
Alimentary and Pharmaceutical Approach to Natural Antimicrobials against Clostridioides difficile Gastrointestinal Infection

 , ,
, ,
Abstract
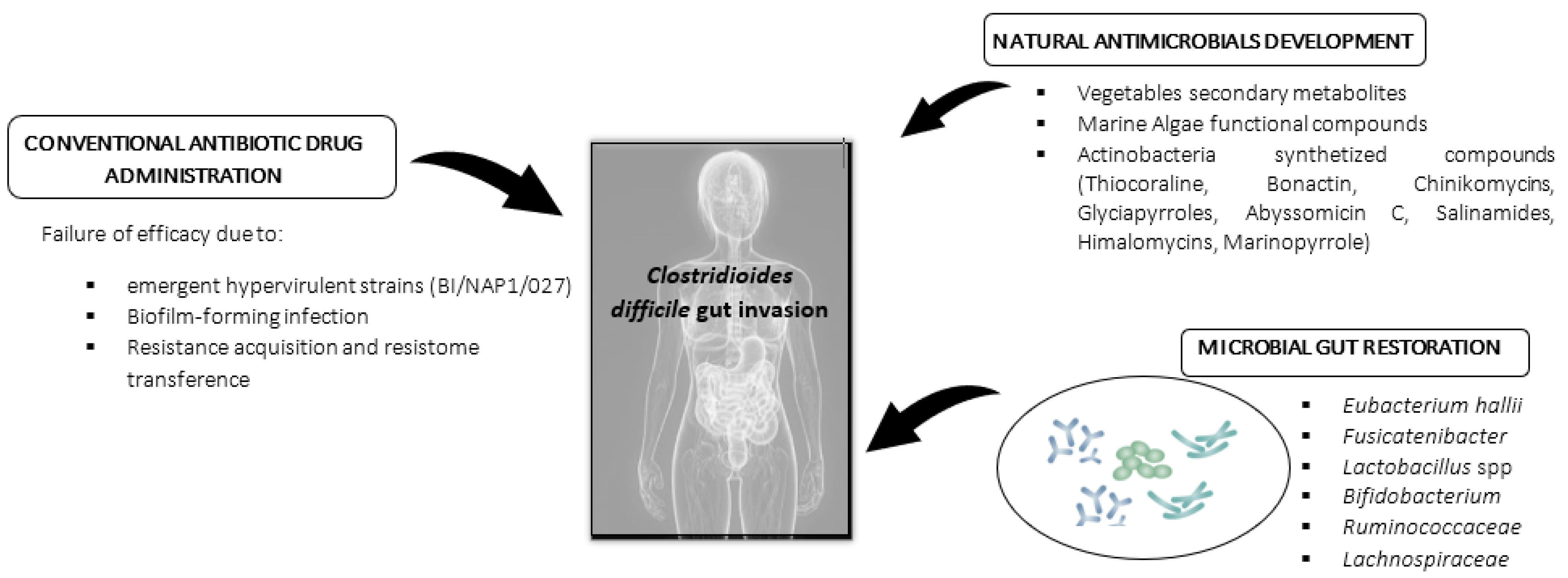
:1. Introduction
Current Antibiotic Therapies Applied against Clostridioides difficile
2. Natural Antimicrobials against Clostridioides difficile Infection (CDI): Nutraceutical and Pharmaceutical Approach
2.1. Vegetable Compounds in Clostridioides difficile Infection Mitigation: Research Advances
2.2. Probiotic Administration, Microbiota Restoration (Fecal Transplantation), and Microbiota Diet Modulation: A Biological Strategy to Improve CDI Resistance
2.3. Marine Natural Compounds as Antimicrobials: Future Niche Strategy against C. difficile
3. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- WHO. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 27 February 2017).
