
One-Day Molecular Detection of Salmonella and Campylobacter in Chicken Meat: A Pilot Study

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Design and Experimental Contamination
2.2. Pathogen Plate Count
2.3. DNA Extraction
2.4. Real-Time PCR
2.5. Colorimetric LAMP
2.6. Statistical Analysis
3. Results
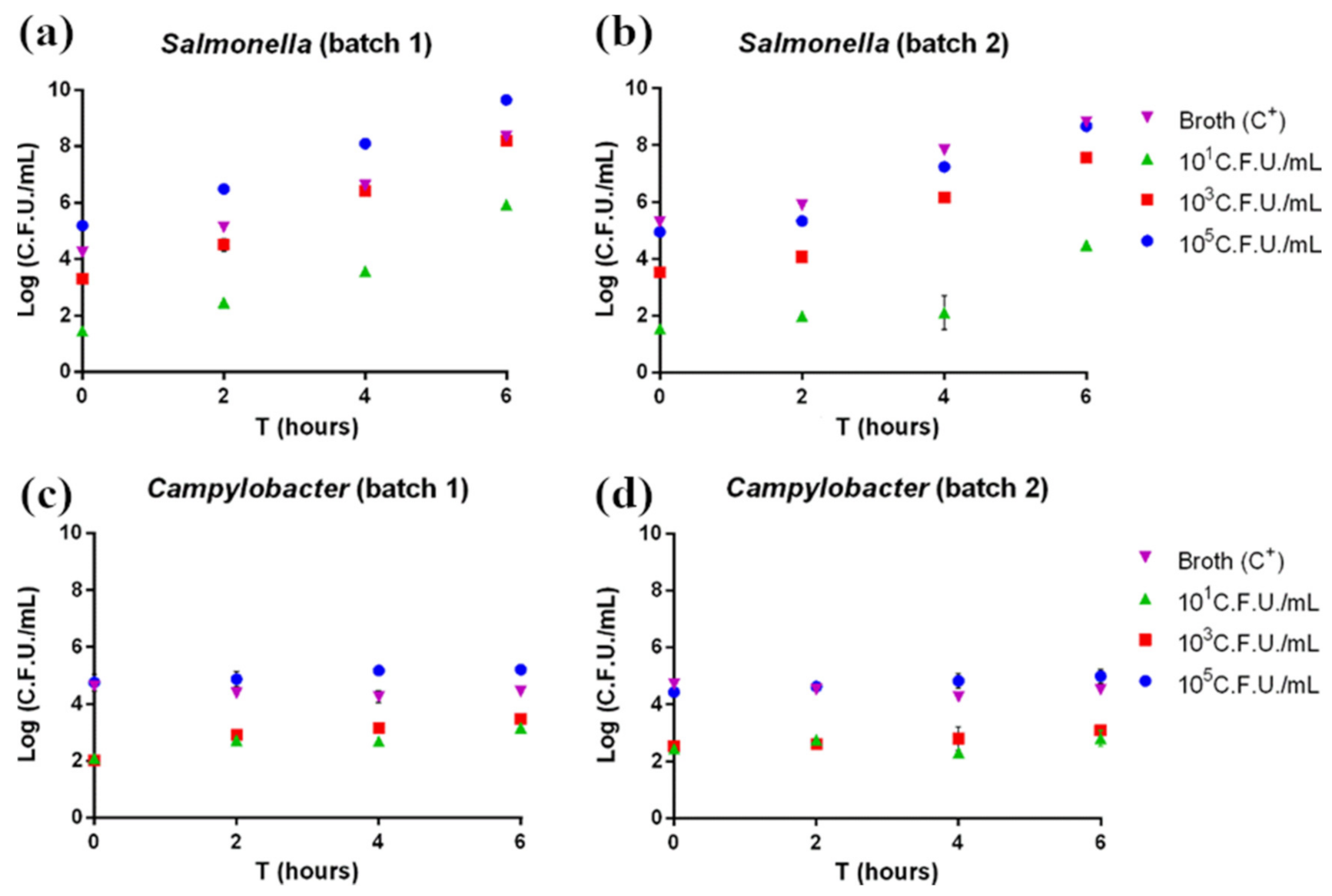
3.1. Pathogen Plate Count
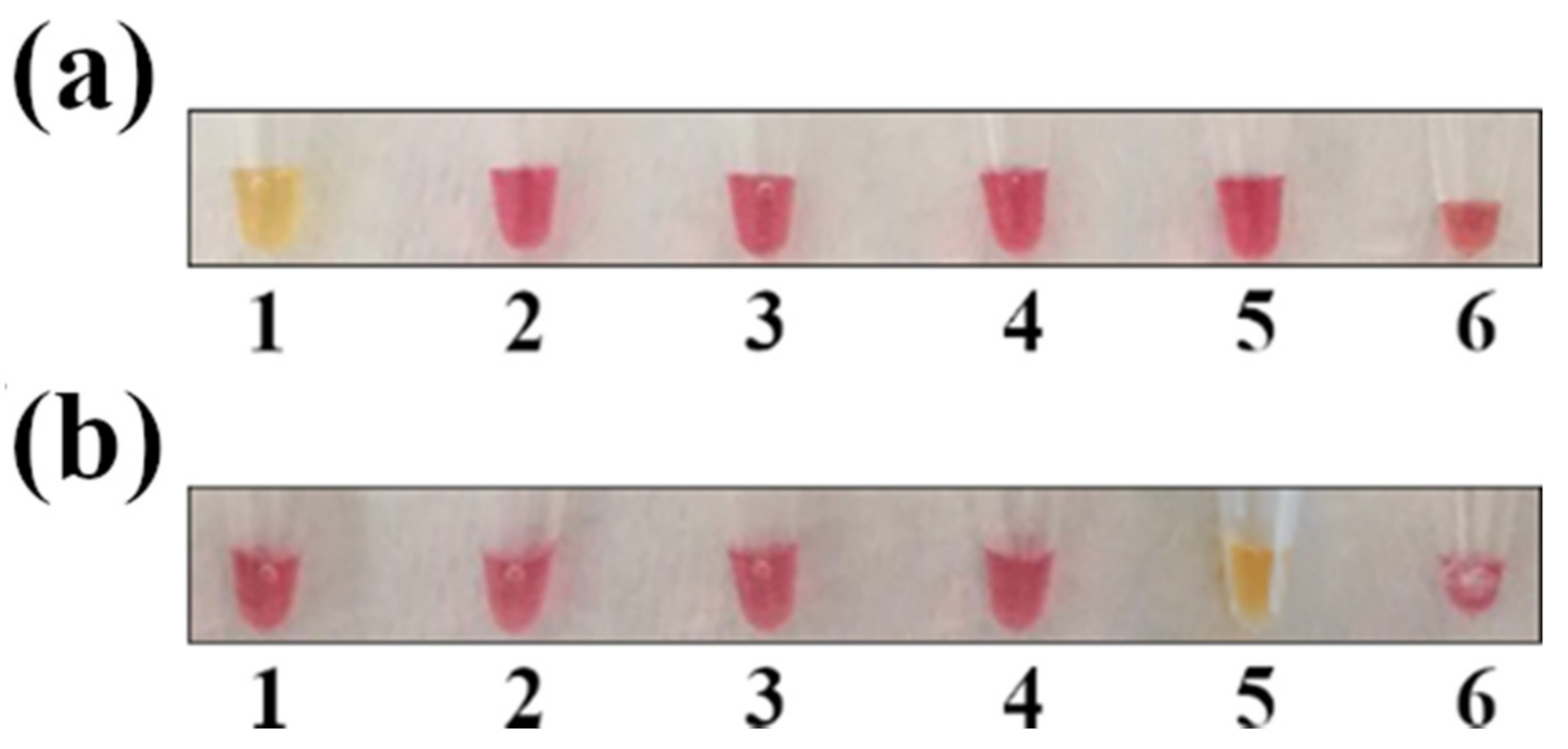
3.2. Colorimetric LAMP Specificity and Inclusivity Tests
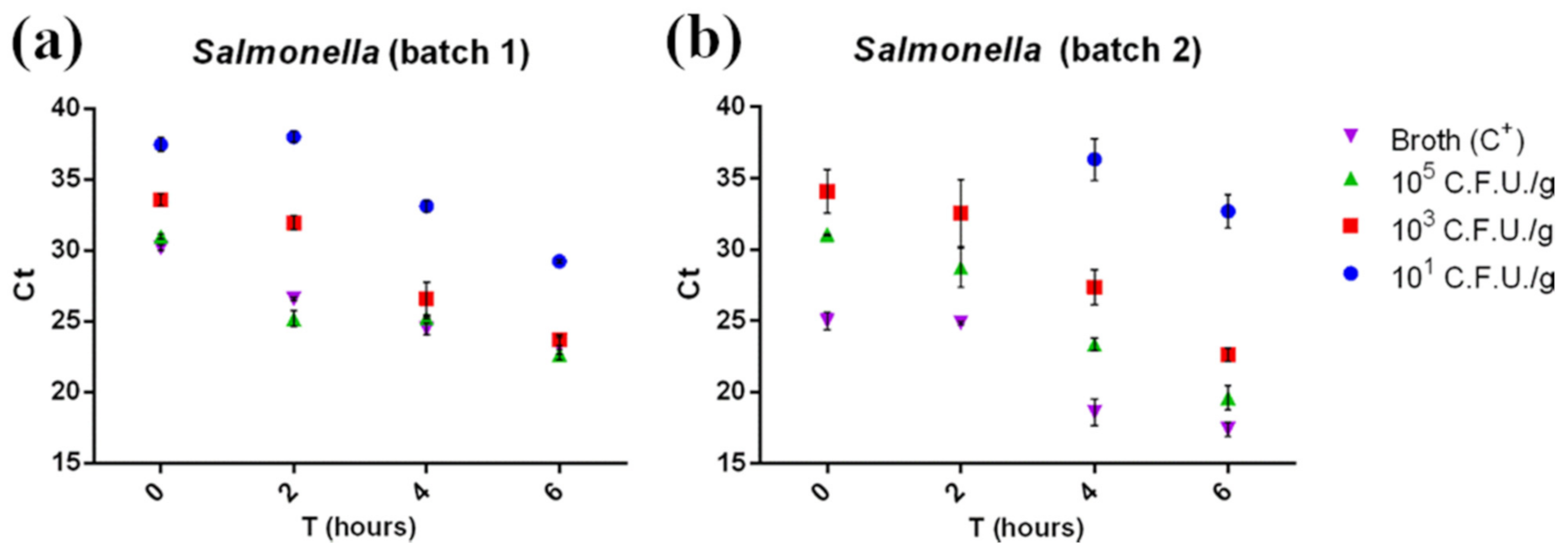
3.3. Real-Time PCR and LAMP for Salmonella
3.4. RT-PCR and LAMP for Campylobacter
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- EFSA; ECDC. The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, e06406. Available online: https://0-efsa-onlinelibrary-wiley-com.brum.beds.ac.uk/doi/abs/10.2903/j.efsa.2021.6406 (accessed on 20 April 2021). [CrossRef]
- Antunes, P.; Mourão, J.; Campos, J.; Peixe, L. Salmonellosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 110–121. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=antunes+2016+salmonellosis (accessed on 20 April 2021). [CrossRef] [Green Version]
- Jajere, S.M. A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and antimicrobial resistance including multidrug resistance. Vet. World 2019, 12, 504–521. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=jajere+2019 (accessed on 20 April 2021). [CrossRef] [PubMed] [Green Version]
- Bryan, F.L.; Doyle, M.P. Health Risks and Consequences of Salmonella and Campylobacter jejuni in Raw Poultry. J. Food Prot. 1995, 58, 326–344. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=bryan+1995+salmonella (accessed on 20 April 2021). [CrossRef] [PubMed]
- Facciolà, A.; Riso, R.; Avventuroso, E.; Visalli, G.; Delia, S.A.; Laganà, P. Campylobacter: From microbiology to prevention. J. Prev. Med. Hyg. 2017, 58, E79–E92. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=facciola+2017 (accessed on 20 April 2021).
