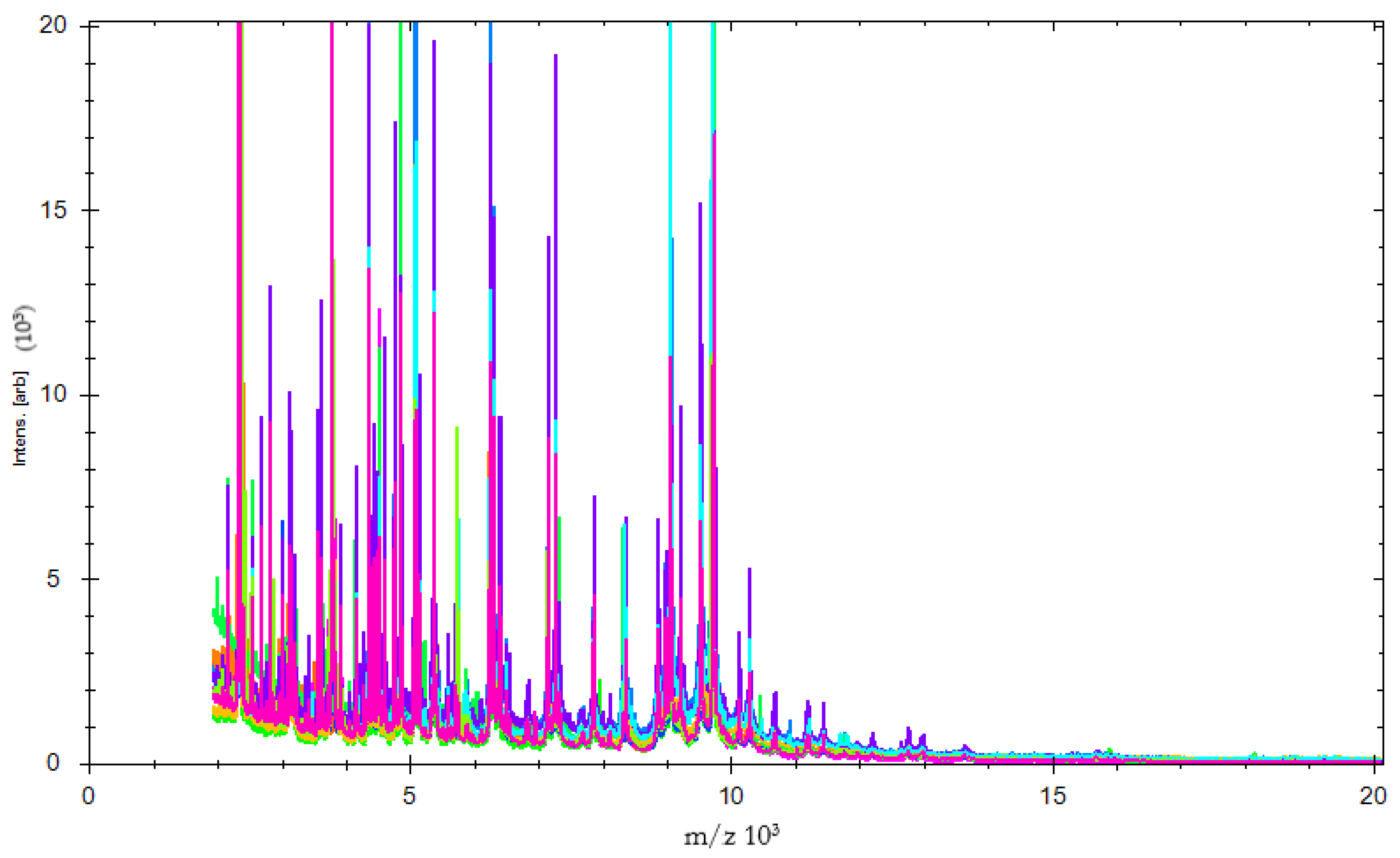
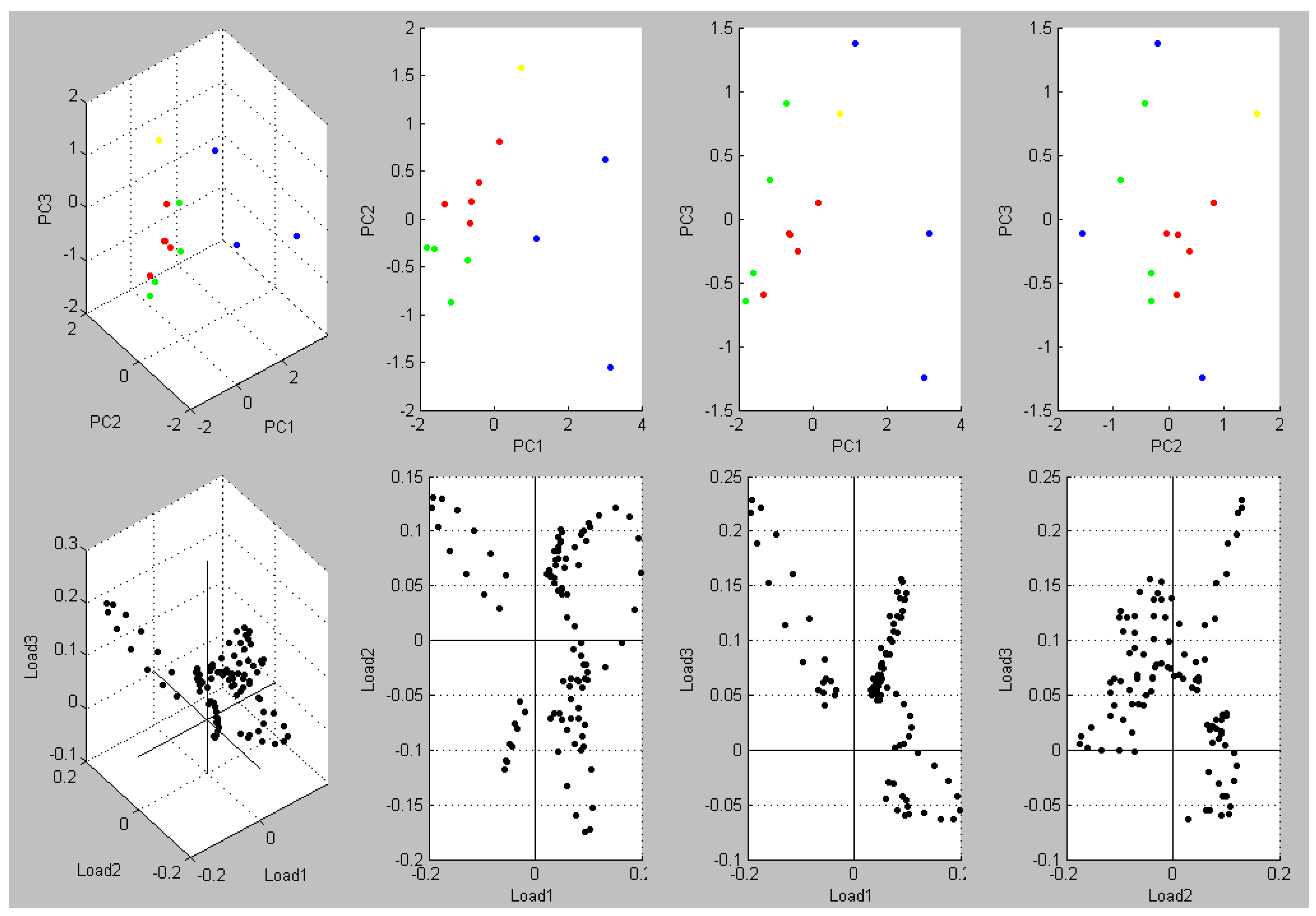
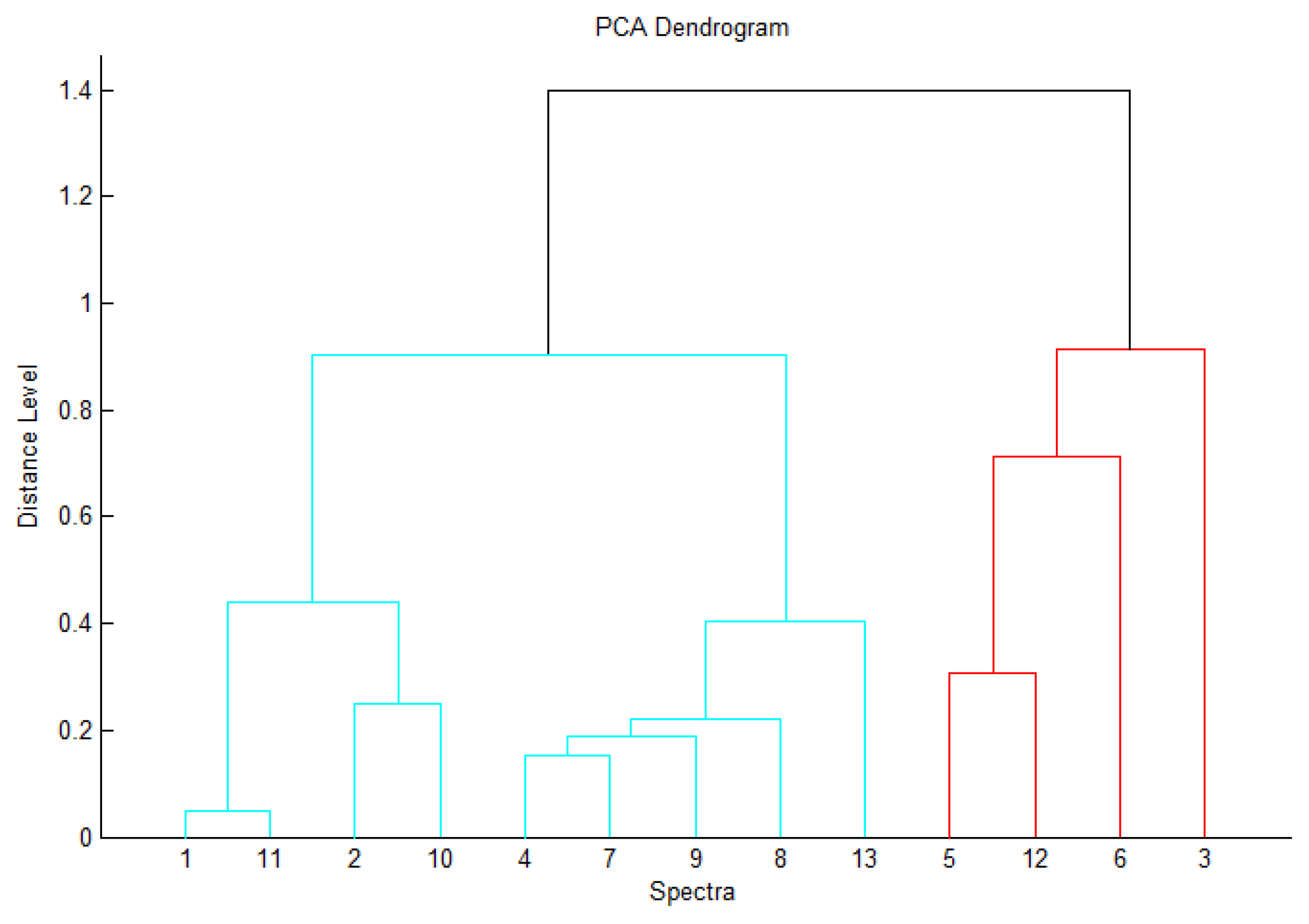
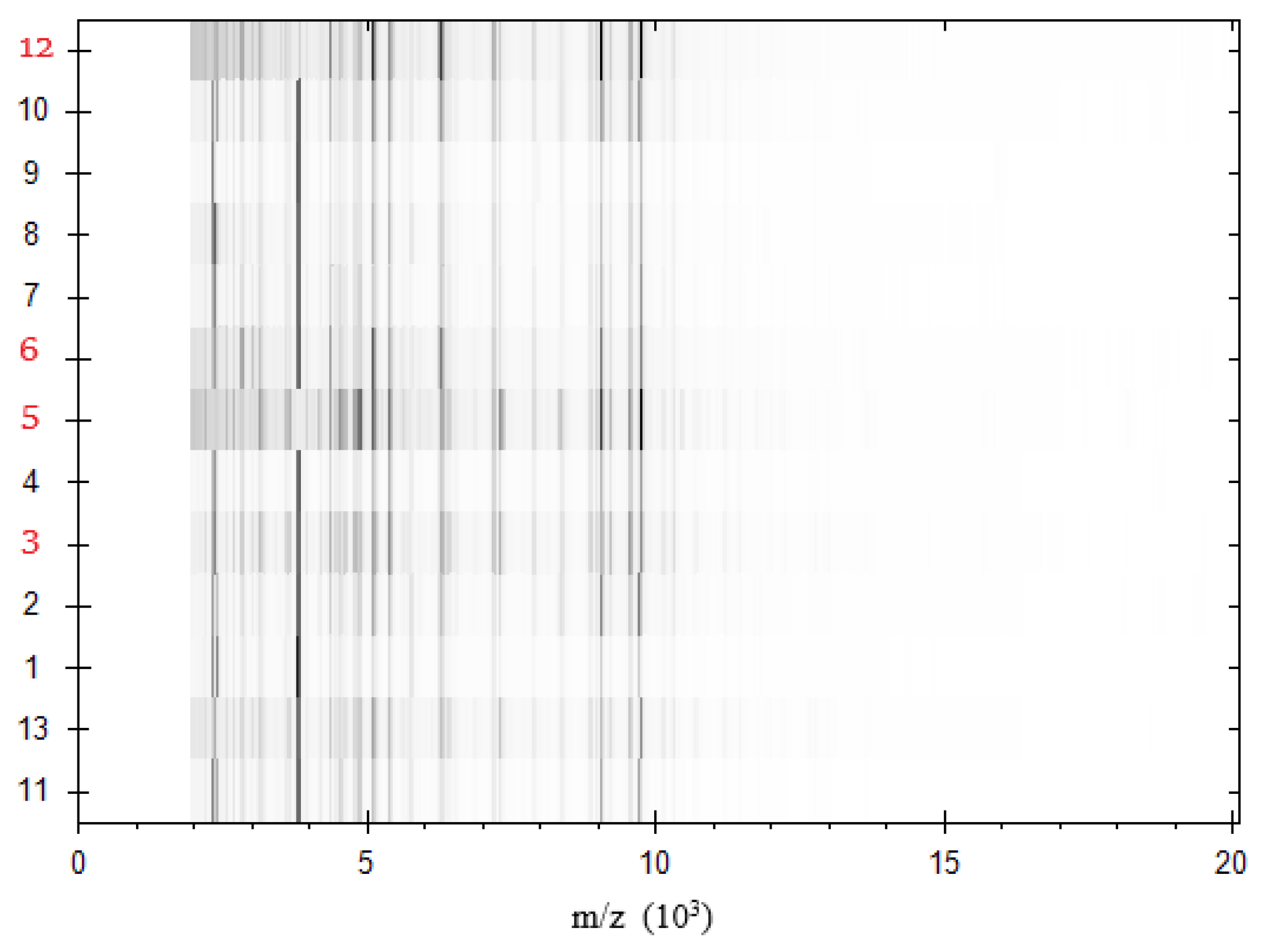
3. Results and Discussion
4. Implications for Practice and Theory
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Käppeli, U.; Hächler, H.; Giezendanner, N.; Beutin, L.; Stephan, R. Human Infections with Non-O157 Shiga