- Peng, Z.; Jin, D.; Kim, H.B.; Stratton, C.W.; Wu, B.; Tang, Y.W.; Sun, X. Update on Antimicrobial Resistance in Clostridium difficile: Resistance Mechanisms and Antimicrobial Susceptibility Testing. J. Clin. Microbiol. 2017, 55, 1998–2008. [Google Scholar] [CrossRef] [Green Version]
- Murray, B.; Wolfe, C.; Marra, A.; Pillar, C.; Shinabarger, D. In vitro activity of the novel antibacterial agent ibezapolstat (ACX-362E) against Clostridioides difficile. J. Antimicrob. Chemother. 2020, 75, 2149–2155. [Google Scholar] [CrossRef]
- Sholeh, M.; Krutova, M.; Forouzesh, M.; Mironov, S.; Sadeghifard, N.; Molaeipour, L.; Maleki, A.; Kouhsari, E. Antimicrobial resistance in Clostridioides (Clostridium) difficile derived from humans: A systematic review and meta-analysis. Antimicrob. Resist. Infect. Control 2020, 9, 158. [Google Scholar] [CrossRef] [PubMed]
- Olsen, M.A.; Stwalley, D.; Demont, C.; Dubberke, E.R. Clostridium difficile infection increases acute and chronic morbidity and mortality. Infect. Control Hosp. Epidemiol. 2019, 40, 65–71. [Google Scholar] [CrossRef] [Green Version]
- ECDC. European Surveillance of Clostridioides (Clostridium) difficile Infections Surveillance Protocol; ECDC: Stockholm, Sweden, 2019; pp. 1–32.
- Zhang, S.; Palazuelos-Munoz, S.; Balsells, E.M.; Harish, N.; Chit, A.; Kyaw, M.H. Cost of hospital management of Clostridium difficile infection in United States—A meta-analysis andmodelling study. BMC Infect. Dis. 2016, 16, 447–465. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, J.R.; Johnson, S.; Gerding, D.N. Clostridium difficile infection caused by the epidemic BI/NAP1/027 strain. Gastroenterology 2009, 136, 1913–1924. [Google Scholar] [CrossRef]
- Ajami, N.J.; Cope, J.L.; Wong, M.C.; Petrosino, J.F.; Chesnel, L. Impact of Oral Fidaxomicin Administration on the Intestinal Microbiota and Susceptibility to Clostridium difficile Colonization in Mice. Antimicrob. Agents Chemother. 2018, 62, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Abou Chakra, C.N.; Pepin, J.; Sirard, S.; Valiquette, L. Risk factors for recurrence, complications and mortality in Clostridium difficile infection: A systematic review. PLoS ONE 2014, 9, e98400. [Google Scholar] [CrossRef] [Green Version]
- Clarkin, C.; Quist, S.; Shamis, R.; King, A.E.; Shah, B.M. Management of Clostridioides difficile Infection. Crit. Care Nurse 2019, 39, e1–e12. [Google Scholar] [CrossRef]
- Skinner, K.; Birchall, S.; Corbett, D.; Thommes, P.; Locher, H.H. Time-kill kinetics of cadazolid and comparator antibacterial agents against different ribotypes of Clostridium difficile. J. Med. Microbiol. 2018, 67, 1402–1409. [Google Scholar] [CrossRef]
- Tsutsumi, L.S.; Owusu, Y.B.; Hurdle, J.G.; Sun, D. Progress in the discovery of treatments for C. difficile infection: A clinical and medicinal chemistry review. Curr. Top. Med. Chem. 2014, 14, 152–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louie, T.J.; Miller, M.A.; Mullane, K.M.; Weiss, K.; Lentek, A. Fidaxomicin versus vancomycin for Clostridium difficile infection. Antimicrob. Agents Chemother. 2011, 58, 901–908. [Google Scholar]