- Mughini-Gras, L.; Enserink, R.; Friesema, I.; Heck, M.; van Duynhoven, Y.; van Pelt, W. Risk factors for human salmonellosis originating from pigs, cattle, broiler chickens and egg laying hens: A combined case-control and source attribution analysis. PLoS ONE 2014, 9, e87933. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=mughini-gras+2014+risk+factors (accessed on 20 April 2021). [CrossRef] [Green Version]
- Bodulev, O.L.; Sakharov, I.Y. Isothermal Nucleic Acid Amplification Techniques and Their Use in Bioanalysis. Biochemistry (Mosc) 2020, 85, 147–166. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=bodulev+2020 (accessed on 20 April 2021). [CrossRef]
- Yang, Q.; Domesle, K.J.; Ge, B. Loop-Mediated Isothermal Amplification for Salmonella Detection in Food and Feed: Current Applications and Future Directions. Foodborne Pathog. Dis. 2018, 15, 309–331. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=yang+2018+loop-mediated+salmonella+ (accessed on 20 April 2021). [CrossRef] [Green Version]
- Maruyama, F.; Kenzaka, T.; Yamaguchi, N.; Tani, K.; Nasu, M. Detection of bacteria carrying the stx2 gene by in situ loop-mediated isothermal amplification. Appl. Environ. Microbiol. 2003, 69, 5023–5028. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, H.; Iida, T.; Aoki, K.; Ohno, S.; Suzutani, T. Sensitive and rapid detection of herpes simplex virus and varicella-zoster virus DNA by loop-mediated isothermal amplification. J. Clin. Microbiol. 2005, 43, 3290–3296. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=kaneko+2005+herpes (accessed on 20 April 2021). [CrossRef] [Green Version]
- Enomoto, Y.; Yoshikawa, T.; Ihira, M.; Akimoto, S.; Miyake, F.; Usui, C.; Suga, S.; Suzuki, K.; Kawana, T.; Nishiyama, Y.; et al. Rapid diagnosis of herpes simplex virus infection by a loop-mediated isothermal amplification method. J. Clin. Microbiol. 2005, 43, 951–955. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=enomoto+2005+herpes (accessed on 20 April 2021). [CrossRef] [Green Version]
- Lau, Y.L.; Meganathan, P.; Sonaimuthu, P.; Thiruvengadam, G.; Nissapatorn, V.; Chen, Y. Specific, sensitive, and rapid diagnosis of active toxoplasmosis by a loop-mediated isothermal amplification method using blood samples from patients. J. Clin. Microbiol. 2010, 48, 3698–3702. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=lau+2010+toxoplasmosis (accessed on 20 April 2021). [CrossRef] [Green Version]
- Li, Y.; Fan, P.; Zhou, S.; Zhang, L. Loop-mediated isothermal amplification (LAMP): A novel rapid detection platform for pathogens. Microb. Pathog. 2017, 107, 54–61. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=li+2017+LAMP (accessed on 20 April 2021). [CrossRef]
- Zhao, X.; Li, Y.; Wang, L.; You, L.; Xu, Z.; Li, L.; He, X.; Liu, Y.; Wang, J.; Yang, L. Development and application of a loop-mediated isothermal amplification method on rapid detection Escherichia coli O157 strains from food samples. Mol. Biol. Rep. 2010, 37, 2183–2188. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=zhao+2010+O157 (accessed on 20 April 2021). [CrossRef] [PubMed]
- Wang, L.; Li, Y.; Chu, J.; Xu, Z.; Zhong, Q. Development and application of a simple loop-mediated isothermal amplification method on rapid detection of Listeria monocytogenes strains. Mol. Biol. Rep. 2012, 39, 445–449. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=wang+2012+development+listeria (accessed on 20 April 2021). [CrossRef] [PubMed]