Toxin–producing Escherichia coli, Switzerland, 2000–2009. Emerg. Infect. Dis. 2011, 17, 180–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Zogibi, O.G.; Mohamed, M.I.; Hessain, A.M.; El-Jakee, J.K.; Kabli, S.A. Molecular and serotyping characterization of shiga toxogenic Escherichia coli associated with food collected from Saudi Arabia. Saudi J. Biol. Sci. 2015, 22, 438–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- USDA; FJFR. Shiga toxin-producing Escherichia coli in certain raw beef products. Fed. Regist. 2011, 76, 58157–58165. [Google Scholar]
- Wilson, J.B.; Clarke, R.C.; Renwick, S.A.; Johnson, R.P.; Karmali, M.A.; Lior, H.; Alves, D.; Gyles, C.L.; Sandhu, K.S.; McEwen, S.A. Vero cytotoxigenic Escherichia coli infection in dairy farm fami-lies. J. Infect. Dis. 1996, 174, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, C.; Doyle, M.E.; Archer, J. White paper on non-O157: H7 Shiga toxin-producing E. coli from meat and non-meat sources. FRI Food Saf. Rev. 2010, 1–26. Available online: http://meatpoultryfoundation.org/research/white-paper-non-o157h7-shiga-toxin-producing-e-coli-meat-and-non-meat-sources (accessed on 30 September 2019).