- Ooijevaar, R.; Van Beurden, Y.; Terveer, E.; Goorhuis, A.; Bauer, M.; Keller, J. Update of treatment algorithms for Clostridium difficile infection. Clin. Microbiol. Infect. 2018, 24, 452–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dapa, T.; Unnikrishnan, M. Biofilm formation by Clostridium difficile. Gut Microbes 2013, 4, 397–402. [Google Scholar] [CrossRef] [Green Version]
- Frost, L.R.; Cheng, J.K.J.; Unnikrishnan, M. Clostridioides difficile biofilms: A mechanism of persistence in the gut? PLoS Pathog. 2021, 17, e1009348. [Google Scholar] [CrossRef]
- Semenyuk, E.G.; Laning, M.L.; Foley, J.; Johnston, P.F.; Knight, K.L.; Gerding, D.N.; Driks, A. Spore formation and toxin production in Clostridium difficile biofilms. PLoS ONE 2014, 9, e87757. [Google Scholar] [CrossRef] [Green Version]
- Pal, R.; Seleem, M.N. Screening of Natural Products and Approved Oncology Drug Libraries for Activity against Clostridioides difficile. Sci. Rep. 2020, 10, 5966. [Google Scholar] [CrossRef] [Green Version]
- Naclerio, G.A.; Abutaleb, N.S.; Li, D.; Seleem, M.N.; Sintim, H.O. Ultrapotent Inhibitor of Clostridioides difficile Growth, Which Suppresses Recurrence in vivo. J. Med. Chem. 2020, 63, 11934–11944. [Google Scholar] [CrossRef]
- Speri, E.; Janardhanan, J.; Masitas, C.; Schroeder, V.A.; Lastochkin, E.; Wolter, W.R.; Fisher, J.F.; Mobashery, S.; Chang, M. Discovery of a Potent Picolinamide Antibacterial Active against Clostridioides difficile. ACS Infect. Dis. 2020, 6, 2362–2368. [Google Scholar] [CrossRef]
- Mefferd, C.C.; Bhute, S.S.; Phan, J.R.; Villarama, J.V.; Do, D.M.; Alarcia, S.; Abel-Santos, E.; Hedlunda, B.P. A High-Fat/High-Protein, Atkins-Type Diet Exacerbates Clostridioides (Clostridium) difficile Infection in Mice, whereas a High-Carbohydrate Diet Protects. Msystems 2020, 5, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Mody, D.; Athamneh, A.I.M.; Seleem, M.N. Curcumin: A natural derivative with antibacterial activity against Clostridium difficile. J. Glob. Antimicrob. Resist. 2020, 21, 154–161. [Google Scholar] [CrossRef]
- Roshan, N.; Riley, T.V.; Hammer, K.A. Antimicrobial activity of natural products against Clostridium difficile in vitro. J. Appl. Microbiol. 2017, 123, 92–103. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Palafox-Rosas, R.; Luna, B.; She, R.C. The Bactericidal Activity and Spore Inhibition Effect of Manuka Honey against Clostridioides Difficile. Antibiotics 2020, 9, 684. [Google Scholar] [CrossRef]
- Num, S.M.; Useh, N.M. Clostridium: Pathogenic roles, industrial uses and medicinal prospects of natural products as ameliorative agents against pathogenic species. Jordan J. Biol. Sci. 2014, 7, 81–94. [Google Scholar]
- Kačániová, M.; Vukovič, N.; Horská, E.; Šalamon, I.; Bobková, A.; Hleba, L.; Mellen, M.; Vatľák, A.; Petrová, J.; Bobko, M. Antibacterial activity against Clostridium genus and antiradical activity of the essential oils from different origin. J. Environ. Sci. Health B 2014, 49, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Mahizan, N.A.; Yang, S.K.; Moo, C.L.; Song, A.A.L.; Chong, C.M.; Chong, C.W.; Abushelaibi, A.; Lim, S.H.E.; Lai, K.S.J.M. Terpene Derivatives as a Potential Agent against Antimicrobial Resistance (AMR). Pathogens 2019, 24, 2631. [Google Scholar] [CrossRef] [Green Version]