- Misawa, Y.; Yoshida, A.; Saito, R.; Yoshida, H.; Okuzumi, K.; Ito, N.; Okada, M.; Moriya, K.; Koike, K. Application of loop-mediated isothermal amplification technique to rapid and direct detection of methicillin-resistant Staphylococcus aureus (MRSA) in blood cultures. J. Infect. Chemother. 2007, 13, 134–140. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=misawa+2007+MRSA (accessed on 20 April 2021). [CrossRef] [PubMed]
- Xu, Z.; Li, L.; Chu, J.; Peters, B.M.; Harris, M.L.; Li, B.; Shi, L.; Shirtliff, M.E. Development and application of loop-mediated isothermal amplification assays on rapid detection of various types of staphylococci strains. Food Res. Int. 2012, 47, 166–173. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=xu+2012+staphylococci (accessed on 20 April 2021). [CrossRef] [Green Version]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, E63. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=notomi+2000 (accessed on 20 April 2021). [CrossRef] [PubMed] [Green Version]
- Domesle, K.J.; Young, S.R.; Yang, Q.; Ge, B. Loop-Mediated Isothermal Amplification for Screening Salmonella in Animal Food and Confirming Salmonella from Culture Isolation. J. Vis. Exp. 2020. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=domesle+2020 (accessed on 20 April 2021). [CrossRef]
- Quyen, T.L.; Nordentoft, S.; Vinayaka, A.C.; Ngo, T.A.; Engelsmenn, P.; Sun, Y.; Madsen, M.; Bang, D.D.; Wolff, A. A Sensitive, Specific and Simple Loop Mediated Isothermal Amplification Method for Rapid Detection of Campylobacter spp. in Broiler Production. Front. Microbiol. 2019, 10, 2443. [Google Scholar] [CrossRef] [PubMed]
- Dalzini, E.; Cosciani-Cunico, E.; Bernini, V.; Bertasi, B.; Losio, M.; Daminelli, P.; Varisco, G. Food control. Food Control 2015, 47, 306–311. Available online: https://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/pii/S0956713514003776 (accessed on 20 April 2021). [CrossRef]
- Zhuang, L.; Gong, J.; Li, Q.; Zhu, C.; Yu, Y.; Dou, X.; Liu, X.; Xu, B.; Wang, C. Detection of Salmonella spp. by a loop-mediated isothermal amplification (LAMP) method targeting bcfD gene. Lett. Appl. Microbiol. 2014, 59, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.R.; D’Agostino, M.; Arias, A.P.; Robles, S.; Casado, C.F.; Iturbe, L.O.; Lerma, O.G.; Andreou, M.; Cook, N. An immunomagnetic separation/loop-mediated isothermal amplification method for rapid direct detection of thermotolerant Campylobacter spp. during poultry production. J. Appl. Microbiol. 2016, 120, 469–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, M.R.; Cook, N. A Rapid LAMP-Based Method for Screening Poultry Samples for Campylobacter without Enrichment. Front. Microbiol. 2018, 9, 2401. [Google Scholar] [CrossRef]
- Vichaibun, V.; Kanchanaphum, P. Quantitative LAMP and PCR Detection of Salmonella in Chicken Samples Collected from Local Markets around Pathum Thani Province, Thailand. Int. J. Food Sci. 2020, 2020, 8833173. [Google Scholar] [CrossRef] [PubMed]
- EUR-Lex—32005R2073—EN—EUR-Lex. Available online: https://eur-lex.europa.eu/legal-content/IT/ALL/?uri=CELEX%3A32005R2073 (accessed on 21 April 2021).