- Osman, K.M.; ElHariri, M.; Mustafa, A.M.; Abdelhamed, G.S. Identification of serotypes and virulence markers of Escherichia coli isolated from human stool and urine samples in Egypt. Indian J. Med Microbiol. 2012, 30, 308–313. [Google Scholar] [CrossRef]
- Kanki, M.; Seto, K.; Harada, T.; Yonogi, S.; Kumeda, Y. Comparison of four enrichment broths for the detection of non-O157 Shiga-toxin-producing Escherichia coli O91, O103, O111, O119, O121, O145 and O165 from pure culture and food samples. Lett. Appl. Microbiol. 2011, 53, 167–173. [Google Scholar] [CrossRef]
- Fratamico, P.M.; Bagi, L.K.; Cray, W.C., Jr.; Narang, N.; Yan, X.; Medina, M.; Liu, Y. Detection by multiplex real-time polymerase chain reaction assays and isolation of Shiga toxin–producing Escherichia coli serogroups O26, O45, O103, O111, O121, and O145 in ground beef. Foodborne Pathog. Dis. 2011, 8, 601–607. [Google Scholar] [CrossRef] [Green Version]
- Madic, J.; Vingadassalon, N.; De Garam, C.P.; Marault, M.; Scheutz, F.; Brugère, H.; Jamet, E.; Auvray, F. Detection of Shiga Toxin-Producing Escherichia coli Serotypes O26:H11, O103:H2, O111:H8, O145:H28, and O157:H7 in Raw-Milk Cheeses by Using Multiplex Real-Time PCR. Appl. Environ. Microbiol. 2011, 77, 2035–2041. [Google Scholar] [CrossRef] [Green Version]
- Verstraete, K.; De Zutter, L.; Messens, W.; Herman, L.; Heyndrickx, M.; De Reu, K. Effect of the enrichment time and immunomagnetic separation on the detection of Shiga toxin-producing Escherichia coli O26, O103, O111, O145 and sorbitol positive O157 from artificially inoculated cattle faeces. Veter Microbiol. 2010, 145, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Beutin, L.; Jahn, S.; Fach, P. Evaluation of the ‘GeneDisc’real--time PCR system for detection of enterohaemorrhagic Escherichia coli (EHEC) O26, O103, O111, O145 and O157 strains according to their virulence markers and their O--and H--antigen--associated genes. J. Appl. Microbiol. 2009, 106, 1122–1132. [Google Scholar] [CrossRef]
- Johnson, R.P.; Clarke, R.C.; Wilson, J.B.; Read, S.C.; Rahn, K.; Renwick, S.A.; Sandhu, K.A.; Alves, D.; Karmali, M.A.; Lior, H. Growing concerns and recent outbreaks involving non-O157: H7 serotypes of verotoxigenic Escherichia coli. J. Food Prot. 1996, 59, 1112–1122. [Google Scholar] [CrossRef] [PubMed]
- Egli, A.; Tschudin-Sutter, S.; Oberle, M.; Goldenberger, D.; Frei, R.; Widmer, A.F. Matrix-Assisted Laser Desorption/Ionization Time of Flight Mass-Spectrometry (MALDI-TOF MS) Based Typing of Extended-Spectrum β-Lactamase Producing E. coli—A Novel Tool for Real-Time Outbreak Investigation. PLoS ONE 2015, 10, e0120624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brusa, V.; Restovich, V.; Galli, L.; Teitelbaum, D.; Signorini, M.; Brasesco, H.; Londero, A.; García, D.; Padola, N.L.; Superno, V.; et al. Isolation and characterization of non-O157 Shiga toxin-producing Escherichia coli from beef carcasses, cuts and trimmings of abattoirs in Argentina. PLoS ONE 2017, 12, e0183248. [Google Scholar] [CrossRef] [PubMed]