- Alghanmi, H.A.; Omran, A.S. Antibacterial activity of ethanol extracts of two algae species against some pathogenic bacteria isolated from hospital patients. Eur. Asian J. Biosci. 2020, 14, 383–394. [Google Scholar]
- Han, Z.L.; Yang, M.; Fu, X.D.; Chen, M.; Su, Q.; Zhao, Y.H.; Mou, H.J. Evaluation of Prebiotic Potential of Three Marine Algae Oligosaccharides from Enzymatic Hydrolysis. Mar. Drugs 2019, 17, 173. [Google Scholar] [CrossRef] [Green Version]
- Poveda-Castillo, G.; Rodrigo, D.; Martínez, A.; Pina-Pérez, M.C. Bioactivity of Fucoidan as an Antimicrobial Agent in a New Functional Beverage. Beverages 2018, 4, 64. [Google Scholar] [CrossRef] [Green Version]
- Abadi, M.S.S.; Mirzaei, E.; Bazargani, A.; Gholipour, A.; Heidari, H.; Hadi, N. Antibacterial activity and mechanism of action of chitosan nanofibers against toxigenic Clostridioides (Clostridium) difficile Isolates. Ann. Ig. 2020, 32, 72–80. [Google Scholar]
- Bundale, S.; Singh, J.; Begde, D.; Nashikkar, N.; Upadhyay, A. Rare actinobacteria: A potential source of bioactive polyketides and peptides. World J. Microbiol. Biotechnol. 2019, 35, 92. [Google Scholar] [CrossRef] [PubMed]
- Stincone, P.; Brandelli, A. Marine bacteria as source of antimicrobial compounds. Crit. Rev. Biotechnol. 2020, 40, 306–319. [Google Scholar] [CrossRef] [PubMed]
- Manivasagana, P.; Kanga, K.H.; Sivakumarb, K.; Li-Chanc, E.C.Y.; Oha, H.M.; Kima, S.K. Marine actinobacteria: An important source of bioactive natural products. Environ. Toxicol. Pharmacol. 2014, 38, 172–188. [Google Scholar] [CrossRef] [PubMed]
- Pina-Pérez, M.C.; Brück, W.M.; Brück, T.; Beyrer, M. Chapter 4—Microalgae as healthy ingredients for functional foods. In The Role of Alternative and Innovative Food Ingredients and Products in Consumer Wellness, 1st ed.; Galanakis, C.M., Ed.; Elsevier: Cambridge, MA, USA, 2019; pp. 103–137. [Google Scholar]
- Pina-Pérez, M.C.; Rivas, A.; Martinez-Lopez, A.; Rodrigo, D. Antimicrobial potential of macro and microalgae against pathogenic and spoilage microorganisms in food. Food Chem. 2017, 235, 34–44. [Google Scholar] [CrossRef]
- Aljarallah, K.M. Inhibition of Clostridium difficile by natural herbal extracts. J. Taibah Univ. Med. Sci. 2016, 11, 427–431. [Google Scholar] [CrossRef] [Green Version]
- Ghaedi, M.; Rezaee, M.A.; Fakhari, S.; Ghorashi, M.; Pirmohamadi, H.; Doost, R.D.; Rahmani, M.R. Effects of ethanol extract of Commiphora myrrha gum against Helicobacter pylori and inhibition of proliferation of the human inhibition of proliferation of the human AGS cell line in vitro. J. Aust. Trad. Med. Sci. 2014, 20, 106–110. [Google Scholar]
- Alhussaini, M.S.; Saadabi, A.M.; Alghonaim, M.I.; Ibrahim, K.E. An Evaluation of the antimicrobial activity of Commiphora myrrha Nees (Engl.) oleo-gum resins from Saudi Arabia. J. Med. Sci. 2015, 15, 198–203. [Google Scholar] [CrossRef] [Green Version]