- Nagamine, K.; Hase, T.; Notomi, T. Accelerated reaction by loop-mediated isothermal amplification using loop primers. Mol. Cell Probes 2002, 16, 223–229. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=nagamine+2002 (accessed on 21 April 2021). [CrossRef] [PubMed]
- Mashooq, M.; Kumar, D.; Niranjan, A.K.; Agarwal, R.K.; Rathore, R. Development and evaluation of probe based real time loop mediated isothermal amplification for Salmonella: A new tool for DNA quantification. J. Microbiol. Methods 2016, 126, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Xie, Z.; Zhao, G.; Liu, J.; Pang, Y.; Deng, X.; Xie, Z.; Fan, Q.; Luo, S. A loop-mediated isothermal amplification assay for the visual detection of duck circovirus. Virol. J. 2014, 11, 76. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=xie+2014+circovirus (accessed on 21 April 2021). [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salmonella spp. | Campylobacter spp. | |
|---|---|---|
| Strain 1 | S. Typhimurium (ATCC 6994) | C. jejuni (ATCC 33291) |
| Strain 2 | S. Enteritidis (ATCC 13076) | C. jejuni (ATCC 49943) |
| Strain 3 | S. Infantis (NCTC 6703) | C. coli (ATCC 43478) |
| 10X Mix Concentration | Primers for Salmonella spp. | |
|---|---|---|
| FIP | 16 µM | 3′-TGCACTTTACCGGTACGCTGAATACAGCGGCAATTTCAACCA-5′ |
| BIP | 16 µM | 3′-CGGTCTGGATTCGCAGGTCAAAGCGATAGCCTGGGGAAC-5′ |
| F3 | 2 µM | 3′-CCGGACAAACGATTCTGGTA-5′ |
| B3 | 2 µM | 3′-CCGACATCGGCATTATCCG-5′ |
| LF | 4 µM | 3′-TACCCCCTCCGGCTTTTG-5′ |
| LB | 4 µM | 3′-ACAATGCGTCTTATCGCTACG-5′ |
| Primers for Campylobacter spp. | ||
| FIP | 16 µM | 3′-GGACCGTGTCTCAGTTCCAGTGTGACGGATGAGACTATATAGTATCAGCTAG-5′ |
| BIP | 16 µM | 3′-CGGGAGGCAGCAGTAGGGAATATTGCTAAGAAAAGGAGTTTACGCTCCG-5′ |
| F3 | 2 µM | 3′-CTGCTTAACACAAGTTGAGTAGG-5′ |
| B3 | 2 µM | 3′-TTCCTTAGGTACCGTCAGAA-5′ |
| LF | 4 µM | 3′-GTTAAGCGTCATAGCCTTGGTAA-5′ |
| LB | 4 µM | 3′-GCGTGGAGGATGACACTT-5′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zendrini, A.; Carta, V.; Filipello, V.; Ragni, L.; Cosciani-Cunico, E.; Arnaboldi, S.; Bertasi, B.; Franceschi, N.; Ajmone-Marsan, P.; De Medici, D.; et al. One-Day Molecular Detection of Salmonella and Campylobacter in Chicken Meat: A Pilot Study. Foods 2021, 10, 1132. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10051132
Zendrini A, Carta V, Filipello V, Ragni L, Cosciani-Cunico E, Arnaboldi S, Bertasi B, Franceschi N, Ajmone-Marsan P, De Medici D, et al. One-Day Molecular Detection of Salmonella and Campylobacter in Chicken Meat: A Pilot Study. Foods. 2021; 10(5):1132. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10051132
Chicago/Turabian StyleZendrini, Andrea, Valentina Carta, Virginia Filipello, Laura Ragni, Elena Cosciani-Cunico, Sara Arnaboldi, Barbara Bertasi, Niccolò Franceschi, Paolo Ajmone-Marsan, Dario De Medici, and et al. 2021. "One-Day Molecular Detection of Salmonella and Campylobacter in Chicken Meat: A Pilot Study" Foods 10, no. 5: 1132. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10051132