- Van Veen, S.Q.; Claas, E.C.J.; Kuijper, E.J. High-Throughput Identification of Bacteria and Yeast by Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry in Conventional Medical Microbiology Laboratories. J. Clin. Microbiol. 2010, 48, 900–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute, CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Christopher, A.F.; Hora, S.; Ali, Z. Investigation of plasmid profile, antibiotic susceptibility pattern multiple antibiotic resistance index calculation of Escherichia coli isolates obtained from different human clinical specimens at tertiary care hospital in Bareilly-India. Ann. Trop. Med. Public Health 2013, 6, 285. [Google Scholar] [CrossRef]
- Vásquez-García, A.; de Oliveira, A.P.S.C.; Mejia-Ballesteros, J.E.; de Godoy, S.H.S.; Barbieri, E.; de Sousa, R.L.M.; Fernandes, A.M.J.A. Escherichia coli detection and identification in shellfish from southeastern Brazil. Aquaculture 2019, 504, 158–163. [Google Scholar] [CrossRef]
- Algammal, A.M.; Hetta, H.F.; Batiha, G.E.; Hozzein, W.N.; El Kazzaz, W.M.; Hashem, H.R.; Tawfik, A.M.; El-Tarabili, R.M. Virulence-determinants and antibiotic-resistance genes of MDR-E. coli isolated from secondary infections following FMD-outbreak in cattle. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Algammal, A.M.; El-Kholy, A.W.; Riad, E.M.; Mohamed, H.E.; Elhaig, M.M.; Yousef, S.A.A.; Hozzein, W.N.; Ghobashy, M.O.J.T. Genes Encoding the Virulence and the Antimicrobial Resistance in Enterotoxigenic and Shiga-Toxigenic E. coli Isolated from Diarrheic Calves. Toxins 2020, 12, 383. [Google Scholar] [CrossRef]
- Sofos, J.N.; Flick, G.; Nychas, G.-J.; O’Bryan, C.A.; Ricke, S.C.; Crandall, P.G. Meat, Poultry, and Seafood. In Food Microbiology; American Society for Microbiology: Washington, DC, USA, 2014; pp. 109–167. [Google Scholar]
- Strejcek, M.; Smrhova, T.; Junkova, P.; Uhlik, O. Whole-Cell MALDI-TOF MS Versus 16S rRNA Gene Analysis for Identification and Dereplication of Recurrent Bacterial Isolates. Front. Microbiol. 2018, 9, 1294. [Google Scholar] [CrossRef] [PubMed]
- Nehoya, K.N.; Hamatui, N.; Shilangale, R.P.; Onywera, H.; Kennedy, J.; Mwapagha, L.M. Characterization of Shiga toxin-producing Escherichia coli in raw beef from informal and commercial abattoirs. PLoS ONE 2020, 15, e0243828. [Google Scholar] [CrossRef] [PubMed]
- Ayalew, H.; Berhanu, A.; Sibhat, B.; Serda, B.J.I.J.o.F.; Sciences, A. Microbiological assessment of meat contact surfaces at abattoir and retail houses in Jigjiga town, Somali National Regional State of Ethiopia. J. Food Agric. Sci. 2015, 5, 21–26. [Google Scholar]
- Elabbasy, M.; Morshdy, A.E. Studies on Retail Chicken Meats with A Special Reference to Antibiotic Resistant Bacteria. SCVMJ 2015, 20, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Adzitey, F. Incidence and antimicrobial susceptibility of Escherichia coli isolated from beef (meat muscle, liver and kidney) samples in Wa Abattoir, Ghana. Cogent Food Agric. 2020, 6, 1718269. [Google Scholar] [CrossRef]
- Rao, T.S.; Gill, J.P.S.; Gvvps, R.K.; Ghatak, S. Multi drug resistance patterns of Shiga toxin” producing Escherichia coli (STEC) and non” STEC isolates from meats, RTE meat foods, drinking water and human diarrhoeic samples of Punjab, India. Arch. Clin. Microbiol. 2011, 2, 1–12. [Google Scholar]
- Gómez-Duarte, O.G.; Kaper, J.B. A plasmid-encoded regulatory region activates chromosomal eaeA expression in enteropathogenic Escherichia coli. Infect. Immun. 1995, 63, 1767–1776. [Google Scholar] [CrossRef] [Green Version]