- Piotrowski, M.; Karpinski, P.; Pituch, H.; van Belkum, A.; Obuch-Woszczatynski, P. Antimicrobial effects of Manuka honey on in vitro biofilm formation by Clostridium difficile. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1661–1664. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.R.M.; Flemer, B.; Joyce, S.A.; Zulquernain, A.; Sheehan, D.; Shanahan, F.; O’Toole, P.W. Changes in microbiota composition, bile and fatty acid metabolism, in successful faecal microbiota transplantation for Clostridioides difficile infection. BMC Gastroenterol. 2018, 18, 131. [Google Scholar] [CrossRef] [Green Version]
- Crobach, M.J.T.; Ducarmon, Q.R.; Terveer, E.M.; Harmanus, C.; Sanders, I.M.J.G.; Verduin, K.M.; Kuijper, E.D.J.; Zwittink, R.D. The Bacterial Gut Microbiota of Adult Patients Infected, Colonized or Noncolonized by Clostridioides difficile. Microorganisms 2020, 8, 677. [Google Scholar] [CrossRef] [PubMed]
- Schnizlein, M.K.; Vendrov, K.C.; Edwards, S.J.; Martens, E.C.; Younga, V.B. Dietary Xanthan Gum Alters Antibiotic Efficacy against the Murine Gut Microbiota and Attenuates Clostridioides difficile Colonization. Am. Soc. Microbiol. 2020, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Theriot, C.M.; Young, V.B. Interactions Between the Gastrointestinal Microbiome and Clostridium difficile. Annu. Rev. Microbiol. 2015, 69, 445–461. [Google Scholar] [CrossRef] [Green Version]
- Maida, M.; Mcilroy, J.; Ianiro, G.; Cammarota, G. Faecal Microbiota Transplantation as Emerging Treatment in European Countries. Adv. Exp. Med. Biol. 2017, 1050, 177–196. [Google Scholar]
- Cammarota, G.; Ianiro, G.; Tilg, H.; Rajilic-Stojanovic, M.; Kump, P.; Satokari, R.; Sokol, H.; Arkkila, P.; Pintus, C.; Hart, A.; et al. European consensus conference on faecal microbiota transplantation in clinical practice. Gut 2017, 66, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, F.; Skally, M.; Brady, M.; Burns, K.; Rooney, C.; Wilcox, M.H. European Practice for CDI Treatment. Adv. Exp. Med. Biol. 2018, 8, 117–135. [Google Scholar]
- Blount, K.F.; Shannon, W.D.; Deych, E.; Jones, C. Restoration of Bacterial Microbiome Composition and Diversity Among Treatment Responders in a Phase 2 Trial of RBX2660: An Investigational Microbiome Restoration Therapeutic. Open Forum. Infect. Dis. 2019, 6, ofz095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGovern, B.H.; Ford, C.B.; Henn, M.R.; Pardi, D.S.; Khanna, S.; Hohmann, E.L.; O’Brien, J.E.; Desjardins, C.A.; Bernardo, P.; Wortman, J.R.; et al. SER-109, an Investigational Microbiome Drug to Reduce Recurrence After Clostridioides difficile Infection: Lessons Learned From a Phase 2 Trial. Clin. Infect. Dis. 2020, ciaa387. [Google Scholar] [CrossRef] [Green Version]
- Auclair, J.; Frappier, M.; Millette, M. Lactobacillus acidophilus CL1285, Lactobacillus casei LBC80R, and Lactobacillus rhamnosus CLR2 (Bio-K+): Characterization, Manufacture, Mechanisms of Action, and Quality Control of a Specific Probiotic Combination for Primary Prevention of Clostridium difficile Infection. Clin. Infect. Dis. 2015, 60, S135–S143. [Google Scholar] [PubMed]
- Valdés-Varela, L.; Gueimonde, M.; Ruas-Madiedo, P. Probiotics for prevention and treatment of Clostridium difficile infection. Adv. Exp. Med. Biol. 2018, 1050, 161–176. [Google Scholar]
- Fredua-Agyeman, M.; Stapleton, P.; Basit, A.W.; Beezer, A.E.; Gaisford, S. In vitro inhibition of Clostridium difficile by commercial probiotics: A microcalorimetric study. Int. J. Pharm. 2017, 517, 96–103. [Google Scholar] [CrossRef]
- Ollech, J.E.; Shen, N.T.; Crawford, C.V.; Ringel, Y. Use of probiotics in prevention and treatment of patients with Clostridium difficile infection. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 111–118. [Google Scholar] [CrossRef]
- Khattab, R.A.; Ahmed, N.A.; Ragab, Y.M.; Rasmy, S.A. Bacteria producing antimicrobials against Clostridium difficile isolated from human stool. Anaerobe 2020, 63, 102206. [Google Scholar] [CrossRef] [PubMed]
- Szajewska, H.; Canani, R.B.; Guarino, A.; Hojsak, I.; Indrio, F.; Kolacek, S.; Orel, R.; Shamir, R.; Vandenplas, Y.; van Goudoever, J.B.; et al. Probiotics for the Prevention of Antibiotic-Associated Diarrhea in Children. J. Pediatr. Gastroenterol. Nutr. 2016, 62, 495–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Zhu, Y.; Zhang, Y.; Hamza, T.; Yu, H.; Saint-Flur, A.; Galen, J.; Yang, Z.; Feng, H. A probiotic yeast-based immunotherapy against Clostridioides difficile infection. Sci. Transl. Med. 2020, 12, 4905–4912. [Google Scholar] [CrossRef]
- Jochems, P.G.M.; Garssen, J.; Rietveld, P.C.S.; Govers, C.; Tomassen, M.M.M.; Wichers, H.J.; van Bergenhenegouwen, J.; Masereeuw, R. Novel Dietary Proteins Selectively Affect Intestinal Health In Vitro after Clostridium difficile-Secreted Toxin A Exposure. Nutrients 2020, 12, 2782. [Google Scholar] [CrossRef] [PubMed]
- Hryckowian, A.J.; Van Treuren, W.; Smits, S.A.; Davis, N.M.; Gardner, J.O.; Bouley, D.M.; Sonnenburg, J.L. Microbiota-accessible carbohydrates suppress Clostridium difficile infection in a murine model. Nat. Microbiol. 2018, 3, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Adarme-Vega, T.C.; Thomas-Hall, S.R.; Schenk, P.M. Towards sustainable sources for omega-3 fatty acids production. Curr. Opin. Biotechnol. 2014, 26, 14–22. [Google Scholar] [CrossRef]
- Pujiastuti, D.Y.; Ghoyatul Amin, M.N.; Alamsjah, M.A.; Hsu, J.L. Marine Organisms as Potential Sources of Bioactive Peptides that Inhibit the Activity of Angiotensin I-Converting Enzyme: A Review. Molecules 2019, 24, 2541. [Google Scholar] [CrossRef] [Green Version]
- Ansar, H.; Suleria, R.; Gobe, G.; Masci, P.; Osborne, S.A. Marine bioactive compounds and health promoting perspectives; innovation pathways for drug discovery. Trends Food Sci. Technol. 2016, 50, 44–55. [Google Scholar]
- Jaiganesh, R.; Sampath Kumar, N.S. Marine bacterial sources of bioactive compounds. Adv. Food Nutr. Res. 2012, 65, 389–408. [Google Scholar]
- Atashrazm, F.; Lowenthal, R.M.; Woods, G.M.; Holloway, A.F.; Dickinson, J.L. Fucoidan and Cancer: A Multifunctional Molecule with Anti-Tumor Potential. Mar. Drugs 2015, 13, 2327–2346. [Google Scholar] [CrossRef] [Green Version]