- Medina, A.; Horcajo, P.; Jurado, S.; De La Fuente, R.; Ruiz-Santa-Quiteria, J.A.; Domínguez-Bernal, G.; Orden, J.A. Phenotypic and genotypic characterization of antimicrobial resistance in enterohemorrhagic Escherichia coli and atypical enteropathogenic E. coli strains from ruminants. J. Veter- Diagn. Investig. 2011, 23, 91–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamed, O.M.; Sabry, M.A.; Hassanain, N.A.; Hamza, E.; Hegazi, A.G.; Salman, M.B. Occurrence of virulent and antibiotic-resistant Shiga toxin-producing Escherichia coli in some food products and human stool in Egypt. Veter World 2017, 10, 1233–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Ye, L.; Kromann, S.; Meng, H. Occurrence of Extended-Spectrum β-Lactamases, Plasmid-Mediated Quinolone Resistance, and Disinfectant Resistance Genes in Escherichia coli Isolated from Ready-To-Eat Meat Products. Foodborne Pathog. Dis. 2017, 14, 109–115. [Google Scholar] [CrossRef]
- Moawad, A.A.; Hotzel, H.; Awad, O.; Tomaso, H.; Neubauer, H.; Hafez, H.M.; El-Adawy, H. Occurrence of Salmonella enterica and Escherichia coli in raw chicken and beef meat in northern Egypt and dissemination of their antibiotic resistance markers. Gut Pathog. 2017, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Seo, K.W.; Lee, Y.J. Prevalence and Characterization of β-Lactamases Genes and Class 1 Integrons in Multidrug-Resistant Escherichia coli Isolates from Chicken Meat in Korea. Microb. Drug Resist. 2018, 24, 1599–1606. [Google Scholar] [CrossRef]
- Bosilevac, J.M.; Gassem, M.A.; Al Sheddy, I.A.; Almaiman, S.A.; Al-Mohizea, I.S.; Alowaimer, A.; Koohmaraie, M. Prevalence of Escherichia coli O157:H7 and Salmonella in Camels, Cattle, Goats, and Sheep Harvested for Meat in Riyadh†. J. Food Prot. 2015, 78, 89–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tóth, I.; Schmidt, H.; Kardos, G.; Lancz, Z.; Creuzburg, K.; Damjanova, I.; Pászti, J.; Beutin, L.; Nagy, B. Virulence Genes and Molecular Typing of Different Groups of Escherichia coli O157 Strains in Cattle. Appl. Environ. Microbiol. 2009, 75, 6282–6291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, H.S.; Bollinger, L.M. Prevalence of Shiga Toxin–Producing Escherichia coli in Beef Cattle. J. Food Prot. 2005, 68, 2224–2241. [Google Scholar] [CrossRef] [PubMed]
- Barkocy-Gallagher, G.A.; Arthur, T.M.; Rivera-Betancourt, M.; Nou, X.; Shackelford, S.D.; Wheeler, T.L.; Koohmaraie, M. Seasonal Prevalence of Shiga Toxin–Producing Escherichia coli, Including O157:H7 and Non-O157 Serotypes, and Salmonella in Commercial Beef Processing Plants†. J. Food Prot. 2003, 66, 1978–1986. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Calvin, S.; Middleton, D. Descriptive epidemiology of verotoxin-producing E. coli reported in Ontario, 1996-2005. Can. Commun. Dis. Rep. 2007, 33, 33. [Google Scholar]
- Gill, A.; Gill, C.O. Non-O157 verotoxigenic Escherichia coli and beef: A Canadian per-spective. Can. J. Vet. Res. 2010, 74, 161–169. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Prevalence |
|---|---|
| Abattoir effluent | 20 (100%) |
| Abattoir floor | 20 (100%) |
| Abattoir wall | 11 (55%) |
| Beef brisket | 15 (75%) |
| Beef shoulder | 16 (80%) |
| Beef thigh | 13 (65%) |
| Knives | 9 (45%) |
| Water | Not detected |
| Worker hands | 6 (30%) |
| O166 | O146 | O44 | O111 | O26 | |
|---|---|---|---|---|---|
| Abattoir effluent | 3 | 2 | 4 | 8 | 3 |
| Abattoir floor | 3 | 4 | 5 | 6 | 2 |
| Abattoir wall | 2 | 3 | 2 | 2 | 2 |
| Beef brisket | 4 | 2 | 3 | 3 | 3 |
| Beef shoulder | 2 | 4 | 1 | 7 | 2 |