- Nikapitiya, C. Bioactive secondary metabolites from marine microbes for drug discovery. Adv. Food Nutr. Res. 2012, 65, 363–387. [Google Scholar]
- Palacios-Gorba, C.; Pina, R.; Tortajada-Girbés, M.; Jiménez-Belenguer, A.; Siguemoto, E.; Ferrús, M.A.; Rodrigo, D.; Pina-Pérez, M.C. Caenorhabditis elegans as an in vivo model to assess fucoidan bioactivity preventing Helicobacter pylori infection. Food Funct. 2020, 11, 4525–4534. [Google Scholar] [CrossRef] [PubMed]
- Aziz, E.; Batool, R.; Khan, M.U.; Rauf, A.; Akhtar, W.; Heydari, M.; Rehman, S.; Shahzad, T.; Malik, A.; Mosavat, S.H.; et al. An overview on red algae bioactive compounds and their pharmaceutical applications. J. Complement. Integr. Med. 2020, 17, 1–21. [Google Scholar] [CrossRef]
- Udayan, A.; Arumugam, M.; Pandey, A. Chapter 4—Nutraceuticals From Algae and Cyanobacteria. In Algal Green Chemistry, Recent Progress in Biotechnology, 1st ed.; Rastogi, R.P., Madamwar, D., Pandey, A., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 65–89. [Google Scholar]
- Silva, A.; Silva, S.A.; Lourenço-Lopes, C.; Jimenez-Lopez, C.; Carpena, M.; Gullón, P.; Fraga-Corral, M.; Domingues, V.F.; Barroso, M.F.; Simal-Gandara, J.; et al. Antibacterial Use of Macroalgae Compounds against Foodborne Pathogens. Antibiotics 2020, 9, 712. [Google Scholar] [CrossRef] [PubMed]
- Salvador, N.; Gomez-Garreta, A.; Lavelli, L.; Ribera, M.A. Antimicrobial activity of Iberian macroalgae. Sci. Mar. 2007, 71, 101–113. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Gupta, C.; Garg, A.P.; Prakash, D. Prebiotic Efficiency of Blue Green Algae on Probiotics Microorganisms. J. Microbiol. Exp. 2017, 4, 00120. [Google Scholar] [CrossRef] [Green Version]
- Guzman, F.; Wong, G.; Román, T.; Cárdenas, C.; Alvárez, C.; Schmit, P.; Albericio, F.; Rojas, V. Identification of Antimicrobial Peptides from the Microalgae Tetraselmis suecica (Kylin) Butcher and Bactericidal Activity Improvement. Mar. Drugs 2019, 17, 453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seghiri, R.; Kharbach, M.; Essamri, A. Functional composition, nutritional properties, and biological activities of Moroccan Spirulina Microalga. J. Food Qual. 2019, 2019, 3707219. [Google Scholar] [CrossRef] [Green Version]
- Syed, S.; Arasu, A.; Ponnuswamy, I. The Uses of Chlorella vulgaris as Antimicrobial Agent and as a Diet: The Presence of Bio-active Compounds which caters the Vitamins, Minerals in General. Int. J. Biosci. Biotechnol. 2015, 7, 185–190. [Google Scholar] [CrossRef]
- Besednova, N.N.; Zaporozhets, T.S.; Somova, L.M.; Kuznetsova, T.A. Review: Prospects for the use of extracts and polysaccharides from marine algae to prevent and treat the diseases caused by Helicobacter pylori. Helicobacter 2015, 20, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Fields, F.J.; Lejzerowicz, F.; Schroeder, D.; Ngoi, S.M.; Tran, M.; McDonald, D.; Jiang, L.; Chang, J.T.; Knight, R.; Mayfield, S. Effects of the microalgae Chlamydomonas on gastrointestinal health. J. Funct. Foods 2020, 65, 103738. [Google Scholar] [CrossRef]