| Beef thigh | 3 | 1 | 2 | 4 | 3 |
| Knives | 3 | - | 2 | 3 | 1 |
| Worker hands | 1 | 2 | 1 | 2 | - |
| Total | 21 (19.10%) | 18 (16.36%) | 20 (18.18%) | 35 (31.81%) | 16 (14.54%) |
| Types | ETEC | EPEC | EPEC | EHEC | EHEC |
| Antimicrobial Agent | Sensitive | Intermediate | Resistant | Serotype | n | Pathotype |
|---|---|---|---|---|---|---|
| Penicillin (P) | - | - | 20 (100%) | O166 O146 O44 O111 O26 | 5 5 5 5 5 | ETEC EPEC EPEC EHEC EHEC |
| Erythromycin (E) | - | 2 (20%) | 18 (80%) | |||
| Oxytetracycline (T) | 3 (15%) | 2 (10%) | 15 (75%) | |||
| Nalidixic acid (NA) | 4 (20%) | 3 (15%) | 13 (65%) | |||
| Ampicillin (AM) | - | 8 (40%) | 12 (60%) | |||
| Sulfamethoxazol (SXT) | 6 (30%) | 3 (15%) | 11 (55%) | |||
| Cephalotin (CN) | 9 (45%) | 2 (10%) | 9 (45%) | |||
| Enrofloxacin (EN) | 10 (50%) | 2 (10%) | 8 (40%) | |||
| Oxacillin (OX) | 12 (60%) | 1 (5%) | 7 (35%) | |||
| Neomycin (N) | 14 (70%) | - | 6 (30%) | |||
| Chloramphenicol (C) | 16 (80%) | - | 4 (20%) | |||
| Kanamycin (K) | 15(75%) | 2 (10%) | 3 (15%) | |||
| Ciprofloxacin (CP) | 16 (80%) | 2 (10%) | 2 (10%) | |||
| Gentamicin (G) | 19 (95%) | - | 1 (5%) | |||
| p value | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| Resistance Pattern | Resistance Profile | Number of Isolates | Number of Antibiotics | MAR |
|---|---|---|---|---|
| i. | P, E, T, NA, AM, SXT, CN, EN, OX, N, C, K, CP, G | 1 | 14 | 1 |
| ii. | P, E, T, NA, AM, SXT, CN, EN, OX, N, C, K, CP | 1 | 13 | 0.92 |
| iii. | P, E, T, NA, AM, SXT, CN, EN, OX, N, C, K | 1 | 12 | 0.85 |
| iv. | P, E, T, NA, AM, SXT, CN, EN, OX, N, C | 1 | 11 | 0.78 |
| v. | P, E, T, NA, AM, SXT, CN, EN, OX, N | 2 | 10 | 0.714 |
| vi. | P, E, T, NA, AM, SXT, CN, EN, OX | 1 | 9 | 0.642 |
| vii. | P, E, T, NA, AM, SXT, CN, EN | 1 | 8 | 0.571 |
| viii. | P, E, T, NA, AM, SXT, CN | 1 | 7 | 0.5 |
| ix. | P, E, T, NA, AM, SXT | 2 | 6 | 0.428 |
| x. | P, E, T, NA, AM | 1 | 5 | 0.357 |
| xi. | P, E, T, NA | 1 | 4 | 0.285 |
| xii. | P, E, T | 2 | 3 | 0.21 |
| xiii. | P, E | 3 | 2 | 0.142 |
| xiv. | P | 2 | 1 | 0.071 |
| Average | 0.533 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elabbasy, M.T.; Hussein, M.A.; Algahtani, F.D.; Abd El-Rahman, G.I.; Morshdy, A.E.; Elkafrawy, I.A.; Adeboye, A.A. MALDI-TOF MS Based Typing for Rapid Screening of Multiple Antibiotic Resistance E. coli and Virulent Non-O157 Shiga Toxin-Producing E. coli Isolated from the Slaughterhouse Settings and Beef Carcasses. Foods 2021, 10, 820. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10040820
Elabbasy MT, Hussein MA, Algahtani FD, Abd El-Rahman GI, Morshdy AE, Elkafrawy IA, Adeboye AA. MALDI-TOF MS Based Typing for Rapid Screening of Multiple Antibiotic Resistance E. coli and Virulent Non-O157 Shiga Toxin-Producing E. coli Isolated from the Slaughterhouse Settings and Beef Carcasses. Foods. 2021; 10(4):820. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10040820
Chicago/Turabian StyleElabbasy, Mohamed Tharwat, Mohamed A. Hussein, Fahad Dhafer Algahtani, Ghada I. Abd El-Rahman, Alaa Eldin Morshdy, Ibrahim A. Elkafrawy, and Adeniyi A. Adeboye. 2021. "MALDI-TOF MS Based Typing for Rapid Screening of Multiple Antibiotic Resistance E. coli and Virulent Non-O157 Shiga Toxin-Producing E. coli Isolated from the Slaughterhouse Settings and Beef Carcasses" Foods 10, no. 4: 820. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10040820