- Cox, A.J.; Cripps, A.W.; Taylor, P.A.; Fitton, H.; West, N. Fucoidan Supplementation Restores Fecal Lysozyme Concentrations in High-Performance Athletes:A Pilot Study. Mar. Drugs 2018, 18, 412. [Google Scholar] [CrossRef] [PubMed]
- Ochoa, J.L.; Sanchez, L.M.; Koo, B.M.; Doherty, J.S.; Rahendram, M.; Huang, K.C.; Gross, A.A.; Linington, R.G. Marine Mammal Microbiota Yields Novel Antibiotic with Potent Activity Against Clostridium difficile. ACS Infect. Dis. 2018, 4, 59–67. [Google Scholar] [CrossRef]
- Shinde, P.; Banerjee, P.; Mandhare, A. Marine natural products as source of new drugs: A patent review (2015–2018). Expert Opin. Therap. Pat. 2019, 29, 283–309. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Antibiotics and Natural Antimicrobials | Concentration | C. difficile (Inhibition mm) | References |
|---|---|---|---|
| Antibiotics | MIC (µg/mL)/MBC (µg/mL) | ||
| Vancomycin (VAN) | 0.5–4 | - | [2,4] |
| Metronidazole (MTZ) | 0.25–16 | - | [2,4] |
| Fidaxomicin (FDX) | 0.015–1 | - | [9,23] |
| Ibezapolstat | 2–4 | - | [3] |
| Cadazolid | 0.12–0.25 | - | [12] |
| Vegetable Origin Matrices | MIC | ||
| Vancomycin (positive control) | 30.3 ± 0.7 | [24] | |
| Onion juice | 100% (v/v) | 10.3 ± 0.6 | |
| Garlic juice | 100% (v/v) | 27.0 ± 1.0 | |
| Ginger juice | 100% (v/v) | - | |
| Garlic powder | (20% w/v) | 26.6 ± 0.6 | |
| Cinnamon powder | (20% w/v) | 20.9 ± 0.9 | |
| Curcumin | 4–32 µg/mL | md | [25] |
| Manuka honey | 50% (v/v) | 11.4 ± 0.5 | [26] |
| Nigella sativa (black seed oil) | 2% (v/v) | >15 | [27] |
| Commiphora myrrha (water extract) | 2% (v/v) | >15 | |
| EOs (Satureia montana, Abies alba Mill., and Thymus vulgaris) | 50 µL/mL | >20 | [28] |
| Marine Antimicrobials | |||
| Chlorella spp. and Spirulina platensis EOs | 300 µg/mL | 8–21 | [29,30] |
| Polysaccharides from Laminaria, Saccharina, Spirulina platensis, Chlorella species, Dunaliella salina, and Scenedesmus | 25–100 µg/mL | md | [31,32] |
| Chitosan | 0.25 mg/mL | md | [33] |
| Phocoenamicin (from Micromonospora auratinigra) | 2.6 µM | >15 | [34] |
| Thiocoraline (peptide) from Micromonospora sp.; bonactin (esters) and chinikomycins A from Streptomyces sp. | 4 µg/mL | >15 | [35] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tortajada-Girbés, M.; Rivas, A.; Hernández, M.; González, A.; Ferrús, M.A.; Pina-Pérez, M.C. Alimentary and Pharmaceutical Approach to Natural Antimicrobials against Clostridioides difficile Gastrointestinal Infection. Foods 2021, 10, 1124. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10051124
Tortajada-Girbés M, Rivas A, Hernández M, González A, Ferrús MA, Pina-Pérez MC. Alimentary and Pharmaceutical Approach to Natural Antimicrobials against Clostridioides difficile Gastrointestinal Infection. Foods. 2021; 10(5):1124. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10051124
Chicago/Turabian StyleTortajada-Girbés, Miguel, Alejandro Rivas, Manuel Hernández, Ana González, Maria A. Ferrús, and Maria C. Pina-Pérez. 2021. "Alimentary and Pharmaceutical Approach to Natural Antimicrobials against Clostridioides difficile Gastrointestinal Infection" Foods 10, no. 5: 1124. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10051124